
Phytoestrogen-Based Hormonal Replacement Therapy Could Benefit Women Suffering Late-Onset Asthma

 , ,
, ,
Abstract
1. Introduction
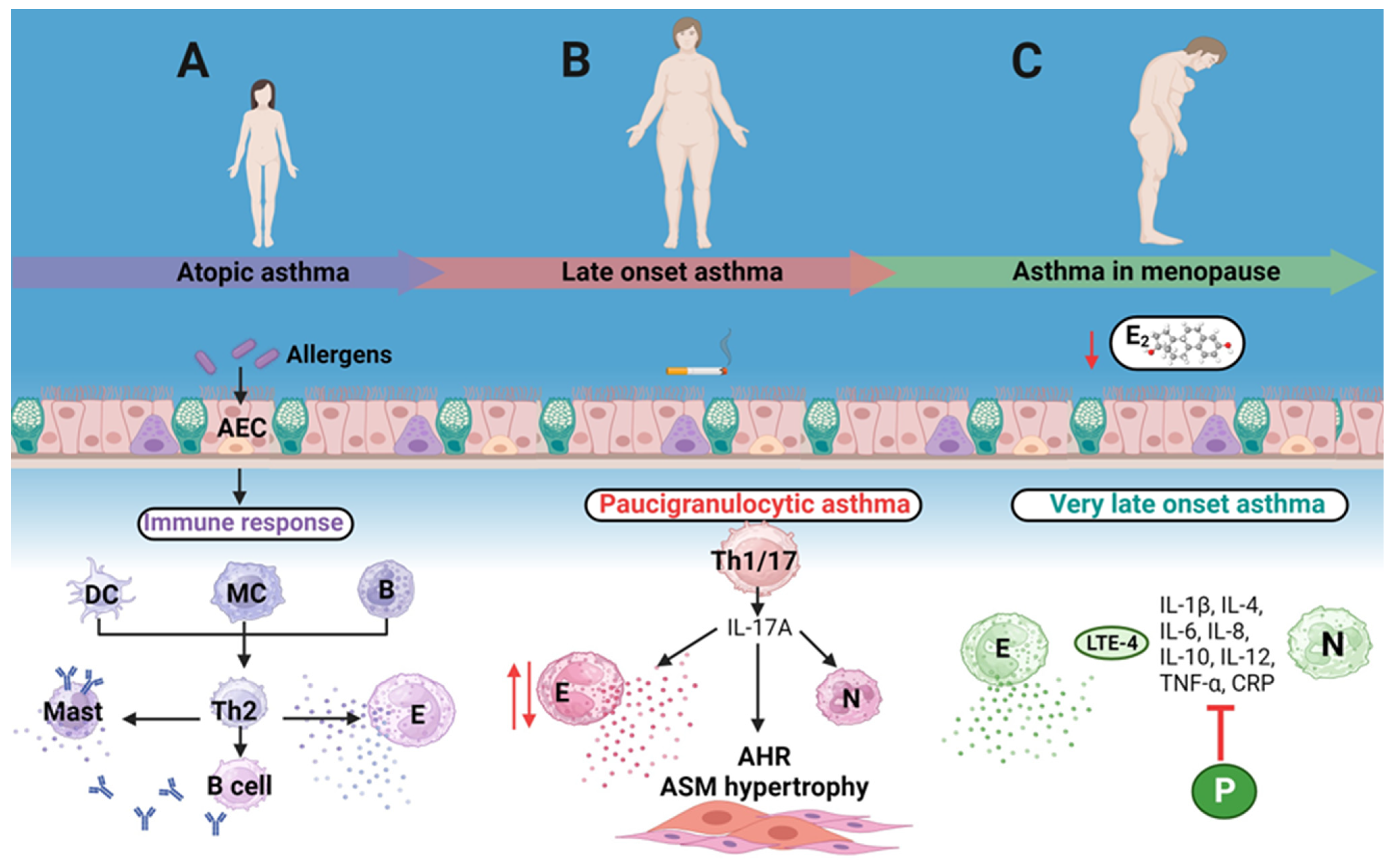
2. Female Humoral Immunity Plays a Key Role in Atopic Asthma
3. Late-Onset Asthma
4. Hormone Replacement Therapy Pros and Cons
5. Menopause Is an Inflammatory State
6. Characteristics of Phytoestrogens
7. Isoflavones and Their Actions in Hormone Replacement Therapy
8. Resveratrol Is an Agonist for the Estrogen Receptor
9. Phytoestrogens Mode of Action
10. Phytoestrogens as HRT in Women with LOA
11. Phytoestrogens Potential in Pulmonary Fibrosis Treatment
12. Final Considerations
13. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barnes, P.J. Cellular and molecular mechanisms of asthma and COPD. Clin. Sci. 2017, 31, 1541–1558. [Google Scholar] [CrossRef] [PubMed]
- Papi, A.; Brightling, C.; Pedersen, S.E.; Reddel, H.K. Asthma. Lancet 2018, 391, 783–800. [Google Scholar] [CrossRef]
- Padem, N.; Saltoun, C. Classification of asthma. Allergy Asthma Proc. 2019, 40, 385–388. [Google Scholar] [CrossRef]
- Han, M.K.; Arteaga-Solis, E.; Blenis, J.; Bourjeily, G.; Clegg, D.J.; Demeo, D.; Duffy, J.; Gaston, B.; Heller, N.M.; Hemnes, A.; et al. Female Sex and Gender in Lung/Sleep Health and Disease. Increased Understanding of Basic Biological, Pathophysiological, and Behavioral Mechanisms Leading to Better Health for Female Patients with Lung Disease. Am. J. Respir. Crit. Care Med. 2018, 198, 850–858. [Google Scholar] [CrossRef]
- Skobeloff, E.M. The Effect of the Menstrual Cycle on Asthma Presentations in the Emergency Department. Arch. Intern. Med. 1996, 156, 1837. [Google Scholar] [CrossRef]
- Vrieze, A.; Postma, D.S.; Kerstjens, H.A. Perimenstrual asthma: A syndrome without known cause or cure. J. Allergy Clin. Immunol. 2003, 112, 271–282. [Google Scholar] [CrossRef]
- Chhabra, S.K. Premenstrual asthma. Indian J. Chest Dis. Allied Sci. 2005, 47, 109–116. [Google Scholar]
- Graziottin, A.; Serafini, A. Perimenstrual asthma: From pathophysiology to treatment strategies. Multidiscip. Respir. Med. 2016, 11, 30. [Google Scholar] [CrossRef]
- Skoczyński, S.; Semik-Orzech, A.; Szanecki, W.; Majewski, M.; Kołodziejczyk, K.; Sozańska, E.; Witek, A.; Pierzchała, W. Perimenstrual Asthma as a Gynecological and Pulmonological Clinical Problem. Adv. Clin. Exp. Med. 2014, 23, 665–668. [Google Scholar] [CrossRef]
- Pereira-Vega, A.; Sánchez Ramos, J.L.; Vázquez Oliva, R.; Maldonado Pérez, J.A.; Olbah, M.A.; Montero Fernández, M.; Vázquez Rico, I.; Robles Rodríguez, J.L.; Ignacio García, J.M.; Romero Palacios, P.; et al. Premenstrual asthma and female sex hormones. J. Investig. Allergol. Clin. Immunol. 2012, 22, 437–439. [Google Scholar]
- Vrachnis, N.; Zygouris, D.; Iliodromiti, Z.; Daniilidis, A.; Valsamakis, G.; Kalantaridou, S. Probing the impact of sex steroids and menopause-related sex steroid deprivation on modulation of immune senescence. Maturitas 2014, 78, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Kovats, S. Estrogen receptors regulate innate immune cells and signaling pathways. Cell. Immunol. 2015, 294, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Giefing-Kröll, C.; Berger, P.; Lepperdinger, G.; Grubeck-Loebenstein, B. How sex and age affect immune responses, susceptibility to infections, and response to vaccination. Aging Cell 2015, 14, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Triebner, K.; Johannessen, A.; Puggini, L.; Benediktsdóttir, B.; Bertelsen, R.J.; Bifulco, E.; Dharmage, S.C.; Dratva, J.; Franklin, K.A.; Gíslason, T.; et al. Menopause as a predictor of new-onset asthma: A longitudinal Northern European population study. J. Allergy Clin. Immunol. 2016, 137, 50–57.e56. [Google Scholar] [CrossRef] [PubMed]
- Dunn, R.M.; Busse, P.J.; Wechsler, M.E. Asthma in the elderly and late-onset adult asthma. Allergy 2018, 73, 284–294. [Google Scholar] [CrossRef]
- Kaur, R.; Chupp, G. Phenotypes and endotypes of adult asthma: Moving toward precision medicine. J. Allergy Clin. Immunol. 2019, 144, 1–12. [Google Scholar] [CrossRef]
- Evaluation of the Efficacy of Respicure® (Resveratrol/Quercetin) in the Management of Respiratory Conditions Including Asthma, COPD and Long COVID. Available online: https://clinicaltrials.gov/study/NCT05601180 (accessed on 29 September 2023).
