
The Effect of Temperature on the Embryo Development of Cephalopod Sepiella japonica Suggests Crosstalk between Autophagy and Apoptosis

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Mortality Rate and Cumulative Survival Rate
2.2. Effects of Different Temperatures on the Embryonic Development Duration
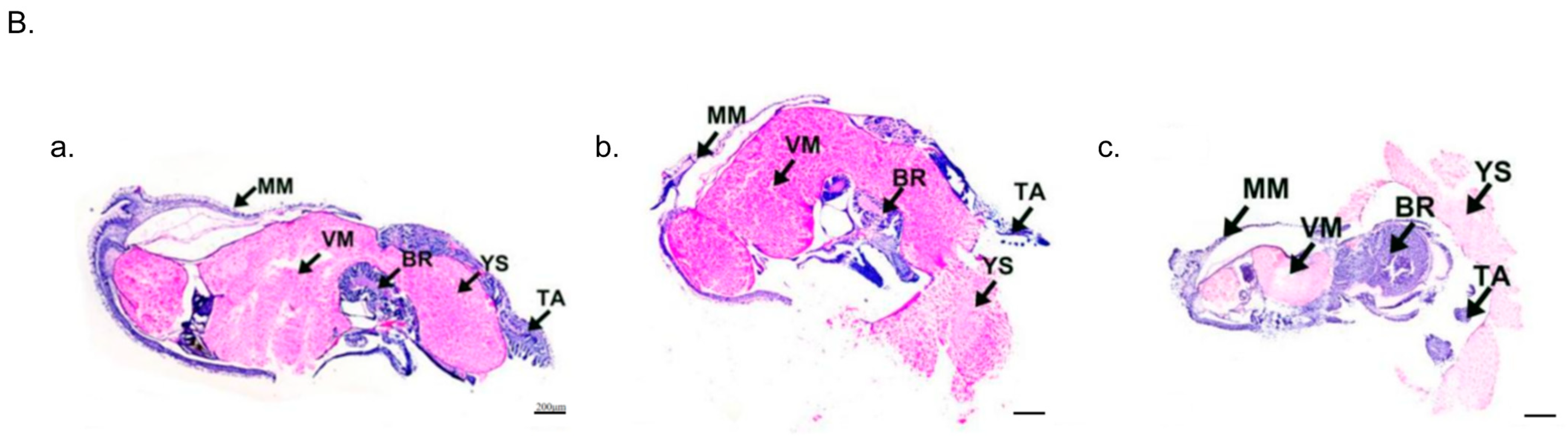
2.3. Embryonic Malformation Caused by Low and High Temperatures
2.4. Expression of Autophagy Marker Genes LC3/BECN1 of the Embryos
2.5. Expression of Autophagy-Related Gene Inx4 of the Embryos
2.6. Expression of Apoptosis Marker Genes Cas3/p53 of the Embryos
2.7. Heat Map of the Gene Expression Elevated Folds
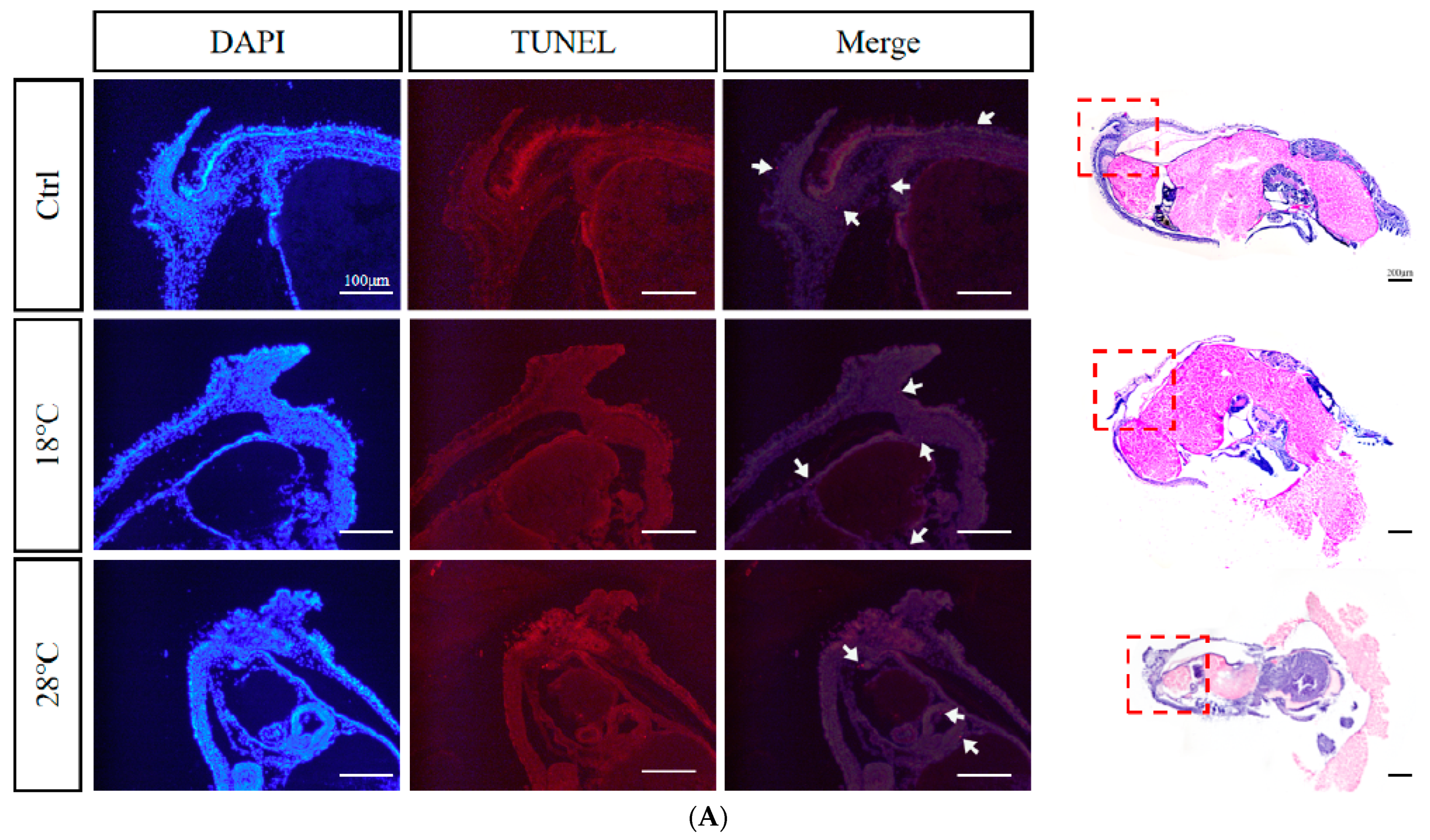
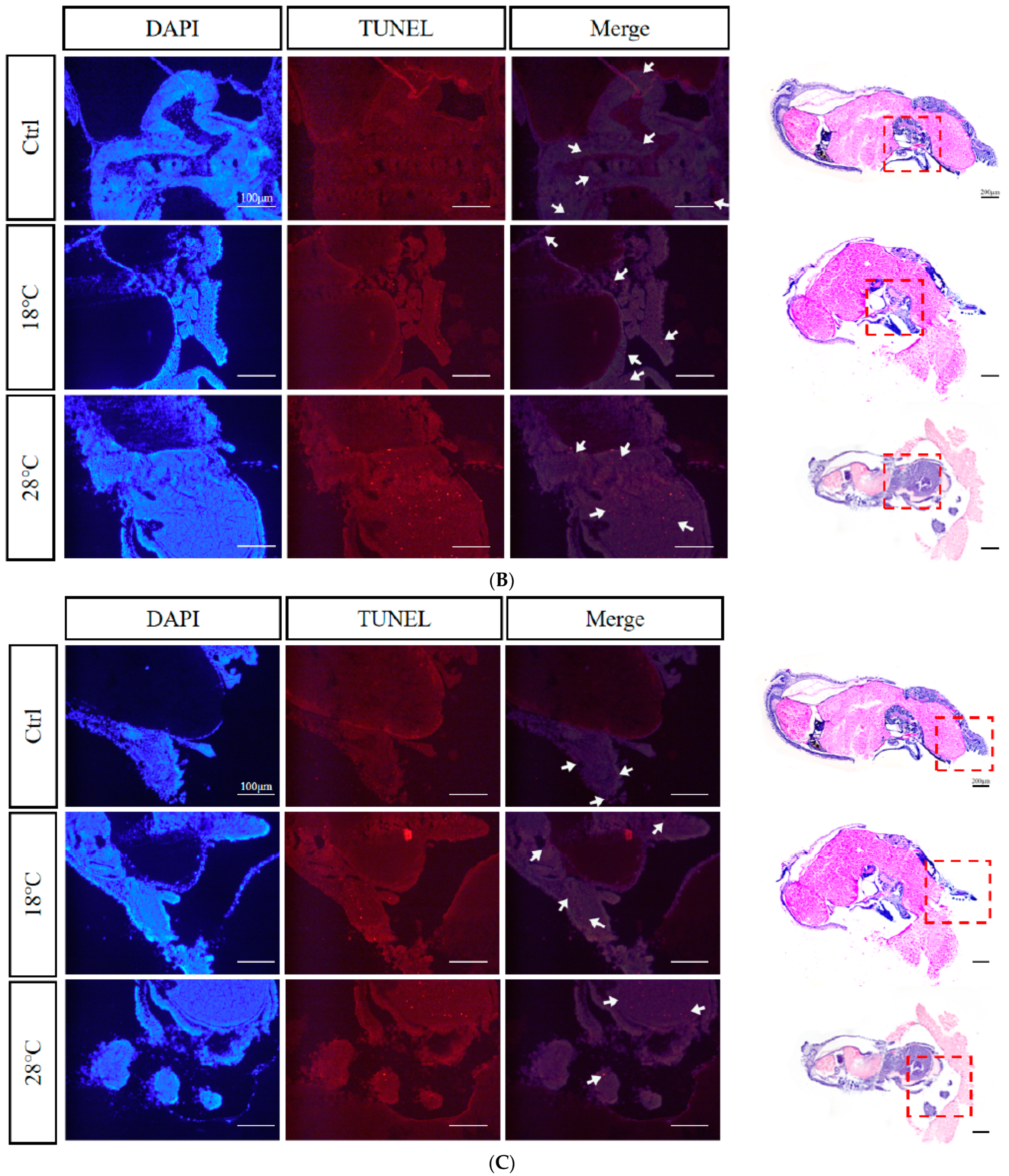
2.8. Apoptotic Staining (TUNEL) of Embryos at Sensitive Stages
3. Discussion
4. Materials and Methods
4.1. Embryo Culture and Sampling
4.2. Embryonic Development Observation and Histological Sections
4.3. RNA Extraction and cDNA Synthesis
4.4. Real-Time Fluorescence Quantitative PCR Assay
4.5. Apoptotic Staining (TUNEL)
4.6. Data Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duan, Z.; Duan, X.; Zhao, S.; Wang, X.; Wang, J.; Liu, Y.; Peng, Y.; Gong, Z.; Wang, L. Barrier function of zebrafish embryonic chorions against microplastics and nanoplastics and its impact on embryo development. J. Hazard. Mater. 2020, 395, 122621. [Google Scholar] [CrossRef]
- Jiang, X.M.; Lu, Z.R.; He, H.J.; Ye, B.L.; Ying, Z.; Wang, C.L. Effects of several ecological factors on the hatching of Sepiella japonica wild and cultured eggs. Chin. J. App. Ecol. 2010, 21, 1321–1326. (In Chinese) [Google Scholar]
- Zhang, J.S.; Chi, C.F.; Wu, C.W. Biological zero temperature and effective accumulated temperature for embryonic development of Sepiella japonica. South China Fish Sci. 2011, 7, 45–49. (In Chinese) [Google Scholar]
- Jiang, D.; Zheng, X.; Qian, Y.; Zhang, Q. Development of Amphioctopus fangsiao (Mollusca: Cephalopoda) from eggs to hatchlings: Indications for the embryonic developmental management. Mar. Life Sci. Technol. 2020, 2, 24–30. [Google Scholar] [CrossRef]
- Repolho, T.; Baptista, M.; Pimentel, M.S.; Dionísio, G.; Trübenbach, K.; Lopes, V.M.; Lopes, A.R.; Calado, R.; Diniz, M.; Rosa, R. Developmental and physiological challenges of octopus (Octopus vulgaris) early life stages under ocean warming. J. Comp. Physiol. B 2014, 184, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.Y.; Su, Y.Z.; Xie, Y.Q.; Zhou, R.F. Preliminary observation on embryonic development of Sepiella japonica. Prog Fish Sci. 2009, 30, 13–19. (In Chinese) [Google Scholar]
- Glick, D.; Barth, S.; MacLeod, K.F. Autophagy: Cellular and molecular mechanisms. J. Pathol. 2010, 221, 3–12. [Google Scholar] [CrossRef]
