

Morphometric Characteristics and Genetic Issr Marker Variability in Rhodiola rosea L. (Crassulaceae) in Different Ecological and Geographic Conditions in the Altai Republic
Abstract
:1. Introduction
2. Results and Discussion
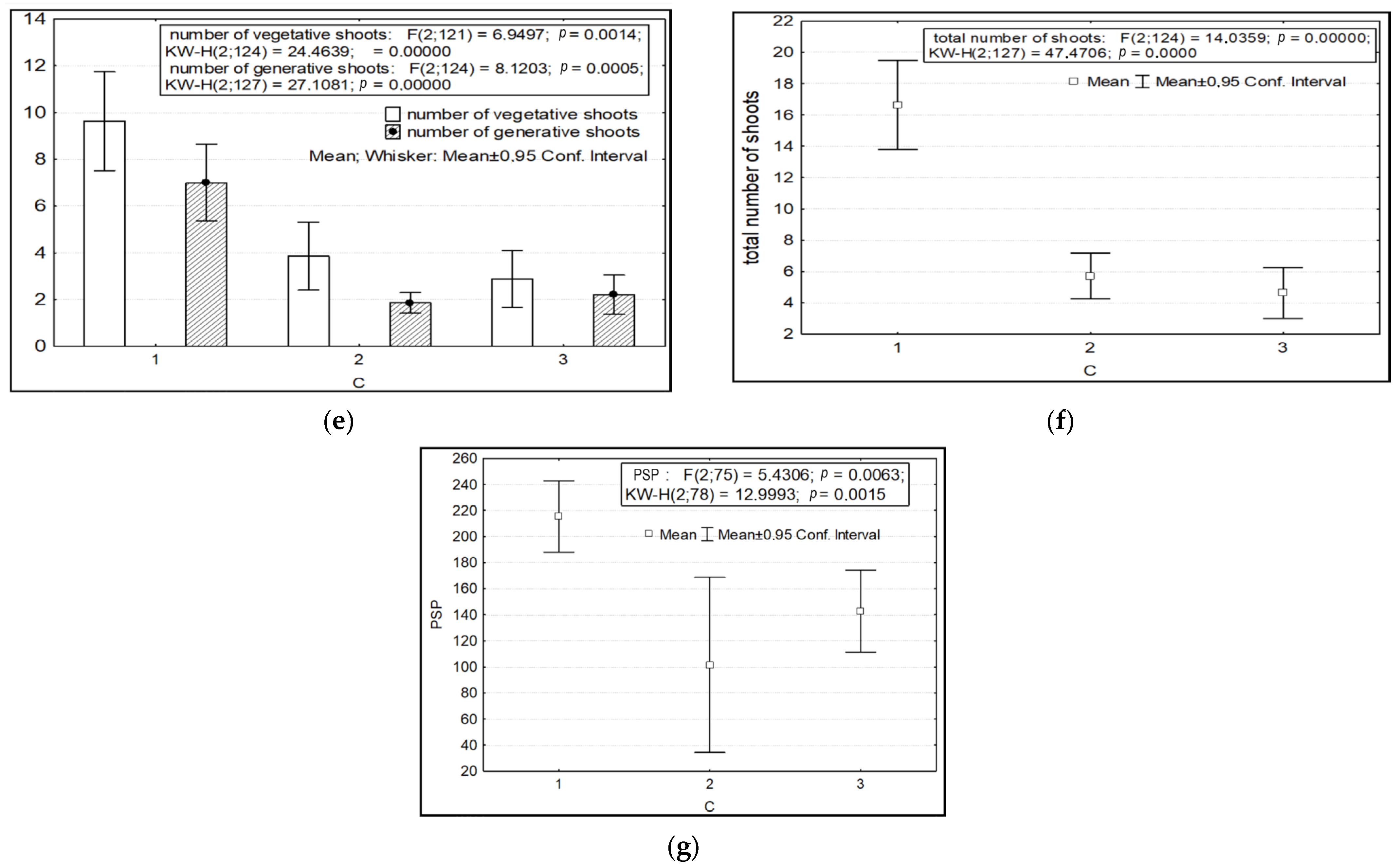
2.1. Results of Morphometric Studies
2.2. Discussion of the Results of Morphometric Studies
2.3. Results of Genetic Analysis and Their Discussion
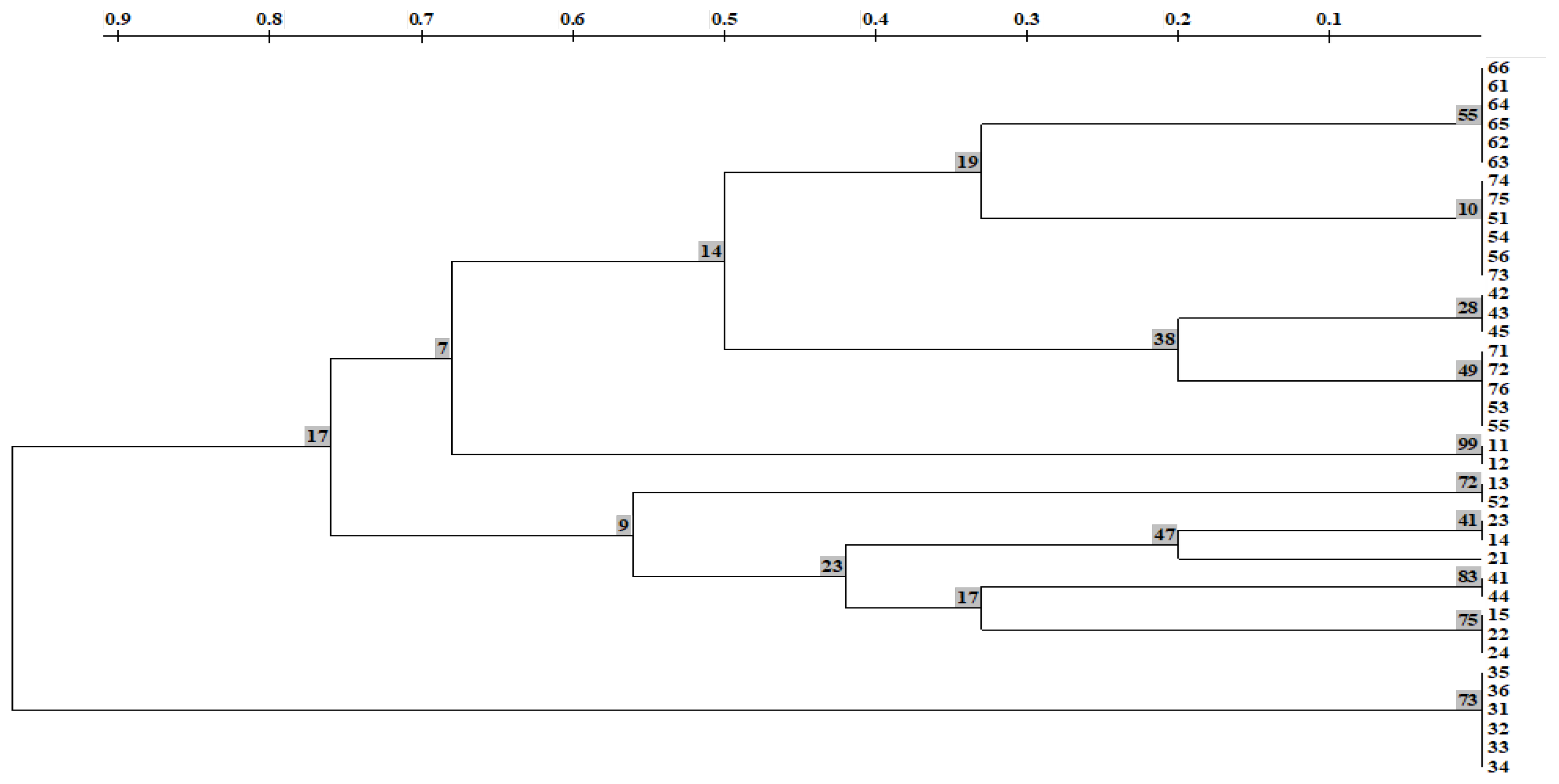
2.3.1. Results of Genetic Analysis
2.3.2. Discussion of the Results of Genetic Analysis
3. Materials and Methods
3.1. Material and Methods of Morphometric Studies
3.2. Statistics of Morphometric Indicators
3.3. Materials and Methods of Genetic Research
3.4. Statistics of Genetic Indicators
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Saratikov, A.S.; Krasnov, E.A. Rhodiola Rosea Is a Valuable Medicinal Plant: Golden Root; Tomsk State University: Tomsk, Russia, 1987; Volume 254. (In Russian) [Google Scholar]
- Yaneva, I.; Balabanski, V.; Karanesheva, T.; Ignatov, I. Some endangered healings plants in Bulgaria-legislative regulation, protection, characteristic description, application, agricultural cultivation. Bulg. J. Agric. Sci. 2020, 26, 847–852. Available online: https://www.agrojournal.org/26/04-20.html (accessed on 21 February 2023).