- Dietary Supplementation with Soy Isoflavones in Asthma. Available online: https://clinicaltrials.gov/study/NCT00277446 (accessed on 29 September 2023).
- The Study of Soy Isoflavones in Asthma. Available online: https://clinicaltrials.gov/study/NCT01052116 (accessed on 29 September 2023).
- Modification of Asthma with Soy Isoflavone. Available online: https://clinicaltrials.gov/study/NCT00741208 (accessed on 29 September 2023).
- Soy Isoflavones for Inner City Infants at Risk for Asthma (SIRA) Study. Available online: https://clinicaltrials.gov/study/NCT05667701 (accessed on 29 September 2023).
- Kanda, N.; Hoashi, T.; Saeki, H. The Roles of Sex Hormones in the Course of Atopic Dermatitis. Int. J. Mol. Sci. 2019, 20, 4660. [Google Scholar] [CrossRef]
- Ridolo, E.; Incorvaia, C.; Martignago, I.; Caminati, M.; Canonica, G.W.; Senna, G. Sex in Respiratory and Skin Allergies. Clin. Rev. Allergy Immunol. 2019, 56, 322–332. [Google Scholar] [CrossRef]
- González, D.A.; Díaz, B.B.; Rodríguez Pérez, M.d.C.; Hernández, A.G.; Chico, B.N.; de León, A.C. Sex hormones and autoimmunity. Immunol. Lett. 2010, 133, 6–13. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Vital signs: Asthma prevalence, disease characteristics, and self-management education: United States, 2001–2009. MMWR Morb. Mortal. Wkly. Rep. 2011, 60, 547–552. [Google Scholar]
- Moorman, J.E.; Zahran, H.; Truman, B.I.; Molla, M.T.; Centers for Disease Control and Prevention (CDC). Current asthma prevalence—United States, 2006–2008. MMWR Suppl. 2011, 60, 84–86. [Google Scholar] [PubMed]
- Siroux, V.; Curt, F.; Oryszczyn, M.-P.; Maccario, J.; Kauffmann, F. Role of gender and hormone-related events on IgE, atopy, and eosinophils in the Epidemiological Study on the Genetics and Environment of Asthma, bronchial hyperresponsiveness and atopy. J. Allergy Clin. Immunol. 2004, 114, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Yancey, S.W.; Keene, O.N.; Albers, F.C.; Ortega, H.; Bates, S.; Bleecker, E.R.; Pavord, I. Biomarkers for severe eosinophilic asthma. J. Allergy Clin. Immunol. 2017, 140, 1509–1518. [Google Scholar] [CrossRef]
- Hollams, E.M.; Deverell, M.; Serralha, M.; Suriyaarachchi, D.; Parsons, F.; Zhang, G.; De Klerk, N.; Holt, B.J.; Ladyman, C.; Sadowska, A.; et al. Elucidation of asthma phenotypes in atopic teenagers through parallel immunophenotypic and clinical profiling. J. Allergy Clin. Immunol. 2009, 124, 463–470.e416. [Google Scholar] [CrossRef]
- Mor, G.; Cardenas, I. The Immune System in Pregnancy: A Unique Complexity. Am. J. Reprod. Immunol. 2010, 63, 425–433. [Google Scholar] [CrossRef]
- Kovats, S. Estrogen receptors regulate an inflammatory pathway of dendritic cell differentiation: Mechanisms and implications for immunity. Horm. Behav. 2012, 62, 254–262. [Google Scholar] [CrossRef]
- Grimaldi, C.M.; Cleary, J.; Dagtas, A.S.; Moussai, D.; Diamond, B. Estrogen alters thresholds for B cell apoptosis and activation. J. Clin. Investig. 2002, 109, 1625–1633. [Google Scholar] [CrossRef] [PubMed]
- Anipindi, V.C.; Bagri, P.; Roth, K.; Dizzell, S.E.; Nguyen, P.V.; Shaler, C.R.; Chu, D.K.; Jiménez-Saiz, R.; Liang, H.; Swift, S.; et al. Estradiol Enhances CD4+ T-Cell Anti-Viral Immunity by Priming Vaginal DCs to Induce Th17 Responses via an IL-1-Dependent Pathway. PLoS Pathog. 2016, 12, e1005589. [Google Scholar] [CrossRef]
- Ku, L.T.; Gercel-Taylor, C.; Nakajima, S.T.; Taylor, D.D. Alterations of T cell activation signalling and cytokine production by postmenopausal estrogen levels. Immun. Ageing 2009, 6, 1. [Google Scholar] [CrossRef]
- Gilmore, W.; Weiner, L.P.; Correale, J. Effect of estradiol on cytokine secretion by proteolipid protein-specific T cell clones isolated from multiple sclerosis patients and normal control subjects. J. Immunol. 1997, 158, 446–451. [Google Scholar] [CrossRef]
- Cai, Y.; Zhou, J.; Webb, D.C. Estrogen stimulates Th2 cytokine production and regulates the compartmentalisation of eosinophils during allergen challenge in a mouse model of asthma. Int. Arch. Allergy Immunol. 2012, 158, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Lauzon-Joset, J.F.; Mincham, K.T.; Abad, A.P.; Short, B.P.; Holt, P.G.; Strickland, D.H.; Leffler, J. Oestrogen amplifies pre-existing atopy-associated Th2 bias in an experimental asthma model. Clin. Exp. Allergy 2020, 50, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Stein, M.M.; Hrusch, C.L.; Gozdz, J.; Igartua, C.; Pivniouk, V.; Murray, S.E.; Ledford, J.G.; Marques Dos Santos, M.; Anderson, R.L.; Metwali, N.; et al. Innate Immunity and Asthma Risk in Amish and Hutterite Farm Children. N. Engl. J. Med. 2016, 375, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Motika, C.A.; Papachristou, C.; Abney, M.; Lester, L.A.; Ober, C. Rising prevalence of asthma is sex-specific in a US farming population. J. Allergy Clin. Immunol. 2011, 128, 774–779. [Google Scholar] [CrossRef] [PubMed]
- Miranda, C.; Busacker, A.; Balzar, S.; Trudeau, J.; Wenzel, S.E. Distinguishing severe asthma phenotypes☆Role of age at onset and eosinophilic inflammation. J. Allergy Clin. Immunol. 2004, 113, 101–108. [Google Scholar] [CrossRef]
- Hirano, T.; Matsunaga, K. Late-onset asthma: Current perspectives. J. Asthma Allergy 2018, 11, 19–27. [Google Scholar] [CrossRef]
- Leynaert, B.; Sunyer, J.; Garcia-Esteban, R.; Svanes, C.; Jarvis, D.; Cerveri, I.; Dratva, J.; Gislason, T.; Heinrich, J.; Janson, C.; et al. Gender differences in prevalence, diagnosis and incidence of allergic and non-allergic asthma: A population-based cohort. Thorax 2012, 67, 625–631. [Google Scholar] [CrossRef]
- Eagan, T.M.; Brøgger, J.C.; Eide, G.E.; Bakke, P.S. The incidence of adult asthma: A review. Int. J. Tuberc. Lung Dis. 2005, 9, 603–612. [Google Scholar]
- Gibson, P.G.; McDonald, V.M.; Marks, G.B. Asthma in older adults. Lancet 2010, 376, 803–813. [Google Scholar] [CrossRef]
- Torén, K.; Palmqvist, M.; Löwhagen, O.; Balder, B.; Tunsäter, A. Self-reported asthma was biased in relation to disease severity while reported year of asthma onset was accurate. J. Clin. Epidemiol. 2006, 59, 90–93. [Google Scholar] [CrossRef]
