
The Root-Colonizing Endophyte Piriformospora indica Supports Nitrogen-Starved Arabidopsis thaliana Seedlings with Nitrogen Metabolites

 , , ,
, , ,  , ,
, ,  ,
,  , and
, and
Abstract
1. Introduction
2. Results
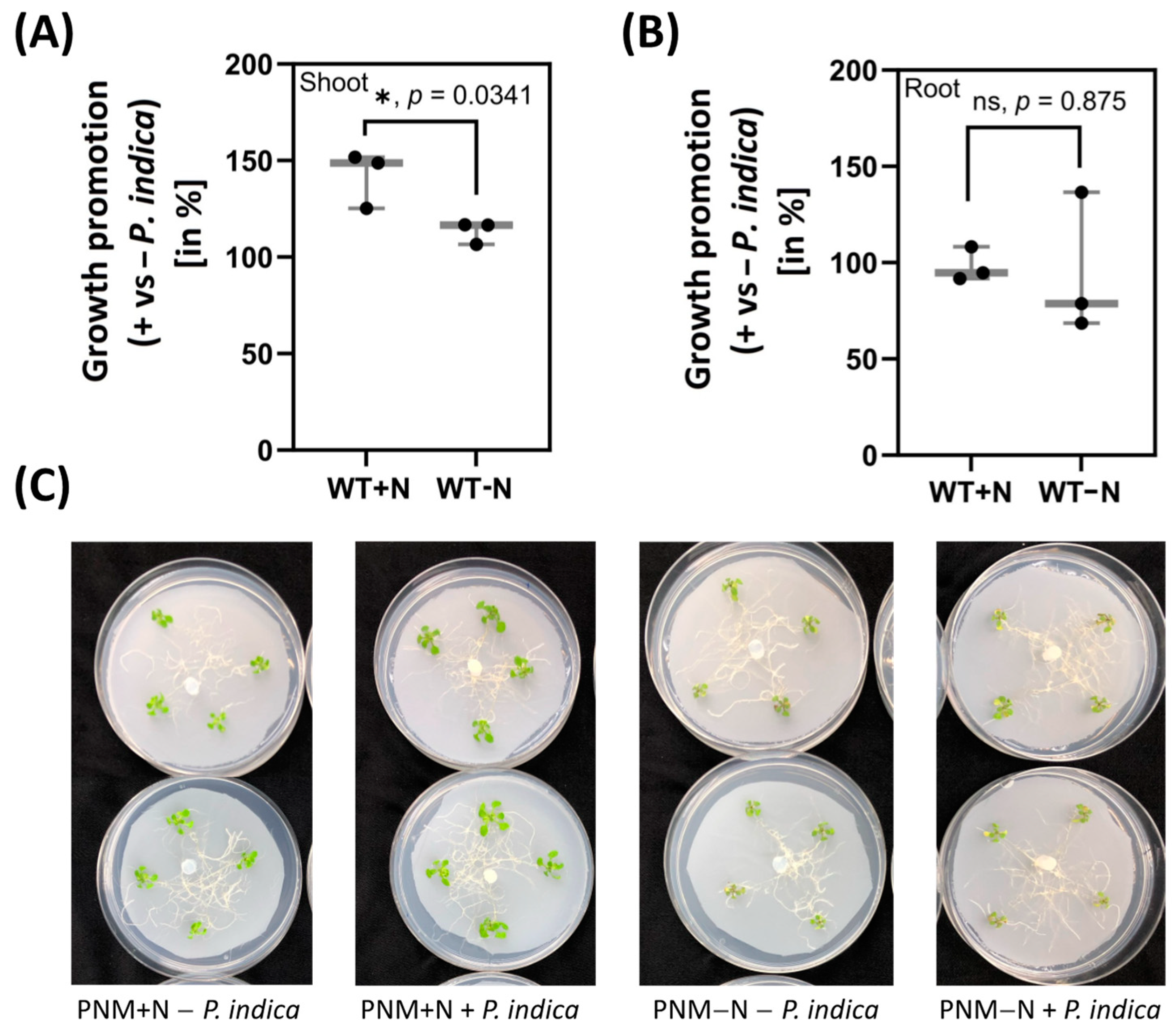
2.1. Shoot Growth Promotion by P. indica Requires External N Supply
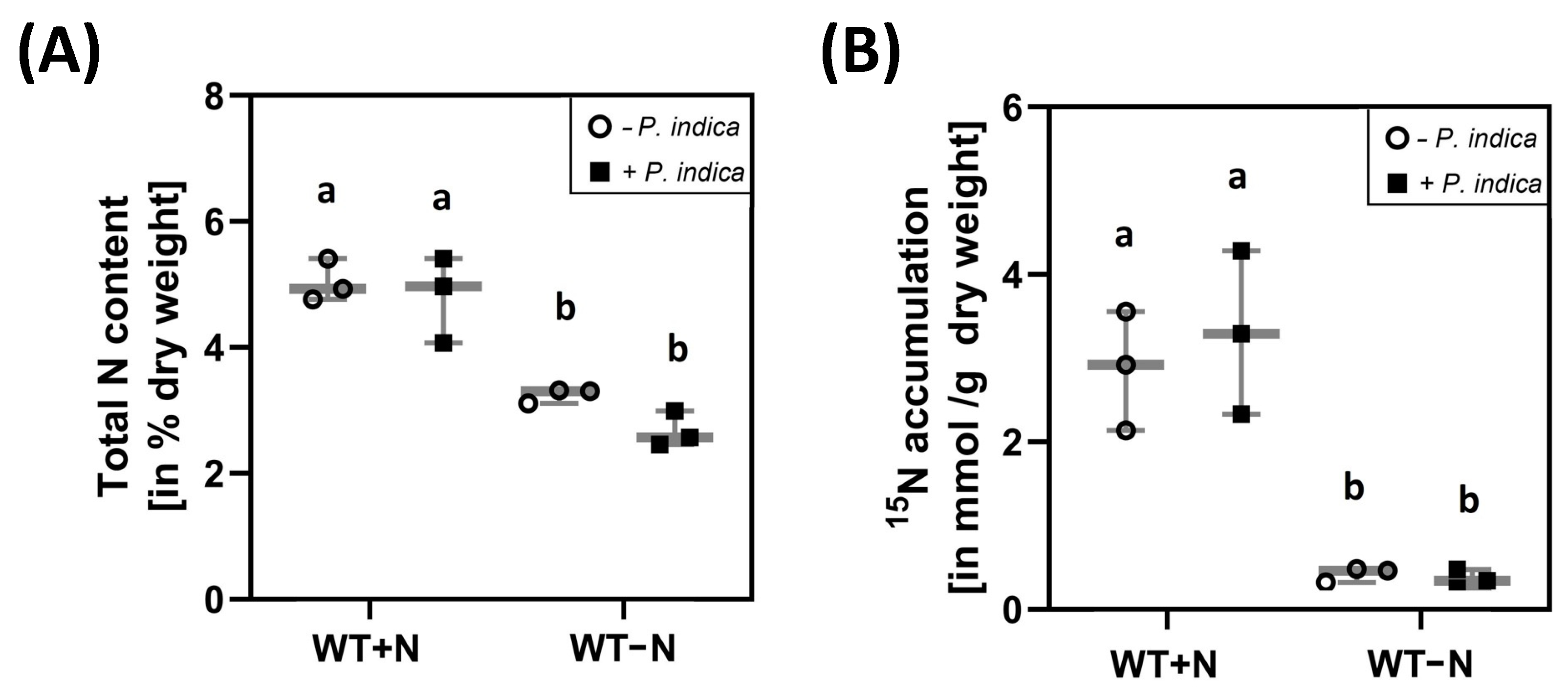
2.2. P. indica Colonisation Did Not Change the Total N Content in the Shoots and Transfer of 15N from the Medium to the Shoots
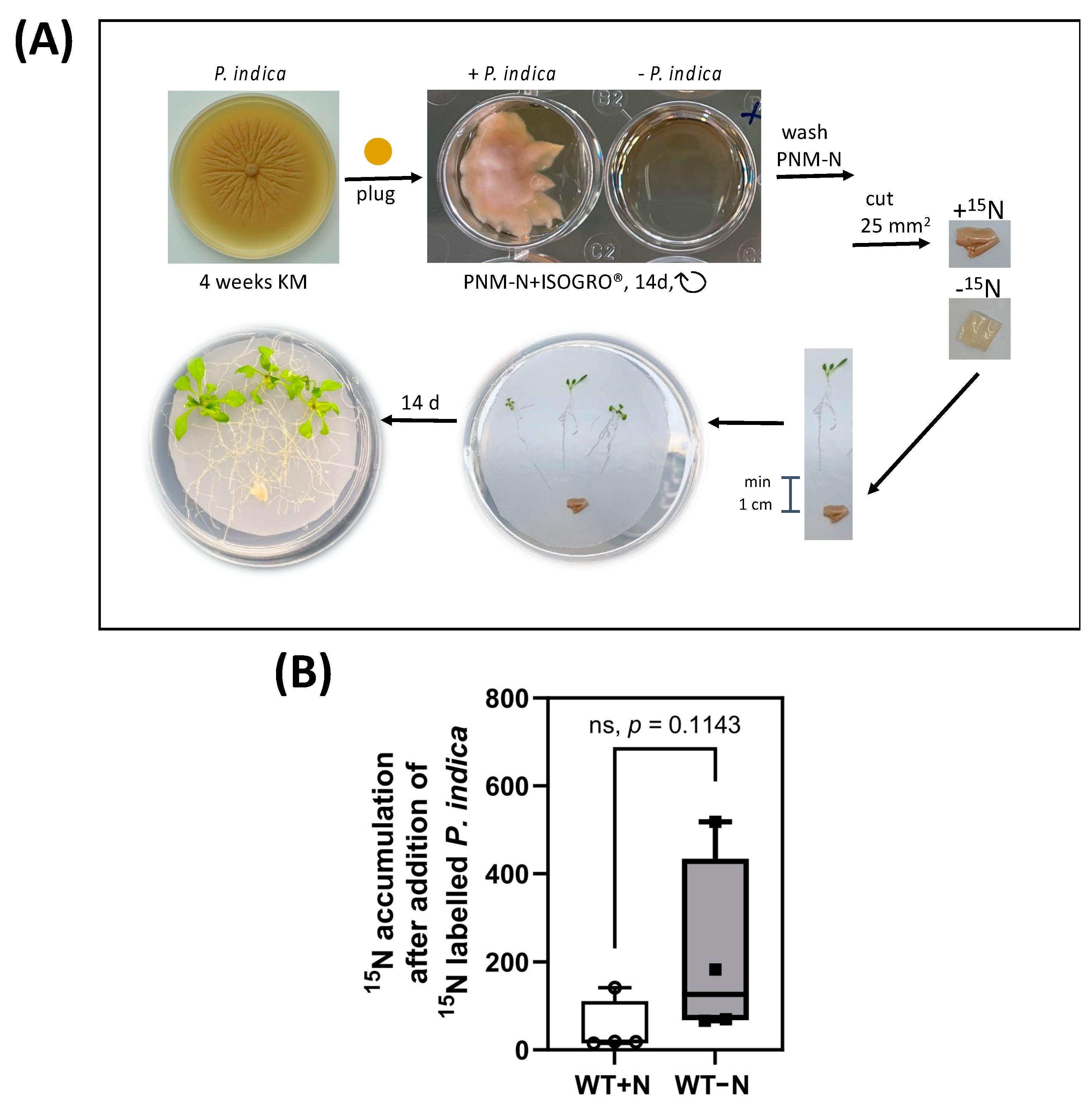
2.3. 15N Label Is Transferred from P. idica to the Host under N-Limiting Conditions
2.4. Reprogramming of the Metabolite Profiles to N Limitation Conditions Is Alleviated by P. indica
2.5. P. indica Stimulates Expression of Specific Host´s Transporter Genes under N Limitation
3. Discussion
4. Materials and Methods
4.1. Plant and Fungus Material and Corresponding Growth Conditions
4.2. Plant-Fungus Co-Cultures and the Determination of Growth Promotion
4.3. 15N Labelling Experiments in the Medium
4.4. 15N Fungus-Labelling Experiments
4.5. Isolation and Clean-Up of RNA
4.6. RNAseq and Data Analysis
4.7. Analysis of Fungal Colonization via qPCR
4.8. Determination of Total Nitrogen and 15N Enrichment
4.9. Metabolomic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Engels, C.; Marschner, H. Plant uptake and utilization of nitrogen. In Nitrogen Fertilization in the Environment; Bacan, P.E., Ed.; Marcel Dekker, Inc.: New York, NY, USA, 1995; pp. 41–81. [Google Scholar]
- Vidal, E.A.; Alvarez, J.M.; Araus, V.; Riveras, E.; Brooks, M.D.; Krouk, G.; Ruffel, S.; Lejay, L.; Crawford, N.M.; Coruzzi, G.M.; et al. Nitrate in 2020: Thirty Years from Transport to Signaling Networks. Plant Cell 2020, 32, 2094–2119. [Google Scholar] [CrossRef] [PubMed]
- Lyzenga, W.J.; Liu, Z.; Olukayode, T.; Zhao, Y.; Kochian, L.V.; Ham, B.-K. Getting to the roots of N, P, and K uptake. J. Exp. Bot. 2023, 74, 1784–1805. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-Y.; Cheng, Y.-H.; Chen, K.-E.; Tsay, Y.-F. Nitrate Transport, Signaling, and Use Efficiency. Annu. Rev. Plant Biol. 2018, 69, 85–122. [Google Scholar] [CrossRef] [PubMed]
- Aluko, O.O.; Kant, S.; Adedire, O.M.; Li, C.; Yuan, G.; Liu, H.; Wang, Q. Unlocking the potentials of nitrate transporters at improving plant nitrogen use efficiency. Front. Plant Sci. 2023, 14, 1074839. [Google Scholar] [CrossRef] [PubMed]
- Guo, K.; Yang, J.; Yu, N.; Luo, L.; Wang, E. Biological nitrogen fixation in cereal crops: Progress, strategies, and perspectives. Plant Commun. 2023, 4, 100499. [Google Scholar] [CrossRef]
- Rui, W.; Mao, Z.; Li, Z. The Roles of Phosphorus and Nitrogen Nutrient Transporters in the Arbuscular Mycorrhizal Symbiosis. Int. J. Mol. Sci. 2022, 23, 11027. [Google Scholar] [CrossRef]
- Chaudhary, P.; Agri, U.; Chaudhary, A.; Kumar, A.; Kumar, G. Endophytes and their potential in biotic stress management and crop production. Front. Microbiol. 2022, 13, 933017. [Google Scholar] [CrossRef]
- Zhang, Y.; Feng, H.; Druzhinina, I.S.; Xie, X.; Wang, E.; Martin, F.; Yuan, Z. Phosphorus/nitrogen sensing and signaling in diverse root-fungus symbioses. Trends Microbiol. 2023. [Google Scholar] [CrossRef]
