
Auxin Involvement in Ceratopteris Gametophyte Meristem Regeneration

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
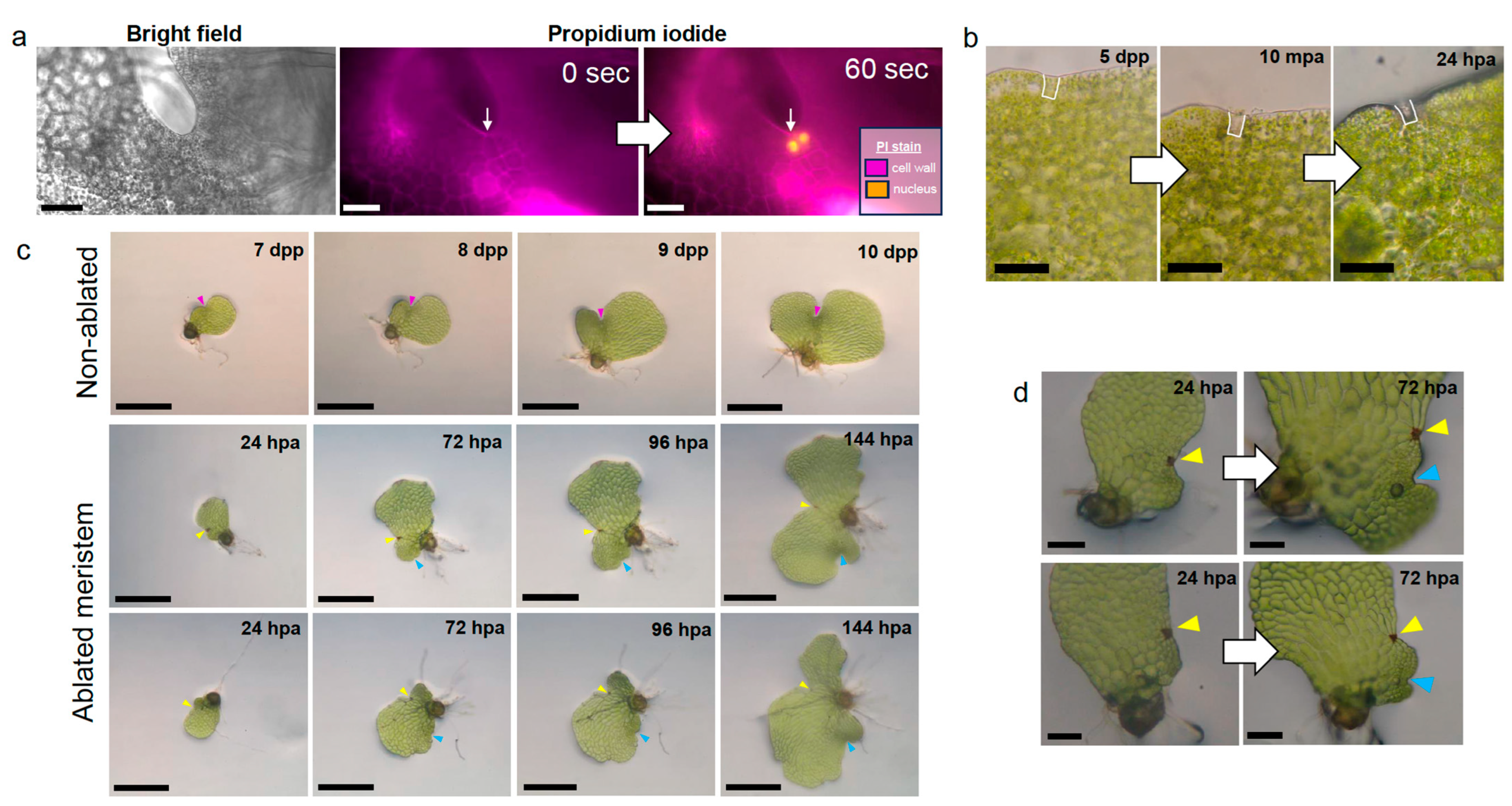
2.1. Laser Ablation of a Single Marginal Meristem Initial Cell Triggers Regeneration
2.2. Meristem Regeneration Occurs within a Predictable Range from Site of Damage
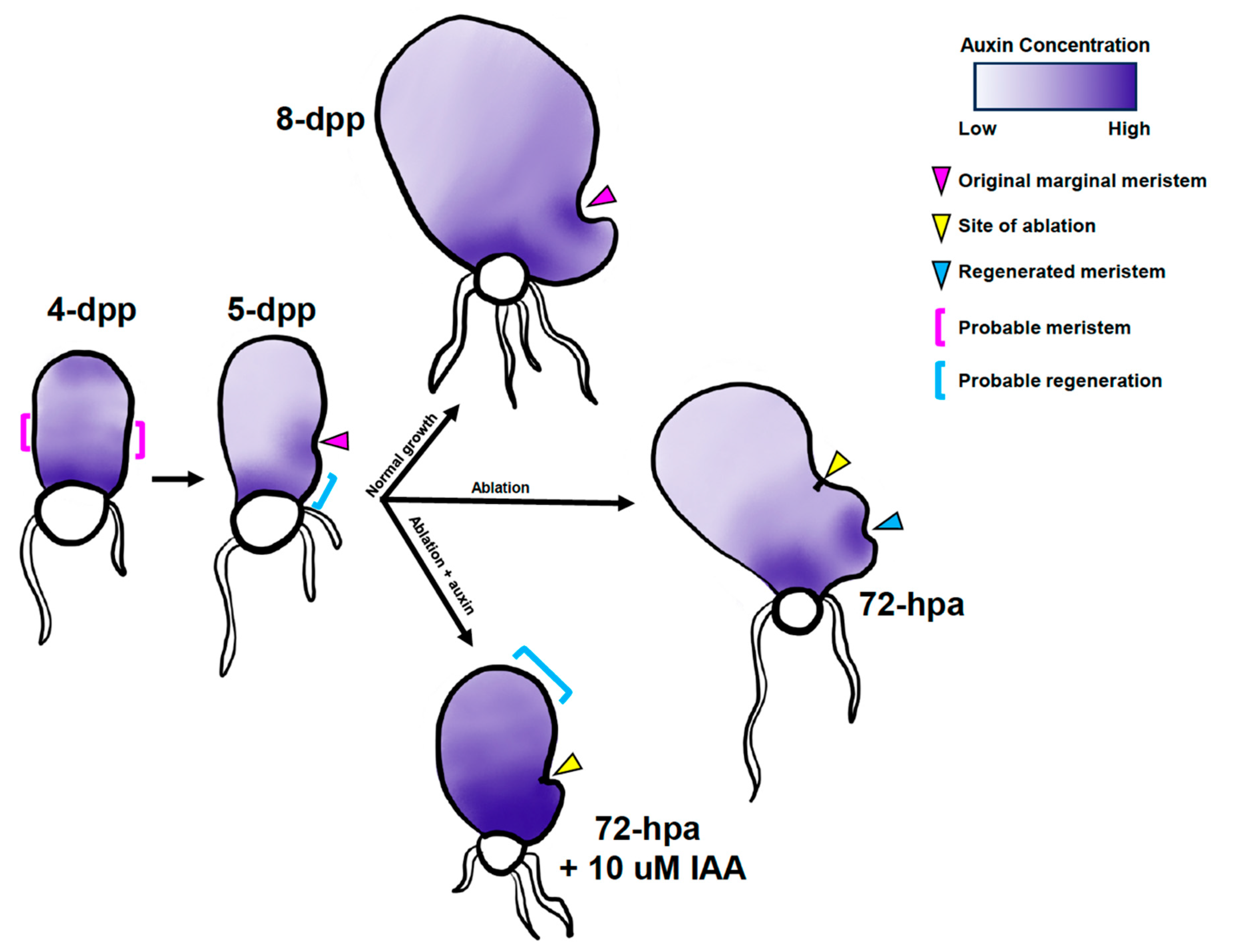
2.3. Ceratopteris Auxin Biosynthesis Gene Expression Localizes to Marginal or Regenerated Meristems
2.4. Exogenous Auxin and Auxin Inhibitors Modulate Gametophyte Morphology
2.5. Auxin Affects the Marginal Meristem Position
2.6. Disruption of Auxin Homeostasis Affects Meristem Regeneration after Ablation
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Laser Ablation of Marginal Meristem Initials
4.3. Phylogeny of Ceratopteris TAA Proteins
4.4. Real-Time Quantitative PCR and Whole Mount In Situ Expression Analyses
4.5. Exogenous Auxin and Auxin Inhibitor Treatments of Gametophytes
4.6. Quantification of Auxin Treatment Response on Non-Ablated and Ablated Hermaphrodites
4.7. Statistical Analyses and Graphical Representations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fouracre, J.P.; Harrison, C.J. How was apical growth regulated in the ancestral land plant? Insights from the development of non-seed plants. Plant Physiol. 2022, 190, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Gregorich, M.; Fisher, R. Auxin regulates lateral meristem activation in developing gametophytes of Ceratopteris richardii. Can. J. Bot. 2006, 84, 1520–1530. [Google Scholar] [CrossRef]
- Imaichi, R. A New Classification of the Gametophyte Development of Homosporous Ferns, Focusing on Meristem Behavior. Fern Gaz. 2013, 19, 141–156. [Google Scholar]
- Wu, X.; Yan, A.; Yang, X.; Banks, J.A.; Zhang, S.; Zhou, Y. Cell growth dynamics in two types of apical meristems in fern gametophytes. Plant J. 2022, 111, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Yan, A.; Zhou, Y. Positional cues and cell division dynamics drive meristem development and archegonium formation in Ceratopteris gametophytes. Commun. Biol. 2022, 5, 650. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Liu, X.; Zhang, S.; Zhou, Y. Cell Division and Meristem Dynamics in Fern Gametophytes. Plants 2023, 12, 209. [Google Scholar] [CrossRef] [PubMed]