- Moore, W.C.; Meyers, D.A.; Wenzel, S.E.; Teague, W.G.; Li, H.; Li, X.; D’Agostino, R.; Castro, M.; Curran-Everett, D.; Fitzpatrick, A.M.; et al. Identification of Asthma Phenotypes Using Cluster Analysis in the Severe Asthma Research Program. Am. J. Respir. Crit. Care Med. 2010, 181, 315–323. [Google Scholar] [CrossRef]
- Hsiao, H.P.; Lin, M.C.; Wu, C.C.; Wang, C.C.; Wang, T.N. Sex-Specific Asthma Phenotypes, Inflammatory Patterns, and Asthma Control in a Cluster Analysis. J. Allergy Clin. Immunol. 2019, 7, 556–567.e515. [Google Scholar] [CrossRef]
- Troisi, R.J.; Speizer, F.E.; Willett, W.C.; Trichopoulos, D.; Rosner, B. Menopause, postmenopausal estrogen preparations, and the risk of adult-onset asthma. A prospective cohort study. Am. J. Respir. Crit. Care Med. 1995, 152, 1183–1188. [Google Scholar] [CrossRef]
- Real, F.G.; Svanes, C.; Omenaas, E.R.; Antò, J.M.; Plana, E.; Jarvis, D.; Janson, C.; Neukirch, F.; Zemp, E.; Dratva, J.; et al. Lung function, respiratory symptoms, and the menopausal transition. J. Allergy Clin. Immunol. 2008, 121, 72–80.e73. [Google Scholar] [CrossRef]
- Jenkins, M.A.; Dharmage, S.C.; Flander, L.B.; Douglass, J.A.; Ugoni, A.M.; Carlin, J.B.; Sawyer, S.M.; Giles, G.G.; Hopper, J.L. Parity and decreased use of oral contraceptives as predictors of asthma in young women. Clin. Exp. Allergy 2006, 36, 609–613. [Google Scholar] [CrossRef] [PubMed]
- Nwaru, B.I.; Pillinger, R.; Tibble, H.; Shah, S.A.; Ryan, D.; Critchley, H.; Price, D.; Hawrylowicz, C.M.; Simpson, C.R.; Soyiri, I.N.; et al. Hormonal contraceptives and onset of asthma in reproductive-age women: Population-based cohort study. J. Allergy Clin. Immunol. 2020, 146, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Newcomb, D.C. Birth control medications decrease asthma incidence. J. Allergy Clin. Immunol. 2020, 146, 283–284. [Google Scholar] [CrossRef] [PubMed]
- Schindler, A.E. Non-contraceptive benefits of hormonal contraceptives. Minerva Ginecol. 2010, 62, 319–329. [Google Scholar] [CrossRef]
- Shah, S.A.; Tibble, H.; Pillinger, R.; Mclean, S.; Ryan, D.; Critchley, H.; Price, D.; Hawrylowicz, C.M.; Simpson, C.R.; Soyiri, I.N.; et al. Hormone replacement therapy and asthma onset in menopausal women: National cohort study. J. Allergy Clin. Immunol. 2021, 147, 1662–1670. [Google Scholar] [CrossRef]
- Abildgaard, J.; Tingstedt, J.; Zhao, Y.; Hartling, H.J.; Pedersen, A.T.; Lindegaard, B.; Dam Nielsen, S. Increased systemic inflammation and altered distribution of T-cell subsets in postmenopausal women. PLoS ONE 2020, 15, e0235174. [Google Scholar] [CrossRef]
- Mccarthy, M.; Raval, A.P. The peri-menopause in a woman’s life: A systemic inflammatory phase that enables later neurodegenerative disease. J. Neuroinflammation 2020, 17, 317. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, M.; Esposito, K.; Vietri, M.T.; Gazzerro, P.; D’Auria, A.; Ardovino, I.; Puca, G.A.; Molinari, A.M. Cytokine pattern in postmenopause. Maturitas 2002, 41, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Pfeilschifter, J.; Köditz, R.; Pfohl, M.; Schatz, H. Changes in proinflammatory cytokine activity after menopause. Endocr. Rev. 2002, 23, 90–119. [Google Scholar] [CrossRef] [PubMed]
- Carlson, C.L.; Cushman, M.; Enright, P.L.; Cauley, J.A.; Newman, A.B.; Group, C.H.S.R. Hormone replacement therapy is associated with higher FEV1 in elderly women. Am. J. Respir. Crit. Care Med. 2001, 163, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Cevrioglu, A.S.; Fidan, F.; Unlu, M.; Yilmazer, M.; Orman, A.; Fenkci, I.V.; Serteser, M. The effects of hormone therapy on pulmonary function tests in postmenopausal women. Maturitas 2004, 49, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Pata, O.; Atiş, S.; Utku Oz, A.; Yazici, G.; Tok, E.; Pata, C.; Kiliç, F.; Camdeviren, H.; Aban, M. The effects of hormone replacement therapy type on pulmonary functions in postmenopausal women. Maturitas 2003, 46, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Mueller, J.E.; Frye, C.; Brasche, S.; Heinrich, J. Association of hormone replacement therapy with bronchial hyper-responsiveness. Respir. Med. 2003, 97, 990–992. [Google Scholar] [CrossRef]
- Boardman, H.M.; Hartley, L.; Eisinga, A.; Main, C.; Roqué I Figuls, M.; Bonfill Cosp, X.; Gabriel Sanchez, R.; Knight, B. Hormone therapy for preventing cardiovascular disease in post-menopausal women. Cochrane Database Syst. Rev. 2015, 2015. [Google Scholar] [CrossRef]
- Gialeraki, A.; Valsami, S.; Pittaras, T.; Panayiotakopoulos, G.; Politou, M. Oral Contraceptives and HRT Risk of Thrombosis. Clin. Appl. Thromb. Hemost. 2018, 24, 217–225. [Google Scholar] [CrossRef]
- Johansson, T.; Fowler, P.; Ek, W.E.; Skalkidou, A.; Karlsson, T.; Johansson, Å. Oral Contraceptives, Hormone Replacement Therapy, and Stroke Risk. Stroke 2022, 53, 3107–3115. [Google Scholar] [CrossRef]
- Santen, R.J.; Heitjan, D.F.; Gompel, A.; Lumsden, M.A.; Pinkerton, J.V.; Davis, S.R.; Stuenkel, C.A. Underlying Breast Cancer Risk and Menopausal Hormone Therapy. J. Clin. Endocrinol. Metab. 2020, 105, dgaa073. [Google Scholar] [CrossRef] [PubMed]
- Mauvais-Jarvis, F. Estrogen and androgen receptors: Regulators of fuel homeostasis and emerging targets for diabetes and obesity. Trends Endocrinol. Metab. 2011, 22, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Mauvais-Jarvis, F. Sex differences in metabolic homeostasis, diabetes, and obesity. Biol. Sex Differ. 2015, 6, 14. [Google Scholar] [CrossRef] [PubMed]
- Barros, R.P.; Gustafsson, J.-Å. Estrogen Receptors and the Metabolic Network. Cell Metab. 2011, 14, 289–299. [Google Scholar] [CrossRef]
- Barros, R.P.A.; Machado, U.F.; Gustafsson, J.-Å. Estrogen receptors: New players in diabetes mellitus. Trends Mol. Med. 2006, 12, 425–431. [Google Scholar] [CrossRef]
- Meyer, M.R.; Clegg, D.J.; Prossnitz, E.R.; Barton, M. Obesity, insulin resistance and diabetes: Sex differences and role of oestrogen receptors. Acta Physiol. 2011, 203, 259–269. [Google Scholar] [CrossRef]
- Abu-Taha, M.; Rius, C.; Hermenegildo, C.; Noguera, I.; Cerda-Nicolas, J.M.; Issekutz, A.C.; Jose, P.J.; Cortijo, J.; Morcillo, E.J.; Sanz, M.J. Menopause and ovariectomy cause a low grade of systemic inflammation that may be prevented by chronic treatment with low doses of estrogen or losartan. J. Immunol. 2009, 183, 1393–1402. [Google Scholar] [CrossRef]
