
A Comprehensive Genetic Study of Microtubule-Associated Gene Clusters for Male Infertility in a Taiwanese Cohort

 and
and
Abstract
:1. Introduction
2. Results
2.1. Semen Analysis
2.2. Y Chromosome Microdeletion
2.3. Variant’s Analysis and Validation in Targeted NGS
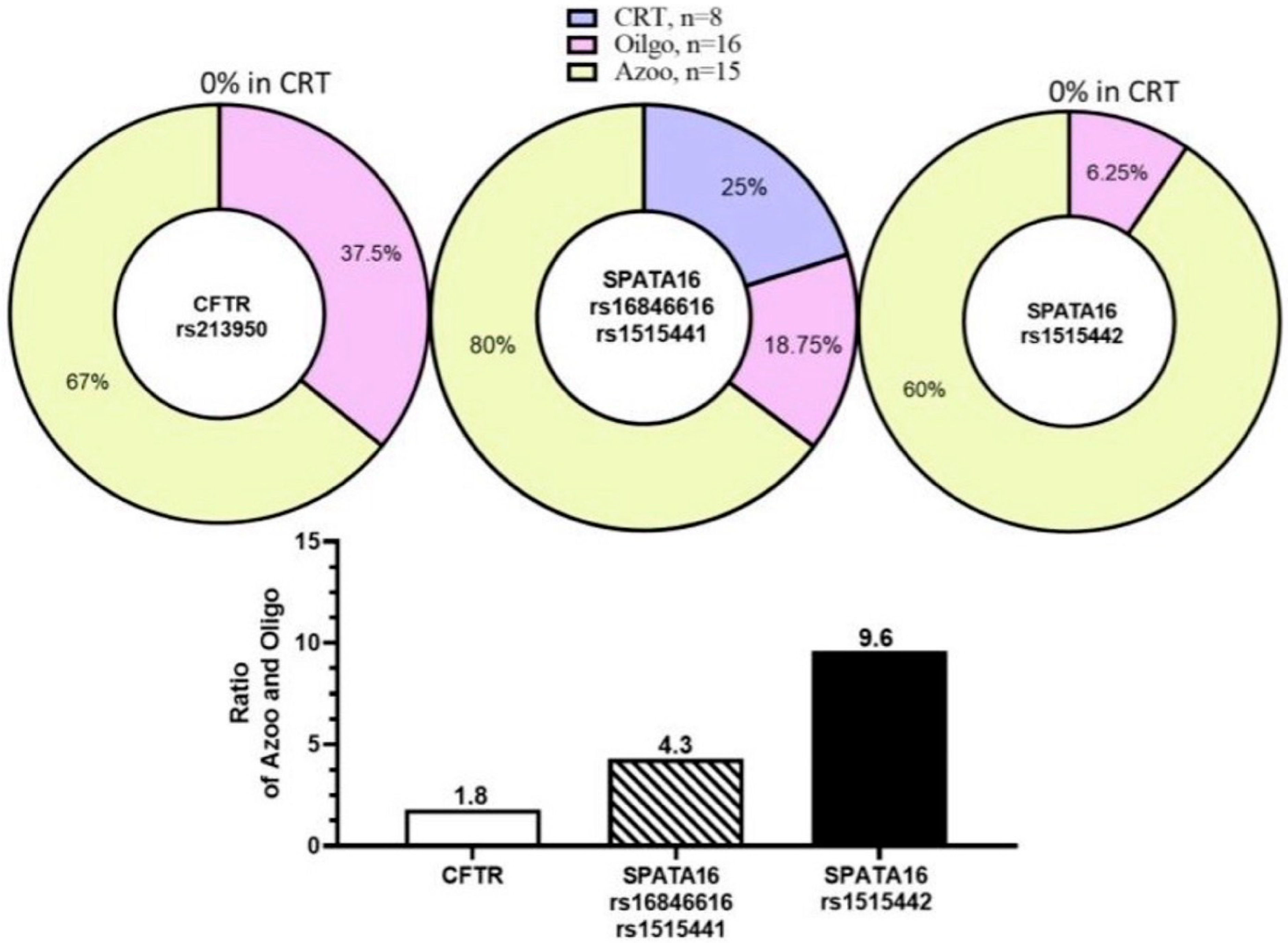
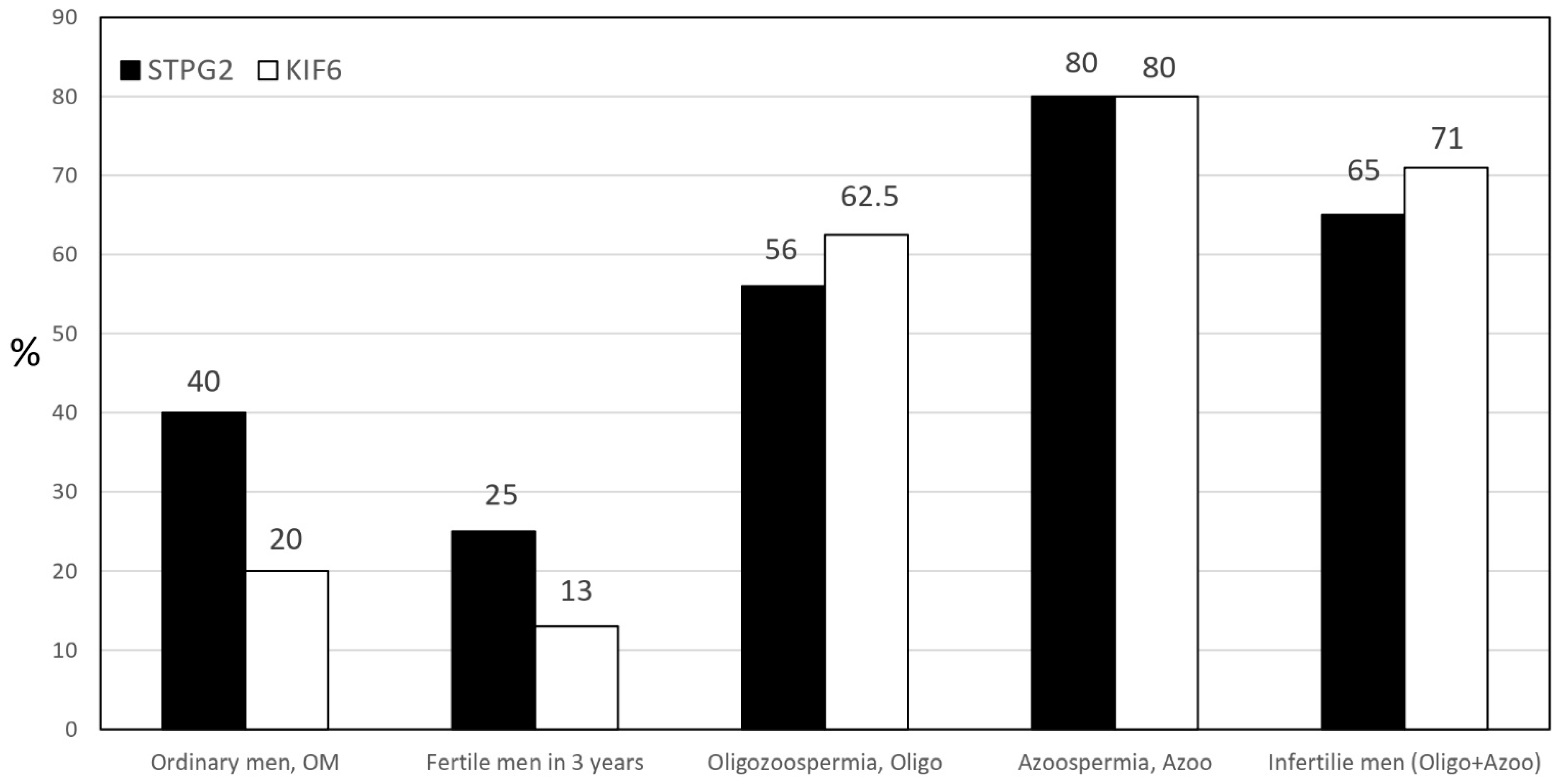
2.4. SNPs of Microtubule-Associated Genes in WES
2.5. Variant Analysis and Validation in WES
3. Discussion
3.1. Microtubule-Associated Genes Affect Spermatogenesis by Variants Analysis
3.2. Other Genes Affect Spermatogenesis by Variants Analysis
4. Materials and Methods
4.1. Patients and Controls
4.2. Semen Analysis
4.3. Y Chromosome Microdeletion (YCMD) Examination
4.4. Targeted Next-Generation Sequencing (NGS) Panel
4.5. Whole-Exome Sequencing (WES) and Variant Analysis
4.6. Validation by Sanger Sequencing
4.7. In Silico Evaluation Workflow
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Smits, R.M.; Mackenzie-Proctor, R.; Yazdani, A.; Stankiewicz, M.T.; Jordan, V.; Showell, M.G. Antioxidants for male subfertility. Cochrane Database Syst. Rev. 2019, 3, CD007411. [Google Scholar] [CrossRef]
- Tahmasbpour, E.; Balasubramanian, D.; Agarwal, A. A multi-faceted approach to understanding male infertility: Gene mutations, molecular defects and assisted reproductive techniques (ART). J. Assist. Reprod. Genet. 2014, 31, 1115–1137. [Google Scholar] [CrossRef]
- Gunes, S.; Sengupta, P.; Henkel, R.; Alguraigari, A.; Sinigaglia, M.M.; Kayal, M.; Joumah, A.; Agarwal, A. Microtubular Dysfunction and Male Infertility. World J. Men’s Health 2020, 38, 9–23. [Google Scholar] [CrossRef]
- Cho, C.; Vale, R.D. The mechanism of dynein motility: Insight from crystal structures of the motor domain. Biochim. Biophys. Acta 2012, 1823, 182–191. [Google Scholar] [CrossRef]
- Lehti, M.S.; Sironen, A. Formation and function of the manchette and flagellum during spermatogenesis. Reproduction 2016, 151, R43–R54. [Google Scholar] [CrossRef]