- Saleem, S.; Sekara, A.; Pokluda, R. Serendipita indica—A Review from Agricultural Point of View. Plants 2022, 11, 3417. [Google Scholar] [CrossRef]
- Del Barrio-Duque, A.; Ley, J.; Samad, A.; Antonielli, L.; Sessitsch, A.; Compant, S. Beneficial Endophytic Bacteria-Serendipita indica Interaction for Crop Enhancement and Resistance to Phytopathogens. Front. Microbiol. 2019, 10, 2888. [Google Scholar] [CrossRef]
- Sherameti, I.; Shahollari, B.; Venus, Y.; Altschmied, L.; Varma, A.; Oelmüller, R. The Endophytic Fungus Piriformospora indica Stimulates the Expression of Nitrate Reductase and the Starch-degrading Enzyme Glucan-water Dikinase in Tobacco and Arabidopsis Roots through a Homeodomain Transcription Factor That Binds to a Conserved Motif in Their Promoters. J. Biol. Chem. 2005, 280, 26241–26247. [Google Scholar] [CrossRef]
- Eliaspour, S.; Sharifi, R.S.; Shirkhani, A. Evaluation of interaction between Piriformospora indica, animal manure and NPK fertilizer on quantitative and qualitative yield and absorption of elements in sunflower. Food Sci. Nutr. 2020, 8, 2789–2797. [Google Scholar] [CrossRef] [PubMed]
- Strehmel, N.; Mönchgesang, S.; Herklotz, S.; Krüger, S.; Ziegler, J.; Scheel, D. Piriformospora indica Stimulates Root Metabolism of Arabidopsis thaliana. Int. J. Mol. Sci. 2016, 17, 1091. [Google Scholar] [CrossRef]
- Ghaffari, M.R.; Ghabooli, M.; Khatabi, B.; Hajirezaei, M.R.; Schweizer, P.; Salekdeh, G.H. Metabolic and transcriptional response of central metabolism affected by root endophytic fungus Piriformospora indica under salinity in barley. Plant Mol. Biol. 2016, 90, 699–717. [Google Scholar] [CrossRef] [PubMed]
- Lahrmann, U.; Ding, Y.; Banhara, A.; Rath, M.; Hajirezaei, M.R.; Döhlemann, S.; von Wirén, N.; Parniske, M.; Zuccaro, A. Host-related metabolic cues affect colonization strategies of a root endophyte. Proc. Natl. Acad. Sci. USA 2013, 110, 13965–13970. [Google Scholar] [CrossRef] [PubMed]
- Hua, M.D.-S.; Kumar, R.S.; Shyur, L.-F.; Cheng, Y.-B.; Tian, Z.; Oelmüller, R.; Yeh, K.-W. Metabolomic compounds identified in Piriformospora indica-colonized Chinese cabbage roots delineate symbiotic functions of the interaction. Sci. Rep. 2017, 7, 9291. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, P.; Yadav, B.G.; Kumar, S.G.; Kumar, R.; Kogel, K.-H.; Kumar, S. Piriformospora indica and Azotobacter chroococcum Consortium Facilitates Higher Acquisition of N, P with Improved Carbon Allocation and Enhanced Plant Growth in Oryza sativa. J. Fungi 2022, 8, 453. [Google Scholar] [CrossRef]
- Mansotra, P.; Sharma, P.; Sharma, S. Bioaugmentation of Mesorhizobium cicer, Pseudomonas spp. and Piriformospora indica for Sustainable Chickpea Production. Physiol. Mol. Biol. Plants 2015, 21, 385–393. [Google Scholar] [CrossRef]
- Hallasgo, A.M.; Spangl, B.; Steinkellner, S.; Hage-Ahmed, K. The Fungal Endophyte Serendipita williamsii Does Not Affect Phosphorus Status But Carbon and Nitrogen Dynamics in Arbuscular Mycorrhizal Tomato Plants. J. Fungi 2020, 6, 233. [Google Scholar] [CrossRef]
- Abdelaziz, M.E.; Kim, D.; Ali, S.; Fedoroff, N.V.; Al-Babili, S. The endophytic fungus Piriformospora indica enhances Arabidopsis thaliana growth and modulates Na+/K+ homeostasis under salt stress conditions. Plant Sci. 2017, 263, 107–115. [Google Scholar] [CrossRef]