- Mauvais-Jarvis, F.; Clegg, D.J.; Hevener, A.L. The role of estrogens in control of energy balance and glucose homeostasis. Endocr. Rev. 2013, 34, 309–338. [Google Scholar] [CrossRef]
- Lee, C.G.; Carr, M.C.; Murdoch, S.J.; Mitchell, E.; Woods, N.F.; Wener, M.H.; Chandler, W.L.; Boyko, E.J.; Brunzell, J.D. Adipokines, inflammation, and visceral adiposity across the menopausal transition: A prospective study. J. Clin. Endocrinol. Metab. 2009, 94, 1104–1110. [Google Scholar] [CrossRef]
- Straub, R.H. The complex role of estrogens in inflammation. Endocr. Rev. 2007, 28, 521–574. [Google Scholar] [CrossRef]
- Kaunitz, A.M.; Manson, J.E. Management of Menopausal Symptoms. Obstet. Gynecol. 2015, 126, 859–876. [Google Scholar] [CrossRef] [PubMed]
- Nabulsi, A.A.; Folsom, A.R.; White, A.; Patsch, W.; Heiss, G.; Wu, K.K.; Szklo, M. Association of Hormone-Replacement Therapy with Various Cardiovascular Risk Factors in Postmenopausal Women. N. Engl. J. Med. 1993, 328, 1069–1075. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, R.; Teixeira, D.; Calhau, C. Estrogen Signaling in Metabolic Inflammation. Mediat. Inflamm. 2014, 2014, 615917. [Google Scholar] [CrossRef] [PubMed]
- Strehlow, K.; Rotter, S.; Wassmann, S.; Adam, O.; Grohé, C.; Laufs, K.; BöHm, M.; Nickenig, G. Modulation of Antioxidant Enzyme Expression and Function by Estrogen. Circ. Res. 2003, 93, 170–177. [Google Scholar] [CrossRef]
- Borrás, C.; Gambini, J.; López-Grueso, R.; Pallardó, F.V.; Viña, J. Direct antioxidant and protective effect of estradiol on isolated mitochondria. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2010, 1802, 205–211. [Google Scholar] [CrossRef]
- Nyenhuis, S.M.; Schwantes, E.A.; Evans, M.D.; Mathur, S.K. Airway neutrophil inflammatory phenotype in older subjects with asthma. J. Allergy Clin. Immunol. 2010, 125, 1163–1165. [Google Scholar] [CrossRef]
- Wood, L.G.; Baines, K.J.; Fu, J.; Scott, H.A.; Gibson, P.G. The neutrophilic inflammatory phenotype is associated with systemic inflammation in asthma. Chest 2012, 142, 86–93. [Google Scholar] [CrossRef]
- Busse, P.J.; Birmingham, J.M.; Calatroni, A.; Manzi, J.; Goryachokovsky, A.; Fontela, G.; Federman, A.D.; Wisnivesky, J.P. Effect of aging on sputum inflammation and asthma control. J. Allergy Clin. Immunol. 2017, 139, 1808–1818.e1806. [Google Scholar] [CrossRef]
- Bauer, B.A.; Reed, C.E.; Yunginger, J.W.; Wollan, P.C.; Silverstein, M.D. Incidence and outcomes of asthma in the elderly. A population-based study in Rochester, Minnesota. Chest 1997, 111, 303–310. [Google Scholar] [CrossRef]
- Kurzer, M.S.; Xu, X. Dietary phytoestrogens. Annu. Rev. Nutr. 1997, 17, 353–381. [Google Scholar] [CrossRef]
- Chang, E.C.; Charn, T.H.; Park, S.H.; Helferich, W.G.; Komm, B.; Katzenellenbogen, J.A.; Katzenellenbogen, B.S. Estrogen Receptors alpha and beta as determinants of gene expression: Influence of ligand, dose, and chromatin binding. Mol. Endocrinol. 2008, 22, 1032–1043. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [PubMed]
- Adlercreutz, H. Lignans and human health. Crit. Rev. Clin. Lab. Sci. 2007, 44, 483–525. [Google Scholar] [CrossRef]
- Zhu, Y.; Kawaguchi, K.; Kiyama, R. Differential and directional estrogenic signaling pathways induced by enterolignans and their precursors. PLoS ONE 2017, 12, e0171390. [Google Scholar] [CrossRef]
- Křížová, L.; Dadáková, K.; Kašparovská, J.; Kašparovský, T. Isoflavones. Molecules 2019, 24, 1076. [Google Scholar] [CrossRef]
- Dakora, F.D.; Phillips, D.A. Diverse functions of isoflavonoids in legumes transcend anti-microbial definitions of phytoalexins. Physiol. Mol. Plant Pathol. 1996, 49, 1–20. [Google Scholar] [CrossRef]
- Hasanah, Y.; Nisa, T.C.; Armidin, H.; Hanum, H.I. Isoflavone content of soybean [Glycine max (L). Merr.] cultivars with different nitrogen sources and growing season under dry land conditions. J. Agric. Environ. Int. Dev. 2015, 109, 5–17. [Google Scholar] [CrossRef]
- Adler, S.A.; Purup, S.; Hansen-Møller, J.; Thuen, E.; Steinshamn, H. Phytoestrogens and Their Metabolites in Bulk-Tank Milk: Effects of Farm Management and Season. PLoS ONE 2015, 10, e0127187. [Google Scholar] [CrossRef]
- Rizzo, G.; Baroni, L. Soy, Soy Foods and Their Role in Vegetarian Diets. Nutrients 2018, 10, 43. [Google Scholar] [CrossRef]
- Andres, S.; Hansen, U.; Niemann, B.; Palavinskas, R.; Lampen, A. Determination of the isoflavone composition and estrogenic activity of commercial dietary supplements based on soy or red clover. Food Funct. 2015, 6, 2017–2025. [Google Scholar] [CrossRef]
- Kanadys, W.; Barańska, A.; Błaszczuk, A.; Polz-Dacewicz, M.; Drop, B.; Kanecki, K.; Malm, M. Evaluation of Clinical Meaningfulness of Red Clover (Trifolium pratense L.) Extract to Relieve Hot Flushes and Menopausal Symptoms in Peri- and Post-Menopausal Women: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2021, 13, 1258. [Google Scholar] [CrossRef] [PubMed]
- Muthyala, R.S.; Ju, Y.H.; Sheng, S.; Williams, L.D.; Doerge, D.R.; Katzenellenbogen, B.S.; Helferich, W.G.; Katzenellenbogen, J.A. Equol, a natural estrogenic metabolite from soy isoflavones: Convenient preparation and resolution of R- and S-equols and their differing binding and biological activity through estrogen receptors alpha and beta. Bioorganic Med. Chem. 2004, 12, 1559–1567. [Google Scholar] [CrossRef] [PubMed]
- Turner, R.; Baron, T.; Wolffram, S.; Minihane, A.M.; Cassidy, A.; Rimbach, G.; Weinberg, P.D. Effect of circulating forms of soy isoflavones on the oxidation of low density lipoprotein. Free Radic. Res. 2004, 38, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Ueno, T.; Uchiyama, S. Identification of the specific intestinal bacteria capable of metabolising soy isoflavone to equol. Ann. Nutr. Metab. 2001, 45, 114. [Google Scholar] [CrossRef]
- Lu, L.J.; Anderson, K.E. Sex and long-term soy diets affect the metabolism and excretion of soy isoflavones in humans. Am. J. Clin. Nutr. 1998, 68, 1500S–1504S. [Google Scholar] [CrossRef]