- Kamelin, R.V. Red Data Book of the Russian Federation (Plants and Fungi); Tovarishchestvo Nauchnyh Izdanij KMK.: Moscow, Russia, 2008; 855p. (In Russian) [Google Scholar]
- Krasnoborov, I.M.; Kim, E.F.; Achimova, A.A. Red Book of the Altai Republic (Plants); Gorno-Altaisk St. University: Gorno-Altajsk, Russia, 2017; pp. 152–153. (In Russian) [Google Scholar]
- György, Z.; Tóth, E.G.; Incze, N.; Molnár, B.; Höhn, M. Intercontinental migration pattern and genetic differentiation of arctic-alpine Rhodiola rosea L.: A chloroplast DNA survey. Ecol. Evol. 2018, 8, 11508–11521. [Google Scholar] [CrossRef] [PubMed]
- Chiang, H.-M.; Chen, H.-C.; Wu, C.-S.; Wu, P.-Y.; Wen, K.-C. Rhodiola plants: Chemistry and biological activity. J. Food Drug. Anal. 2015, 23, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Jafari, M.; Arabit, J.J.G.; Courville, R. The impact of Rhodiola rosea on biomarkers of diabetes, inflammation, and microbiota in a leptin receptor-knockout mouse model. Sci. Rep. 2022, 12, 10581. [Google Scholar] [CrossRef]
- Stojcheva, E.I.; Quintela, J.C. The Effectiveness of Rhodiola rosea L. Preparations in Alleviating Various Aspects of Life-Stress Symptoms and Stress-Induced Conditions—Encouraging Clinical Evidence. Molecules 2022, 27, 3902. [Google Scholar] [CrossRef]
- Brinckmann, J.A.; Cunningham, A.B.; Harter, D.E. Running out of time to smell the roseroots: Reviewing threats and trade in wild Rhodiola rosea L. J. Ethnopharmacol. 2021, 269, 113710. [Google Scholar] [CrossRef]
- Valuiskih, O.E.; Dubrovski, Y.A.; Kuliugina, E.E.; Kanev, V.A. Rare plants of Halmersale mountain vicinities (Northern Urals): Eco-phytocoenotic preferences, population structure and protection. Tomsk. State Univ. J. Biol. 2017, 40, 66–87. [Google Scholar] [CrossRef]
- Kubentayev, S.A.; Zhumagul, M.Z.; Kurmanbayeva, M.S. Current state of populations of Rhodiola rosea L. (Crassulaceae) in East Kazakhstan. Bot. Stud. 2021, 62, 19. [Google Scholar] [CrossRef]
- Chavhan, R.L.; Sable, S.; Narwade, A.V.; Hinge, V.R.; Kalbande, B.B.; Mukherjee, A.K.; Chakrabarty, P.K.; Kadam, U.S. Multiplex molecular marker-assisted analysis of significant pathogens of cotton (Gossypium sp.). Biocatal. Agric. Biotechnol. 2023, 47, 102557. [Google Scholar] [CrossRef]
- Upadhyay, A.; Kadam, U.S.; Chacko, P.; Karibasappa, G.S. Microsatellite and RAPD analysis of grape (Vitis spp.) accessions and identification of duplicates/misnomers in germplasm collection. Indian J. Hortic. 2010, 67, 8–15. [Google Scholar]
- Upadhyay, A.; Kadam, U.S.; Priya, M.C.; Aher, L.; Karibasappa, G.M. Microsatellite analysis to differentiate clones of Thompson seedless grapevine. Indian J. Hortic. 2010, 67, 260–263. [Google Scholar]
- Kylin, M. Genetic Diversity of Roseroot (Rhodiola rosea L.) from Sweden. Master’s Thesis, University of Agricultural Sciences The Faculty of Landscape Planning, Qeqqata Municipality, Greenland, 2010; 58p. [Google Scholar]
- György, Z.; Fjelldal, E.; Szabo, A.; Aspholm, P.E.; Pedryc, A. Genetic diversity of golden root (Rhodiola rosea L.) in northern Norway based on recently developed SSR markers. Turk. J. Biol. 2013, 37, 655–660. [Google Scholar] [CrossRef]
- Zhang, J.-Q.; Meng, S.-Y.; Wen, J.; Rao, G.-Y. Phylogenetic Relationships and Character Evolution of Rhodiola (Crassulaceae) based on Nuclear Ribosomal ITS and Plastid trnL-F and psbA-trnH Sequences. Syst. Bot. 2014, 39, 441–451. [Google Scholar] [CrossRef]
- György, Z.; Vouillamoz, J.; Ladányi, M.; Pedryc, A. Genetic survey of Rhodiola rosea L. populations from the Swiss Alps based on SSR markers. Biochem. Syst. Ecol. 2014, 54, 137–143. [Google Scholar] [CrossRef]
- Zhao, K.; Li, L.; Quan, H. Comparative analyses of chloroplast genomes from Six Rhodiola species: Variable DNA markers identification and phylogenetic relationships within the genus. BMC Genom. 2022, 23, 577. [Google Scholar] [CrossRef] [PubMed]
- Tigimanova, D.; Khapilina, O.; Danilova, A.; Kalendar, R. Genetic diversity of Rhodiola sp. in Altai as assessed by intron polymorphism markers. Biotechnol. Theory Pract. 2017, 2, 42–46. [Google Scholar] [CrossRef]
- Shvydkov, A.V.; Mryasova, K.P.; Asming, S.V.; Tsvetov, N.S.; Nikolaev, V.G. Assessment of the prospects of growing Rhodiola rosea L. (Crassulaceae A. DC.) for the needs of pharmaceutical and food-processing industry Murmansk region. Vestn. Ural. Meditsinskoi Akad. Nauk. J. Ural Med. Acad. Sci. 2019, 16, 296–302. (In Russian) [Google Scholar] [CrossRef]
- Bakhtushkina, A.I.; Podkorytov, A.T. Economically valuable features of yaks of the Altai population. Bull. Altai State Agric. Univ. 2015, 9, 109–112. Available online: https://cyberleninka.ru/article/n/hozyaystvenno-poleznye-priznaki-yakov-altayskoy-populyatsii (accessed on 13 February 2023). (In Russian).
