
Mechanism of Salt Tolerance and Plant Growth Promotion in Priestia megaterium ZS-3 Revealed by Cellular Metabolism and Whole-Genome Studies
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
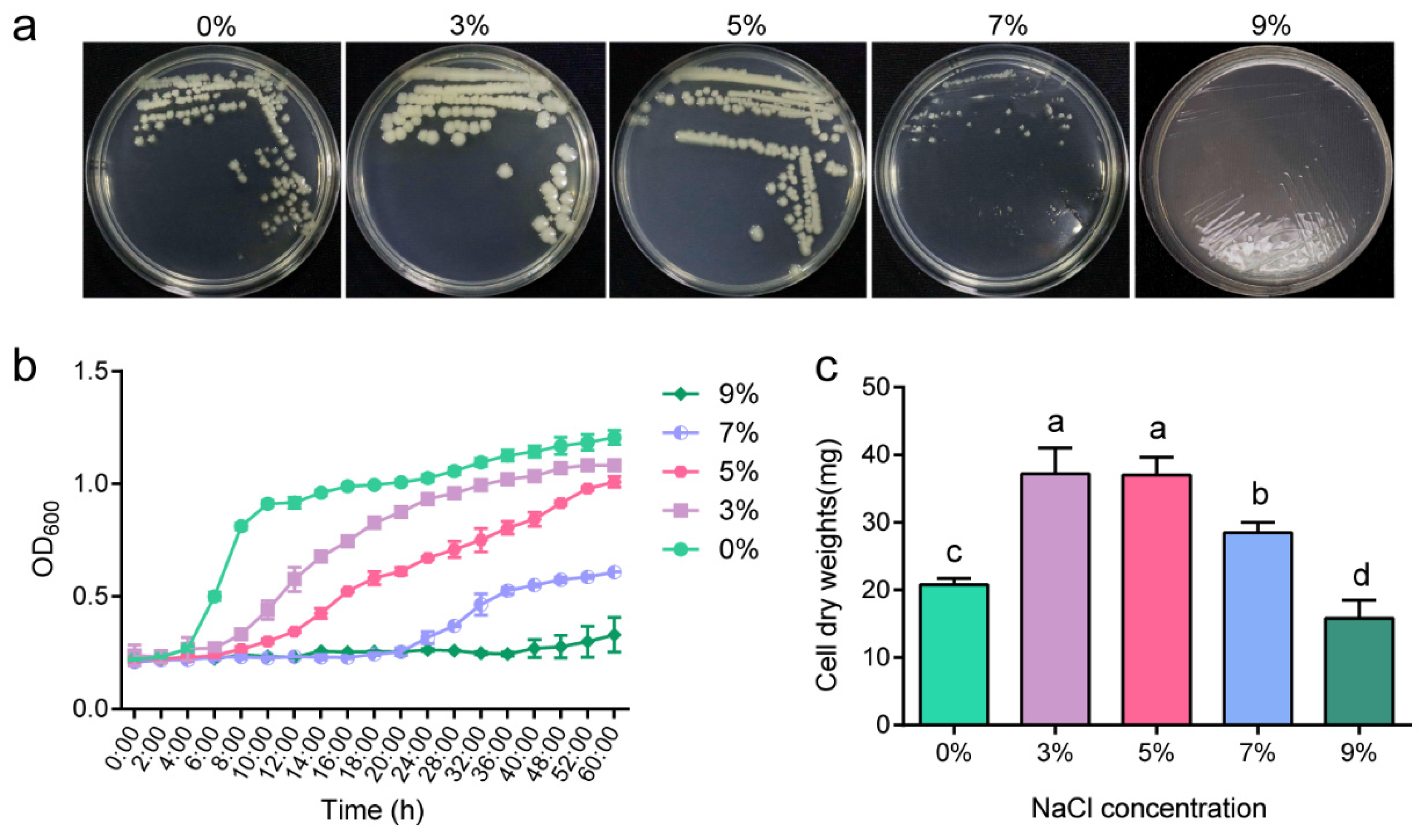
2.1. Salt-Tolerant Growth Characteristics of P. megaterium ZS-3
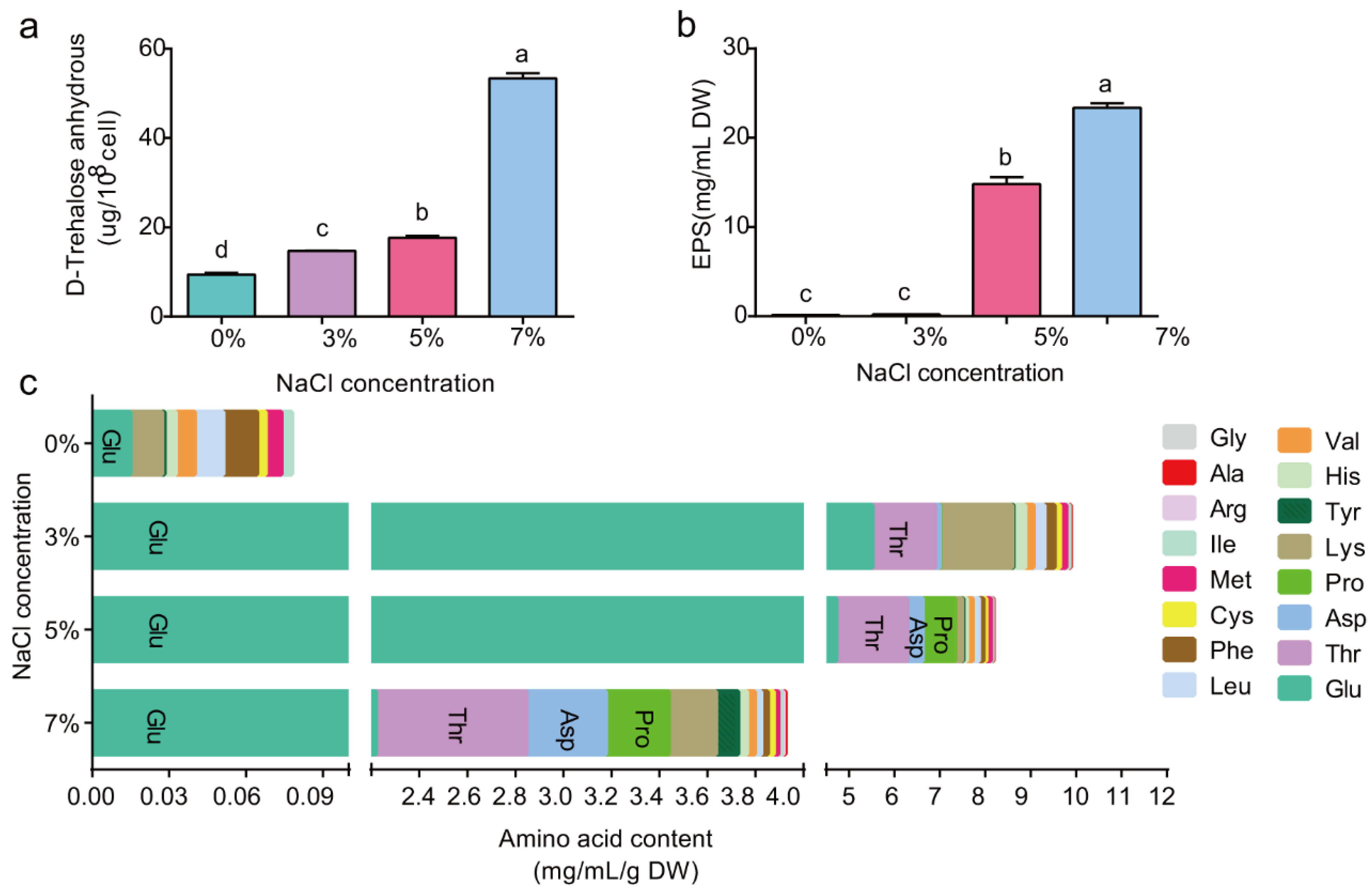
2.2. High Salt Causes P. megaterium ZS-3 Metabolites to Accumulate
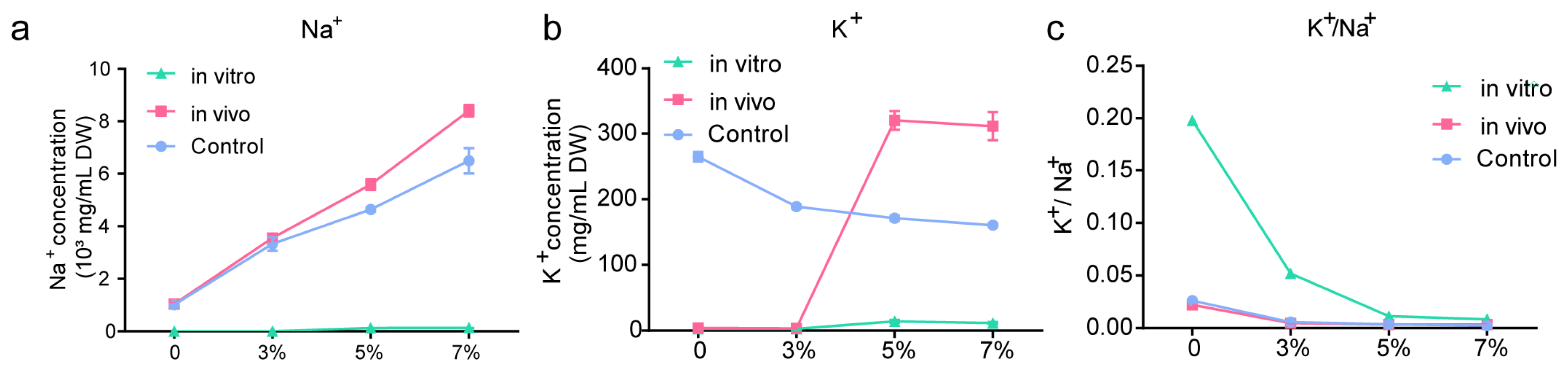
2.3. Ion Content In Vivo and In Vitro
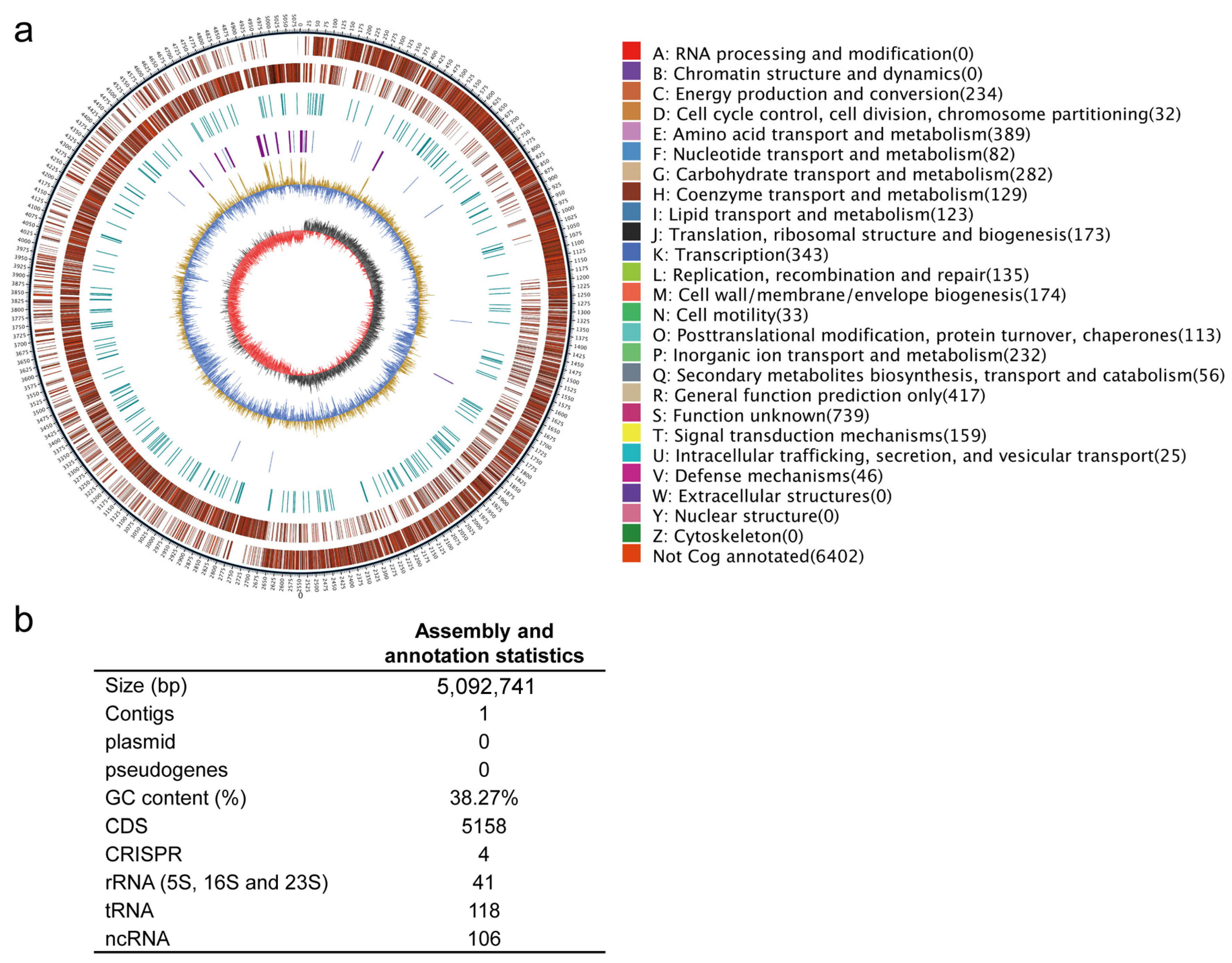
2.4. General Characteristics of the Whole-Genome Sequence of P. megaterium ZS-3
2.5. Genome Mining for Stress Survival/Alleviation
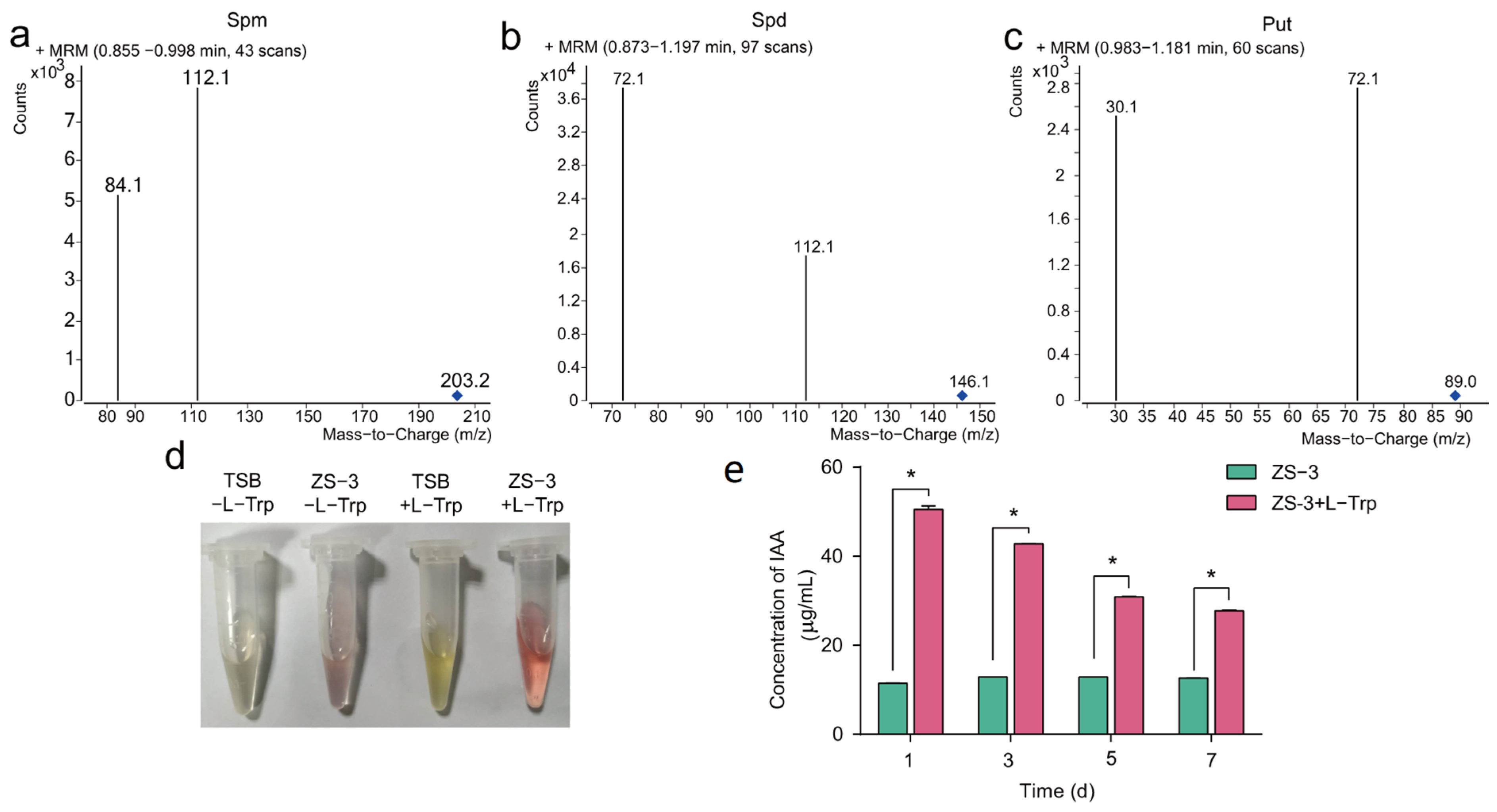
2.6. Genes Involved in Pgp
2.7. Biocontrol Activities
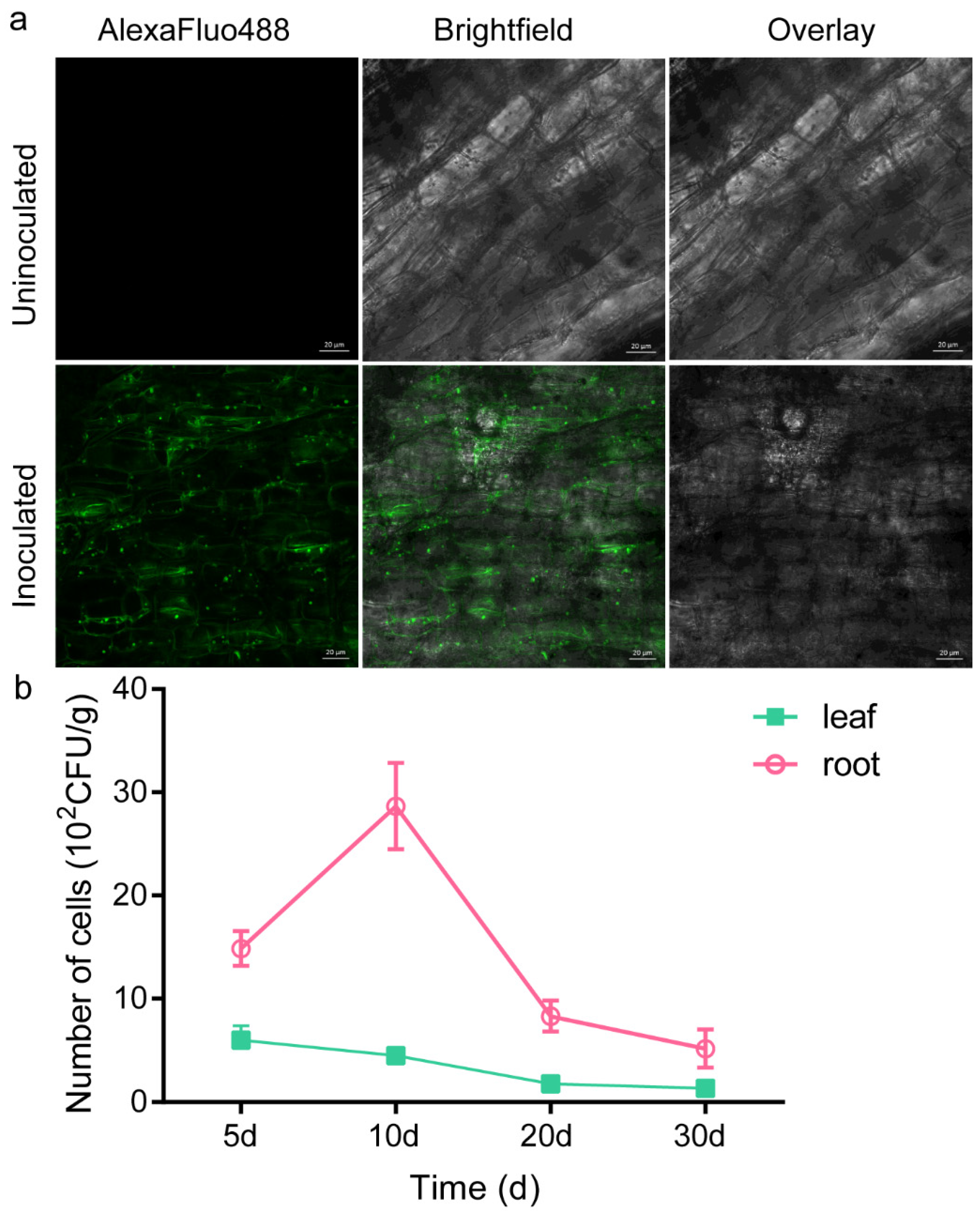
2.8. Root Colonisation Study: GFP Marker Analysis
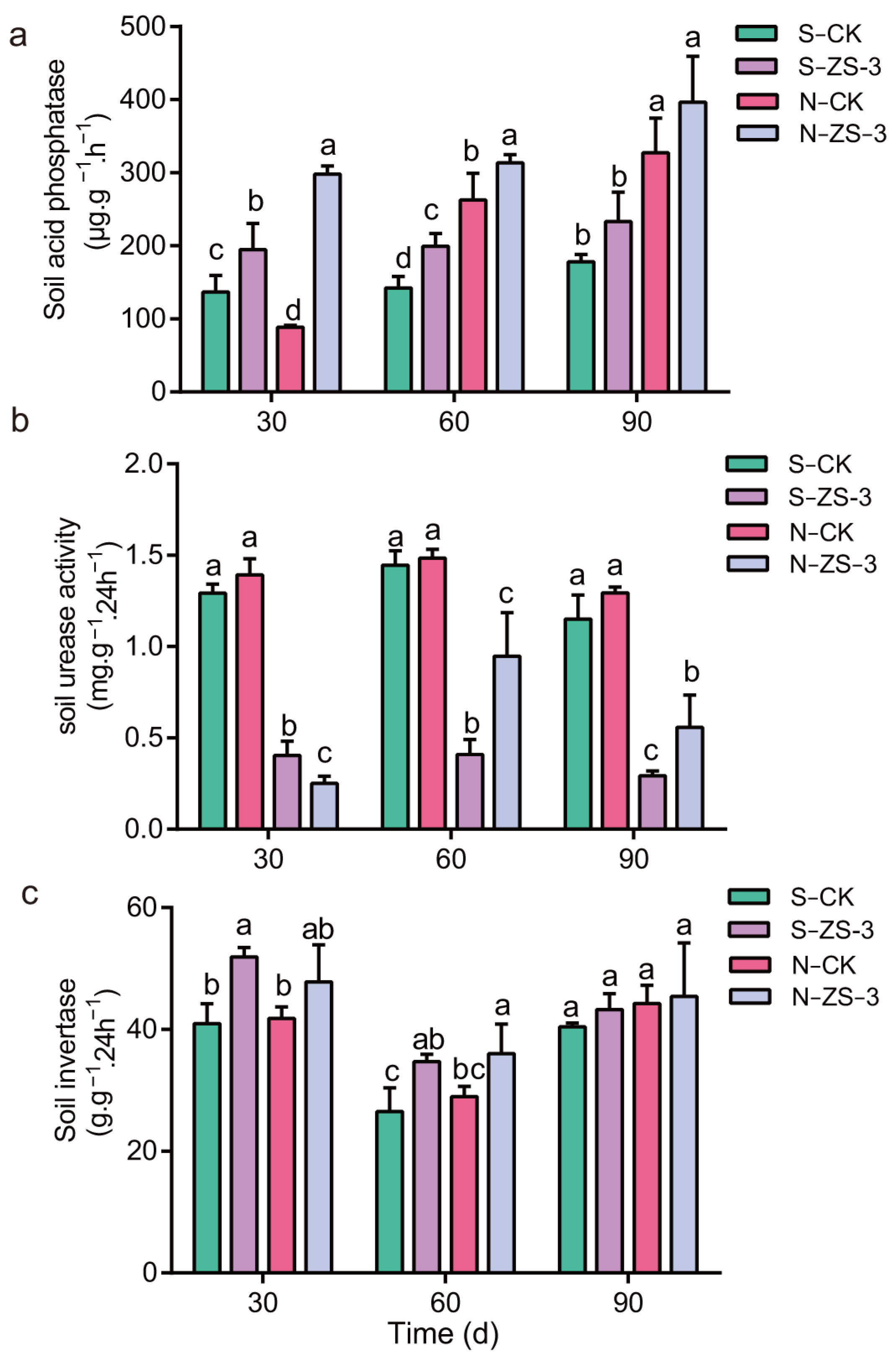
2.9. Regulation of Soil Enzyme Activity by P. megaterium ZS-3
3. Discussion
4. Materials and Methods
4.1. Bacterial Cultures and Inoculation
4.2. Extraction and Measurement of Intracellular Trehalose
4.3. Extraction and Measurement of the Intracellular Free aa Content
4.4. Determination of the EPS Content
4.5. Measurement of the Intracellular Na+ and K+ Ion Content
4.6. Determination of the Polyamine Content
4.7. Phytohormone Quantification and Visualisation by the Salkowski Method
4.8. Root Colonisation Assay
4.9. Estimation of Soil Enzyme Activity
4.10. Genome Sequencing and Bioinformatic Analysis
4.11. Data Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abbas, R.; Rasul, S.; Aslam, K.; Baber, M.; Shahid, M.; Mubeen, F.; Naqqash, T. Halotolerant PGPR: A hope for cultivation of saline soils. J. King Saud Univ.-Sci. 2019, 31, 1195–1201. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Finkel, O.M.; Castrillo, G.; Paredes, S.H.; González, I.S.; Dangl, J.L. Understanding and exploiting plant beneficial microbes. Curr. Opin. Plant Biol. 2017, 38, 155–163. [Google Scholar] [CrossRef]
- Kumar, A.; Patel, J.S.; Meena, V.S.; Srivastava, R. Recent advances of PGPR based approaches for stress tolerance in plants for sustainable agriculture. Biocatal. Agric. Biotechnol. 2019, 20, 101271. [Google Scholar] [CrossRef]
- Saghafi, D.; Delangiz, N.; Lajayer, B.A.; Ghorbanpour, M. An overview on improvement of crop productivity in saline soils by halotolerant and halophilic PGPRs. 3 Biotech 2019, 9, 261. [Google Scholar] [CrossRef]
- Ramadoss, D.; Lakkineni, V.K.; Bose, P.; Ali, S.; Annapurna, K. Mitigation of salt stress in wheat seedlings by halotolerant bacteria isolated from saline habitats. SpringerPlus 2013, 2, 6. [Google Scholar] [CrossRef]