- Helmke, K.J.; Heald, R.; Wilbur, J.D. Interplay Between Spindle Architecture and Function. Int. Rev. Cell Mol. Biol. 2013, 306, 83–125. [Google Scholar] [CrossRef]
- Pasek, R.C.; Malarkey, E.; Berbari, N.F.; Sharma, N.; Kesterson, R.A.; Tres, L.L.; Kierszenbaum, A.L.; Yoder, B.K. Coiled-coil domain containing 42 (Ccdc42) is necessary for proper sperm development and male fertility in the mouse. Dev. Biol. 2016, 412, 208–218. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Haidl, G.; Schaefer, M.; Egner, U.; Mandal, A.; Herr, J.C. Compartmentalization of a unique ADP/ATP carrier protein SFEC (Sperm Flagellar Energy Carrier, AAC4) with glycolytic enzymes in the fibrous sheath of the human sperm flagellar principal piece. Dev. Biol. 2007, 302, 463–476. [Google Scholar] [CrossRef]
- O’Donnell, L.; Rhodes, D.; Smith, S.J.; Merriner, D.J.; Clark, B.J.; Borg, C.; Whittle, B.; O’Connor, A.E.; Smith, L.B.; McNally, F.J.; et al. An Essential Role for Katanin p80 and Microtubule Severing in Male Gamete Production. PLoS Genet. 2012, 8, e1002698. [Google Scholar] [CrossRef]
- Smith, L.B.; Milne, L.; Nelson, N.; Eddie, S.; Brown, P.; Atanassova, N.; O’Bryan, M.K.; O’Donnell, L.; Rhodes, D.; Wells, S.; et al. KATNAL1 Regulation of Sertoli Cell Microtubule Dynamics Is Essential for Spermiogenesis and Male Fertility. PLoS Genet. 2012, 8, e1002697. [Google Scholar] [CrossRef]
- Cerván-Martín, M.; Bossini-Castillo, L.; Guzmán-Jiménez, A.; Rivera-Egea, R.; Garrido, N.; Lujan, S.; Romeu, G.; Santos-Ribeiro, S.; Castilla, J.A.; Gonzalvo, M.d.C.; et al. Common genetic variation in KATNAL1 non-coding regions is involved in the susceptibility to severe phenotypes of male infertility. Andrology 2022, 10, 1339–1350. [Google Scholar] [CrossRef]
- Wu, X.; Yun, D.; Sang, M.; Liu, J.; Zhou, L.; Shi, J.; Wang, L.; Bu, T.; Li, L.; Huang, Y.; et al. Defects of microtubule cytoskeletal organization in NOA human testes. Reprod. Biol. Endocrinol. 2022, 20, 154. [Google Scholar] [CrossRef]