- Banks, J.A. Gametophyte Development in Ferns. Annu. Rev. Plant Biol. 1999, 50, 163–186. [Google Scholar] [CrossRef] [PubMed]
- Bartz, M.; Gola, E.M. Meristem development and activity in gametophytes of the model fern, Ceratopteris richardii. Dev. Biol. 2018, 444, 107–115. [Google Scholar] [CrossRef]
- Conway, S.J.; Di Stilio, V.S. An ontogenetic framework for functional studies in the model fern Ceratopteris richardii. Dev. Biol. 2020, 457, 20–29. [Google Scholar] [CrossRef]
- Wu, X.; Yan, A.; Mcadam SA, M.; Banks, J.A.; Zhang, S.; Zhou, Y. Timing of meristem initiation and maintenance determines the morphology of fern gametophytes. J. Exp. Bot. 2021, 72, 6990–7001. [Google Scholar] [CrossRef]
- Plackett, A.R.; Conway, S.J.; Hewett Hazelton, K.D.; Rabbinowitsch, E.H.; Langdale, J.A.; Di Stilio, V.S. LEAFY maintains apical stem cell activity during shoot development in the fern Ceratopteris richardii. eLife 2018, 7, e39625. [Google Scholar] [CrossRef] [PubMed]
- Korn, R.W. Computer simulation of the early development of the gametophyte of Dryopteris thelypteris (L.) Gray. Bot. J. Linn. Soc. 1974, 68, 163–171. [Google Scholar] [CrossRef]
- Albaum, H.G. Inhibitions Due to Growth Hormones in Fern Prothallia and Sporophytes. Am. J. Bot. 1938, 25, 124–133. [Google Scholar] [CrossRef]
- Youngstrom, C.E.; Geadelmann, L.F.; Irish, E.E.; Cheng, C.L. A fern WUSCHEL-RELATED HOMEOBOX gene functions in both gametophyte and sporophyte generations. BMC Plant Biol. 2019, 19, 416. [Google Scholar] [CrossRef] [PubMed]
- Ruegger, M.; Dewey, E.; Hobbie, L.; Brown, D.; Bernasconi, P.; Turner, J.; Muday, G.; Estelle, M. Reduced naphthylphthalamic acid binding in the tir3 mutant of Arabidopsis is associated with a reduction in polar auxin transport and diverse morphological defects. Plant Cell 1997, 9, 745–757. [Google Scholar]
