
The Impact of p70S6 Kinase-Dependent Phosphorylation of Gemin2 in UsnRNP Biogenesis
 , ,
, ,  , ,
, , 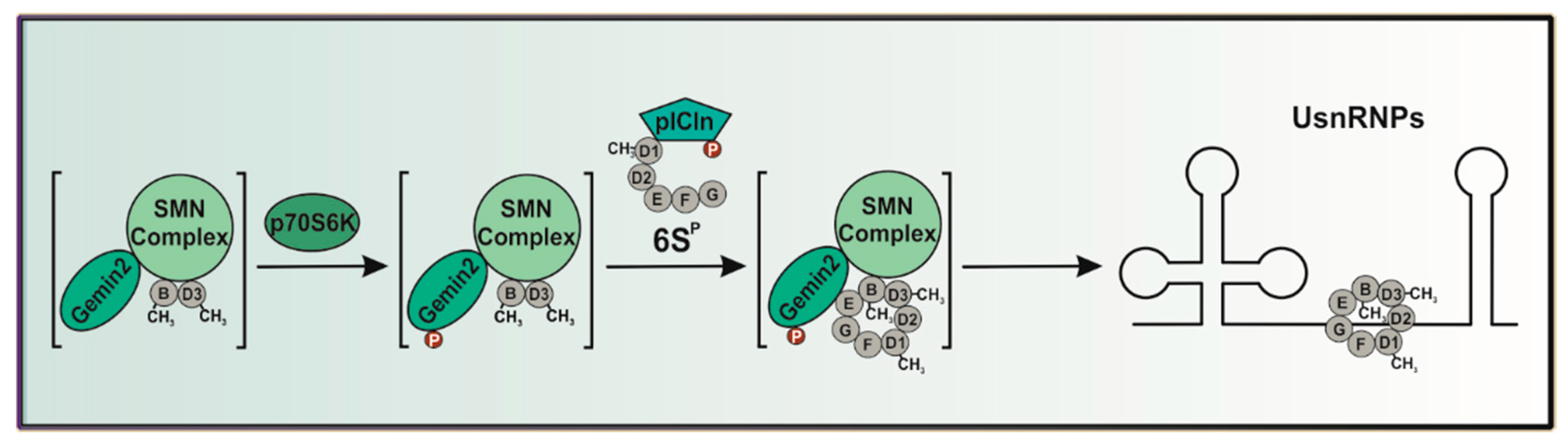{kind=link}
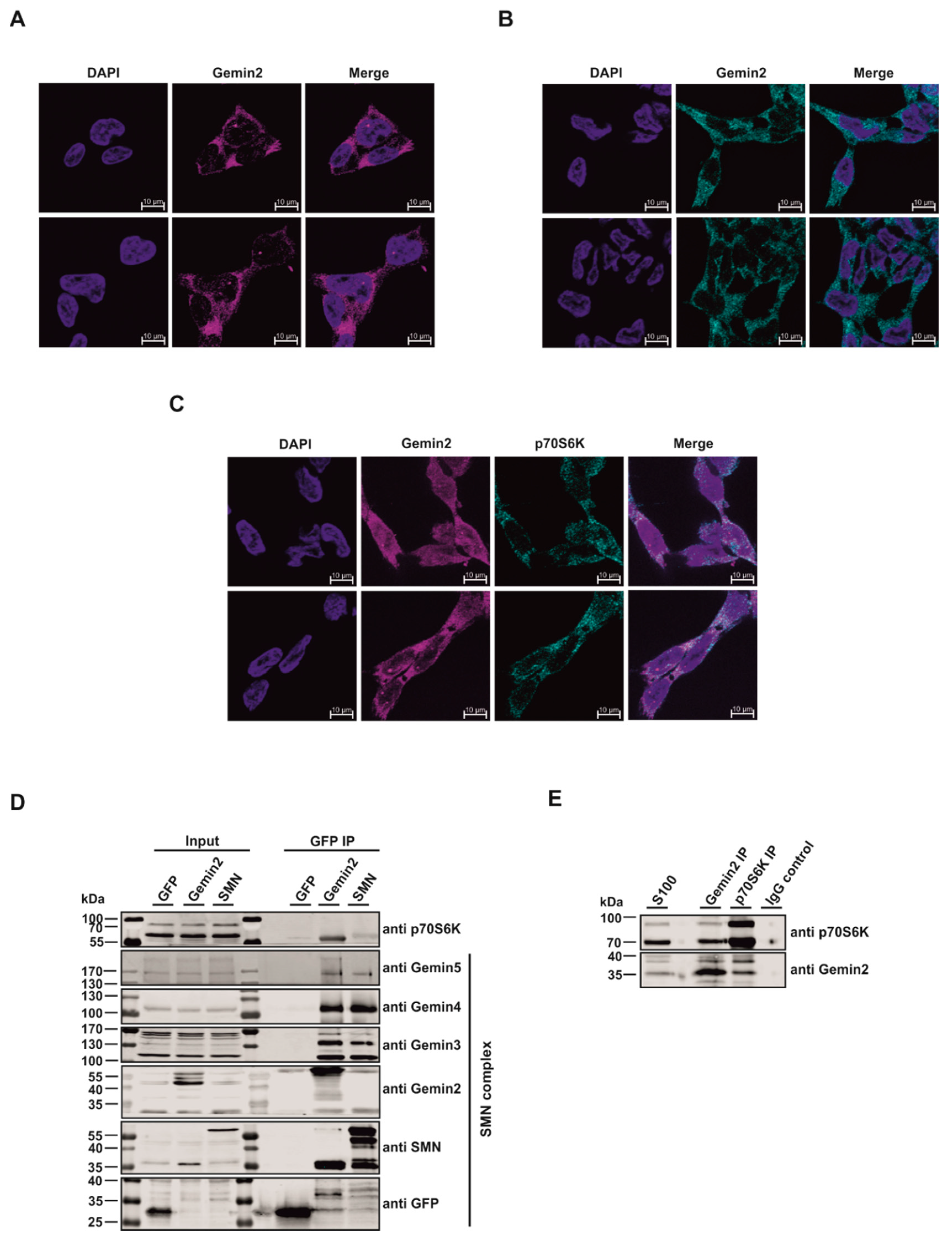{kind=link}
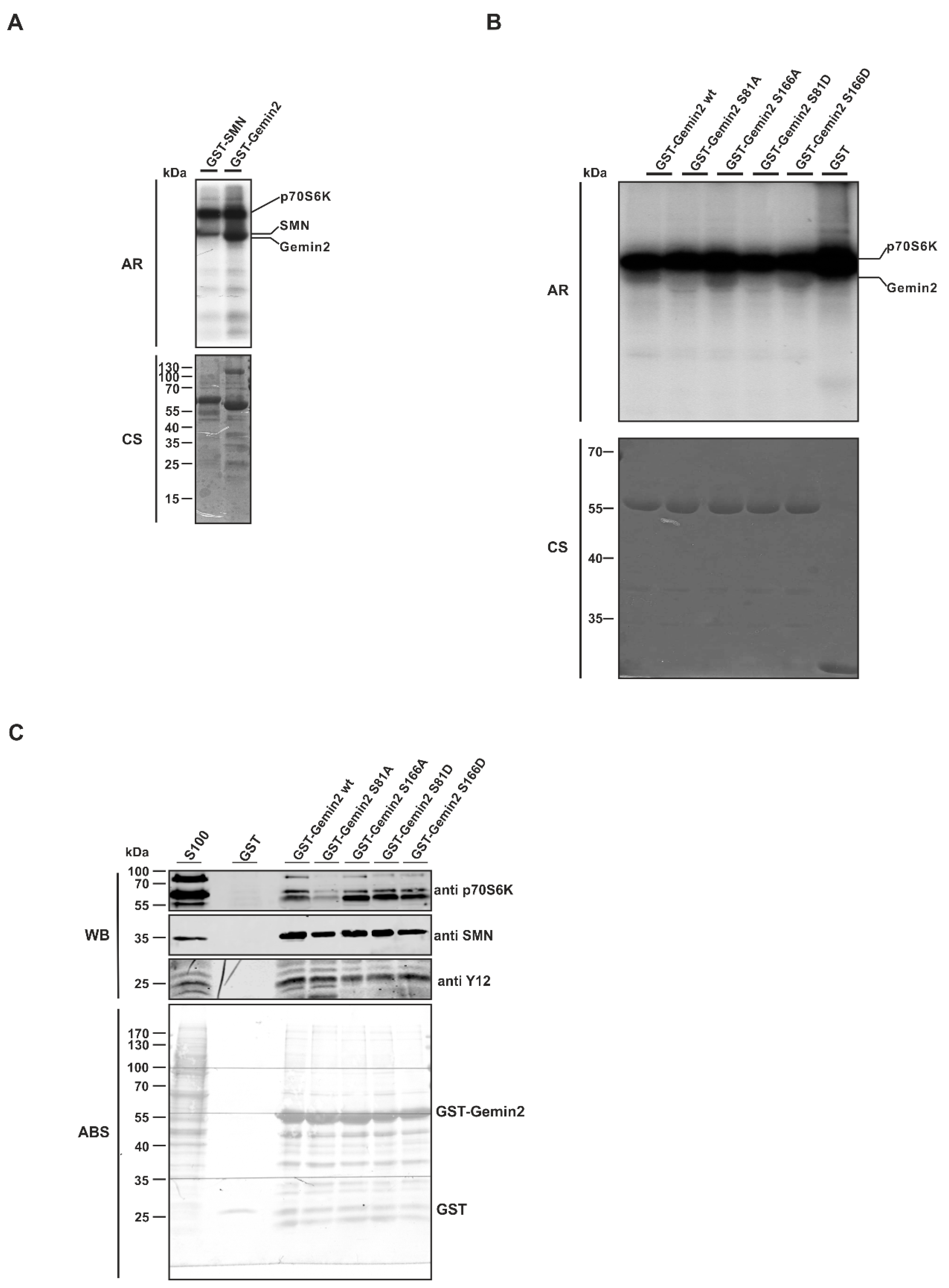{kind=link}
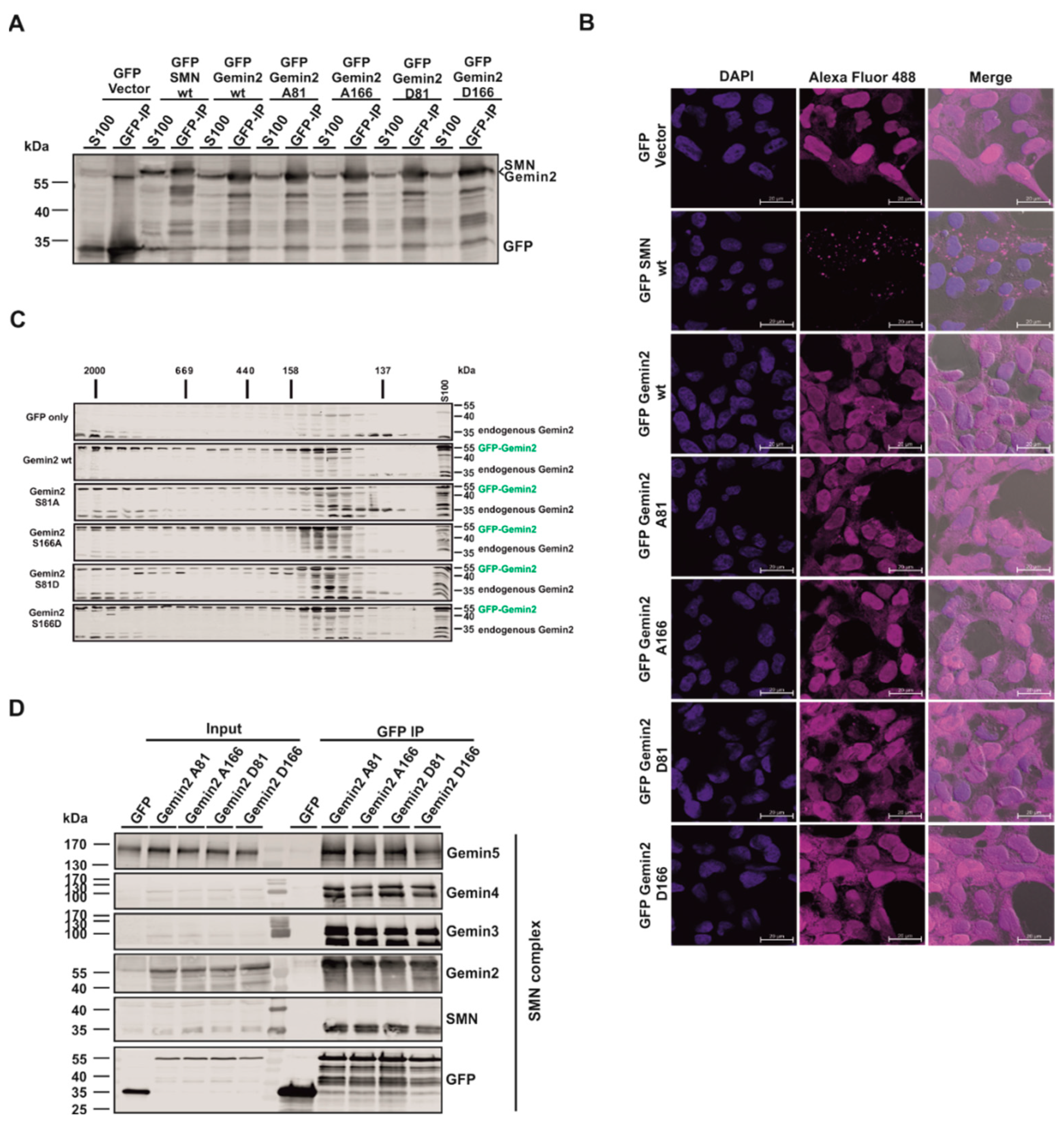{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
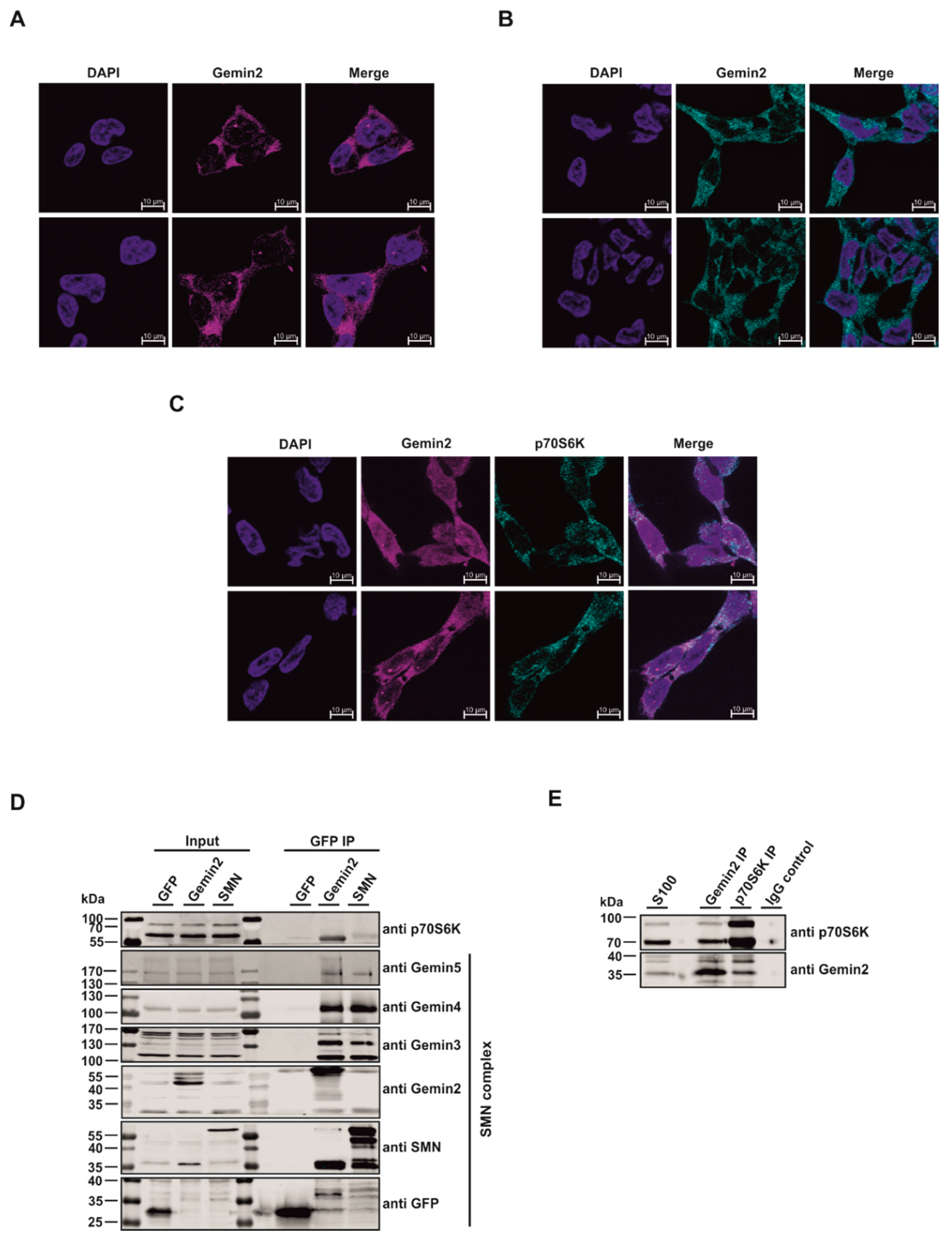
2.1. The p70S6 Kinase Is a New Interaction Partner of the SMN Complex
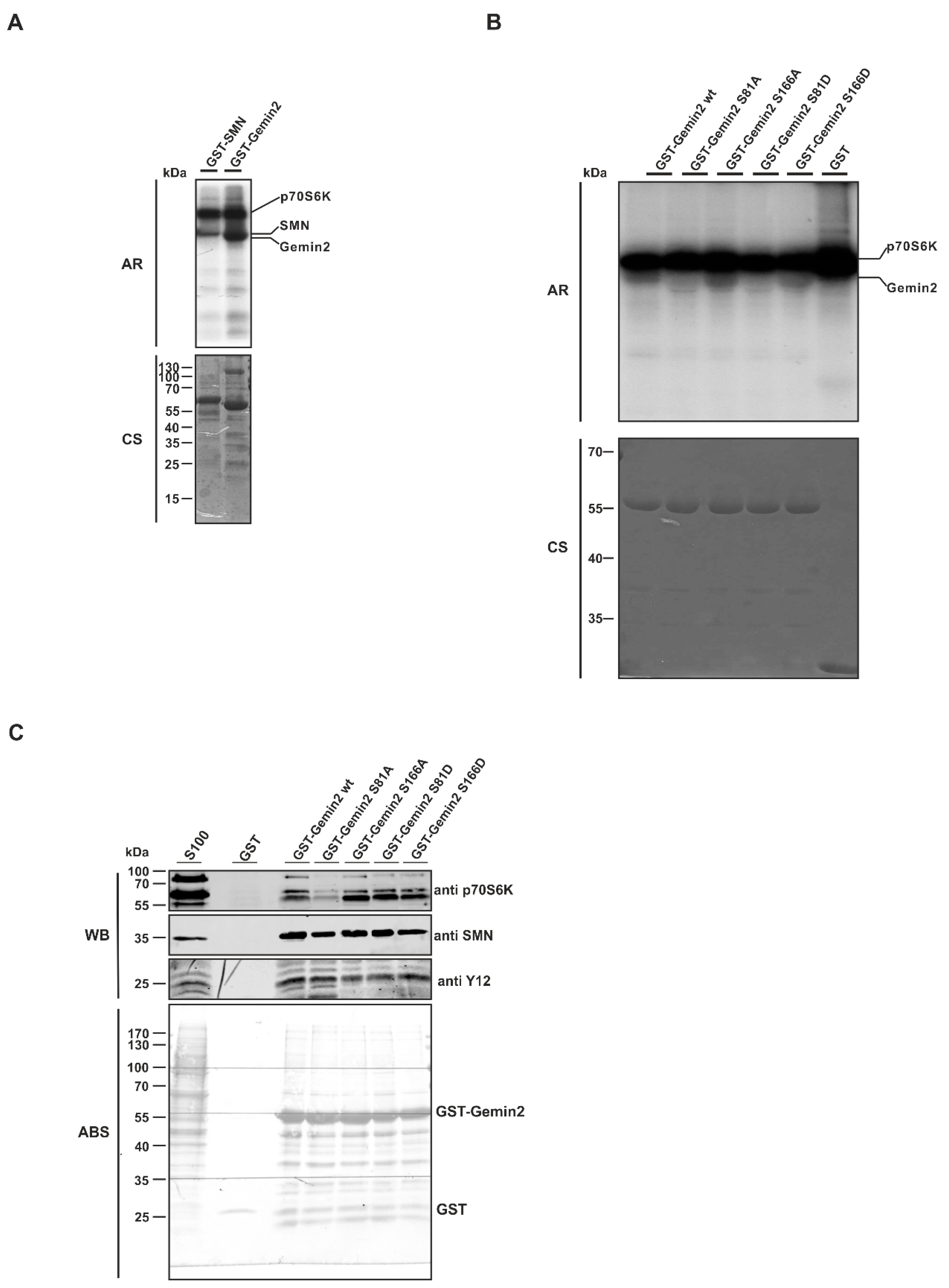
2.2. The SMN Complex Core Subunit Gemin2 Is a New Substrate of the p70S6 Kinase
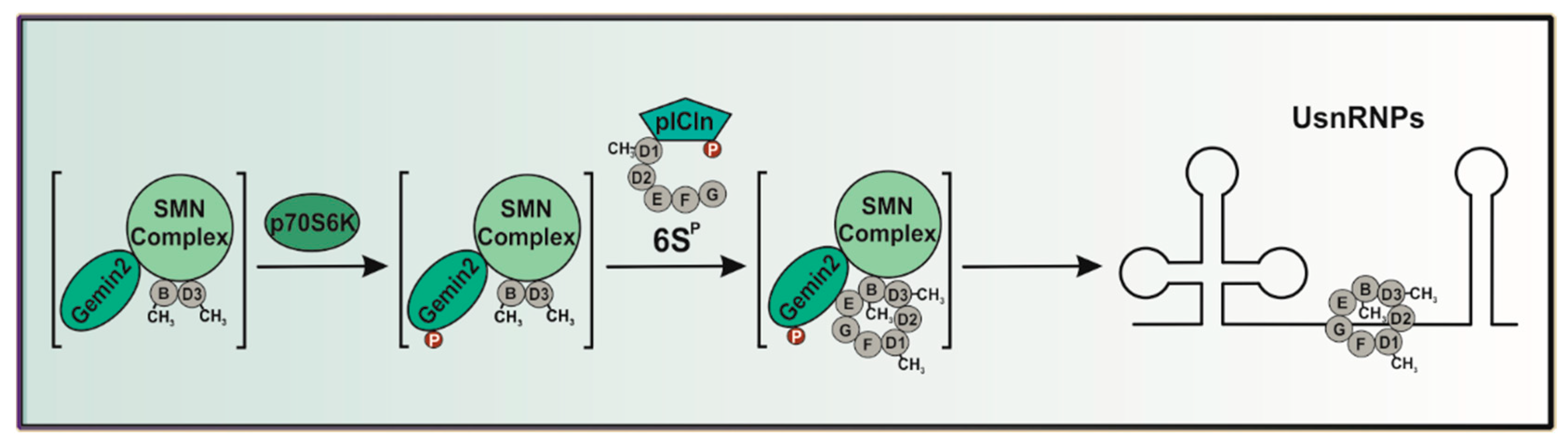
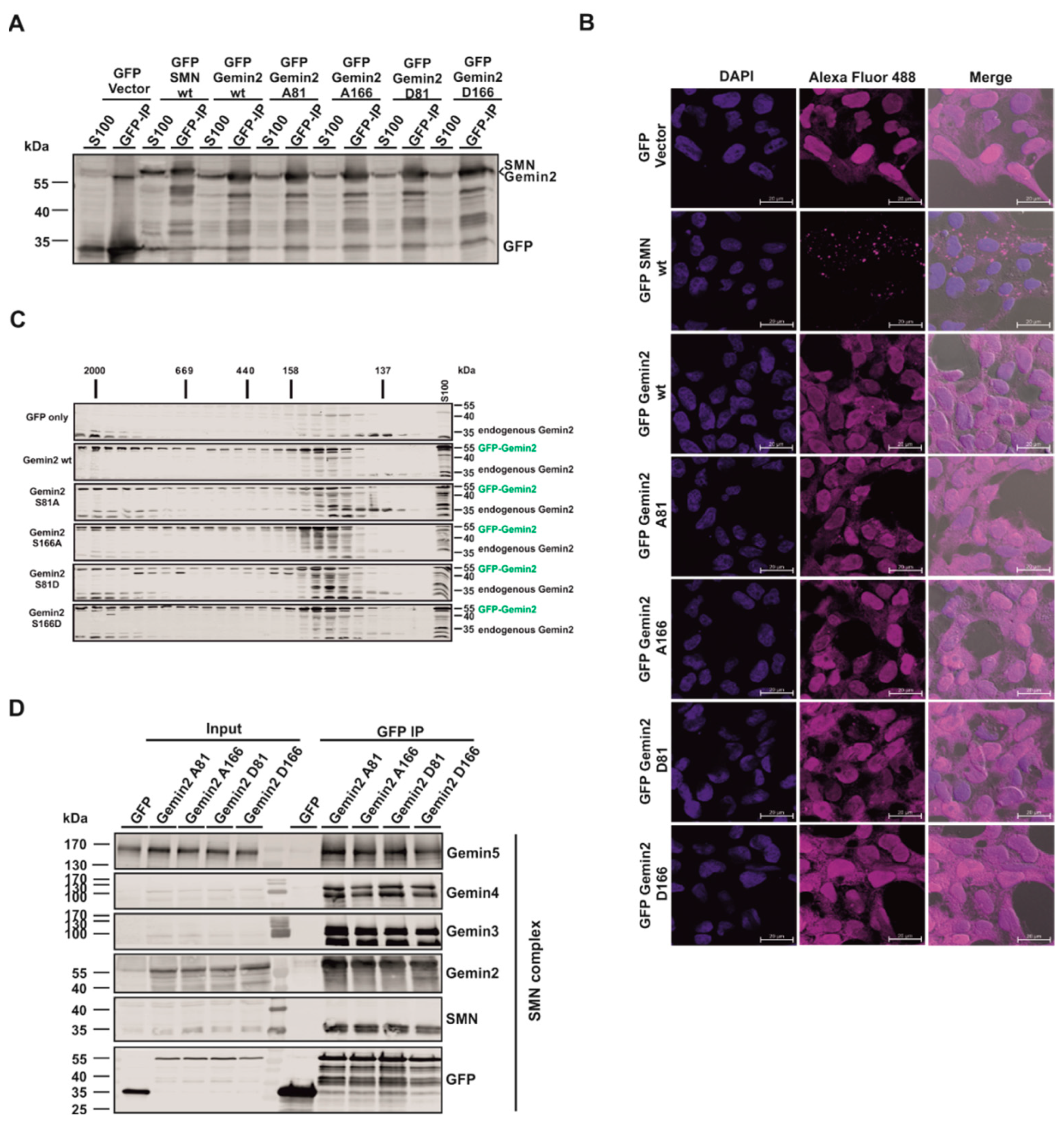
2.3. The p70S6 Kinase-Dependent Phosphorylation Influences Binding towards Gemin2
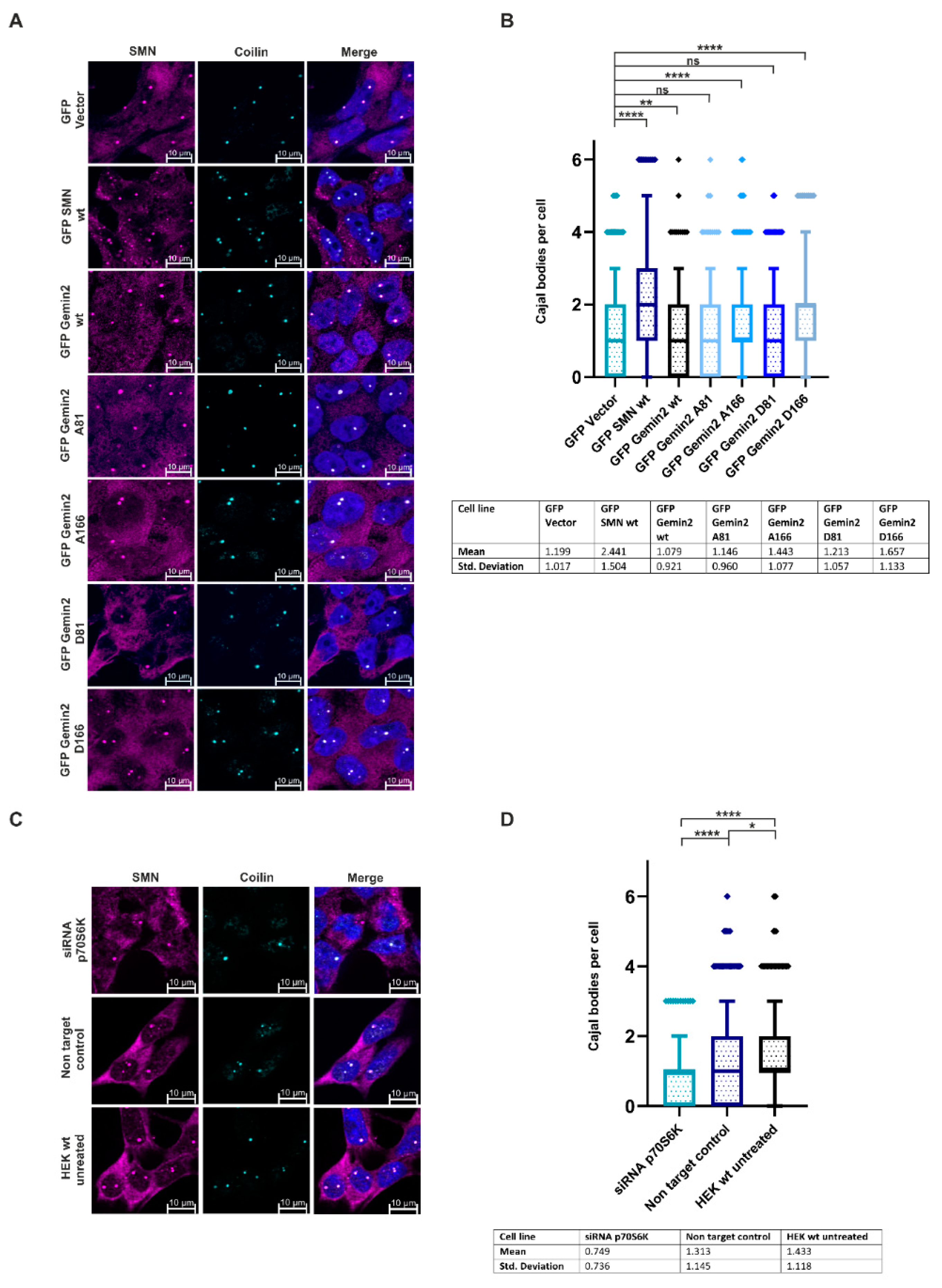
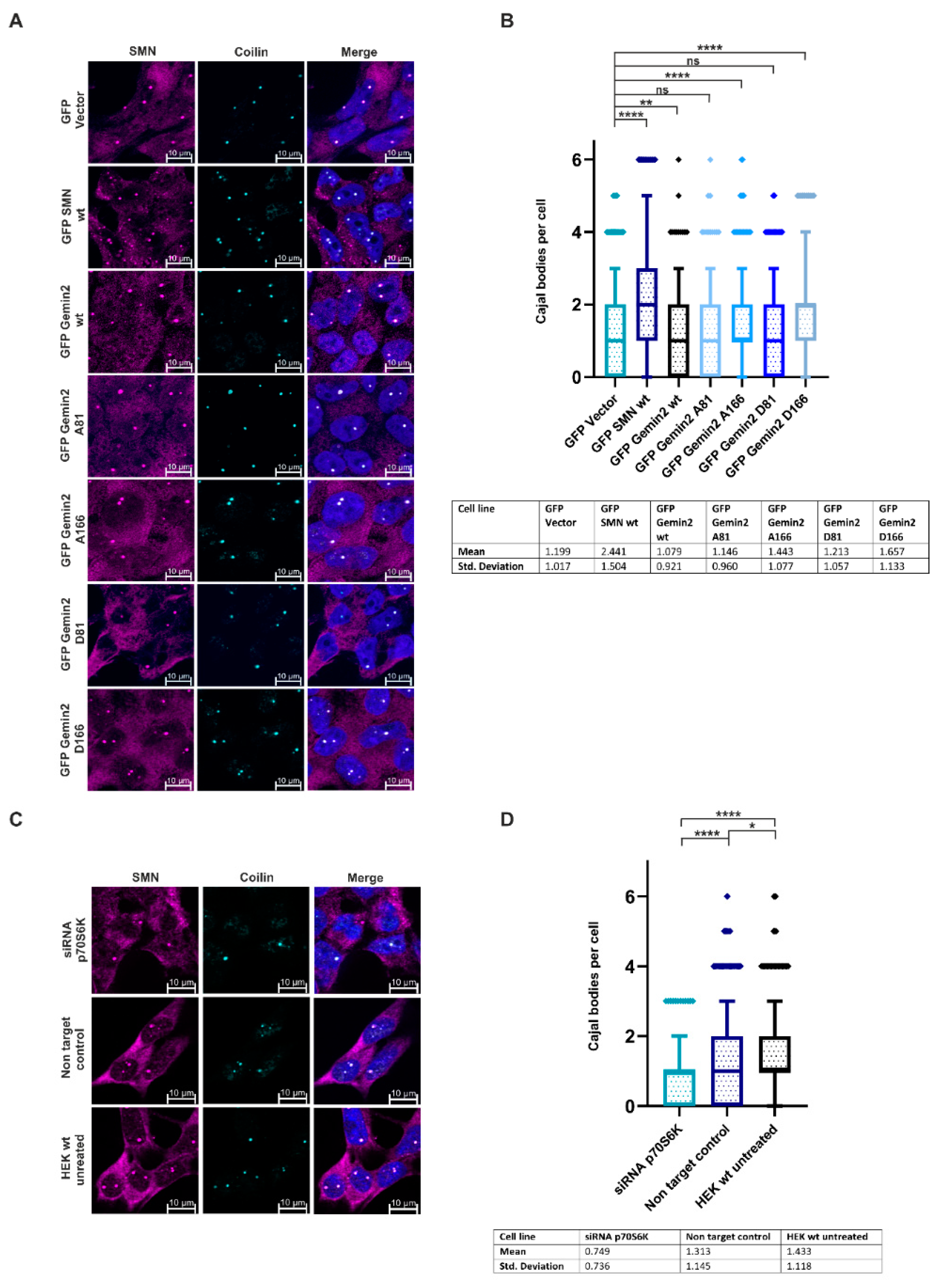
2.4. The Phosphorylation Status of Gemin2 Influences UsnRNP Biogenesis and Inhibition of p70S6K Results in a Decreased Number of Cajal Bodies
3. Discussion
4. Materials and Methods
4.1. Antibodies
4.2. Plasmids and Proteins
4.3. Cell Lines and Cell Culture
4.4. Protein Expression and Purification
4.5. Cytoplasm Extraction (S100) and Size Exclusion Chromatography
4.6. Immunoblotting and Immunopurification
4.7. Immunofluorescence Microscopy
4.8. Immunofluorescence Quantification with Fiji
4.9. In Vitro Phosphorylation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wahl, M.C.; Will, C.L.; Lührmann, R. The spliceosome: Design principles of a dynamic RNP machine. Cell 2009, 136, 701–718. [Google Scholar] [CrossRef] [PubMed]