- Khan, R.; Zaman, Q.; Chen, J.; Khan, M.; Ma, A.; Zhou, J.; Zhang, B.; Ali, A.; Naeem, M.; Zubair, M.; et al. Novel Loss-of-Function Mutations in DNAH1 Displayed Different Phenotypic Spectrum in Humans and Mice. Front. Endocrinol. 2021, 12, 765639. [Google Scholar] [CrossRef]
- O’Donnell, L.; O’Bryan, M.K. Microtubules and spermatogenesis. Semin. Cell Dev. Biol. 2014, 30, 45–54. [Google Scholar] [CrossRef]
- Dunleavy, J.E.M.; O’Bryan, M.K. Regulation of microtubules in the seminiferous epithelium. Microtubule Dyn. Methods Protoc. 2019, 1926, 119–134. [Google Scholar]
- Rudnik-Schöneborn, S.; Messner, M.; Vockel, M.; Wirleitner, B.; Pinggera, G.-M.; Witsch-Baumgartner, M.; Murtinger, M.; Kliesch, S.; Swoboda, M.; Sänger, N.; et al. Andrological findings in infertile men with two (biallelic) CFTR mutations: Results of a multicentre study in Germany and Austria comprising 71 patients. Hum. Reprod. 2021, 36, 551–559. [Google Scholar] [CrossRef]
- Lehti, M.S.; Kotaja, N.; Sironen, A. KIF3A is essential for sperm tail formation and manchette function. Mol. Cell. Endocrinol. 2013, 377, 44–55. [Google Scholar] [CrossRef]
- Sweeney, H.L.; Holzbaur, E.L. Motor Proteins. Cold Spring Harb. Perspect. Biol. 2018, 10, a021931. [Google Scholar] [CrossRef]
- Miki, H.; Setou, M.; Kaneshiro, K.; Hirokawa, N. All kinesin superfamily protein, KIF, genes in mouse and human. Proc. Natl. Acad. Sci. USA 2001, 98, 7004–7011. [Google Scholar] [CrossRef]
- Marx, A.; Hoenger, A.; Mandelkow, E. Structures of kinesin motor proteins. Cell Motil. Cytoskelet. 2009, 66, 958–966. [Google Scholar] [CrossRef]
- Hirokawa, N.; Tanaka, Y. Kinesin superfamily proteins (KIFs): Various functions and their relevance for important phenomena in life and diseases. Exp. Cell Res. 2015, 334, 16–25. [Google Scholar] [CrossRef]
- Hirokawa, N.; Noda, Y.; Tanaka, Y.; Niwa, S. Kinesin superfamily motor proteins and intracellular transport. Nat. Rev. Mol. Cell Biol. 2009, 10, 682–696. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.F.; Lefebvre, P.A. The role of central apparatus components in flagellar motility and microtubule assembly. Cell Motil. Cytoskelet. 1997, 38, 1–8. [Google Scholar] [CrossRef]