- Stepanova, A.N.; Robertson-Hoyt, J.; Yun, J.; Benavente, L.M.; Xie, D.-Y.; Doležal, K.; Schlereth, A.; Jürgens, G.; Alonso, J.M. TAA1-Mediated Auxin Biosynthesis Is Essential for Hormone Crosstalk and Plant Development. Cell 2008, 133, 177–191. [Google Scholar] [CrossRef] [PubMed]
- Ishida, S.; Suzuki, H.; Iwaki, A.; Kawamura, S.; Yamaoka, S.; Kojima, M.; Takebayashi, Y.; Yamaguchi, K.; Shigenobu, S.; Sakakibara, H.; et al. Diminished Auxin Signaling Triggers Cellular Reprogramming by Inducing a Regeneration Factor in the Liverwort Marchantia polymorpha. Plant Cell Physiol. 2022, 63, 384–400. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Zhang, N.; Gao, T.; Jin, J.; Jing, T.; Wang, J.; Wu, Y.; Wan, X.; Schwab, W.; Song, C. Sesquiterpene glucosylation mediated by glucosyltransferase UGT91Q2 is involved in the modulation of cold stress tolerance in tea plants. New Phytol. 2020, 226, 362–372. [Google Scholar] [CrossRef]
- Zhao, Y. Essential Roles of Local Auxin Biosynthesis in Plant Development and in Adaptation to Environmental Changes. Annu. Rev. Plant Biol. 2018, 69, 417–435. [Google Scholar] [CrossRef]
- He, W.; Brumos, J.; Li, H.; Ji, Y.; Ke, M.; Gong, X.; Zeng, Q.; Li, W.; Zhang, X.; An, F.; et al. A small-molecule screen identifies L-kynurenine as a competitive inhibitor of TAA1/TAR activity in ethylene-directed auxin biosynthesis and root growth in Arabidopsis. Plant Cell 2011, 23, 3944–3960. [Google Scholar] [CrossRef]
- Hoffmann, O.L.; Smith, A.E. A New Group of Plant Growth Regulators. Science 1949, 109, 588. [Google Scholar] [CrossRef] [PubMed]
- Abas, L.; Kolb, M.; Stadlmann, J.; Janacek, D.P.; Lukic, K.; Schwechheimer, C.; Sazanov, L.A.; Mach, L.; Friml, J.; Hammes, U.Z. Naphthylphthalamic acid associates with and inhibits PIN auxin transporters. Proc. Natl. Acad. Sci. USA 2021, 118, e2020857118. [Google Scholar] [CrossRef] [PubMed]