- Krapp, A.; Berthomé, R.; Orsel, M.; Mercey-Boutet, S.; Yu, A.; Castaings, L.; Elftieh, S.; Major, H.; Renou, J.-P.; Daniel-Vedele, F. Arabidopsis Roots and Shoots Show Distinct Temporal Adaptation Patterns toward Nitrogen Starvation. Plant Physiol. 2011, 157, 1255–1282. [Google Scholar] [CrossRef]
- Hodge, A.; Stewart, J.; Robinson, D.; Griffiths, B.S.; Fitter, A.H. Competition between roots and soil microorganisms for nutrientsfrom nitrogen-rich patches of varying complexity. J. Ecol. 2000, 88, 150–164. [Google Scholar] [CrossRef]
- Hodge, A.; Campbell, C.D.; Fitter, A.H. An arbuscular mycorrhizal fungus accelerates decomposition and acquires nitrogen directly from organic material. Nature 2001, 413, 297–299. [Google Scholar] [CrossRef]
- Thirkell, T.J.; Cameron, D.D.; Hodge, A. Resolving the ‘nitrogen paradox’ of arbuscular mycorrhizas: Fertilisation with organic matter brings considerable benefits for plant nutrition and growth. Plant Cell Environ. 2016, 39, 1683–1690. [Google Scholar] [CrossRef] [PubMed]
- Hoysted, G.A.; Field, K.J.; Sinanaj, B.; Bell, C.A.; Bidartondo, M.I.; Pressel, S. Direct nitrogen, phosphorus and carbon exchanges between Mucoromycotina ‘fine root endophyte’ fungi and a flowering plant in novel monoxenic cultures. New Phytol. 2023, 238, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Breuillin-Sessoms, F.; Floss, D.S.; Gomez, S.K.; Pumplin, N.; Ding, Y.; Levesque-Tremblay, V.; Noar, R.D.; Daniels, D.A.; Bravo, A.; Eaglesham, J.B.; et al. Suppression of Arbuscule Degeneration in Medicago truncatula phosphate transporter4 Mutants Is Dependent on the Ammonium Transporter 2 Family Protein AMT2;3. Plant Cell 2015, 27, 1352–1366. [Google Scholar] [CrossRef]
- Paul, E.A.; Kucey, R.M.N. Carbon flow in plant microbial associations incriminated in perinatal morbidity and mortality. Science 1981, 213, 473–474. [Google Scholar] [CrossRef]
- Wright, D.P.; Read, D.J.; Scholes, J.D. Mycorrhizal sink strength influences whole plant carbon balance of Trifolium repens L. Plant Cell Environ. 1998, 21, 881–891. [Google Scholar] [CrossRef]
- Lendenmann, M.; Thonar, C.; Barnard, R.L.; Salmon, Y.; Werner, R.A.; Frossard, E.; Jansa, J. Symbiont identity matters: Carbon and phosphorus fluxes between Medicago truncatula and different arbuscular mycorrhizal fungi. Mycorrhiza 2011, 21, 689–702. [Google Scholar] [CrossRef]
- Cope, K.R.; Kafle, A.; Yakha, J.K.; Pfeffer, P.E.; Strahan, G.D.; Garcia, K.; Subramanian, S.; Bücking, H. Physiological and transcriptomic response of Medicago truncatula to colonization by high- or low-benefit arbuscular mycorrhizal fungi. Mycorrhiza 2022, 32, 281–303. [Google Scholar] [CrossRef]
- Belmondo, S.; Fiorilli, V.; Pérez-Tienda, J.; Ferrol, N.; Marmeisse, R.; Lanfranco, L. A dipeptide transporter from the arbuscular mycorrhizal fungus Rhizophagus irregularis is upregulated in the intraradical phase. Front. Plant Sci. 2014, 5, 436. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, T.; Zhang, C.; Zhang, W.; Deng, N.; Dirk, L.M.A.; Downie, A.B.; Zhao, T. Raffinose positively regulates maize drought tolerance by reducing leaf transpiration. Plant J. 2023, 114, 55–67. [Google Scholar] [CrossRef] [PubMed]