- Mizushima, N.; Levine, B.; Cuervo, A.M.; Klionsky, D.J. Autophagy fights disease through cellular selfdigestion. Nature 2008, 451, 1069–1075. [Google Scholar] [CrossRef]
- Wirawan, E.; Vanden Berghe, T.; Lippens, S.; Agostinis, P.; Vandenabeele, P. Autophagy: For better or for worse. Cell Res. 2012, 22, 43–61. [Google Scholar] [CrossRef]
- Varshavsky, A. The Ubiquitin System, Autophagy, and Regulated Protein Degradation. Annu. Rev. Biochem. 2017, 86, 123–128. [Google Scholar] [CrossRef]
- Chen, Y.; Culetto, E.; Legouis, R. A DRP-1 dependent autophagy process facilitates rebuilding of the mitochondrial network and modulates adaptation capacity in response to acute heat stress during C. elegans development. Autophagy 2021, 17, 2654–2655. [Google Scholar] [CrossRef] [PubMed]
- Kassis, S.; Grondin, M.; Averill-Bates, D.A. Heat shock increases levels of reactive oxygen species, autophagy and apoptosis. BBA-Mol. Cell Res. 2020, 1868, 118924. [Google Scholar] [CrossRef] [PubMed]
- Mccormick, J.J.; King, K.E.; Côté, M.D.; Meade, R.D.; Akerman, A.P.; Kenny, G.P. Impaired autophagy following ex vivo heating at physiologically relevant temperatures in peripheral blood mononuclear cells from elderly adults. J. Therm. Biol. 2020, 95, 102790. [Google Scholar] [CrossRef] [PubMed]
- Dündar, G.; Shao, Z.; Higashitani, N.; Kikuta, M.; Izumi, M.; Higashitani, A. Autophagy mitigates high-temperature injury in pollen development of Arabidopsis thaliana. Dev. Biol. 2019, 456, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.L.; Chen, L.B.; Hu, C.F.; Han, B.S. Research on the role of autophagy in cell cold stress. Chin. J. Cell Biol. 2016, 38, 1077–1083. (In Chinese) [Google Scholar]
- Molina, A.; Dettleff, P.; Valenzuela-Muñoz, V.; Gallardo-Escarate, C.; Valdés, J.A. High-Temperature Stress Induces Autophagy in Rainbow Trout Skeletal Muscle. Fishes 2023, 8, 303. [Google Scholar] [CrossRef]
- Ruperez, C.; Blasco-Roset, A.; Kular, D.; Cairo, M.; Ferrer-Curriu, G.; Villarroya, J.; Zamora, M.; Crispi, F.; Villarroya, F.; Planavila, A. Autophagy is Involved in Cardiac Remodeling in Response to Environmental Temperature Change. Front. Physiol. 2022, 13, 864427. [Google Scholar] [CrossRef]
- Das, S.; Shukla, N.; Singh, S.S.; Kushwaha, S.; Shrivastava, R. Mechanism of interaction between autophagy and apoptosis in cancer. Apoptosis 2021, 26, 512–533. [Google Scholar] [CrossRef]
- Qu, X.; Zou, Z.; Sun, Q.; Luby-Phelps, K.; Cheng, P.; Hogan, R.N.; Gilpin, C.; Levine, B. Autophagy gene-dependent clearance of apoptotic cells during embryonic development. Cell 2007, 128, 833–836. [Google Scholar] [CrossRef]
- Xu, D.; Jiang, X.; He, H.; Liu, D.; Yang, L.; Chen, H.; Wu, L.; Geng, G.; Li, Q. SIRT2 functions in aging, autophagy, and apoptosis in post-maturation bovine oocytes. Life Sci. 2019, 232, 116639. [Google Scholar] [CrossRef]