- Kaida, D.; Berg, M.G.; Younis, I.; Kasim, M.; Singh, L.N.; Wan, L.; Dreyfuss, G. U1 snRNP protects pre-mRNAs from premature cleavage and polyadenylation. Nature 2010, 468, 664–668. [Google Scholar] [CrossRef]
- Staley, J.P.; Guthrie, C. Mechanical devices of the spliceosome: Motors, clocks, springs, and things. Cell 1998, 92, 315–326. [Google Scholar] [CrossRef]
- Mattaj, I.W.; De Robertis, E.M. Nuclear segregation of U2 snRNA requires binding of specific snRNP proteins. Cell 1985, 40, 111–118. [Google Scholar] [CrossRef]
- Kambach, C.; Walke, S.; Young, R.; Avis, J.M.; de la Fortelle, E.; Raker, V.A.; Luhrmann, R.; Li, J.; Nagai, K. Crystal structures of two Sm protein complexes and their implications for the assembly of the spliceosomal snRNPs. Cell 1999, 96, 375–387. [Google Scholar] [CrossRef]
- Will, C.L.; Lührmann, R. Spliceosomal UsnRNP biogenesis, structure and function. Curr. Opin. Cell Biol. 2001, 13, 290–301. [Google Scholar] [CrossRef]
- Weber, G.; Trowitzsch, S.; Kastner, B.; Lührmann, R.; Wahl, M.C. Functional organization of the Sm core in the crystal structure of human U1 snRNP. EMBO J. 2010, 29, 4172–4184. [Google Scholar] [CrossRef] [PubMed]
- Newman, A.J.; Nagai, K. Structural studies of the spliceosome: Blind men and an elephant. Curr. Opin. Struct. Biol. 2010, 20, 82–89. [Google Scholar] [CrossRef]