- Donovan, M.; Ross, M. Impacts of grazing on ground cover, soil physical properties and soil loss via surface erosion: A novel geospatial modelling approach. J. Environ. Manag. 2021, 287, 112206. [Google Scholar] [CrossRef]
- Zhmud, E.V.; Dorogina, O.V.; Achimova, A.A. Morphological plasticity of Hedysarum austrosibiricum B. Fedtsch (Fabaceae) shoots under different ecological and geographical conditions. Contemp. Probl. Ecol. 2018, 11, 159–167. [Google Scholar] [CrossRef]
- Zhmud, E.V.; Dorogina, O.V. Ecological Plasticity of plants Astragalus mongholicus (Fabaceae) in the Mountains of Southern Siberia. Contemp. Probl. Ecol. 2015, 8, 351–357. [Google Scholar] [CrossRef]
- Zhmud, E.V.; Achimova, A.A.; Kuban, I.N.; Jamtyrov, M.B.; Dorogina, O.V. Rhaponticum carthamoides (Asteraceae) in the Altai Republic: Assessment of the State of the Plant Affected by Human Activities. J. Sib. Fed. Univ. Biol. 2022, 15, 92–106. [Google Scholar] [CrossRef]
- Robertus, Y.V. (Ed.) Report on the State and Protection of the Environment of the Altai Republic in 2018; Gorno-Altay Branch Institute for Water and Environmental Problems: Gorno-Altaisk, Russia, 2019; 119p, Available online: https://ecoportaltai.ru/page25529994.html (accessed on 25 April 2023).
- Robertus, Y.V. (Ed.) Report on the State and Protection of the Environment of the Altai Republic in 2020; Gorno-Altay branch Institute for Water and Environmental Problems: Gorno-Altaisk, Russia, 2021; 119p, Available online: https://ecoportaltai.ru/page25529994.html (accessed on 27 April 2023).
- Nei, M. Genetic Distance between Populations. Am. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Nei, M.; Li, W.H. Mathematical Model for Studying Genetic Variation in Terms of Restriction Endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [PubMed]
- Barkalov, A.V. Hover-flies (Diptera, Syrphidae) of the high mountains of Altai. Part 1. Fauna distribution. Eur. J. Entomol. 2011, 10, 507–511. (In Russian) [Google Scholar]
- Vedernikova, O.P. Ontogenesis of Rhodiola rosea. In Ontogeneticheskij Atlas Lekarstvennyh Rastenij; MarGU: Joshkar-Ola, Russia, 2000; pp. 199–206. (In Russian) [Google Scholar]
- Modina, T.D. Climates of Altai Republic; Nauka: Novosibirsk, Russia, 1997; 177p. (In Russian) [Google Scholar]
- Plants of the World Online (POWO 2022). Available online: https://powo.science.kew.org/ (accessed on 2 March 2023).
- Available online: https://earth.google.com/web/@50.47470772,87.57177033,2051.47331335a,473472.93127544d,35y (accessed on 20 March 2023).