- Kanduc, D.; Mittelman, A.; Serpico, R.; Sinigaglia, E.; Sinha, A.A.; Natale, C.; Santacroce, R.; Di Corcia, M.G.; Lucchese, A.; Dini, L.; et al. Cell death: Apoptosis versus necrosis (Review). Int. J. Oncol. 2022, 21, 165–170. [Google Scholar] [CrossRef]
- Kaur, A.P.; Agrawal, S. A review of the molecular mechanism of apoptosis and its role in pathological conditions. Int. J. Pharm. Bio Sci. 2019, 10, 124–131. [Google Scholar] [CrossRef]
- Cheng, C.-H.; Yang, F.-F.; Liao, S.-A.; Miao, Y.-T.; Ye, C.-X.; Wang, A.-L.; Tan, J.-W.; Chen, X.-Y. High temperature induces apoptosis and oxidative stress in pufferfish (Takifugu obscurus) blood cells. J. Therm. Biol. 2015, 53, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Kim, J.; Lee, D.-W.; Park, Y.-S.; Kim, J.-H.; Choi, C.Y. Oxidative stress and apoptosis in disk abalone (Haliotis discus hannai) caused by water temperature and pH changes. Antioxidants 2023, 12, 1003. [Google Scholar] [CrossRef]
- Li, A.J.; Leung, P.T.; Bao, V.W.; Lui, G.C.; Leung, K.M. Temperature-dependent physiological and biochemical responses of the marine medaka Oryzias melastigma with consideration of both low and high thermal extremes. J. Therm. Biol. 2014, 54, 98–105. [Google Scholar] [CrossRef]
- Shahabad, Z.A.; Avci, C.B.; Bani, F.; Zarebkohan, A.; Sadeghizadeh, M.; Salehi, R.; Ghafarkhani, M.; Rahbarghazi, R.; Bagca, B.G.; Ozates, N.P. Photothermal effect of albumin-modified gold nanorods diminished neuroblastoma cancer stem cells dynamic growth by modulating autophagy. Sci. Rep. 2022, 12, 11774. [Google Scholar] [CrossRef]
- Bauer, R.; Löer, B.; Ostrowski, K.; Martini, J.; Weimbs, A.; Lechner, H.; Hoch, M. Intercellular communication: The Drosophila innexin multiprotein family of gap junction proteins. Chem. Biol. 2005, 12, 515–526. [Google Scholar] [CrossRef]
- Beyer, E.C.; Berthoud, V.M. Gap junction gene and protein families: Connexins, innexins, and pannexins. Biochim. Biophys. Acta Biomembr. 2017, 1860, 5–8. [Google Scholar] [CrossRef]
- Akopian, A.; Atlasz, T.; Pan, F.; Wong, S.; Zhang, Y.; Völgyi, B.; Paul, D.L.; Bloomfield, S.A. Gap junction-mediated death of retinal neurons is connexin and insult specific: A potential target for neuroprotection. J. Neurosci. 2014, 34, 10582–10591. [Google Scholar] [CrossRef]
- Huang, R.P.; Hossain, M.Z.; Huang, R.; Gano, J.; Fan, Y.; Boynton, A.L. Connexin 43 (Cx43) enhances chemotherapy-induced apoptosis in human glioblastoma cells. Int. J. Cancer 2001, 92, 130–138. [Google Scholar] [CrossRef]