- Oren, A. Salinity: Environment-Plants-Molecules. In Adaptation of Halophilic Archaea to Life at High Salt Concentrations; Läuchli, A., Lüttge, U., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 81–96. [Google Scholar]
- Schroeter, R.; Hoffmann, T.; Voigt, B.; Meyer, H.; Bleisteiner, M.; Muntel, J.; Jürgen, B.; Albrecht, D.; Becher, D.; Lalk, M.; et al. Stress Responses of the Industrial Workhorse Bacillus licheniformis to Osmotic Challenges. PLoS ONE 2013, 8, e80956. [Google Scholar] [CrossRef] [PubMed]
- Brill, J.; Hoffmann, T.; Bleisteiner, M.; Bremer, E. Osmotically Controlled Synthesis of the Compatible Solute Proline Is Critical for Cellular Defense of Bacillus subtilis against High Osmolarity. J. Bacteriol. 2011, 193, 5335–5346. [Google Scholar] [CrossRef] [PubMed]
- Dave, S.R.; Vaishnav, A.M.; Upadhyay, K.H.; Tipre, D.R. Microbial Exopolysaccharide—An Inevitable Product for Living Beings and Environment. J. Bacteriol. Mycol. Open Access 2016, 2, 109–111. [Google Scholar] [CrossRef]
- Srivastava, S.; Sharma, S. Insight into Exopolysaccharide-Mediated Stress Tolerance in Plants: A Feasible Approach Towards the Development of Next-Generation Bioformulations. J. Soil Sci. Plant Nutr. 2023, 23, 22–33. [Google Scholar] [CrossRef]
- Romantsov, T.; Guan, Z.; Wood, J.M. Cardiolipin and the osmotic stress responses of bacteria. Biochim. Biophys. Acta (BBA)-Biomembr. 2009, 1788, 2092–2100. [Google Scholar] [CrossRef] [PubMed]
- Biedendieck, R.; Knuuti, T.; Moore, S.J.; Jahn, D. The “beauty in the beast”—The multiple uses of Priestia megaterium in biotechnology. Appl. Microbiol. Biotechnol. 2021, 105, 5719–5737. [Google Scholar] [CrossRef] [PubMed]
- Motaleb, N.A.A.; Elhady, S.A.A.; Ghoname, A.A. AMF and Bacillus megaterium Neutralize the Harmful Effects of Salt Stress On Bean Plants. Gesunde Pflanz. 2020, 72, 29–39. [Google Scholar] [CrossRef]
- Vásconez, R.D.A.; Moya, E.M.T.; Yépez, L.A.C.; Chiluisa-Utreras, V.P.; Suquillo, I.d.L.Á.V. Evaluation of Bacillus megaterium strain AB4 as a potential biocontrol agent of Alternaria japonica, a mycopathogen of Brassica oleracea var. italica. Biotechnol. Rep. 2020, 26, e00454. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhang, D.; Chu, S.; Zhi, Y.; Liu, X.; Zhou, P. Genomic Analysis of Bacillus megaterium NCT-2 Reveals Its Genetic Basis for the Bioremediation of Secondary Salinization Soil. Int. J. Genom. 2020, 2020, 4109186. [Google Scholar] [CrossRef]
- Shi, L.-N.; Lu, L.-X.; Ye, J.-R.; Shi, H.-M. The Endophytic Strain ZS-3 Enhances Salt Tolerance in Arabidopsis thaliana by Regulating Photosynthesis, Osmotic Stress, and Ion Homeostasis and Inducing Systemic Tolerance. Front. Plant Sci. 2022, 13, 820837. [Google Scholar] [CrossRef]