- Mamaev, S.A. Some questions of the formation of the species of woody plants population structure. Russ. J. Ecol. 1975, 1, 39–49. (In Russian) [Google Scholar]
- Nabieva, A.Y.; Zhmud’, E.V.; Kuban, I.N.; Dorogina, O.V. Morfometricheskii i molekulyarnyi analiz populyatsii Cypripedium × ventricosum (Orchidaceae) v Novosibirskoi oblasti [Morphometric and molecular analysis of the population of Cypripedium × ventricosum (Orchidaceae) in the Novosibirsk region]. Bot. Z. 2020, 105, 78–85. (In Russian) [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Indicators | CP 1 N = 15 | CP 2 N = 15 | CP 3 N = 20 | CP 4 N = 16 | CP 5 N = 20 | CP 6 N = 20 | CP 7 N = 19 | Total N = 125 |
|---|---|---|---|---|---|---|---|---|---|
| 1 | M * ± m Cv,% min-max | 47.2 a ± 2.2 18.9 31.7–64.0 | 44.4 a ± 3.4 30.1 4.5–60.5 | 21.3 b ± 1.0 25.6 13.0–35.0 | 23.4 ab ± 1.3 21.4 14.0–34.0 | 34.8 ab ± 1.5 19.9 25.0–48.0 | 20.7 b ± 0.8 22.3 13.0–32.0 | 28.6 b ± 1.1 18.0 17.0–37.5 | 30.8 ± 1.1 39.4 4.5–64.0 |
| 2 | M ± m Cv, % min-max | 26.8 a ± 2.9 42.5 12.0–57.0 | 19.4 a ± 2.5 49.2 6.0–39.0 | 8.3 ab ± 0.5 32.5 4.0–14.0 | 8.4 ab ± 0.8 36.7 4.0–14.0 | 7.5 ab ± 0.7 41.6 4.0–14.0 | 4.2 b ± 0.4 47.3 1.5–9.0 | 12.5 ab ± 1.1 37.7 6.0–26.5 | 11.9 ± 0.8 77.3 1.5–57.0 |
| 3 | M ± m Cv, % min-max | 2.7 ab ± 0.1 17.2 2.0–3.5 | 3.0 ab ± 0.1 16.6 2.0–4.0 | 2.4 ab ± 0.1 20.5 1.5–3.2 | 2.9 ab ± 0.2 28.5 1.5–4.6 | 2.8 ab ± 0.2 27.3 2.0–4.5 | 1.7 b ± 0.1 31.0 1.0–3.0 | 2.3 ab ± 0.1 19.0 1.7–3.5 | 2.5 ± 0.1 28.4 1.0–4.6 |
| 4 ** | M ± m Cv, % min-max | 1.0 ± 0.1 24.3 0.7–1.5 | 1.1 ± 0.1 19.8 0.8–1.5 | 0.9 ± 0.0 28.3 0.5–1.5 | 1.1 ± 0.1 23.6 0.7–1.6 | 1.0 ± 0.1 32.4 0.5–1.5 | 0.9 ± 0.1 44.8 0.5–1.8 | 0.88 ± 0.04 18.3 0.7–1.2 | 1.00 ± 0.01 29.1 0.5–1.8 |
| 5 | M ± m Cv, % min-max | 55.6 ab ± 6.8 48.8 3.0–116.0 | 81.3 a ± 4.8 22.7 58.0–132.0 | 27.2 b ± 0.8 16.3 19.0–34.0 | 49.0 ab ± 2.4 19.5 34.0–65.0 | 44.8 ab ± 2.3 23.5 22.0–63.0 | 35.9 ab ± 0.9 14.4 27.0–47.0 | 42.8 ab ± 1.6 16.3 29.0–55.0 | 47.0 ± 1.7 41.3 19.0–132.0 |
| 6 | M ± m Cv, % min-max | 10.8 a ± 2.2 80.9 2.0–32.0 | 11.9 ab ± 3.8 123.0 2.0–57.0 | 6.8 ab ± 1.0 82.4 1.0–26.0 | 7.3 ab ± 1.2 67.5 1.0–18.0 | 3.9 b ± 0.7 82.9 0–12.0 | 2.9 b ± 0.4 79.2 1.0–10.0 | 11.8 ab ± 2.8 106.2 0–40.0 | 7.8 ± 0.8 114.5 0–57.0 |
| 7 | M ± m Cv, % min-max | 5.3 ab ± 1.9 141.6 1.0–31.0 | 4.7 ab ± 1.0 78.0 1.0–14.0 | 5.5 ab ± 0.7 72.4 1.0–16.0 | 2.5 ab ± 0.4 61.2 1.0–5.0 | 1.9 b ± 0.2 51.9 1.0–4.0 | 2.2 ab ± 0.3 79.2 1.0–7.0 | 15.2 a ± 2.2 66.1 4.0–40.0 | 5.4 ± 0.6 125.6 1.0–40.0 |
| 8 ** | M ± m Cv, % min-max | 0.8 ± 0.3 150.8 0.1–4.5 | 1.3 ± 0.5 159.9 0.1–7.0 | 1.5 ± 0.4 106.3 0–5.3 | 0.5 ± 0.1 112.0 0.1–2.0 | 1.0 ± 0.2 110.4 0.1–4.0 | 0.9 ± 0.2 87.8 0.2–3.5 | 3.6 ± 1.6 159.3 0.19–20.0 | 1.4 ± 0.2 173.1 0.04–20.0 |