- Morohoshi, A.; Miyata, H.; Shimada, K.; Nozawa, K.; Matsumura, T.; Yanase, R.; Shiba, K.; Inaba, K.; Ikawa, M. Nexin-Dynein regulatory complex component DRC7 but not FBXL13 is required for sperm flagellum formation and male fertility in mice. PLoS Genet. 2020, 16, e1008585. [Google Scholar] [CrossRef] [PubMed]
- Kubo, S.; Black, C.S.; Joachimiak, E.; Yang, S.K.; Legal, T.; Peri, K.; Khalifa, A.A.Z.; Ghanaeian, A.; McCafferty, C.L.; Valente-Paterno, M.; et al. Native doublet microtubules from Tetrahymena thermophila reveal the importance of outer junction proteins. Nat. Commun. 2023, 14, 2168. [Google Scholar] [CrossRef] [PubMed]
- Singharajkomron, N.; Yodsurang, V.; Seephan, S.; Kungsukool, S.; Petchjorm, S.; Maneeganjanasing, N.; Promboon, W.; Dangwilailuck, W.; Pongrakhananon, V. Evaluating the Expression and Prognostic Value of Genes Encoding Microtubule-Associated Proteins in Lung Cancer. Int. J. Mol. Sci. 2022, 23, 14724. [Google Scholar] [CrossRef] [PubMed]
- Holloway, K.; Roberson, E.C.; Corbett, K.L.; Kolas, N.K.; Nieves, E.; Cohen, P.E. NEK1 Facilitates Cohesin Removal during Mammalian Spermatogenesis. Genes 2011, 2, 260–279. [Google Scholar] [CrossRef]
- Ghédir, H.; Braham, A.; Viville, S.; Saad, A.; Ibala-Romdhane, S. Comparison of sperm morphology and nuclear sperm quality in SPATA16- and DPY19L2-mutated globozoospermic patients. Andrologia 2019, 51, e13277. [Google Scholar] [CrossRef]
- Xu, X.; Toselli, P.A.; Russell, L.D.; Seldin, D.C. Globozoospermia in mice lacking the casein kinase II alpha’ catalytic subunit. Nat. Genet. 1999, 23, 118–121. [Google Scholar] [CrossRef]
- ElInati, E.; Fossard, C.; Okutman, O.; Ghédir, H.; Ibala-Romdhane, S.; Ray, P.F.; Saad, A.; Hennebicq, S.; Viville, S. A new mutation identified in SPATA16 in two globozoospermic patients. J. Assist. Reprod. Genet. 2016, 33, 815–820. [Google Scholar] [CrossRef]