- Nardmann, J.; Werr, W. The invention of WUS-like stem cell-promoting functions in plants predates leptosporangiate ferns. Plant Mol. Biol. 2012, 78, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhang, Y.; Liu, W.; Wang, H.; Wen, S.; Zhang, Y.; Xu, L. Molecular Evolution of Auxin-Mediated Root Initiation in Plants. Mol. Biol. Evol. 2020, 37, 1387–1393. [Google Scholar] [CrossRef] [PubMed]
- Youngstrom, C.E.; Withers, K.A.; Irish, E.E.; Cheng, C.-L. Vascular function of the T3/modern clade WUSCHEL-Related HOMEOBOX transcription factor genes predate apical meristem-maintenance function. BMC Plant Biol. 2022, 22, 210. [Google Scholar] [CrossRef] [PubMed]
- Hickok, L.G.; Kiriluk, R.M. Effects of Auxins on Gametophyte Development and Sexual Differentiation in the Fern Ceratopteris thalictroides (L.) Brongn. Bot. Gaz. 1984, 145, 37–42. [Google Scholar] [CrossRef]
- Tanaka, H.; Hashimoto, N.; Kawai, S.; Yumoto, E.; Shibata, K.; Tameshige, T.; Yamamoto, Y.; Sugimoto, K.; Asahina, M.; Ikeuchi, M. Auxin-Induced WUSCHEL-RELATED HOMEOBOX13 Mediates Asymmetric Activity of Callus Formation upon Cutting. Plant Cell Physiol. 2023, 64, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Jones, V.A.; Dolan, L. The evolution of root hairs and rhizoids. Ann. Bot. 2012, 110, 205–212. [Google Scholar] [CrossRef]
- Withers, K.A.; Falls, K.; Youngstrom, C.E.; Nguyen, T.; Dewald, A.; Yarvis, R.M.; Simons, G.P.; Flanagan, R.; Bui, L.T.; Irish, E.E.; et al. A Ceratopteris EXCESS MICROSPOROCYTES1 suppresses reproductive transition in the fern vegetative leaves. Plant Sci. 2023, 335, 111812. [Google Scholar] [CrossRef]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Connor, R.; Funk, K.; Kelly, C.; Kim, S.; et al. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2022, 50, D20–D26. [Google Scholar] [CrossRef]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef] [PubMed]
- Li, F.W.; Brouwer, P.; Carretero-Paulet, L.; Cheng, S.; De Vries, J.; Delaux, P.M.; Eily, A.; Koppers, N.; Kuo, L.Y.; Li, Z.; et al. Fern genomes elucidate land plant evolution and cyanobacterial symbioses. Nat. Plants 2018, 4, 460–472. [Google Scholar] [CrossRef] [PubMed]