- ElSayed, A.I.; Rafudeen, M.S.; Golldack, D. Physiological aspects of raffinose family oligosaccharides in plants: Protection against abiotic stress. Plant Biol. 2014, 16, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Jiang, T.; Zhang, C.; Li, X.; Wang, C.; Zhang, Y.; Li, T.; Dirk, L.M.A.; Downie, A.B.; Zhao, T. Maize HSFA2 and HSBP2 antagonistically modulate raffinose biosynthesis and heat tolerance in Arabidopsis. Plant J. 2019, 100, 128–142. [Google Scholar] [CrossRef]
- Han, Q.; Qi, J.; Hao, G.; Zhang, C.; Wang, C.; Dirk, L.M.A.; Downie, A.B.; Zhao, T. ZmDREB1A Regulates RAFFINOSE SYNTHASE Controlling Raffinose Accumulation and Plant Chilling Stress Tolerance in Maize. Plant Cell Physiol. 2020, 61, 331–341. [Google Scholar] [CrossRef]
- Li, C.; Tien, H.; Wen, M.; Yen, H.E. Myo-inositol transport and metabolism participate in salt tolerance of halophyte ice plant seedlings. Physiol. Plant. 2021, 172, 1619–1629. [Google Scholar] [CrossRef]
- Yang, J.; Ling, C.; Liu, Y.; Zhang, H.; Hussain, Q.; Lyu, S.; Wang, S.; Liu, Y. Genome-Wide Expression Profiling Analysis of Kiwifruit GolS and RFS Genes and Identification of AcRFS4 Function in Raffinose Accumulation. Int. J. Mol. Sci. 2022, 23, 8836. [Google Scholar] [CrossRef]
- Wang, S.; Chen, A.; Xie, K.; Yang, X.; Luo, Z.; Chen, J.; Zeng, D.; Ren, Y.; Yang, C.; Wang, L.; et al. Functional analysis of the OsNPF4.5 nitrate transporter reveals a conserved mycorrhizal pathway of nitrogen acquisition in plants. Proc. Natl. Acad. Sci. USA 2020, 117, 16649–16659. [Google Scholar] [CrossRef]
- Kiba, T.; Feria-Bourrellier, A.-B.; Lafouge, F.; Lezhneva, L.; Boutet-Mercey, S.; Orsel, M.; Bréhaut, V.; Miller, A.; Daniel-Vedele, F.; Sakakibara, H.; et al. The Arabidopsis Nitrate Transporter NRT2.4 Plays a Double Role in Roots and Shoots of Nitrogen-Starved Plants. Plant Cell 2012, 24, 245–258. [Google Scholar] [CrossRef]
- Chen, H.-Y.; Chen, Y.-N.; Wang, H.-Y.; Liu, Z.-T.; Frommer, W.B.; Ho, C.-H. Feedback inhibition of AMT1 NH4+-transporters mediated by CIPK15 kinase. BMC Biol. 2020, 18, 196. [Google Scholar] [CrossRef]
- Li, J.-Y.; Fu, Y.-L.; Pike, S.M.; Bao, J.; Tian, W.; Zhang, Y.; Chen, C.-Z.; Zhang, Y.; Li, H.-M.; Huang, J.; et al. The Arabidopsis Nitrate Transporter NRT1.8 Functions in Nitrate Removal from the Xylem Sap and Mediates Cadmium Tolerance. Plant Cell 2010, 22, 1633–1646. [Google Scholar] [CrossRef] [PubMed]
- Dechorgnat, J.; Patrit, O.; Krapp, A.; Fagard, M.; Daniel-Vedele, F. Characterization of the Nrt2.6 Gene in Arabidopsis thaliana: A Link with Plant Response to Biotic and Abiotic Stress. PLoS ONE 2012, 7, e42491. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-Y.; Tsay, Y.-F. Arabidopsis Nitrate Transporter NRT1.9 Is Important in Phloem Nitrate Transport. Plant Cell 2011, 23, 1945–1957. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.-T.; Liu, D.-F.; Wen, T.-T.; Fang, Z.-J.; Chen, S.-Y.; Li, H.; Gong, J.-M. Vacuolar nitrate efflux requires multiple functional redundant nitrate transporter in Arabidopsis thaliana. Front. Plant Sci. 2022, 13, 926809. [Google Scholar] [CrossRef]