- Liu, Q.; Fischer, U.; Wang, F.; Dreyfuss, G. The spinal muscular atrophy disease gene product, SMN, and its associated protein SIP1 are in a complex with spliceosomal snRNP proteins. Cell 1997, 90, 1013–1021. [Google Scholar] [CrossRef]
- Friesen, W.J.; Paushkin, S.; Wyce, A.; Massenet, S.; Pesiridis, G.S.; Van Duyne, G.; Rappsilber, J.; Mann, M.; Dreyfuss, G. The methylosome, a 20S complex containing JBP1 and pICln, produces dimethylarginine-modified Sm proteins. Mol. Cell Biol. 2001, 21, 8289–8300. [Google Scholar] [CrossRef]
- Meister, G.; Buhler, D.; Pillai, R.; Lottspeich, F.; Fischer, U. A multiprotein complex mediates the ATP-dependent assembly of spliceosomal U snRNPs. Nat. Cell Biol. 2001, 3, 945–949. [Google Scholar] [CrossRef] [PubMed]
- Charroux, B.; Pellizzoni, L.; Perkinson, R.A.; Shevchenko, A.; Mann, M.; Dreyfuss, G. Gemin3: A novel DEAD box protein that interacts with SMN, the spinal muscular atrophy gene product, and is a component of gems. J. Cell Biol. 1999, 147, 1181–1194. [Google Scholar] [CrossRef]
- Baccon, J.; Pellizzoni, L.; Rappsilber, J.; Mann, M.; Dreyfuss, G. Identification and characterization of Gemin7, a novel component of the survival of motor neuron complex. J. Biol. Chem. 2002, 277, 31957–31962. [Google Scholar] [CrossRef] [PubMed]
- Carissimi, C.; Baccon, J.; Straccia, M.; Chiarella, P.; Maiolica, A.; Sawyer, A.; Rappsilber, J.; Pellizzoni, L. Unrip is a component of SMN complexes active in snRNP assembly. FEBS Lett. 2005, 579, 2348–2354. [Google Scholar] [CrossRef] [PubMed]