- Yu, J.; Chen, Z.; Ni, Y.; Li, Z. CFTR mutations in men with congenital bilateral absence of the vas deferens (CBAVD): A systemic review and meta-analysis. Hum. Reprod. 2012, 27, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Bieniek, J.M.; Lapin, C.D.; Jarvi, K.A. Genetics of CFTR and male infertility. Transl. Androl. Urol. 2021, 10, 1391–1400. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.W.; Yin, H.Q.; Li, Q.; Zhao, Y.P.; Brandes, K.; Bai, W.J.; Xu, T. CMTM2 is involved in spermiogenesis in mice. Beijing Da Xue Xue Bao Yi Xue Ban. 2019, 51, 228–293. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Chromosome | Number of Amplicons | Total_Bp | Covered_Bp | Missed_Bp | Coverage |
|---|---|---|---|---|---|---|---|
| G1 | FSHB | chr11 | 3 | 410 | 410 | 0 | 1 |
| G2 | AURKC | chr19 | 9 | 1011 | 1011 | 0 | 1 |
| G3 | LHB | chr19 | 3 | 456 | 191 | 265 | 0.419 |
| G4 | PICK1 | chr22 | 17 | 1368 | 1368 | 0 | 1 |
| G5 | SPATA16 | chr3 | 16 | 1810 | 1810 | 0 | 1 |
| G6 | DAZL | chr3 | 15 | 1071 | 1071 | 0 | 1 |
| G7 | ESR1 | chr6 | 15 | 1874 | 1874 | 0 | 1 |
| G8 | CFTR | chr7 | 46 | 4713 | 4709 | 4 | 0.999 |
| G9 | KAL2 | chr8 | 28 | 2825 | 2825 | 0 | 1 |
| G10 | GNRH1 | chr8 | 4 | 321 | 321 | 0 | 1 |
| G11 | NR5A1 | chr9 | 14 | 1446 | 1424 | 22 | 0.985 |
| G12 | AR | chrx | 19 | 2873 | 2873 | 0 | 1 |
| G13 | TEX11 | chrx | 34 | 3160 | 3068 | 92 | 0.971 |
| G14 | USP26 | chrx | 15 | 2752 | 2752 | 0 | 1 |
| G15 | KAL1 (ANOS1) | chrx | 19 | 2183 | 2000 | 183 | 0.916 |
| Gene | Chr. | SNP ID | Allele Variation | Oligo | Azoo | Fertile Men | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S06 | S07 | S08 | S11 | S02 | S09 | S10 | S-2017 | N1 | CRT-1 | ||||
| SPATA16 | 3 | rs16846616 | T→C | ■ | ■ | ■ | ■ | ■ | |||||
| rs1515442 | C→T | ■ | ■ | ■ | ■ | ■ | |||||||
| rs1515441 | C→T | ■ | ■ | ■ | ■ | ■ | |||||||
| CFTR | 7 | rs213950 | G→A | ■ | ■ | ■ | ■ | ■ | ■ | ■ | ■ | ||
| ESR1 | 6 | rs17847065 | C→A | ■ | ■ | ■ | ■ | ■ | |||||