- Seul, K.H.; Kang, K.Y.; Lee, K.S.; Kim, S.H.; Beyer, E.C. Adenoviral delivery of human connexin37 induces endothelial cell death through apoptosis. Biochem. Biophys. Res. Commun. 2004, 319, 1144–1151. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Fraizer, G.; De La Cerda, J.; Cristiano, R.; Liebert, M.; Grossman, H. Connexin 26 enhances the bystander effect in HSVtk/GCV gene therapy for human bladder cancer by adenovirus/PLL/DNA gene delivery. Gene Ther. 2001, 8, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Bejarano, E.; Yuste, A.; Patel, B.; Stout, R.F., Jr.; Spray, D.C.; Cuervo, A.M. Connexins modulate autophagosome biogenesis. Nat. Cell Biol. 2014, 16, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Gao, J.; Zhao, M.; Cui, J.; Li, Y.; Yang, X.; Jing, X.; Wu, Z. A novel cognitive impairment mechanism that astrocytic p-connexin 43 promotes neuronic autophagy via activation of P2X7R and down-regulation of GLT-1 expression in the hippocampus following traumatic brain injury in rats. Behav. Brain Res. 2015, 291, 315–324. [Google Scholar] [CrossRef]
- MolluscaBase. Cephalopoda. 2023. Available online: http://www.molluscabase.org (accessed on 8 August 2023).
- Domingues, P.M.; Bettencourt, V.; Guerra, A. Growth of Sepia officinalis in captivity and in nature. Vie Milieu 2006, 56, 109–120. [Google Scholar] [CrossRef]
- Liu, L.; Zhao, S.; Zhang, Y.; Wang, M.; Yan, Y.; Lv, Z.; Gong, L.; Liu, B.; Dong, Y.; Xu, Z. Identification of Vitellogenin 1 Potentially Related to Reproduction in the Cephalopod, Sepiella japonica. J. Shellfish. Res. 2022, 41, 235–241. [Google Scholar] [CrossRef]
- Lv, Z.; Zhu, K.; Pang, Z.; Liu, L.; Jiang, L.; Liu, B.; Shi, H.; Ping, H.; Chi, C.; Gong, L. Identification, characterization and mRNA transcript abundance profiles of estrogen related receptor (ERR) in Sepiella japonica imply its possible involvement in female reproduction. Anim. Reprod. Sci. 2019, 211, 106231. [Google Scholar] [CrossRef]
- Pang, Z.; Zhang, Y.; Liu, L. Identification and functional characterization of interferon-γ-inducible lysosomal thiol reductase (GILT) gene in common Chinese cuttlefish Sepiella japonica. Fish Shellfish. Immunol. 2019, 86, 627–634. [Google Scholar] [CrossRef]
- Wada, T.; Takegaki, T.; Mori, T.; Natsukari, Y. Reproductive Behavior of the Japanese Spineless Cuttlefish Sepiella japonica. Venus 2018, 65, 221–228. [Google Scholar] [CrossRef]
- Vagner, M.; Zambonino-Infante, J.-L.; Mazurais, D. Fish facing global change: Are early stages the lifeline? Mar. Environ. Res. 2019, 147, 159–178. [Google Scholar] [CrossRef]
- Soman, M.; Chadha, N.K.; Madhu, K.; Madhu, R.; Sawant, P.B.; Francis, B. Optimization of temperature improves embryonic development and hatching efficiency of false clown fish, Amphiprion ocellaris Cuvier, 1830 under captive condition. Aquaculture 2021, 536, 736417. [Google Scholar] [CrossRef]
- Li, D.; Dorber, M.; Barbarossa, V.; Verones, F. Global characterization factors for quantifying the impacts of increasing water temperature on freshwater fish. Ecol. Indic. 2022, 142, 109201. [Google Scholar] [CrossRef]