- Carissimi, C.; Saieva, L.; Baccon, J.; Chiarella, P.; Maiolica, A.; Sawyer, A.; Rappsilber, J.; Pellizzoni, L. Gemin8 is a novel component of the survival motor neuron complex and functions in small nuclear ribonucleoprotein assembly. J. Biol. Chem. 2006, 281, 8126–8134. [Google Scholar] [CrossRef]
- Grimmler, M.; Otter, S.; Peter, C.; Muller, F.; Chari, A.; Fischer, U. Unrip, a factor implicated in cap-independent translation, associates with the cytosolic SMN complex and influences its intracellular localization. Hum. Mol. Genet. 2005, 14, 3099–3111. [Google Scholar] [CrossRef]
- Kroiss, M.; Schultz, J.; Wiesner, J.; Chari, A.; Sickmann, A.; Fischer, U. Evolution of an RNP assembly system: A minimal SMN complex facilitates formation of UsnRNPs in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2008, 105, 10045–10050. [Google Scholar] [CrossRef]
- Zhang, R.; So, B.R.; Li, P.; Yong, J.; Glisovic, T.; Wan, L.; Dreyfuss, G. Structure of a key intermediate of the SMN complex reveals Gemin2’s crucial function in snRNP assembly. Cell 2011, 146, 384–395. [Google Scholar] [CrossRef]
- Pellizzoni, L.; Yong, J.; Dreyfuss, G. Essential role for the SMN complex in the specificity of snRNP assembly. Science 2002, 298, 1775–1779. [Google Scholar] [CrossRef]
- Fischer, U.; Lührmann, R. An essential signaling role for the m3G cap in the transport of U1 snRNP to the nucleus. Science 1990, 249, 786–790. [Google Scholar] [CrossRef]
- Liu, Q.; Dreyfuss, G. A novel nuclear structure containing the survival of motor neurons protein. EMBO J. 1996, 15, 3555–3565. [Google Scholar] [CrossRef]