- Berardini, T.Z.; Reiser, L.; Li, D.; Mezheritsky, Y.; Muller, R.; Strait, E.; Huala, E. The arabidopsis information resource: Making and mining the “gold standard” annotated reference plant genome. Genesis 2015, 53, 474–485. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin DM, A.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Hejatko, J.; Blilou, I.; Brewer, P.B.; Friml, J.; Scheres, B.; Benkova, E. In situ hybridization technique for mRNA detection in whole mount Arabidopsis samples. Nat. Protoc. 2006, 1, 1939–1946. [Google Scholar] [CrossRef]
- GRAPHPAD-SOFTWARE 2023; GraphPad Prism: Boston, MA, USA, 2023; Available online: www.graphpad.com (accessed on 27 September 2023).





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Withers, K.A.; Kvamme, A.; Youngstrom, C.E.; Yarvis, R.M.; Orpano, R.; Simons, G.P.; Irish, E.E.; Cheng, C.-L. Auxin Involvement in Ceratopteris Gametophyte Meristem Regeneration. Int. J. Mol. Sci. 2023, 24, 15832. https://doi.org/10.3390/ijms242115832
Withers KA, Kvamme A, Youngstrom CE, Yarvis RM, Orpano R, Simons GP, Irish EE, Cheng C-L. Auxin Involvement in Ceratopteris Gametophyte Meristem Regeneration. International Journal of Molecular Sciences. 2023; 24(21):15832. https://doi.org/10.3390/ijms242115832
Chicago/Turabian StyleWithers, Kelley A., Athena Kvamme, Christopher E. Youngstrom, Rebekah M. Yarvis, Rachel Orpano, Gabriel P. Simons, Erin E. Irish, and Chi-Lien Cheng. 2023. "Auxin Involvement in Ceratopteris Gametophyte Meristem Regeneration" International Journal of Molecular Sciences 24, no. 21: 15832. https://doi.org/10.3390/ijms242115832
APA StyleWithers, K. A., Kvamme, A., Youngstrom, C. E., Yarvis, R. M., Orpano, R., Simons, G. P., Irish, E. E., & Cheng, C.-L. (2023). Auxin Involvement in Ceratopteris Gametophyte Meristem Regeneration. International Journal of Molecular Sciences, 24(21), 15832. https://doi.org/10.3390/ijms242115832