- Aminian-Dehkordi, J.; Mousavi, S.M.; Jafari, A.; Mijakovic, I.; Marashi, S.-A. Manually curated genome-scale reconstruction of the metabolic network of Bacillus megaterium DSM319. Sci. Rep. 2019, 9, 18762. [Google Scholar] [CrossRef]
- Zou, W.; Zhou, M.; Liu, L.; Chen, J. Reconstruction and analysis of the industrial strain Bacillus megaterium WSH002 genome-scale in silico metabolic model. J. Biotechnol. 2013, 164, 503–509. [Google Scholar] [CrossRef]
- Maghembe, R.S.; Mdoe, F.P.; Makaranga, A.; Mpemba, J.A.; Mark, D.; Mlay, C.; Moto, E.A.; Mtewa, A.G. Complete genome sequence data of Priestia megaterium strain MARUCO02 isolated from marine mangrove-inhabited sediments of the Indian Ocean in the Bagamoyo Coast. Data Brief 2023, 48, 109119. [Google Scholar] [CrossRef]
- Carles, L.; Donnadieu, F.; Wawrzyniak, I.; Besse-Hoggan, P.; Batisson, I. Genomic analysis of the Bacillus megaterium Mes11: New insights into nitroreductase genes associated with the degradation of mesotrione. Int. Biodeterior. Biodegrad. 2021, 162, 105254. [Google Scholar] [CrossRef]
- Feng, N.-X.; Yu, J.; Mo, C.-H.; Zhao, H.-M.; Li, Y.-W.; Wu, B.-X.; Cai, Q.-Y.; Li, H.; Zhou, D.-M.; Wong, M.-H. Biodegradation of di-n-butyl phthalate (DBP) by a novel endophytic Bacillus megaterium strain YJB3. Sci. Total. Environ. 2018, 616–617, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Vary, P.S.; Biedendieck, R.; Fuerch, T.; Meinhardt, F.; Rohde, M.; Deckwer, W.-D.; Jahn, D. Bacillus megaterium—From simple soil bacterium to industrial protein production host. Appl. Microbiol. Biotechnol. 2007, 76, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Nau-Wagner, G.; Opper, D.; Rolbetzki, A.; Boch, J.; Kempf, B.; Hoffmann, T.; Bremer, E. Genetic Control of Osmoadaptive Glycine Betaine Synthesis in Bacillus subtilis through the Choline-Sensing and Glycine Betaine-Responsive GbsR Repressor. J. Bacteriol. 2012, 194, 2703–2714. [Google Scholar] [CrossRef] [PubMed]
- Belaouni, H.A.; Compant, S.; Antonielli, L.; Nikolic, B.; Zitouni, A.; Sessitsch, A. In-depth genome analysis of Bacillus sp. BH32, a salt stress-tolerant endophyte obtained from a halophyte in a semiarid region. Appl. Microbiol. Biotechnol. 2022, 106, 3113–3137. [Google Scholar] [CrossRef]
- Huang, J.; Qiao, Z.X.; Tang, J.W.; Wang, G. High quality draft genome sequence of the moderately halophilic bacterium Pontibacillus yanchengensis Y32(T) and comparison among Pontibacillus genomes. Stand. Genom. Sci. 2015, 10, 93. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Y.; Ma, Y.; Wang, X.; Zhang, B.; Zhang, G.; Bahadur, A.; Chen, T.; Liu, G.; Zhang, W.; et al. Research progress regarding the role of halophilic and halotolerant microorganisms in the eco-environmental sustainability and conservation. J. Clean. Prod. 2023, 418, 138054. [Google Scholar] [CrossRef]