- Bellini, M.; Gall, J.G. Coilin shuttles between the nucleus and cytoplasm in Xenopus oocytes. Mol. Biol. Cell 1999, 10, 3425–3434. [Google Scholar] [CrossRef] [PubMed]
- Carmo-Fonseca, M.; Pepperkok, R.; Carvalho, M.T.; Lamond, A.I. Transcription-dependent colocalization of the U1, U2, U4/U6, and U5 snRNPs in coiled bodies. J. Cell Biol. 1992, 117, 1–14. [Google Scholar] [CrossRef]
- Chari, A.; Golas, M.M.; Klingenhager, M.; Neuenkirchen, N.; Sander, B.; Englbrecht, C.; Sickmann, A.; Stark, H.; Fischer, U. An assembly chaperone collaborates with the SMN complex to generate spliceosomal SnRNPs. Cell 2008, 135, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Meister, G.; Eggert, C.; Buhler, D.; Brahms, H.; Kambach, C.; Fischer, U. Methylation of Sm proteins by a complex containing PRMT5 and the putative U snRNP assembly factor pICln. Curr. Biol. 2001, 11, 1990–1994. [Google Scholar] [CrossRef] [PubMed]
- Grimmler, M.; Bauer, L.; Nousiainen, M.; Korner, R.; Meister, G.; Fischer, U. Phosphorylation regulates the activity of the SMN complex during assembly of spliceosomal U snRNPs. EMBO Rep. 2005, 6, 70–76. [Google Scholar] [CrossRef]
- Husedzinovic, A.; Oppermann, F.; Draeger-Meurer, S.; Chari, A.; Fischer, U.; Daub, H.; Gruss, O.J. Phosphoregulation of the human SMN complex. Eur. J. Cell Biol. 2014, 93, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, K.; Cox, J.; Esser, L.M.; Voss, M.; Sander, K.; Loffler, A.; Hillebrand, F.; Erkelenz, S.; Schaal, H.; Kahne, T.; et al. An essential role of the autophagy activating kinase ULK1 in snRNP biogenesis. Nucleic Acids Res. 2021, 49, 6437–6455. [Google Scholar] [CrossRef]