- Fopp-Bayat, D.; Chandra, G.; Nitkiewicz, A. How Cold Shock Affects Ploidy Level and Early Ontogenetic Development of the Sterlet, A. ruthenus L. Int. J. Mol. Sci. 2022, 23, 494. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.-P.; Wei, C.-Q.; Zhao, W.-H.; Du, H.-J.; Chen, Y.-L.; Gui, J.-F. Effects of incubation temperatures on embryonic development in the Asian yellow pond turtle. Aquaculture 2006, 259, 243–248. [Google Scholar] [CrossRef]
- Braga, R.; Van der Molen, S.; Pontones, J.; Ortiz, N. Embryonic development, hatching time and newborn juveniles of Octopus tehuelchus under two culture temperatures. Aquaculture 2020, 530, 735778. [Google Scholar] [CrossRef]
- Dahlke, F.T.; Wohlrab, S.; Butzin, M.; Pörtner, H.-O. Thermal bottlenecks in the life cycle define climate vulnerability of fish. Science 2020, 369, 65–70. [Google Scholar] [CrossRef]
- Guiet, J.; Aumont, O.; Poggiale, J.-C.; Maury, O. Effects of lower trophic level biomass and water temperature on fish communities: A modelling study. Prog. Oceanogr. 2016, 146, 22–37. [Google Scholar] [CrossRef]
- Gao, W.-J.; Li, H.-X.; Feng, J.; Lu, X.-R.; Yin, P.-L.; Jia, H.; Ma, W.-Z. Transcriptome Analysis in High Temperature Inhibiting Spermatogonial Stem Cell Differentiation In Vitro. Reprod. Sci. 2023, 30, 1938–1951. [Google Scholar] [CrossRef]
- Li, X.; Shi, C.; Yang, B.; Li, Q.; Liu, S. High temperature aggravates mortalities of the Pacific oyster (Crassostrea gigas) infected with Vibrio: A perspective from homeostasis of digestive microbiota and immune response. Aquaculture 2023, 568, 739309. [Google Scholar] [CrossRef]
- Obrador, E.; Jihad-Jebbar, A.; Salvador-Palmer, R.; López-Blanch, R.; Oriol-Caballo, M.; Moreno-Murciano, M.P.; Navarro, E.A.; Cibrian, R.; Estrela, J.M. Externally applied electromagnetic fields and hyperthermia irreversibly damage cancer cells. Cancers 2023, 15, 3413. [Google Scholar] [CrossRef]
- Boletzky, S. Biology of early life stages in cephalopod molluscs. Adv. Mar. Biol. 2003, 44, 143–203. [Google Scholar] [CrossRef]
- Boletzky, S.V. Embryonic phase. In Cephalopod Life Cycles; Boyle, P.R., Ed.; Acad Press: London, UK, 1987; Volume 2, pp. 5–31. [Google Scholar]
- Boletzky, S. Embryonic development of cephalopods at low temperatures. Antarct. Sci. 1994, 6, 139–142. [Google Scholar] [CrossRef]
- Radonic, M.; López, A.V.; Oka, M.; Aristizábal, E. Effect of the incubation temperature on the embryonic development and hatching time of eggs of the red porgy Pagrus pagrus (Linne, 1758) (Pisces: Sparidae). Rev. Biol. Mar. Oceanogr. 2005, 40, 91–99. [Google Scholar] [CrossRef]
- Pype, C.; Verbueken, E.; Saad, M.A.; Casteleyn, C.R.; Van Ginneken, C.J.; Knapen, D.; Van Cruchten, S.J. Incubation at 32.5 °C and above causes malformations in the zebrafish embryo. Reprod. Toxicol. 2015, 56, 56–63. [Google Scholar] [CrossRef]