- Setchell, K.D.; Brown, N.M.; Lydeking-Olsen, E. The clinical importance of the metabolite equol—A clue to the effectiveness of soy and its isoflavones. J. Nutr. 2002, 132, 3577–3584. [Google Scholar] [CrossRef]
- Atkinson, C.; Frankenfeld, C.L.; Lampe, J.W. Gut bacterial metabolism of the soy isoflavone daidzein: Exploring the relevance to human health. Exp. Biol. Med. 2005, 230, 155–170. [Google Scholar] [CrossRef]
- Shutt, D.A.; Cox, R.I. Steroid and phyto-oestrogen binding to sheep uterine receptors in vitro. J. Endocrinol. 1972, 52, 299–310. [Google Scholar] [CrossRef]
- Leonard, L.M.; Choi, M.S.; Cross, T.-W.L. Maximizing the Estrogenic Potential of Soy Isoflavones through the Gut Microbiome: Implication for Cardiometabolic Health in Postmenopausal Women. Nutrients 2022, 14, 553. [Google Scholar] [CrossRef]
- Lethaby, A.; Marjoribanks, J.; Kronenberg, F.; Roberts, H.; Eden, J.; Brown, J. Phytoestrogens for menopausal vasomotor symptoms. Cochrane Database Syst. Rev. 2013. [Google Scholar] [CrossRef]
- Krebs, E.E.; Ensrud, K.E.; MacDonald, R.; Wilt, T.J. Phytoestrogens for treatment of menopausal symptoms: A systematic review. Obstet. Gynecol. 2004, 104, 824–836. [Google Scholar] [CrossRef] [PubMed]
- Howes, L.G.; Howes, J.B.; Knight, D.C. Isoflavone therapy for menopausal flushes: A systematic review and meta-analysis. Maturitas 2006, 55, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Chandrareddy, A.; Muneyyirci-Delale, O.; McFarlane, S.I.; Murad, O.M. Adverse effects of phytoestrogens on reproductive health: A report of three cases. Complement. Ther. Clin. Pract. 2008, 14, 132–135. [Google Scholar] [CrossRef] [PubMed]
- Gehm, B.D.; Mcandrews, J.M.; Chien, P.-Y.; Jameson, J.L. Resveratrol, a polyphenolic compound found in grapes and wine, is an agonist for the estrogen receptor. Proc. Natl. Acad. Sci. USA 1997, 94, 14138–14143. [Google Scholar] [CrossRef] [PubMed]
- Siemann, H.; Creasy, L.L. Concentration of the phytoalexin resveratrol in wine. Am. J. Enol. Vitic. 1992, 43, 49–52. [Google Scholar] [CrossRef]
- Hrelia, S.; Di Renzo, L.; Bavaresco, L.; Bernardi, E.; Malaguti, M.; Giacosa, A. Moderate Wine Consumption and Health: A Narrative Review. Nutrients 2022, 15, 175. [Google Scholar] [CrossRef]
- Martínez-Ortega, M.V.; Carcía-Parrilla, M.C.; Troncoso, A.M. Resveratrol content in wines and musts from the south of Spain. Nahr. Food 2000, 44, 253–256. [Google Scholar] [CrossRef]
- Muñoz-Bernal, Ó.A.; Coria-Oliveros, A.J.; de la Rosa, L.A.; Rodrigo-García, J.; Del Rocío Martínez-Ruiz, N.; Sayago-Ayerdi, S.G.; Alvarez-Parrilla, E. Cardioprotective effect of red wine and grape pomace. Food Res. Int. 2021, 140, 110069. [Google Scholar] [CrossRef]
- Castaldo, L.; Narváez, A.; Izzo, L.; Graziani, G.; Gaspari, A.; Di Minno, G.; Ritieni, A. Red Wine Consumption and Cardiovascular Health. Molecules 2019, 24, 3626. [Google Scholar] [CrossRef]
- Haseeb, S.; Alexander, B.; Baranchuk, A. Wine and Cardiovascular Health. Circulation 2017, 136, 1434–1448. [Google Scholar] [CrossRef]
- Donnelly, L.E.; Newton, R.; Kennedy, G.E.; Fenwick, P.S.; Leung, R.H.; Ito, K.; Russell, R.E.; Barnes, P.J. Anti-inflammatory effects of resveratrol in lung epithelial cells: Molecular mechanisms. Am. J. Physiol. Lung Cell. Mol. Physiol. 2004, 287, L774–L783. [Google Scholar] [CrossRef] [PubMed]
- Meng, T.; Xiao, D.; Muhammed, A.; Deng, J.; Chen, L.; He, J. Anti-Inflammatory Action and Mechanisms of Resveratrol. Molecules 2021, 26, 229. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Li, X.; Li, N.; Liu, T.; Liu, J.; Li, Z.; Xiao, H.; Li, J. Long-term resveratrol treatment prevents ovariectomy-induced osteopenia in rats without hyperplastic effects on the uterus. Br. J. Nutr. 2014, 111, 836–846. [Google Scholar] [CrossRef]
- Wong, R.H.; Thaung Zaw, J.J.; Xian, C.J.; Howe, P.R. Regular Supplementation With Resveratrol Improves Bone Mineral Density in Postmenopausal Women: A Randomized, Placebo-Controlled Trial. J. Bone Miner. Res. 2020, 35, 2121–2131. [Google Scholar] [CrossRef]
- Qasem, R.J. The estrogenic activity of resveratrol: A comprehensive review of in vitro and in vivo evidence and the potential for endocrine disruption. Crit. Rev. Toxicol. 2020, 50, 439–462. [Google Scholar] [CrossRef]
- Figtree, G.A.; Noonan, J.E.; Bhindi, R.; Collins, P. Estrogen receptor polymorphisms: Significance to human physiology, disease and therapy. Recent Pat. DNA Gene Seq. 2009, 3, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Hilakivi-Clarke, L.; de Assis, S. Fetal origins of breast cancer. Trends Endocrinol. Metab. 2006, 17, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Man Gho, W.; Chan, F.L.; Chen, S.; Leung, L.K. The red clover (Trifolium pratense) isoflavone biochanin A inhibits aromatase activity and expression. Br. J. Nutr. 2008, 99, 303–310. [Google Scholar] [CrossRef]
- Vitale, D.C.; Piazza, C.; Melilli, B.; Drago, F.; Salomone, S. Isoflavones: Estrogenic activity, biological effect and bioavailability. Eur. J. Drug Metab. Pharmacokinet. 2013, 38, 15–25. [Google Scholar] [CrossRef]
- Messina, M.; Kucuk, O.; Lampe, J.W. An overview of the health effects of isoflavones with an emphasis on prostate cancer risk and prostate-specific antigen levels. J. AOAC Int. 2006, 89, 1121–1134. [Google Scholar] [CrossRef]
- Evers, N.M.; van de Klundert, T.M.; van Aesch, Y.M.; Wang, S.; de Roos, W.K.; Romano, A.; de Haan, L.H.; Murk, A.J.; Ederveen, A.G.; Rietjens, I.M.; et al. Human T47D-ERβ breast cancer cells with tetracycline-dependent ERβ expression reflect ERα/ERβ ratios in rat and human breast tissue. Toxicol. In Vitro 2013, 27, 1753–1761. [Google Scholar] [CrossRef] [PubMed]
- Aravamudan, B.; Goorhouse, K.J.; Unnikrishnan, G.; Thompson, M.A.; Pabelick, C.M.; Hawse, J.R.; Prakash, Y.S.; Sathish, V. Differential Expression of Estrogen Receptor Variants in Response to Inflammation Signals in Human Airway Smooth Muscle. J. Cell. Physiol. 2017, 232, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Ambhore, N.S.; Katragadda, R.; Raju Kalidhindi, R.S.; Thompson, M.A.; Pabelick, C.M.; Prakash, Y.S.; Sathish, V. Estrogen receptor beta signaling inhibits PDGF induced human airway smooth muscle proliferation. Mol. Cell. Endocrinol. 2018, 476, 37–47. [Google Scholar] [CrossRef]