- Schilling, M.; Prusty, A.B.; Boysen, B.; Oppermann, F.S.; Riedel, Y.L.; Husedzinovic, A.; Rasouli, H.; König, A.; Ramanathan, P.; Reymann, J.; et al. TOR signaling regulates liquid phase separation of the SMN complex governing snRNP biogenesis. Cell Rep. 2021, 35, 109277. [Google Scholar] [CrossRef]
- Buhler, D.; Raker, V.; Luhrmann, R.; Fischer, U. Essential role for the tudor domain of SMN in spliceosomal U snRNP assembly: Implications for spinal muscular atrophy. Hum. Mol. Genet. 1999, 8, 2351–2357. [Google Scholar] [CrossRef]
- Pu, W.T.; Krapivinsky, G.B.; Krapivinsky, L.; Clapham, D.E. pICln inhibits snRNP biogenesis by binding core spliceosomal proteins. Mol. Cell Biol. 1999, 19, 4113–4120. [Google Scholar] [CrossRef] [PubMed]
- Pellizzoni, L.; Baccon, J.; Rappsilber, J.; Mann, M.; Dreyfuss, G. Purification of native survival of motor neurons complexes and identification of Gemin6 as a novel component. J. Biol. Chem. 2002, 277, 7540–7545. [Google Scholar] [CrossRef] [PubMed]
- Gubitz, A.K.; Mourelatos, Z.; Abel, L.; Rappsilber, J.; Mann, M.; Dreyfuss, G. Gemin5, a novel WD repeat protein component of the SMN complex that binds Sm proteins. J. Biol. Chem. 2002, 277, 5631–5636. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, C.; Usui, K.; Aoki, M.; Ito, F.; Itoh, M.; Kai, C.; Kanamori-Katayama, M.; Hayashizaki, Y.; Suzuki, H. Gemin2 plays an important role in stabilizing the survival of motor neuron complex. J. Biol. Chem. 2007, 282, 11122–11134. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, T.; Almeida, F.; Calapez, A.; Lafarga, M.; Berciano, M.T.; Carmo-Fonseca, M. The spinal muscular atrophy disease gene product, SMN: A link between snRNP biogenesis and the Cajal (coiled) body. J. Cell Biol. 1999, 147, 715–728. [Google Scholar] [CrossRef]
- Esser, L.M.; Schmitz, K.; Hillebrand, F.; Erkelenz, S.; Schaal, H.; Stork, B.; Grimmler, M.; Wesselborg, S.; Peter, C. Phosphorylation of pICln by the autophagy activating kinase ULK1 regulates snRNP biogenesis and splice activity of the cell. Comput. Struct. Biotechnol. J. 2023, 21, 2100–2109. [Google Scholar] [CrossRef]
- Meister, G.; Fischer, U. Assisted RNP assembly: SMN and PRMT5 complexes cooperate in the formation of spliceosomal UsnRNPs. EMBO J. 2002, 21, 5853–5863. [Google Scholar] [CrossRef]
- Lefebvre, S.; Burlet, P.; Liu, Q.; Bertrandy, S.; Clermont, O.; Munnich, A.; Dreyfuss, G.; Melki, J. Correlation between severity and SMN protein level in spinal muscular atrophy. Nat. Genet. 1997, 16, 265–269. [Google Scholar] [CrossRef]
- Coovert, D.D.; Le, T.T.; McAndrew, P.E.; Strasswimmer, J.; Crawford, T.O.; Mendell, J.R.; Coulson, S.E.; Androphy, E.J.; Prior, T.W.; Burghes, A.H. The survival motor neuron protein in spinal muscular atrophy. Hum. Mol. Genet. 1997, 6, 1205–1214. [Google Scholar] [CrossRef]