| 9 | M ± m Cv, % min-max | 16.1 a ± 3.6 89.3 5.0–57.0 | 16.6 a ± 3.8 88.4 4.0–60.0 | 12.3 ab ± 1.2 43.4 5.0–27.0 | 9.8 ab ± 1.5 62.7 3.0–22.0 | 5.7 b ± 0.7 56.7 2.0–14.0 | 4.6 b ± 0.8 72.8 2.0–16.0 | 33.2 a ± 4.6 49.7 15.0–58.0 | 13.0 ± 1.1 95.4 2.0–60.0 |
| 10 | M ± m Cv, % min-max | 2.8 ab ± 0.2 27.6 1.8–4.3 | 2.9 ab ± 0.2 20.9 2.0–4.3 | 2.7 ab ± 0.2 27.7 1.7–5.0 | 2.6 ab ± 0.1 17.8 1.7–3.4 | 3.1 ab ± 0.2 35.4 1.9–6.6 | 2.1 b ± 0.2 31.8 0.8–3.0 | 2.7 ab ± 0.1 18.0 1.8–3.5 | 2.7 ± 0.1 28.9 0.8–6.6 |
| Characteristics | Indicators | CP 1 N = 10 | CP 2 N = 15 | CP 3 N = 15 | CP 4 N = 17 | CP 5 N = 4 | CP 6 N = 4 | CP 7 N = 13 | Total N = 78 |
|---|---|---|---|---|---|---|---|---|---|
| Id, ** sm | M * ± m Cv, % min-max | 2.9 ± 0.2 32.2 1.0–5.0 | 2.5 ± 0.3 44.4 1.0–5.0 | 1.9 ± 0.1 26.3 1.1–3.0 | 2.5 ±0.1 19.2 1.5–3.0 | 2.4 ± 0.2 23.8 1.5–3.3 | 2.1 ± 0.2 37.3 1.0–4.0 | 2.4 ± 0.1 32.5 1.0–5.0 | 2.2 ± 0.1 40.7 0.7–5.0 |
| Nf | M ± m Cv, % Min-max | 46.7 ab ± 4.0 26.9 33.0–71.0 | 66.2 a ± 9.1 53.4 20.0–139.0 | 35.3 ab ± 3.5 38.3 18.0–58.0 | 43.1 b ± 3.5 33.4 22.0–79.0 | 23.5 ab ± 4.7 40.1 15.0–37.0 | 28.3 ab ± 4.0 28.1 20.0–39.0 | 55.7 a ± 4.3 27.7 31.0–81.0 | 46.8 ± 2.6 49.0 15.0–139.0 |
| Nc ** | M± m Cv, % Min-max | 4.1 ± 0.0 3.4 4.0–4.4 | 4.3 ± 0.1 6.5 4.0–4.6 | 4.0 ± 0.0 1.7 4.0–4.2 | 4.3 ± 0.1 8.7 3.8–5.0 | 3.5 ± 0.3 16.5 3.0–4.0 | 3.5 ± 0.3 16.5 3.0–4.0 | 4.3 ± 0.1 9.0 4.0–5.0 | 4.10 ± 0.04 9.5 3.0–5.0 |
| PSP | M ± m Cv, % Min-max | 189.9 ab ± 15.4 25.6 132.0–284.0 | 283.0 a ± 40.5 55.4 88.0–639.4 | 142.8 ab ± 14.6 39.7 72.0–243.6 | 185.4 ab ± 15.2 33.8 88.0–316.0 | 85.0 b ± 22.5 52.8 45.0–148.0 | 101.5 b ± 21.1 41.6 60.0–156.0 | 235.3 a ± 16.2 24.8 155.0–324.0 | 195.4 ± 11.5 101.2 45.0–639.4 |
| CP No | Na | Ne | I | h | uh | P |
|---|---|---|---|---|---|---|
| M ± m | M ± m | M ± m | M ± m | M ± m | M ± m | |
| CP 1 | 1.70 ± 0.33 | 1.69 ± 0.16 | 0.53 ± 0.11 | 0.37 ± 0.08 | 0.47 ± 0.09 | 83.33 |
| CP 2 | 1.17 ± 0.40 | 1.43 ± 0.20 | 0.32 ± 0.15 | 0.23 ± 0.10 | 0.31 ± 0.14 | 50.00 |
| CP 3 | 0.17 ± 0.16 | 1 ± 0 | 0 | 0 | 0 | 0 |
| CP 4 | 1.67 ± 0.33 | 1.77 ± 0.15 | 0.56 ± 0.11 | 0.40 ± 0.08 | 0.50 ± 0.10 | 83.33 |
| CP 5 | 1.00 ± 0.44 | 1.36 ± 0.18 | 0.29 ± 0.14 | 0.20 ± 0.09 | 0.24 ± 0.11 | 50.00 |
| CP 6 | 0.33 ± 0.21 | 1.00 ± 0.01 | 0 | 0 | 0 | 0 |
| CP 7 | 0.83 ± 0.40 | 1.33 ± 0.21 | 0.23 ± 0.15 | 0.17 ± 0.10 | 0.2 ± 0.12 | 33.33 |
| M | 0.98 ± 0.15 | 1.37 ± 0.07 | 0.28 ± 0.05 | 0.20 ± 0.04 | 0.25 ± 0.05 | 42.90 |
| CP No | Pairwise Population Matrix of Nei Unbiased Genetic Distance | ||||||
|---|---|---|---|---|---|---|---|
| CP 1 | CP 2 | CP 3 | CP 4 | CP 5 | CP 6 | CP 7 | |
| CP 1 | 0.000 | - | - | - | - | - | - |
| CP 2 | 0.000 | 0.000 | - | - | - | ||
| CP 3 | 0.418 | 0.799 | 0.000 | - | - | ||
| CP 4 | 0.000 | 0.181 | 0.187 | 0.000 | - | - | - |
| CP 5 | 0.103 | 0.317 | 0.448 | 0.000 | 0.000 | - | - |