| TEX11 | X | rs6525433 | T→C | ■ | ■ | ||||||||
| rs4844247 | C→T | ■ | |||||||||||
| LHB | 19 | rs146251380 | G→A,T | ■ | |||||||||
| USP26 | X | rs61741870 | A→G | ■ | |||||||||
| rs41299088 | G→A | ■ | |||||||||||
| AR | X | rs777131133 | C→A,G,T | ■ | ■ | ||||||||
| GNRH1 | 8 | rs6185 | C→A,G,T | ■ | ■ | ■ | ■ | ■ | ■ | ||||
| NR5A1 | 9 | rs1110061 | C→A,G | ■ | ■ | ■ | ■ | ■ | ■ | ||||
| ANOS1 | X | rs808119 | C→A,T | ■ | ■ | ■ | ■ | ■ | |||||
| rs2229013 | C→A,T | ■ | |||||||||||
| Phenotype | Patient ID | CFTR | SPATA16 | ||
|---|---|---|---|---|---|
| rs213950 | rs16846616 | rs1515442 | rs1515441 | ||
| G→A | T→C | C→T | C→T | ||
| CRT-1 | |||||
| N1 | |||||
| N2 | |||||
| CRT, Fertile | N3 | T→C | G→A | ||
| (n = 9) | N4 | T→C | G→A | ||
| N5 | |||||
| N8 | |||||
| N27(NN7) | |||||
| N31(sample 1) | |||||
| S01 | |||||
| S03 | C→T | T→C | G→A | ||
| S04 | |||||
| S05 | |||||
| S06 | C→T | ||||
| S07 | C→T | ||||
| S08 | C→T | T→C | C→T | G→A | |
| Oligo, sperm counts | S11 | C→T | |||
| ≦15 million (n = 16) | S15 | ||||
| S18 | |||||
| S20 | |||||
| CA21 | |||||
| CA22 | C→T | T→C | G→A | ||
| CA24 | |||||
| CA25 | |||||
| S31 | |||||
| P1 (S-2017) | C→T | T→C | C→T | G→A | |
| P2 | T→C | C→T | G→A | ||
| S02 | C→T | T→C | C→T | G→A | |
| S09 | C→T | T→C | C→T | G→A | |
| S10 | C→T | T→C | C→T | G→A | |
| S13 | T→C | G→A | |||
| Azoo, sperm counts | S14 | ||||
| <0.1 million (n = 15) | S16 | T→C | G→A | ||
| S17 | C→T | ||||
| S26 | C→T | T→C | C→T | G→A | |
| S32 | C→T | T→C | C→T | G→A | |
| CA23 | T→C | G→A | |||
| CA26 | C→T | ||||
| TCA26 | C→T | T→C | C→T | G→A | |
| TS1 | C→T | T→C | C→T | G→A | |
| Phenotype | Patient | STPG2 | KIF6 |
|---|---|---|---|
| N1 | A/G | X | |
| N2 | X | G/A | |
| N3 | A/G | G/A | |
| N4 | X | X | |
| N5 | X | X | |
| N6 | A/G | X | |
| N7 | X | X | |
| N8 | X | X | |
| N9 | A/G | X | |
| Ordinary Men | N10 | A/G | G/A |
| n = 20 | N11 | X | X |
| N12 | X | G/A | |
| N13 | A/G | X | |
| N14 | X | X | |
| N15 | X | X | |
| N16 | X | X | |