- Ambhore, N.S.; Kalidhindi, R.S.R.; Loganathan, J.; Sathish, V. Role of Differential Estrogen Receptor Activation in Airway Hyperreactivity and Remodeling in a Murine Model of Asthma. Am. J. Respir. Cell Mol. Biol. 2019, 61, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Kalidhindi, R.S.R.; Ambhore, N.S.; Bhallamudi, S.; Loganathan, J.; Sathish, V. Role of Estrogen Receptors α and β in a Murine Model of Asthma: Exacerbated Airway Hyperresponsiveness and Remodeling in ERβ Knockout Mice. Front. Pharmacol. 2020, 10, 1499. [Google Scholar] [CrossRef] [PubMed]
- Prossnitz, E.R.; Arterburn, J.B.; Smith, H.O.; Oprea, T.I.; Sklar, L.A.; Hathaway, H.J. Estrogen signaling through the transmembrane G protein-coupled receptor GPR30. Annu. Rev. Physiol. 2008, 70, 165–190. [Google Scholar] [CrossRef] [PubMed]
- Levin, E.R. Extranuclear Steroid Receptors Are Essential for Steroid Hormone Actions. Annu. Rev. Med. 2015, 66, 271–280. [Google Scholar] [CrossRef]
- Owman, C.; Blay, P.; Nilsson, C.; Lolait, S.J. Cloning of human cDNA encoding a novel heptahelix receptor expressed in Burkitt’s lymphoma and widely distributed in brain and peripheral tissues. Biochem. Biophys. Res. Commun. 1996, 228, 285–292. [Google Scholar] [CrossRef]
- Pandey, D.P.; Lappano, R.; Albanito, L.; Madeo, A.; Maggiolini, M.; Picard, D. Estrogenic GPR30 signalling induces proliferation and migration of breast cancer cells through CTGF. EMBO J. 2009, 28, 523–532. [Google Scholar] [CrossRef]
- Prossnitz, E.R.; Barton, M. Estrogen biology: New insights into GPER function and clinical opportunities. Mol. Cell. Endocrinol. 2014, 389, 71–83. [Google Scholar] [CrossRef]
- Vivacqua, A.; De Marco, P.; Santolla, M.F.; Cirillo, F.; Pellegrino, M.; Panno, M.L.; Abonante, S.; Maggiolini, M. Estrogenic gper signaling regulates mir144 expression in cancer cells and cancer-associated fibroblasts (cafs). Oncotarget 2015, 6, 16573–16587. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.; Pang, Y.; Filardo, E.J.; Dong, J. Identity of an estrogen membrane receptor coupled to a G protein in human breast cancer cells. Endocrinology 2005, 146, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Filardo, E.J.; Quinn, J.A.; Frackelton, A.R., Jr.; Bland, K.I. Estrogen action via the G protein-coupled receptor, GPR30: Stimulation of adenylyl cyclase and cAMP-mediated attenuation of the epidermal growth factor receptor-to-MAPK signaling axis. Mol. Endocrinol. 2002, 16, 70–84. [Google Scholar] [CrossRef]
- Revankar, C.M.; Cimino, D.F.; Sklar, L.A.; Arterburn, J.B.; Prossnitz, E.R. A Transmembrane Intracellular Estrogen Receptor Mediates Rapid Cell Signaling. Science 2005, 307, 1625–1630. [Google Scholar] [CrossRef] [PubMed]
- Lucas, T.F.; Royer, C.; Siu, E.R.; Lazari, M.F.; Porto, C.S. Expression and signaling of G protein-coupled estrogen receptor 1 (GPER) in rat sertoli cells. Biol. Reprod. 2010, 83, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Filardo, E.J.; Quinn, J.A.; Bland, K.I.; Frackelton, A.R., Jr. Estrogen-induced activation of Erk-1 and Erk-2 requires the G protein-coupled receptor homolog, GPR30, and occurs via trans-activation of the epidermal growth factor receptor through release of HB-EGF. Mol. Endocrinol. 2000, 14, 1649–1660. [Google Scholar] [CrossRef]
- Deng, Q.; Jiang, G.; Wu, Y.; Li, J.; Liang, W.; Chen, L.; Su, Q.; Li, W.; Du, J.; Wong, C.K.C.; et al. GPER/Hippo-YAP signal is involved in Bisphenol S induced migration of triple negative breast cancer (TNBC) cells. J. Hazard. Mater. 2018, 355, 1–9. [Google Scholar] [CrossRef]
- Thomas, P.; Dong, J. Binding and activation of the seven-transmembrane estrogen receptor GPR30 by environmental estrogens: A potential novel mechanism of endocrine disruption. J. Steroid. Biochem. Mol. Biol. 2006, 102, 175–179. [Google Scholar] [CrossRef]
- Zhou, C.; Li, P.; Han, M.; Gao, X. Daidzein stimulates fatty acid-induced fat deposition in C2C12 myoblast cells via the G protein-coupled receptor 30 pathway. Anim. Biotechnol. 2022, 33, 851–863. [Google Scholar] [CrossRef]
- Moriyama, M.; Hashimoto, A.; Satoh, H.; Kawabe, K.; Ogawa, M.; Takano, K.; Nakamura, Y. S-Equol, a Major Isoflavone from Soybean, Inhibits Nitric Oxide Production in Lipopolysaccharide-Stimulated Rat Astrocytes Partially via the GPR30-Mediated Pathway. Int. J. Inflamm. 2018, 2018, 8496973. [Google Scholar] [CrossRef]
- Moore, D.D. A Conversation with Elwood Jensen. Annu. Rev. Physiol. 2012, 74, 1–11. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bologa, C.G.; Revankar, C.M.; Young, S.M.; Edwards, B.S.; Arterburn, J.B.; Kiselyov, A.S.; Parker, M.A.; Tkachenko, S.E.; Savchuck, N.P.; Sklar, L.A.; et al. Virtual and biomolecular screening converge on a selective agonist for GPR30. Nat. Chem. Biol. 2006, 2, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Natale, C.A.; Li, J.; Pitarresi, J.R.; Norgard, R.J.; Dentchev, T.; Capell, B.C.; Seykora, J.T.; Stanger, B.Z.; Ridky, T.W. Pharmacologic Activation of the G Protein–Coupled Estrogen Receptor Inhibits Pancreatic Ductal Adenocarcinoma. Cell. Mol. Gastroenterol. Hepatol. 2020, 10, 868–880.e861. [Google Scholar] [CrossRef] [PubMed]
- Muller, C.; Brown-Glaberman, U.A.; Chaney, M.F.; Garyantes, T.; LoRusso, P.; McQuade, J.L.; Mita, A.C.; Mita, M.M.; Natale, C.; Orloff, M.; et al. Phase 1 trial of a novel, first-in-class G protein-coupled estrogen receptor (GPER) agonist, LNS8801 in patients with advanced or recurrent treatment-refractory solid malignancies. J. Clin. Oncol. 2021, 39, 3084. [Google Scholar] [CrossRef]
- Muller, C.; Chaney, M.F.; Cohen, J.V.; Garyantes, T.; Lin, J.J.; Lorusso, P.; Mita, A.C.; Mita, M.M.; Natale, C.; Orloff, M.M.; et al. Phase 1b study of the novel first-in-class G protein-coupled estrogen receptor (GPER) agonist, LNS8801, in combination with pembrolizumab in patients with immune checkpoint inhibitor (ICI)-relapsed and refractory solid malignancies and dose escalation upd. J. Clin. Oncol. 2022, 40 (Suppl. S16), 2574. [Google Scholar] [CrossRef]