| CP 6 | 0.224 | 0.431 | 0.693 | 0.381 | 0.265 | 0.000 | - |
| CP 7 | 0.173 | 0.399 | 0.582 | 0.000 | 0.000 | 0.294 | 0.000 |
| CP 1 | CP 2 | CP 3 | CP 4 | CP 5 | CP 6 | CP 7 | |
|---|---|---|---|---|---|---|---|
| CP 1 | 0 | - | - | - | - | - | - |
| CP 2 | 2.6 | 0 | - | - | - | - | - |
| CP 3 | 171.1 | 173.7 | 0 | - | - | - | - |
| CP 4 | 50.8 | 48.2 | 221.7 | 0 | - | - | - |
| CP 5 | 150.5 | 149.2 | 280.2 | 128.6 | 0 | - | - |
| CP 6 | 175.3 | 177.9 | 12.5 | 225.8 | 278.4 | 0 | - |
| CP 7 | 237.3 | 239.6 | 112.9 | 282.5 | 291.6 | 100.4 | 0 |
| CP No | Location, Year of Research | Location Altitude (m Above Sea Level); Slope Exposure, Coordinates | Phytocenosis; Surrounding Plant Species |
|---|---|---|---|
| 1. | Ust’-Koksinskij district, Katunskij nature reserve (KNR), lake shore Poperechnoe, 2020 | 1998 m; southwestern slope, 49.921111° N; 85.889444° E | Subalpine meadow; Aconitum septentrionale Koelle, Betula rotundifolia Regel & Tiling, Carex sp., Chamaenerion angustifolium (L.) Scop., Cotoneaster uniflorus Bunge Dianthus superbus L., Poa pratensis L., Veratrum lobelianum Bernh., Hedysarum neglectum Ledeb., Rumex confertus Willd., Solidago virgaurea L., Salix sp. |
| 2. | Ust’-Koksinskij district, the KNR, water flow between lakes Suroch’e and Verhnee Mul’tinskoe, 2020 | 2070 m; western slope, 49.928056° N; 85.854722° E | Subalpine meadow; Poa pratensis L., Veratrum lobelianum Bernh., Hedysarum neglectum Ledeb., Rumex confertus Willd., Solidago virgaurea L., Aconitum septentrionale Koelle, Salix sp., Saussurea alpina (L.) DC., Sanguisorba alpina Bunge, Delphinium laxiflorum DC. |
| 3. | Kosh-agachskij district; Sailyugemsky National Park (SNP), along the watercourse river Usaj | 2522 m; 49.456944° N; 88.156667° E | Mountain tundra; Dryas oxyodonta Juz., Rumex sp., Ranunculus sp., Dasiphora parvifolia (Fisch.) Juz., Dasiphora parvifolia (Fisch.) Juz., Galium sp., Potentilla sp., Aconitum sp., Gentiana algida Pall., Betula rotundifolia Spach, Artemisia sp., Swertia obtusa Ledeb., Cotoneaster sp., Rhodiola rosea L., Spiraea alpine Pall., Rhodiola coccinea (Royle) Boriss. |
| 4. | Ust’-Koksinskij district, vicinity of the village Kajtanak, slope of Krasnaja Mountain, 2020 | 2000 m; rocky southeastern slope; glacial lake shore 50.076944° N; 85.221667° E | Sparse cedar forest; Betula rotundifolia Spach; Dryas oxyodonta Juz.; Salix KNRylovii E. Wolf.; Salix reticulata L.; Aquilegia sibirica Lam.; Bergenia crassifolia; Bistorta vivipara (L.) Delarb.; Carex melanantha C.A. Mey.; Doronicum altaicum Pall.; Dracocephalum grandiflorum L.; Hedysarum austrosibiricum B. Fedtsch.; Oxyria digyna (L.) Hill.; Platanthera bifolia (L.) Rich.; Potentilla gelida C.A. Mey.; Ranunculus altaicus Laxm.; Rhodiola rosea L.; Swertia obtusa K. Koch; Trollius asiaticus L.; Viola altaica L. |