- Wu, X.; Liu, T.; Zhang, Y.; Duan, F.; Neuhäuser, B.; Ludewig, U.; Schulze, W.X.; Yuan, L. Ammonium and nitrate regulate NH4+ uptake activity of Arabidopsis ammonium transporter AtAMT1;3 via phosphorylation at multiple C-terminal sites. J. Exp. Bot. 2019, 70, 4919–4930. [Google Scholar] [CrossRef]
- Yuan, L.; Loqué, D.; Kojima, S.; Rauch, S.; Ishiyama, K.; Inoue, E.; Takahashi, H.; von Wirén, N. The Organization of High-Affinity Ammonium Uptake in Arabidopsis Roots Depends on the Spatial Arrangement and Biochemical Properties of AMT1-Type Transporters. Plant Cell 2007, 19, 2636–2652. [Google Scholar] [CrossRef]
- Yuan, L.; Graff, L.; Loqué, D.; Kojima, S.; Tsuchiya, Y.N.; Takahashi, H.; von Wirén, N. AtAMT1;4, a Pollen-Specific High-Affinity Ammonium Transporter of the Plasma Membrane in Arabidopsis. Plant Cell Physiol. 2009, 50, 13–25. [Google Scholar] [CrossRef]
- Zanin, L.; Tomasi, N.; Wirdnam, C.; Meier, S.; Komarova, N.Y.; Mimmo, T.; Cesco, S.; Rentsch, D.; Pinton, R. Isolation and functional characterization of a high affinity urea transporter from roots of Zea mays. BMC Plant Biol. 2014, 14, 222. [Google Scholar] [CrossRef]
- Wang, W.-H.; Köhler, B.; Cao, F.-Q.; Liu, G.-W.; Gong, Y.-Y.; Sheng, S.; Song, Q.-C.; Cheng, X.-Y.; Garnett, T.; Okamoto, M.; et al. Rice DUR3 mediates high-affinity urea transport and plays an effective role in improvement of urea acquisition and utilization when expressed in Arabidopsis. New Phytol. 2012, 193, 432–444. [Google Scholar] [CrossRef]
- Yu, S.; Pratelli, R.; Denbow, C.; Pilot, G. Suppressor mutations in the Glutamine Dumper1 protein dissociate disturbance in amino acid transport from other characteristics of the Gdu1D phenotype. Front. Plant Sci. 2015, 6, 593. [Google Scholar] [CrossRef]
- Hirner, A.; Ladwig, F.; Stransky, H.; Okumoto, S.; Keinath, M.; Harms, A.; Frommer, W.B.; Koch, W. Arabidopsis LHT1 Is a High-Affinity Transporter for Cellular Amino Acid Uptake in Both Root Epidermis and Leaf Mesophyll. Plant Cell 2006, 18, 1931–1946. [Google Scholar] [CrossRef] [PubMed]
- Havé, M.; Marmagne, A.; Chardon, F.; Masclaux-Daubresse, C. Nitrogen remobilisation during leaf senescence: Lessons from Arabidopsis to crops. J. Exp. Bot. 2017, 68, 2513–2529. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Eom, S.; Shin, K.; Lee, R.-A.; Choi, S.; Lee, J.-H.; Lee, S.; Soh, M.-S. Identification of Lysine Histidine Transporter 2 as an 1-Aminocyclopropane Carboxylic Acid Transporter in Arabidopsis thaliana by Transgenic Complementation Approach. Front. Plant Sci. 2019, 10, 1092. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, T.; Hirai, M.Y.; Chino, M.; Komeda, Y.; Naito, S. Effects of Sulfur Nutrition on Expression of the Soybean Seed Storage Protein Genes in Transgenic Petunia. Plant Physiol. 1992, 99, 263–268. [Google Scholar] [CrossRef]
- Johnson, J.M.; Sherameti, I.; Ludwig, A.; Nongbri, P.L.; Sun, C.; Varma, A.; Oelmüller, R. Protocols for Arabidopsis thaliana and Piriformospora indica co-cultivation: A model system to study plant beneficial traits. Endocyt. Cell Res. 2011, 21, 101–113. [Google Scholar]