- Tamaki, M.; Konno, Y.; Kobayashi, Y.; Takeda, M.; Itoga, M.; Moritoki, Y.; Oyamada, H.; Kayaba, H.; Chihara, J.; Ueki, S. Expression and functional roles of G-protein-coupled estrogen receptor (GPER) in human eosinophils. Immunol. Lett. 2014, 160, 72–78. [Google Scholar] [CrossRef]
- Itoga, M.; Konno, Y.; Moritoki, Y.; Saito, Y.; Ito, W.; Tamaki, M.; Kobayashi, Y.; Kayaba, H.; Kikuchi, Y.; Chihara, J.; et al. G-Protein-Coupled Estrogen Receptor Agonist Suppresses Airway Inflammation in a Mouse Model of Asthma through IL-10. PLoS ONE 2015, 10, e0123210. [Google Scholar] [CrossRef]
- Brunsing, R.L.; Owens, K.S.; Prossnitz, E.R. The G Protein-coupled Estrogen Receptor (GPER) Agonist G-1 Expands the Regulatory T-cell Population Under TH17-polarizing Conditions. J. Immunother. 2013, 36, 190–196. [Google Scholar] [CrossRef]
- Brunsing, R.L.; Prossnitz, E.R. Induction of interleukin-10 in the T helper type 17 effector population by the G protein coupled estrogen receptor (GPER) agonist G-1. Immunology 2011, 134, 93–106. [Google Scholar] [CrossRef]
- Sharma, G.; Hu, C.; Staquicini, D.I.; Brigman, J.L.; Liu, M.; Mauvais-Jarvis, F.; Pasqualini, R.; Arap, W.; Arterburn, J.B.; Hathaway, H.J.; et al. Preclinical efficacy of the GPER-selective agonist G-1 in mouse models of obesity and diabetes. Sci. Transl. Med. 2020, 12, eaau5956. [Google Scholar] [CrossRef]
- Sharma, G.; Mauvais-Jarvis, F.; Prossnitz, E.R. Roles of G protein-coupled estrogen receptor GPER in metabolic regulation. J. Steroid Biochem. Mol. Biol. 2018, 176, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Kim, S.R.; Kim, J.O.; Lee, Y.C. The Roles of Phytochemicals in Bronchial Asthma. Molecules 2010, 15, 6810–6834. [Google Scholar] [CrossRef] [PubMed]
- Grudzien, M.; Rapak, A. Effect of Natural Compounds on NK Cell Activation. J. Immunol. Res. 2018, 2018, 4868417. [Google Scholar] [CrossRef] [PubMed]
- Tasneem, S.; Liu, B.; Li, B.; Choudhary, M.I.; Wang, W. Molecular pharmacology of inflammation: Medicinal plants as anti-inflammatory agents. Pharmacol. Res. 2019, 139, 126–140. [Google Scholar] [CrossRef] [PubMed]
- Mollazadeh, H.; Cicero, A.F.G.; Blesso, C.N.; Pirro, M.; Majeed, M.; Sahebkar, A. Immune modulation by curcumin: The role of interleukin-10. Crit. Rev. Food Sci. Nutr. 2019, 59, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Wohlmuth, H.; Leach, D.N.; Smith, M.K.; Myers, S.P. Gingerol content of diploid and tetraploid clones of ginger (Zingiber officinale Roscoe). J. Agric. Food Chem. 2005, 53, 5772–5778. [Google Scholar] [CrossRef]
- Luković, E.; Perez-Zoghbi, J.F.; Zhang, Y.; Zhu, Y.; Sang, S.; Emala, C.W. Ginger metabolites and metabolite-inspired synthetic products modulate intracellular calcium and relax airway smooth muscle. Am. J. Physiol. Lung Cell. Mol. Physiol. 2021, 321, L912–L924. [Google Scholar] [CrossRef]
- Townsend, E.A.; Zhang, Y.; Xu, C.; Wakita, R.; Emala, C.W. Active Components of Ginger Potentiate β-Agonist–Induced Relaxation of Airway Smooth Muscle by Modulating Cytoskeletal Regulatory Proteins. Am. J. Respir. Cell Mol. Biol. 2014, 50, 115–124. [Google Scholar] [CrossRef]
- Rachoń, D.; Rimoldi, G.; Wuttke, W. In vitro effects of genistein and resveratrol on the production of interferon-gamma (IFNgamma) and interleukin-10 (IL-10) by stimulated murine splenocytes. Phytomedicine 2006, 13, 419–424. [Google Scholar] [CrossRef]
- Campbell, J.D.; Buckland, K.F.; Mcmillan, S.J.; Kearley, J.; Oldfield, W.L.G.; Stern, L.J.; Grönlund, H.; Van Hage, M.; Reynolds, C.J.; Boyton, R.J.; et al. Peptide immunotherapy in allergic asthma generates IL-10–dependent immunological tolerance associated with linked epitope suppression. J. Exp. Med. 2009, 206, 1535–1547. [Google Scholar] [CrossRef]
- Meiler, F.; Zumkehr, J.; Klunker, S.; RüCkert, B.; Akdis, C.A.; Akdis, M.B. In vivo switch to IL-10–secreting T regulatory cells in high dose allergen exposure. J. Exp. Med. 2008, 205, 2887–2898. [Google Scholar] [CrossRef]
- Golebski, K.; Layhadi, J.A.; Sahiner, U.; Steveling-Klein, E.H.; Lenormand, M.M.; Li, R.C.Y.; Bal, S.M.; Heesters, B.A.; Vilà-Nadal, G.; Hunewald, O.; et al. Induction of IL-10-producing type 2 innate lymphoid cells by allergen immunotherapy is associated with clinical response. Immunity 2021, 54, 291–307.e297. [Google Scholar] [CrossRef] [PubMed]
- Kearley, J.; Barker, J.E.; Robinson, D.S.; Lloyd, C.M. Resolution of airway inflammation and hyperreactivity after in vivo transfer of CD4+CD25+ regulatory T cells is interleukin 10 dependent. J. Exp. Med. 2005, 202, 1539–1547. [Google Scholar] [CrossRef] [PubMed]
- Schülke, S. Induction of Interleukin-10 Producing Dendritic Cells As a Tool to Suppress Allergen-Specific T Helper 2 Responses. Front. Immunol. 2018, 9, 455. [Google Scholar] [CrossRef] [PubMed]
- Hawrylowicz, C.; Richards, D.; Loke, T.K.; Corrigan, C.; Lee, T. A defect in corticosteroid-induced IL-10 production in T lymphocytes from corticosteroid-resistant asthmatic patients. J. Allergy Clin. Immunol. 2002, 109, 369–370. [Google Scholar] [CrossRef]
- Gupta, A.; Dimeloe, S.; Richards, D.F.; Chambers, E.S.; Black, C.; Urry, Z.; Ryanna, K.; Xystrakis, E.; Bush, A.; Saglani, S.; et al. Defective IL-10 expression and in vitro steroid-induced IL-17A in paediatric severe therapy-resistant asthma. Thorax 2014, 69, 508–515. [Google Scholar] [CrossRef]
- Chi, X.-X.; Zhang, T. The effects of soy isoflavone on bone density in north region of climacteric Chinese women. J. Clin. Biochem. Nutr. 2013, 53, 102–107. [Google Scholar] [CrossRef][Green Version]
- Nadadur, M.; Stanczyk, F.Z.; Tseng, C.C.; Kim, L.; Wu, A.H. The Effect of Reduced Dietary Fat and Soy Supplementation on Circulating Adipocytokines in Postmenopausal Women: A Randomized Controlled 2-Month Trial. Nutr. Cancer 2016, 68, 554–559. [Google Scholar] [CrossRef]
- Llaneza, P.; González, C.; Fernandez-Iñarrea, J.; Alonso, A.; Diaz, F.; Arnott, I.; Ferrer-Barriendos, J. Soy isoflavones, diet and physical exercise modify serum cytokines in healthy obese postmenopausal women. Phytomedicine 2011, 18, 245–250. [Google Scholar] [CrossRef]