- Ventosa, A.; Nieto, J.J.; Oren, A. Biology of Moderately Halophilic Aerobic Bacteria. Microbiol. Mol. Biol. Rev. 1998, 62, 504–544. [Google Scholar] [CrossRef]
- Rangseekaew, P.; Barros-Rodríguez, A.; Pathom-Aree, W.; Manzanera, M. Plant Beneficial Deep-Sea Actinobacterium, Dermacoccus abyssi MT1.1T Promote Growth of Tomato (Solanum lycopersicum) under Salinity Stress. Biology 2022, 11, 191. [Google Scholar] [CrossRef]
- Krulwich, T.A.; Hicks, D.B.; Ito, M. Cation/proton antiporter complements of bacteria: Why so large and diverse? Mol. Microbiol. 2009, 74, 257–260. [Google Scholar] [CrossRef]
- Zamanzadeh-Nasrabadi, S.M.; Mohammadiapanah, F.; Sarikhan, S.; Shariati, V.; Saghafi, K.; Hosseini-Mazinani, M. Comprehensive genome analysis of Pseudomonas sp. SWRIQ11, a new plant growth-promoting bacterium that alleviates salinity stress in olive. 3 Biotech 2023, 13, 347. [Google Scholar] [CrossRef]
- Haroon, U.; Khizar, M.; Liaquat, F.; Ali, M.; Akbar, M.; Tahir, K.; Batool, S.S.; Kamal, A.; Chaudhary, H.J.; Munis, M.F.H. Halotolerant Plant Growth-Promoting Rhizobacteria Induce Salinity Tolerance in Wheat by Enhancing the Expression of SOS Genes. J. Plant Growth Regul. 2022, 41, 2435–2448. [Google Scholar] [CrossRef]
- Liu, X.; Chai, J.; Zhang, Y.; Zhang, C.; Lei, Y.; Li, Q.; Yao, T. Halotolerant rhizobacteria mitigate the effects of salinity stress on maize growth by secreting exopolysaccharides. Environ. Exp. Bot. 2022, 204, 105098. [Google Scholar] [CrossRef]
- López-Ortega, M.A.; Chavarría-Hernández, N.; del Rocio Lopez-Cuellar, M.; Rodríguez-Hernández, A.I. A review of extracellular polysaccharides from extreme niches: An emerging natural source for the biotechnology. From the adverse to diverse! Int. J. Biol. Macromol. 2021, 177, 559–577. [Google Scholar] [CrossRef] [PubMed]
- Sandhya, V.; Ali, S.Z. The production of exopolysaccharide by Pseudomonas putida GAP-P45 under various abiotic stress conditions and its role in soil aggregation. Microbiology 2015, 84, 512–519. [Google Scholar] [CrossRef]
- Duca, D.; Lorv, J.; Patten, C.L.; Rose, D.; Glick, B.R. Indole-3-acetic acid in plant–microbe interactions. Antonie Van Leeuwenhoek 2014, 106, 85–125. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.-H.; Chen, F.-F.; Wang, C.-E.; Fu, H.-H.; Fang, X.-Q.; Ye, J.-R.; Shi, J.-Y. Indole-3-Acetic Acid in Burkholderia pyrrocinia JK-SH007: Enzymatic Identification of the Indole-3-Acetamide Synthesis Pathway. Front. Microbiol. 2019, 10, 2559. [Google Scholar] [CrossRef]