| N17 | X | X | |
| N18 | A/G | X | |
| N19 | X | X | |
| N20 | A/G | X | |
| PN1 | X | X | |
| PN2 | X | G/A | |
| PN3 | X | X | |
| CRT Fertile | PN4 | X | X |
| n = 8 | PN5 | A/G | X |
| PN6 | X | X | |
| PN7 | A/G | X | |
| NN7 | X | X | |
| S01 | A/G | G/A | |
| S03 | A/G | G/A | |
| S04 | A/G | G/A | |
| S05 | X | X | |
| S06 | X | G/A | |
| S7 | A/G | G/A | |
| Oligozoospermia | S8 | A/G | A/A |
| n = 16 | S11 | X | G/A |
| S12 | A/G | X | |
| S15 | A/G | G/A | |
| S18 | X | G/A | |
| S20 | X | X | |
| CA22 | X | X | |
| CA24 | X | X | |
| CA25 | A/G | A/A | |
| S31 | X | X | |
| P1 | A/G | G/A | |
| P2 | A/G | G/A | |
| S02 | A/G | G/A | |
| S09 | A/G | G/A | |
| S10 | A/G | G/A | |
| S13 | A/G | G/A | |
| Azoospermia | S14 | A/G | G/A |
| n = 15 | S16 | A/G | G/A |
| S17 | X | X | |
| S26 | X | X | |
| S32 | A/G | G/A | |
| CA23 | A/G | X | |
| CA26 | A/G | G/A | |
| TCA26 | X | G/A | |
| TS1 | A/G | G/A |
| Group by | Patient | Volume | Sperm Counts | Age | Motility | Morphology |
|---|---|---|---|---|---|---|
| Phenotype | ID | (mL) | (M) | (%) | (%) | |
| CRT-1 | 3.2 | normal | 40 | 45 | 60 | |
| CRT Fertile (n = 9) | N1 | 2.1 | normal | 32 | 35 | 36 |
| N2 | nd | normal | 33 | nd | nd | |
| N3 | nd | normal | 35 | nd | nd | |
| N4 | nd | normal | 42 | nd | nd | |
| N5 | nd | normal | 38 | nd | nd | |
| N8 | nd | normal | 29 | nd | nd | |
| N27(NN7) | nd | normal | 40 | nd | nd | |
| N31 | nd | normal | 50 | 61 | 61 | |
| Oligo, sperm counts ≤ 15 million (n = 16) | S01 | 1.9 | 15 | 42 | 13 | 58 |
| S03 | 1.8 | 11.6 | 40 | 65 | 72 | |
| S04 | 1.9 | 11.9 | 36 | 47 | 52 | |
| S05 | 0.5 | 5.6 | 37 | 45 | 60 | |
| S06 | 3.4 | 2.1 | 32 | 90 | 30 | |
| S07 | 4.5 | 4.5 | 41 | 20 | 58 | |
| S08 | 4.1 | 1.4 | 45 | 21 | 40 | |
| S11 | 1.5 | 0.3 | 36 | 67 | 61 | |
| S15 | 0.7 | 3.6 | 38 | 38 | 55 | |
| S18 | 1.5 | 7.7 | 44 | 71 | 53 | |
| S20 | 2.6 | 4.2 | 34 | 52 | 45 | |
| CA21 | 3.2 | 4.4 | 32 | 52 | 33 | |
| CA22 | 3.5 | 7.1 | 36 | 81 | 53 | |
| CA24 | 1.6 | 7 | 39 | 54 | 45 | |