- Loffler, A.S.; Alers, S.; Dieterle, A.M.; Keppeler, H.; Franz-Wachtel, M.; Kundu, M.; Campbell, D.G.; Wesselborg, S.; Alessi, D.R.; Stork, B. Ulk1-mediated phosphorylation of AMPK constitutes a negative regulatory feedback loop. Autophagy 2011, 7, 696–706. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esser, L.M.; Li, Q.; Jüdt, M.; Kähne, T.; Stork, B.; Grimmler, M.; Wesselborg, S.; Peter, C. The Impact of p70S6 Kinase-Dependent Phosphorylation of Gemin2 in UsnRNP Biogenesis. Int. J. Mol. Sci. 2023, 24, 15552. https://doi.org/10.3390/ijms242115552
Esser LM, Li Q, Jüdt M, Kähne T, Stork B, Grimmler M, Wesselborg S, Peter C. The Impact of p70S6 Kinase-Dependent Phosphorylation of Gemin2 in UsnRNP Biogenesis. International Journal of Molecular Sciences. 2023; 24(21):15552. https://doi.org/10.3390/ijms242115552
Chicago/Turabian StyleEsser, Lea Marie, Qiaoping Li, Maximilian Jüdt, Thilo Kähne, Björn Stork, Matthias Grimmler, Sebastian Wesselborg, and Christoph Peter. 2023. "The Impact of p70S6 Kinase-Dependent Phosphorylation of Gemin2 in UsnRNP Biogenesis" International Journal of Molecular Sciences 24, no. 21: 15552. https://doi.org/10.3390/ijms242115552
APA StyleEsser, L. M., Li, Q., Jüdt, M., Kähne, T., Stork, B., Grimmler, M., Wesselborg, S., & Peter, C. (2023). The Impact of p70S6 Kinase-Dependent Phosphorylation of Gemin2 in UsnRNP Biogenesis. International Journal of Molecular Sciences, 24(21), 15552. https://doi.org/10.3390/ijms242115552