- Boss, B.L.; Wanees, A.E.; Zaslow, S.J.; Normile, T.G.; Izquierdo, J.A. Comparative genomics of the plant-growth promoting bacterium Sphingobium sp. strain AEW4 isolated from the rhizosphere of the beachgrass Ammophila breviligulata. BMC Genom. 2022, 23, 508. [Google Scholar] [CrossRef]
- Zhang, P.; Jin, T.; Sahu, S.K.; Xu, J.; Shi, Q.; Liu, H.; Wang, Y. The Distribution of Tryptophan-Dependent Indole-3-Acetic Acid Synthesis Pathways in Bacteria Unraveled by Large-Scale Genomic Analysis. Molecules 2019, 24, 1411. [Google Scholar] [CrossRef]
- Naveed, M.; Qureshi, M.A.; Zahir, Z.A.; Hussain, M.B.; Sessitsch, A.; Mitter, B. L-Tryptophan-dependent biosynthesis of indole-3-acetic acid (IAA) improves plant growth and colonization of maize by Burkholderia phytofirmans PsJN. Ann. Microbiol. 2015, 65, 1381–1389. [Google Scholar] [CrossRef]
- Ali, B.; Sabri, A.N.; Ljung, K.; Hasnain, S. Quantification of indole-3-acetic acid from plant associated Bacillus spp. and their phytostimulatory effect on Vigna radiata (L.). World J. Microbiol. Biotechnol. 2009, 25, 519–526. [Google Scholar] [CrossRef]
- Zhu, X.; Liao, J.; Xia, X.; Xiong, F.; Li, Y.; Shen, J.; Wen, B.; Ma, Y.; Wang, Y.; Fang, W. Physiological and iTRAQ-based proteomic analyses reveal the function of exogenous γ-aminobutyric acid (GABA) in improving tea plant (Camellia sinensis L.) tolerance at cold temperature. BMC Plant Biol. 2019, 19, 43. [Google Scholar] [CrossRef]
- Kamilova, F.; Kravchenko, L.V.; Shaposhnikov, A.I.; Azarova, T.; Makarova, N.; Lugtenberg, B.; Liu, X.; Xie, Z.; Wang, Y.; Sun, Y.; et al. Organic Acids, Sugars, and l-Tryptophane in Exudates of Vegetables Growing on Stonewool and Their Effects on Activities of Rhizosphere Bacteria. Mol. Plant-Microbe Interact. 2006, 19, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Jangra, A.; Chaturvedi, S.; Kumar, N.; Singh, H.; Sharma, V.; Thakur, M.; Tiwari, S.; Chhokar, V. Polyamines: The Gleam of Next-Generation Plant Growth Regulators for Growth, Development, Stress Mitigation, and Hormonal Crosstalk in Plants—A Systematic Review. J. Plant Growth Regul. 2022, 42, 5167–5191. [Google Scholar] [CrossRef]
- Moselhy, S.S.; Asami, T.; Abualnaja, K.O.; Al-Malki, A.L.; Yamano, H.; Akiyama, T.; Wada, R.; Yamagishi, T.; Hikosaka, M.; Iwakawa, J.; et al. Spermidine, a polyamine, confers resistance to rice blast. J. Pestic. Sci. 2016, 41, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.R.; Jalil, S.U.; Chopra, P.; Chhillar, H.; Ferrante, A.; Khan, N.A.; Ansari, M.I. Role of GABA in plant growth, development and senescence. Plant Gene 2021, 26, 100283. [Google Scholar] [CrossRef]