| CA25 | 1.2 | 6.5 | 41 | 57 | 51 | |
| S31 | 2.1 | 14 | 32 | 35 | 45 | |
| Azoo, sperm counts < 0.1 million (n = 15) | P1 (S-2017) | 3.5 | <0.1 | 40 | nd | nd |
| P2 | 4 | <0.1 | 38 | nd | nd | |
| S02 | 0.5 | <0.1 | 43 | 50 | 50 | |
| S09 | 1.3 | <0.1 | 32 | 50 | 50 | |
| S10 | 0.5 | <0.1 | 30 | 0 | 30 | |
| S13 | 0.9 | <0.1 | 42 | 0 | nd | |
| S14 | <0.1 | <0.1 | 39 | 0 | 0 | |
| S16 | 6.7 | <0.1 | 39 | 42 | 30 | |
| S17 | 0.3 | <0.1 | 35 | nd | nd | |
| S26 | 1.2 | <0.1 | 36 | 57 | 52 | |
| S32 | 1.5 | <0.1 | 43 | nd | nd | |
| CA23 | 0.4 | <0.1 | 39 | nd | nd | |
| CA26 | 0.9 | <0.1 | 36 | nd | nd | |
| TCA26 | 0.7 | <0.1 | 30 | nd | nd | |
| TS1 | 0.6 | <0.1 | 34 | nd | nd |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chan, C.-C.; Yen, T.-H.; Tseng, H.-C.; Mai, B.; Ho, P.-K.; Chou, J.-L.; Wu, G.-J.; Huang, Y.-C. A Comprehensive Genetic Study of Microtubule-Associated Gene Clusters for Male Infertility in a Taiwanese Cohort. Int. J. Mol. Sci. 2023, 24, 15363. https://doi.org/10.3390/ijms242015363
Chan C-C, Yen T-H, Tseng H-C, Mai B, Ho P-K, Chou J-L, Wu G-J, Huang Y-C. A Comprehensive Genetic Study of Microtubule-Associated Gene Clusters for Male Infertility in a Taiwanese Cohort. International Journal of Molecular Sciences. 2023; 24(20):15363. https://doi.org/10.3390/ijms242015363
Chicago/Turabian StyleChan, Chying-Chyuan, Te-Hsin Yen, Hao-Chen Tseng, Brang Mai, Pin-Kuan Ho, Jian-Liang Chou, Gwo-Jang Wu, and Yu-Chuan Huang. 2023. "A Comprehensive Genetic Study of Microtubule-Associated Gene Clusters for Male Infertility in a Taiwanese Cohort" International Journal of Molecular Sciences 24, no. 20: 15363. https://doi.org/10.3390/ijms242015363
APA StyleChan, C.-C., Yen, T.-H., Tseng, H.-C., Mai, B., Ho, P.-K., Chou, J.-L., Wu, G.-J., & Huang, Y.-C. (2023). A Comprehensive Genetic Study of Microtubule-Associated Gene Clusters for Male Infertility in a Taiwanese Cohort. International Journal of Molecular Sciences, 24(20), 15363. https://doi.org/10.3390/ijms242015363