- Paredes-Molina, F.J.; Chaparro, O.R.; Navarro, J.M.; Cubillos, V.M.; Montory, J.A.; Pechenik, J.A. Embryonic encapsulated development of the gastropod Acanthina monodon is impacted by future environmental changes of temperature and pCO2. Mar. Environ. Res. 2023, 187, 105971. [Google Scholar] [CrossRef] [PubMed]
- Uriarte, I.; Espinoza, V.; Herrera, M.; Zúñiga, O.; Olivares, A.; Carbonell, P.; Pino, S.; Farías, A.; Rosas, C. Effect of temperature on embryonic development of Octopus mimus under controlled conditions. J. Exp. Mar. Biol. Ecol. 2012, 416–417, 168–175. [Google Scholar] [CrossRef]
- Nande, M.; Domingues, P.; Rosas, C. Effects of Temperature on the Embryonic Development of Octopus vulgaris. J. Shellfish. Res. 2018, 37, 1013–1019. [Google Scholar] [CrossRef]
- Aritaki, M.; Ohta, K.; Hotta, Y.; Tagawa, M.; Tanaka, M. Temperature effects on larval development and occurrence of metamorphosis-related morphological abnormalities in hatchery-reared spotted halibut Verasper variegatus juveniles. Nippon. Suisan Gakkaishi 2004, 70, 8–15, (In Japanese with English abstract). [Google Scholar] [CrossRef][Green Version]
- Tsuji, M.; Abe, H.; Hanyuu, K.; Kuriyama, I.; Tsuchihashi, Y.; Tsumoto, K.; Nigou, T.; Kasuya, T.; Katou, T.; Kawamura, T.; et al. Effect of temperature on survival, growth and malformation of cultured larvae and juveniles of the seven-band grouper Epinephelus septemfasciatus. Fish. Sci. 2013, 80, 69–81. [Google Scholar] [CrossRef]
- Doxa, C.K.; Sfakianakis, D.; Sterioti, A.; Kentouri, M. Effect of temperature on the development of deformities during the embryonic stages of Charonia seguenzae (Aradas & Benoit, 1870). J. Therm. Biol. 2021, 100, 103046. [Google Scholar] [CrossRef]
- Liu, D.J.; Chen, J.M. Study on temperature effect of early embryonic development of Coelomactra antiquata. Chin. J. Zool. 1998, 2, 2–4. (In Chinese) [Google Scholar]
- Jing, H.-R.; Luo, F.-W.; Liu, X.-M.; Tian, X.-F.; Zhou, Y. Fish oil alleviates liver injury induced by intestinal ischemia/reperfusion via AMPK/SIRT-1/autophagy pathway. World J. Gastroenterol. 2018, 24, 833–843. [Google Scholar] [CrossRef]
- Wei, J.; Ma, Z.; Li, Y.; Zhao, B.; Wang, D.; Jin, Y.; Jin, Y. miR-143 inhibits cell proliferation by targeting autophagy-related 2B in non-small cell lung cancer H1299 cells. Mol. Med. Rep. 2014, 11, 571–576. [Google Scholar] [CrossRef]
- Chen, H.; Zhao, F.; Chen, K.; Guo, Y.; Liang, Y.; Zhao, H.; Chen, S. Exposure of zebrafish to a cold environment triggered cellular autophagy in zebrafish liver. J. Fish Dis. 2022, 45, 991–1000. [Google Scholar] [CrossRef]