- Johnson, J.M.; Sherameti, I.; Nongbri, P.L.; Oelmüller, R. Standardized conditions to study beneficial and nonbeneficial traits in the Piriformospora indica/Arabidopsis thaliana interaction. In Piriformospora indica: Sebacinales and Their Biotechnological Applications; Varma, A., Kost, G., Oelmüller, R., Eds.; Soil Biology; Springer: Berlin/Heidelberg, Germany, 2013; Volume 33, pp. 325–343. [Google Scholar]
- Pérez-Alonso, M.; Guerrero-Galán, C.; Ortega-Villaizán, A.G.; Ortiz-García, P.; Scholz, S.S.; Ramos, P.; Sakakibara, H.; Kiba, T.; Ludwig-Müller, J.; Krapp, A.; et al. The calcium sensor CBL7 is required for Serendipita indica -induced growth stimulation in Arabidopsis thaliana, controlling defense against the endophyte and K+ homoeostasis in the symbiosis. Plant Cell Environ. 2022, 45, 3367–3382. [Google Scholar] [CrossRef]
- Chen, A.; Gu, M.; Wang, S.; Chen, J.; Xu, G. Transport properties and regulatory roles of nitrogen in arbuscular mycorrhizal symbiosis. Semin. Cell Dev. Biol. 2018, 74, 80–88. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. feature Counts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Bütehorn, B.; Rhody, D.; Franken, P. Isolation and Characterisation of Pitef1 Encoding the Translation Elongation Factor EF-1α of the Root Endophyte Piriformospora indica. Plant Biol. 2000, 2, 687–692. [Google Scholar] [CrossRef]
- Scholz, S.S.; Vadassery, J.; Heyer, M.; Reichelt, M.; Bender, K.W.; Snedden, W.A.; Boland, W.; Mithöfer, A. Mutation of the Arabidopsis Calmodulin-Like Protein CML37 Deregulates the Jasmonate Pathway and Enhances Susceptibility to Herbivory. Mol. Plant 2014, 7, 1712–1726. [Google Scholar] [CrossRef]
- Forzani, C.; Duarte, G.T.; Van Leene, J.; Clément, G.; Huguet, S.; Paysant-Le-Roux, C.; Mercier, R.; De Jaeger, G.; Leprince, A.-S.; Meyer, C. Mutations of the AtYAK1 Kinase Suppress TOR Deficiency in Arabidopsis. Cell Rep. 2019, 27, 3696–3708.e5. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| (A) | Metabolite Ratio Limiting vs. Sufficient N Supply | |||
|---|---|---|---|---|
| without P. indica | with P. indica | |||
| Compound | Mean | SE | Mean | SE |
| Aspartate | 0.36 | 0.07 | 0.44 | 0.17 |
| Alanine | 0.50 | 0.06 | 0.66 | 0.14 |
| Homoserine | 0.69 | 0.17 | 0.55 | 0.10 |
| Glutamine | 0.89 | 0.24 | 0.64 | 0.19 |
| Glutamate | 0.98 | 0.21 | 0.76 | 0.07 |
| Glycine | 1.10 | 0.46 | 0.72 | 0.12 |
| Asparagine | 1.22 | 0.39 | 0.67 | 0.11 |
| Proline | 1.45 | 0.19 | 1.42 | 0.05 |
| Cystein | 1.45 | 0.30 | 1.20 | 0.04 |
| Methionine | 1.51 | 0.71 | 0.72 | 0.04 |
| Agmatine(-NH3) | 1.64 | 0.27 | 1.40 | 0.52 |
| beta-Alanine | 1.67 | 0.46 | 1.25 | 0.12 |
| Threonine | 1.86 | 0.57 | 2.64 | 0.73 |
| Valine | 2.05 | 0.59 | 1.55 | 0.22 |
| Arginine | 2.08 | 0.86 | 0.98 | 0.33 |
| Leucine | 2.31 | 0.50 | 1.82 | 0.41 |
| Histidine | 2.46 | 0.88 | 2.13 | 0.97 |
| Tyrosine | 2.64 | 0.38 | 2.46 | 0.65 |
| Phenylalanine | 2.65 | 0.99 | 1.46 | 0.22 |
| Tryptophan | 2.70 | 0.99 | 2.02 | 0.48 |
| Lysine | 2.78 | 0.76 | 2.03 | 0.65 |
| Isoleucine | 3.24 | 1.10 | 2.04 | 0.45 |
| Serine | 4.40 | 0.42 | 6.00 | 2.75 |
| (B) | Metabolite Ratio Limiting vs. Sufficient N Supply | |||
| without P.indica | with P. indica | |||
| Compound | Mean | SE | Mean | SE |