- Zhu, J.; Li, C.; Sun, L.; Cheng, Y.; Hou, J.; Fan, Y.; Ge, Y. Application of γ-aminobutyric acid induces disease resistance in apples through regulation of polyamine metabolism, GABA shunt and reactive oxygen species metabolism. Sci. Hortic. 2022, 291, 110588. [Google Scholar] [CrossRef]
- Akçay, N.; Bor, M.; Karabudak, T.; Özdemir, F.; Türkan, I. Contribution of Gamma amino butyric acid (GABA) to salt stress responses of Nicotiana sylvestris CMSII mutant and wild type plants. J. Plant Physiol. 2012, 169, 452–458. [Google Scholar] [CrossRef]
- Vijayakumari, K.; Puthur, J.T. γ-Aminobutyric acid (GABA) priming enhances the osmotic stress tolerance in Piper nigrum Linn. plants subjected to PEG-induced stress. Plant Growth Regul. 2016, 78, 57–67. [Google Scholar] [CrossRef]
- Ramesh, S.A.; Tyerman, S.D.; Gilliham, M.; Xu, B. γ-Aminobutyric acid (GABA) signalling in plants. Cell. Mol. Life Sci. 2017, 74, 1577–1603. [Google Scholar] [CrossRef]
- Ryu, C.-M.; Farag, M.A.; Hu, C.-H.; Reddy, M.S.; Wei, H.-X.; Paré, P.W.; Kloepper, J.W. Bacterial volatiles promote growth in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 4927–4932. [Google Scholar] [CrossRef]
- Dimkić, I.; Janakiev, T.; Petrović, M.; Degrassi, G.; Fira, D. Plant-associated Bacillus and Pseudomonas antimicrobial activities in plant disease suppression via biological control mechanisms—A review. Physiol. Mol. Plant Pathol. 2022, 117, 101754. [Google Scholar] [CrossRef]
- Ghazy, N.; El-Nahrawy, S. Siderophore production by Bacillus subtilis MF497446 and Pseudomonas koreensis MG209738 and their efficacy in controlling Cephalosporium maydis in maize plant. Arch. Microbiol. 2021, 203, 1195–1209. [Google Scholar] [CrossRef]
- Dimopoulou, A.; Theologidis, I.; Benaki, D.; Koukounia, M.; Zervakou, A.; Tzima, A.; Diallinas, G.; Hatzinikolaou, D.G.; Skandalis, N. Direct Antibiotic Activity of Bacillibactin Broadens the Biocontrol Range of Bacillus amyloliquefaciens MBI600. mSphere 2021, 6, e0037621. [Google Scholar] [CrossRef]
- Ali, I.; Sultan, T.; Subhan, F.; Haleem, K.S.; Sultana, N.; Tauseef, I. PGPRs of plum (Prunus domestica) rhizosphere enhance plant growth and antagonise fungal activityin vitro. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2018, 68, 367–378. [Google Scholar] [CrossRef]
- Kong, W.-L.; Wang, W.-Y.; Zuo, S.-H.; Wu, X.-Q. Genome Sequencing of Rahnella victoriana JZ-GX1 Provides New Insights into Molecular and Genetic Mechanisms of Plant Growth Promotion. Front. Microbiol. 2022, 13, 828990. [Google Scholar] [CrossRef] [PubMed]