- Yue, Z.Y.; Horton, A.; Bravin, M.; Dejager, P.L.; Selimi, F.; Heintz, N. A novel protein complex linking the delta 2 glutamate receptor and autophagy: Implications for neurodegeneration in lurcher mice. Neuron. 2002, 35, 921–933. [Google Scholar] [CrossRef]
- Yue, Z.Y.; Jin, S.K.; Yang, C.W.; Levine, A.J.; Heintz, N. Beclin 1, an autophagy gene essential for early embryonic development, is a haploinsufficient tumor suppressor. Proc. Natl. Acad. Sci. USA 2003, 100, 15077–15082. [Google Scholar] [CrossRef]
- Hong, S.-M.; Kang, S.-W.; Goo, T.-W.; Kim, N.-S.; Lee, J.-S.; Kim, K.-A.; Nho, S.-K. Two gap junction channel (innexin) genes of the Bombyx mori and their expression. J. Insect Physiol. 2007, 54, 180–191. [Google Scholar] [CrossRef]
- Wang, S.; Chen, F.; Zhang, Y.; Ma, X.; Qiao, K. Gap junction gene innexin3 being highly expressed in the nervous system and embryonic stage of the mud crab Scylla paramamosain. J. Oceanol. Limnol. 2019, 37, 1649–1658. [Google Scholar] [CrossRef]
- Pang, Z.; Li, M.; Yu, D.; Yan, Z.; Liu, X.; Ji, X.; Yang, Y.; Hu, J.; Luo, K. Two innexins of Spodoptera litura influences hemichannel and gap junction functions in cellular immune responses. Arch. Insect Biochem. Physiol. 2015, 90, 43–57. [Google Scholar] [CrossRef]
- Liu, T.; Li, M.; Zhang, Y.; Pang, Z.; Xiao, W.; Yang, Y.; Luo, K. A role for Innexin2 and Innexin3 proteins from Spodoptera litura in apoptosis. PLoS ONE 2013, 8, e70456. [Google Scholar] [CrossRef]
- Wang, S.-P.; Chen, F.-Y.; Dong, L.-X.; Zhang, Y.-Q.; Chen, H.-Y.; Qiao, K.; Wang, K.-J. A novel innexin2 forming membrane hemichannel exhibits immune responses and cell apoptosis in Scylla paramamosain. Fish Shellfish. Immunol. 2015, 47, 485–499. [Google Scholar] [CrossRef]
- Yuan, S.; Akey, C.W. Apoptosome Structure, Assembly, and Procaspase Activation. Structure 2013, 21, 501–515. [Google Scholar] [CrossRef] [PubMed]
- Xi, H.; Wang, S.; Wang, B.; Hong, X.; Liu, X.; Li, M.; Shen, R.; Dong, Q. The role of interaction between autophagy and apoptosis in tumorigenesis. Oncol. Rep. 2022, 48, 208. [Google Scholar] [CrossRef] [PubMed]
- Maejima, Y.; Kyoi, S.; Zhai, P.; Tong, L.; Sadoshima, J. Mst1 inhibits autophagy by pro-moting Beclin1-Bcl-2 interaction. Nat. Med. 2013, 19, 1478–1488. [Google Scholar] [CrossRef] [PubMed]
- Pattingre, S.; Tassa, A.; Qu, X.; Garuti, R.; Liang, X.H.; Mizushima, N.; Packer, M.; Schneider, M.D.; Levine, B. Bcl-2 Antiapoptotic proteins inhibit beclin 1-dependent autophagy. Cell 2005, 122, 927–939. [Google Scholar] [CrossRef]
- Qiang, J.; Tao, Y.-F.; Zhu, J.-H.; Lu, S.-Q.; Cao, Z.-M.; Ma, J.-L.; He, J.; Xu, P. Effects of heat stress on follicular development and atresia in Nile tilapia (Oreochromis niloticus) during one reproductive cycle and its potential regulation by autophagy and apoptosis. Aquaculture 2022, 555, 738171. [Google Scholar] [CrossRef]