| 5. | Ongudajskij district, Seminskij pass, 2020 | 1850 m; northern slope; 51.230833° N; 85.351944° E | Sparse cedar forest; Rhaponticum carthamoides (Willd.) Iljin, Geranium albiflorum Ledeb., Bupleurum aureum Fisch. ex Hoffm., Poa palustris L., Trollius altaicus C.A. Mey., Hedysarum neglectum Ledeb. |
| 6. | Kosh-agachskij district; SNP shore of lake Tunguryuk, 2020 | 2412 M; eastern slope; 49.541389° N; 88.256111° E | Grain-grass forb meadow; Ligularia altaica DC., Achillea asiatica Serg., Spiraea alpina Pall., Dasiphora parvifolia (Fisch.) Juz., Gentiana macrophylla Pall., Antennaria dioica (L.) Gaertn., Achillea sp., Veronica sp., Potentilla argentea L., Dracocephalum grandiflorum L., Rhodiola rosea L., Leontopodium ochroleucum Beauverd |
| 7. | Ulagansky district, territory of the Altai Nature Reserve (ANR) in the valley of the Bogoyash River, 2018 | 2260 m; 50.2281° N; 89.1807° E | Mountain tundra; Achillea asiatica, Bistorta vivipara, Gentiana frigida Haenke, Gentiana pneumonante L., Hedysarum consanquineum DC., Phlojodicarpus sibiricus (Fisch. ex Spreng.) Koso-Pol., Poa palustris, Potentilla parvifolia, Rumex acetosa, Rhodiola algida, Saxifraga hirculus L., Schulzia crinita (Pall.) Spreng., Spiraea alpina Pall., Trollius altaicus, Campanula rotundifolia L. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dorogina, O.V.; Kuban, I.N.; Achimova, A.A.; Williams, N.; Lashchinskiy, N.N.; Zhmud, E.V. Morphometric Characteristics and Genetic Issr Marker Variability in Rhodiola rosea L. (Crassulaceae) in Different Ecological and Geographic Conditions in the Altai Republic. Int. J. Mol. Sci. 2023, 24, 15224. https://doi.org/10.3390/ijms242015224
Dorogina OV, Kuban IN, Achimova AA, Williams N, Lashchinskiy NN, Zhmud EV. Morphometric Characteristics and Genetic Issr Marker Variability in Rhodiola rosea L. (Crassulaceae) in Different Ecological and Geographic Conditions in the Altai Republic. International Journal of Molecular Sciences. 2023; 24(20):15224. https://doi.org/10.3390/ijms242015224
Chicago/Turabian StyleDorogina, Olga V., Irina N. Kuban, Altynai A. Achimova, Natasha Williams, Nicolay N. Lashchinskiy, and Elena V. Zhmud. 2023. "Morphometric Characteristics and Genetic Issr Marker Variability in Rhodiola rosea L. (Crassulaceae) in Different Ecological and Geographic Conditions in the Altai Republic" International Journal of Molecular Sciences 24, no. 20: 15224. https://doi.org/10.3390/ijms242015224
APA StyleDorogina, O. V., Kuban, I. N., Achimova, A. A., Williams, N., Lashchinskiy, N. N., & Zhmud, E. V. (2023). Morphometric Characteristics and Genetic Issr Marker Variability in Rhodiola rosea L. (Crassulaceae) in Different Ecological and Geographic Conditions in the Altai Republic. International Journal of Molecular Sciences, 24(20), 15224. https://doi.org/10.3390/ijms242015224