- Rikame, T.; Borde, M. Whole Genome, Functional Annotation and Comparative Genomics of Plant Growth-Promoting Bacteria Pseudomonas aeruginosa (NG61) with Potential Application in Agro-Industry. Curr. Microbiol. 2022, 79, 169. [Google Scholar] [CrossRef]
- You, M.; Fang, S.; MacDonald, J.; Xu, J.; Yuan, Z.-C. Isolation and characterization of Burkholderia cenocepacia CR318, a phosphate solubilizing bacterium promoting corn growth. Microbiol. Res. 2020, 233, 126395. [Google Scholar] [CrossRef]
- Sharma, S.; Gang, S.; Schumacher, J.; Buck, M.; Saraf, M. Genomic appraisal of Klebsiella PGPB isolated from soil to enhance the growth of barley. Genes Genom. 2021, 43, 869–883. [Google Scholar] [CrossRef]
- Bach, E.; Rangel, C.P.; Ribeiro, I.D.A.; Passaglia, L.M.P. Pangenome analyses of Bacillus pumilus, Bacillus safensis, and Priestia megaterium exploring the plant-associated features of bacilli strains isolated from canola. Mol. Genet. Genom. 2022, 297, 1063–1079. [Google Scholar] [CrossRef]
- Ribeiro, I.D.A.; Bach, E.; da Silva Moreira, F.; Müller, A.R.; Rangel, C.P.; Wilhelm, C.M.; Barth, A.L.; Passaglia, L.M.P. Antifungal potential against Sclerotinia sclerotiorum (Lib.) de Bary and plant growth promoting abilities of Bacillus isolates from canola (Brassica napus L.) roots. Microbiol. Res. 2021, 248, 126754. [Google Scholar] [CrossRef]
- Eppinger, M.; Bunk, B.; Johns, M.A.; Edirisinghe, J.N.; Kutumbaka, K.K.; Koenig, S.S.K.; Creasy, H.H.; Rosovitz, M.J.; Riley, D.R.; Daugherty, S.; et al. Genome Sequences of the Biotechnologically Important Bacillus megaterium Strains QM B1551 and DSM319. J. Bacteriol. 2011, 193, 4199–4213. [Google Scholar] [CrossRef]
- Wang, Z.; Lian, H.; Sun, Y.; Ma, Z.; Tian, P.; Qi, H.; Jiang, Y. Effects of Straw Return Depth on Soil Organic Carbon, Nitrogen Content, and Soil Enzyme Activity of Spring Maize Field. J. Agric. Resour. Environ. 2021, 4, 636–646. [Google Scholar]
- Yang, W.-N.; Yu, L.; Luo, D.-H.; Xiong, Z.-Y.; Wang, Y.-Y.; Xu, M.; Wang, Z.-F.; Gao, M. Effect of Combined Application of Biochar with Chemical Fertilizer and Organic Fertilizer on Soil Phosphatase Activity and Microbial Community. Environ. Sci. 2022, 43, 540–549. [Google Scholar] [CrossRef]
- Haddar, A.; Hamed, M.; Bouallegue, A.; Bastos, R.; Coelho, E.; Coimbra, M.A. Structural elucidation and interfacial properties of a levan isolated from Bacillus mojavensis. Food Chem. 2021, 343, 128456. [Google Scholar] [CrossRef] [PubMed]
- Sahar, S.M.; Shaimaa, K.A.; Manal, S.S.; Hala, M.R. Characterization and applications of exopolysaccharide produced by marine Bacillus altitudinis MSH2014 from Ras Mohamed, Sinai, Egypt. Egypt. J. Basic Appl. Sci. 2018, 5, 204–209. [Google Scholar] [CrossRef]