- Baghdiguian, S.; Martinand-Mari, C.; Mangeat, P. Using Ciona to study developmental programmed cell death. Semin. Cancer Biol. 2007, 17, 147–153. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, H.W.; Yao, C.L. Molecular and acute temperature stress response characterizations of caspase-8 gene in two mussels, Mytilus coruscus and Mytilus galloprovincialis. Comp. Biochem. Physiol. B. Biochem. Mol. Biol. 2014, 177–178, 10–20. [Google Scholar] [CrossRef]
- Ren, J.Y.; Shi, M.A.; Liu, R.S.; Yang, Q.H.; Johnson, T.; Skarnes, W.C.; Du, C.Y. The Birc6 (Bruce) gene regulates p53 and the mitochondrial pathway of apoptosis and is essential for mouse embryonic development. Proc. Natl. Acad. Sci. USA 2005, 102, 565–570. [Google Scholar] [CrossRef]
- Naef, A. Cephalopoda embryology. In Fauna and Flora of the Bay of Naples: Part I; Boletzky, S.V., Ed.; Smithsonian Institution Libraries: Washington, DC, USA, 1928; Volume 2, pp. 1–461. [Google Scholar]
- Zhao, X.; Yin, X.; Ma, T.; Song, W.; Jiang, L.; Zhang, X.; Liu, B.; Liu, Y.; Yan, X. The effect of chloroquine on large yellow croaker (Larimichthys crocea): From autophagy, inflammation, to apoptosis. Aquac. Rep. 2023, 28, 101457. [Google Scholar] [CrossRef]
- Huo, L.; Bao, M.; Lv, Z.; Chi, C.; Wang, T.; Liu, H. Identification, functional characterization and expression pattern of myeloid differentiation factor 88 (MyD88) in Sepiella japonica. Fish Shellfish. Immunol. 2018, 79, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Chen, L.; Meng, F.; Zhang, T.; Luo, J.; Chen, S.; Shi, H.; Liu, B.; Lv, Z. The Effect of Temperature on the Embryo Development of Cephalopod Sepiella japonica Suggests Crosstalk between Autophagy and Apoptosis. Int. J. Mol. Sci. 2023, 24, 15365. https://doi.org/10.3390/ijms242015365
Liu Y, Chen L, Meng F, Zhang T, Luo J, Chen S, Shi H, Liu B, Lv Z. The Effect of Temperature on the Embryo Development of Cephalopod Sepiella japonica Suggests Crosstalk between Autophagy and Apoptosis. International Journal of Molecular Sciences. 2023; 24(20):15365. https://doi.org/10.3390/ijms242015365
Chicago/Turabian StyleLiu, Yifan, Long Chen, Fang Meng, Tao Zhang, Jun Luo, Shuang Chen, Huilai Shi, Bingjian Liu, and Zhenming Lv. 2023. "The Effect of Temperature on the Embryo Development of Cephalopod Sepiella japonica Suggests Crosstalk between Autophagy and Apoptosis" International Journal of Molecular Sciences 24, no. 20: 15365. https://doi.org/10.3390/ijms242015365
APA StyleLiu, Y., Chen, L., Meng, F., Zhang, T., Luo, J., Chen, S., Shi, H., Liu, B., & Lv, Z. (2023). The Effect of Temperature on the Embryo Development of Cephalopod Sepiella japonica Suggests Crosstalk between Autophagy and Apoptosis. International Journal of Molecular Sciences, 24(20), 15365. https://doi.org/10.3390/ijms242015365