- Podolanczuk, A.J.; Thomson, C.C.; Remy-Jardin, M.; Richeldi, L.; Martinez, F.J.; Kolb, M.; Raghu, G. Idiopathic pulmonary fibrosis: State of the art for 2023. Eur. Respir. J. 2023, 61, 2200957. [Google Scholar] [CrossRef]
- Andugulapati, S.B.; Gourishetti, K.; Tirunavalli, S.K.; Shaikh, T.B.; Sistla, R. Biochanin-A ameliorates pulmonary fibrosis by suppressing the TGF-β mediated EMT, myofibroblasts differentiation and collagen deposition in in vitro and in vivo systems. Phytomedicine 2020, 78, 153298. [Google Scholar] [CrossRef]
- Solopov, P.; Colunga Biancatelli, R.M.L.; Dimitropoulou, C.; Catravas, J.D. Dietary Phytoestrogens Ameliorate Hydrochloric Acid-Induced Chronic Lung Injury and Pulmonary Fibrosis in Mice. Nutrients 2021, 13, 3599. [Google Scholar] [CrossRef]
- Zhao, X.; Qu, G.; Song, C.; Li, R.; Liu, W.; Lv, C.; Song, X.; Zhang, J.; Li, M. Novel formononetin-7-sal ester ameliorates pulmonary fibrosis via MEF2c signaling pathway. Toxicol. Appl. Pharmacol. 2018, 356, 15–24. [Google Scholar] [CrossRef]
- White, J.; Dubey, S. Eosinophilic granulomatosis with polyangiitis: A review. Autoimmun. Rev. 2023, 22, 103219. [Google Scholar] [CrossRef]
- Nguyen, Y.; Guillevin, L. Eosinophilic Granulomatosis with Polyangiitis (Churg-Strauss). Semin. Respir. Crit. Care Med. 2018, 39, 471–481. [Google Scholar] [CrossRef]
- Jariwala, M.; Laxer, R.M. Childhood GPA, EGPA, and MPA. Clin. Immunol. 2020, 211, 108325. [Google Scholar] [CrossRef]
- Springer, J.M.; Kalot, M.A.; Husainat, N.M.; Byram, K.W.; Dua, A.B.; James, K.E.; Chang Lin, Y.; Turgunbaev, M.; Villa-Forte, A.; Abril, A.; et al. Eosinophilic Granulomatosis with Polyangiitis: A Systematic Review and Meta-Analysis of Test Accuracy and Benefits and Harms of Common Treatments. ACR Open Rheumatol. 2021, 3, 101–110. [Google Scholar] [CrossRef]
- Comarmond, C.; Pagnoux, C.; Khellaf, M.; Cordier, J.F.; Hamidou, M.; Viallard, J.F.; Maurier, F.; Jouneau, S.; Bienvenu, B.; Puéchal, X.; et al. Eosinophilic granulomatosis with polyangiitis (Churg-Strauss): Clinical characteristics and long-term followup of the 383 patients enrolled in the French Vasculitis Study Group cohort. Arthritis Rheum. 2013, 65, 270–281. [Google Scholar] [CrossRef]
- Moosig, F.; Bremer, J.P.; Hellmich, B.; Holle, J.U.; Holl-Ulrich, K.; Laudien, M.; Matthis, C.; Metzler, C.; Nölle, B.; Richardt, G.; et al. A vasculitis centre based management strategy leads to improved outcome in eosinophilic granulomatosis and polyangiitis (Churg-Strauss, EGPA): Monocentric experiences in 150 patients. Ann. Rheum. Dis. 2013, 72, 1011–1017. [Google Scholar] [CrossRef]
- Bottero, P.; Bonini, M.; Vecchio, F.; Grittini, A.; Patruno, G.M.; Colombo, B.; Sinico, R.A. The common allergens in the Churg-Strauss syndrome. Allergy 2007, 62, 1288–1294. [Google Scholar] [CrossRef]
- Amelink, M.; De Groot, J.C.; De Nijs, S.B.; Lutter, R.; Zwinderman, A.H.; Sterk, P.J.; Ten Brinke, A.; Bel, E.H. Severe adult-onset asthma: A distinct phenotype. J. Allergy Clin. Immunol. 2013, 132, 336–341. [Google Scholar] [CrossRef]
- Salles, G.; Barrett, M.; Foà, R.; Maurer, J.; O’Brien, S.; Valente, N.; Wenger, M.; Maloney, D.G. Rituximab in B-Cell Hematologic Malignancies: A Review of 20 Years of Clinical Experience. Adv. Ther. 2017, 34, 2232–2273. [Google Scholar] [CrossRef]
- Noga, O.; Hanf, G.; Brachmann, I.; Klucken, A.; Kleinetebbe, J.; Rosseau, S.; Kunkel, G.; Suttorp, N.; Seybold, J. Effect of omalizumab treatment on peripheral eosinophil and T-lymphocyte function in patients with allergic asthma. J. Allergy Clin. Immunol. 2006, 117, 1493–1499. [Google Scholar] [CrossRef]
- Haldar, P.; Brightling, C.E.; Hargadon, B.; Gupta, S.; Monteiro, W.; Sousa, A.; Marshall, R.P.; Bradding, P.; Green, R.H.; Wardlaw, A.J.; et al. Mepolizumab and Exacerbations of Refractory Eosinophilic Asthma. N. Engl. J. Med. 2009, 360, 973–984. [Google Scholar] [CrossRef]
- Agache, I.; Rocha, C.; Beltran, J.; Song, Y.; Posso, M.; Solà, I.; Alonso-Coello, P.; Akdis, C.; Akdis, M.; Canonica, G.W.; et al. Efficacy and safety of treatment with biologicals (benralizumab, dupilumab and omalizumab) for severe allergic asthma: A systematic review for the EAACI Guidelines—Recommendations on the use of biologicals in severe asthma. Allergy 2020, 75, 1043–1057. [Google Scholar] [CrossRef]
- Levy, M.L.; Bacharier, L.B.; Bateman, E.; Boulet, L.-P.; Brightling, C.; Buhl, R.; Brusselle, G.; Cruz, A.A.; Drazen, J.M.; Duijts, L.; et al. Key recommendations for primary care from the 2022 Global Initiative for Asthma (GINA) update. NPJ Prim. Care Respir. Med. 2023, 33, 7. [Google Scholar] [CrossRef]
- Fijolek, J.; Radzikowska, E. Eosinophilic granulomatosis with polyangiitis—Advances in pathogenesis, diagnosis, and treatment. Front. Med. 2023, 10, 1145257. [Google Scholar] [CrossRef]


{kind=link}
| Atopic Asthma | Late-Onset Asthma (LOA) | Factors Contributing to LOA during Menopause |
|---|---|---|
|
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sommer, B.; González-Ávila, G.; Flores-Soto, E.; Montaño, L.M.; Solís-Chagoyán, H.; Romero-Martínez, B.S. Phytoestrogen-Based Hormonal Replacement Therapy Could Benefit Women Suffering Late-Onset Asthma. Int. J. Mol. Sci. 2023, 24, 15335. https://doi.org/10.3390/ijms242015335
Sommer B, González-Ávila G, Flores-Soto E, Montaño LM, Solís-Chagoyán H, Romero-Martínez BS. Phytoestrogen-Based Hormonal Replacement Therapy Could Benefit Women Suffering Late-Onset Asthma. International Journal of Molecular Sciences. 2023; 24(20):15335. https://doi.org/10.3390/ijms242015335
Chicago/Turabian StyleSommer, Bettina, Georgina González-Ávila, Edgar Flores-Soto, Luis M. Montaño, Héctor Solís-Chagoyán, and Bianca S. Romero-Martínez. 2023. "Phytoestrogen-Based Hormonal Replacement Therapy Could Benefit Women Suffering Late-Onset Asthma" International Journal of Molecular Sciences 24, no. 20: 15335. https://doi.org/10.3390/ijms242015335
APA StyleSommer, B., González-Ávila, G., Flores-Soto, E., Montaño, L. M., Solís-Chagoyán, H., & Romero-Martínez, B. S. (2023). Phytoestrogen-Based Hormonal Replacement Therapy Could Benefit Women Suffering Late-Onset Asthma. International Journal of Molecular Sciences, 24(20), 15335. https://doi.org/10.3390/ijms242015335