- Nagata, S.; Sasaki, H.; Oshima, A.; Takeda, S.; Hashimoto, Y.; Ishida, A. Effect of Proline and K+on the Stimulation of Cellular Activities in Escherichia coli K-12 under High Salinity. Biosci. Biotechnol. Biochem. 2005, 69, 740–746. [Google Scholar] [CrossRef][Green Version]
- Santos, J.T.D.C.; Petry, F.C.; Tobaruela, E.d.C.; Mercadante, A.Z.; Gloria, M.B.A.; Costa, A.M.; Lajolo, F.M.; Hassimotto, N.M.A. Brazilian native passion fruit (Passiflora tenuifila Killip) is a rich source of proanthocyanidins, carotenoids, and dietary fiber. Food Res. Int. 2021, 147, 110521. [Google Scholar] [CrossRef]
- Xie, S.-S.; Wu, H.-J.; Zang, H.-Y.; Wu, L.-M.; Zhu, Q.-Q.; Gao, X.-W. Plant Growth Promotion by Spermidine-Producing Bacillus subtilis OKB105. Mol. Plant-Microbe Interact. 2014, 27, 655–663. [Google Scholar] [CrossRef]
- Fett, W.F.; Osman, S.F.; Dunn, M.F. Auxin Production by Plant-Pathogenic Pseudomonads and Xanthomonads. Appl. Environ. Microbiol. 1987, 53, 1839–1845. [Google Scholar] [CrossRef]
- Meinhardt, F.; Stahl, U.; Ebeling, W. Highly efficient expression of homologous and heterologous genes in Bacillus megaterium. Appl. Microbiol. Biotechnol. 1989, 30, 343–350. [Google Scholar] [CrossRef]
- Chu, S.; Zhang, D.; Zhi, Y.; Wang, B.; Chi, C.-P.; Zhang, D.; Liu, Y.; Zhou, P. Enhanced removal of nitrate in the maize rhizosphere by plant growth-promoting Bacillus megaterium NCT-2, and its colonization pattern in response to nitrate. Chemosphere 2018, 208, 316–324. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, L.; Zhu, X.; Qian, T.; Du, J.; Du, Y.; Ye, J. Mechanism of Salt Tolerance and Plant Growth Promotion in Priestia megaterium ZS-3 Revealed by Cellular Metabolism and Whole-Genome Studies. Int. J. Mol. Sci. 2023, 24, 15751. https://doi.org/10.3390/ijms242115751
Shi L, Zhu X, Qian T, Du J, Du Y, Ye J. Mechanism of Salt Tolerance and Plant Growth Promotion in Priestia megaterium ZS-3 Revealed by Cellular Metabolism and Whole-Genome Studies. International Journal of Molecular Sciences. 2023; 24(21):15751. https://doi.org/10.3390/ijms242115751
Chicago/Turabian StyleShi, Lina, Xiaoxia Zhu, Ting Qian, Jiazhou Du, Yuanyuan Du, and Jianren Ye. 2023. "Mechanism of Salt Tolerance and Plant Growth Promotion in Priestia megaterium ZS-3 Revealed by Cellular Metabolism and Whole-Genome Studies" International Journal of Molecular Sciences 24, no. 21: 15751. https://doi.org/10.3390/ijms242115751
APA StyleShi, L., Zhu, X., Qian, T., Du, J., Du, Y., & Ye, J. (2023). Mechanism of Salt Tolerance and Plant Growth Promotion in Priestia megaterium ZS-3 Revealed by Cellular Metabolism and Whole-Genome Studies. International Journal of Molecular Sciences, 24(21), 15751. https://doi.org/10.3390/ijms242115751