| Rhamnose | 1.22 | 0.08 | 1.20 | 0.09 |
| Arabinose | 1.24 | 0.19 | 1.56 | 0.32 |
| Gentiobiose | 1.29 | 0.17 | 1.41 | 0.13 |
| Ribose | 1.32 | 0.07 | 1.61 | 0.24 |
| Xylose | 1.42 | 0.32 | 2.21 | 0.58 |
| Mannose | 1.43 | 0.28 | 1.89 | 0.15 |
| Galactose | 1.44 | 0.34 | 2.02 | 0.22 |
| Sucrose | 1.55 | 0.37 | 2.30 | 0.77 |
| Glucose | 1.59 | 0.67 | 3.34 | 0.73 |
| Fructose | 1.72 | 0.63 | 3.32 | 0.75 |
| Maltose | 2.22 | 1.20 | 1.45 | 0.29 |
| Trehalose | 2.88 | 2.21 | 2.02 | 0.54 |
| Raffinose | 3.46 | 1.55 | 10.21 | 3.77 |
 | ||||
| DEGs | Shoot | Root | ||||
|---|---|---|---|---|---|---|
| Category | Gene | Atg Number | −N vs. +N | −N vs. +N | −N vs. +N | −N vs. +N |
| w/o P. indica | w P. indica | w/o P. indica | w P. indica | |||
| Nitrate (NRT2 family) | AtNRT2.2 | At1g08100 | x | 3.68 | x | x |
| Nitrate (NRT2 family) | AtNRT2.3 | At5G60780 | −1.55 | −5.8 | x | −5.22 |
| Nitrate (NRT2 family) | AtNRT2.4 | At5g60770 | x | 3.61 | 2.9 | 2.58 |
| Nitrate (NRT2 family) | AtNRT2.5 | At1g12940 | 2.17 | 3.85 | 5.63 | 3.61 |
| Nitrate (NRT2 family) | AtNRT2.6 | At3g45060 | x | x | −3.85 | −5.14 |
| Nitrate (NPF family) | NPF2.6 | At3g45660 | x | 3.34 | x | x |
| Nitrate (NPF family) | NPF2.8/NRT1.9 | At5g28470 | x | −4.49 | x | x |
| Nitrate (NPF family) | NPF2.13/NRT1.7 | At1g69870 | x | 2.48 | x | x |
| Nitrate (NPF family) | NPF4.1/AIT3 | At3g25260 | x | x | x | 2.3 |
| Nitrate (NPF family) | NPF5.3/NRT1.8 | At5g46040 | x | 4.1 | x | x |
| Nitrate (NPF family) | NPF5.6 | At2g37900 | x | x | −3.57 | x |
| Nitrate (NPF family) | NPF5.12 | At1g72140 | −2.03 | x | x | x |
| Nitrate (NPF family) | NPF5.14/NRT1.15 | At1g72120 | x | 1.83 | x | x |
| Nitrate (NPF family) | NPF6.2/NRT1.4 | At2g26690 | x | x | −2.11 | −1.61 |
| Ammonium (AMT family) | AMT1−3 | At3g24300 | x | 2.42 | x | X |
| Ammonium (AMT family) | AMT1−4 | At4g28700 | x | x | x | 2.51 |
| Ammonium (AMT family) | AMT1−5 | At3g24290 | x | 4.18 | 4.12 | 2.51 |
| Urea | DUR3 | At5g45380 | x | 2.46 | 2.87 | 2.04 |
| Amino acid (GDU family) | GDU1 | At4g31730 | x | x | −2.1 | x |
| Amino acid (GDU family) | GDU4 | At2g24762 | −1.77 | −1.99 | −2.54 | −1.96 |
| Amino acid (GDU family) | GDU5 | At5g24920 | x | x | −1.8 | x |
| Amino acid (GDU family) | GDU6 | At3g30725 | x | x | −2.89 | −2.03 |
| Amino acid (GDU family) | GDU7 | At5g38770 | x | x | −1.81 | x |
| Amino acid (LHT family) | LHT1 | At5g40780 | x | 2.16 | x | x |
| Amino acid (LHT family) | LHT2/AATL2 | At1g24400 | x | x | x | −2.06 |
| Amino acid (LHT family) | LHT3 | At1g61270 | x | x | x | 1.53 |
| Amino acid (LHT family) | LHT7 | At4g35180 | 2.09 | 2.52 | x | X |
| Amino acid (AAP family) | AAP3 | At1g77380 | x | 1.84 | x | X |
| Amino acid (AAP family) | AAP4 | At5g63850 | x | 2.15 | x | X |
| Amino acid (AAP family) | AAP6 | At5g49630 | x | 1.9 | x | X |
| Amino acid (AAP family) | AAP7 | At5g23810 | x | x | x | 1.53 |
| Amino acid (AVT family) | AVT1E | At5g02170 | x | −4.51 | −1.77 | X |
| Amino acid (AVT family) | AVT1H | At5g16740 | 6.41 | 7.5 | 2.25 | 2.72 |
| Amino acid (AVT family) | AVT3B | At2g42005 | −2.89 | −1.65 | x | X |
| Amino acid (CAT family) | GAT1/BAT1 | At1g08230 | x | 2.02 | x | X |
| Amino acid (CAT family) | CAT1/AAT1 | At4g21120 | 1.65 | 3.39 | x | X |
| Amino acid (CAT family) | CAT5 | At2g34960 | x | 2.18 | x | X |
| Amino acid (UmamiT family) | UmamiT 4 | At3G18200 | x | 4.31 | x | X |
| Amino acid (UmamiT family) | UmamiT 8 | At4G16620 | x | 1.99 | −1.69 | X |
| Amino acid (UmamiT family) | UmamiT 10 | At3G56620 | x | 1.89 | x | X |
| Amino acid (UmamiT family) | UmamiT 13 | At2G37450 | x | −2.07 | x | −1.74 |
| Amino acid (UmamiT family) | UmamiT 14 | At2G39510 | x | x | −1.54 | X |
| Amino acid (UmamiT family) | UmamiT 17 | At4G08300 | x | 1.76 | x | X |
| Amino acid (UmamiT family) | UmamiT 19 | At1G21890 | x | 2.27 | x | 3.02 |
| Amino acid (UmamiT family) | UmamiT 20 | At4G08290 | x | 2 | −2.01 | −1.57 |
| Amino acid (UmamiT family) | UmamiT 25 | At1G09380 | x | 2.4 | x | X |
| Amino acid (UmamiT family) | UmamiT 26 | At1G11460 | x | −1.9 | x | X |
| Amino acid (UmamiT family) | UmamiT 29 | At4G01430 | x | 1.57 | x | X |
| Amino acid (UmamiT family) | UmamiT 35 | At1G60050 | x | 1.75 | −2.5 | X |
| Amino acid (UmamiT family) | UmamiT 36 | At1G70260 | x | x | x | 1.95 |
| Amino acid (UmamiT family) | UmamiT 40 | At5G40240 | x | 2.14 | x | X |
| Amino acid (UmamiT family) | UmamiT 42 | At5G40210 | x | 1.84 | x | X |
| Amino acid (UmamiT family) | UmamiT 43 | At3G28060 | x | −2.34 | x | X |
| Amino acid (UmamiT family) | UmamiT 45 | At3G28100 | x | 1.85 | x | X |
| Amino acid (UmamiT family) | UmamiT 46 | At3G28070 | x | x | −5.76 | X |
| Amino acid (UmamiT family) | UmamiT 47 | At3G28080 | x | x | −2.94 | X |
 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scholz, S.S.; Barth, E.; Clément, G.; Marmagne, A.; Ludwig-Müller, J.; Sakakibara, H.; Kiba, T.; Vicente-Carbajosa, J.; Pollmann, S.; Krapp, A.; et al. The Root-Colonizing Endophyte Piriformospora indica Supports Nitrogen-Starved Arabidopsis thaliana Seedlings with Nitrogen Metabolites. Int. J. Mol. Sci. 2023, 24, 15372. https://doi.org/10.3390/ijms242015372
Scholz SS, Barth E, Clément G, Marmagne A, Ludwig-Müller J, Sakakibara H, Kiba T, Vicente-Carbajosa J, Pollmann S, Krapp A, et al. The Root-Colonizing Endophyte Piriformospora indica Supports Nitrogen-Starved Arabidopsis thaliana Seedlings with Nitrogen Metabolites. International Journal of Molecular Sciences. 2023; 24(20):15372. https://doi.org/10.3390/ijms242015372
Chicago/Turabian StyleScholz, Sandra S., Emanuel Barth, Gilles Clément, Anne Marmagne, Jutta Ludwig-Müller, Hitoshi Sakakibara, Takatoshi Kiba, Jesús Vicente-Carbajosa, Stephan Pollmann, Anne Krapp, and et al. 2023. "The Root-Colonizing Endophyte Piriformospora indica Supports Nitrogen-Starved Arabidopsis thaliana Seedlings with Nitrogen Metabolites" International Journal of Molecular Sciences 24, no. 20: 15372. https://doi.org/10.3390/ijms242015372
APA StyleScholz, S. S., Barth, E., Clément, G., Marmagne, A., Ludwig-Müller, J., Sakakibara, H., Kiba, T., Vicente-Carbajosa, J., Pollmann, S., Krapp, A., & Oelmüller, R. (2023). The Root-Colonizing Endophyte Piriformospora indica Supports Nitrogen-Starved Arabidopsis thaliana Seedlings with Nitrogen Metabolites. International Journal of Molecular Sciences, 24(20), 15372. https://doi.org/10.3390/ijms242015372