

Tick Salivary Kunitz-Type Inhibitors: Targeting Host Hemostasis and Immunity to Mediate Successful Blood Feeding

 and
and 
Abstract
1. Introduction
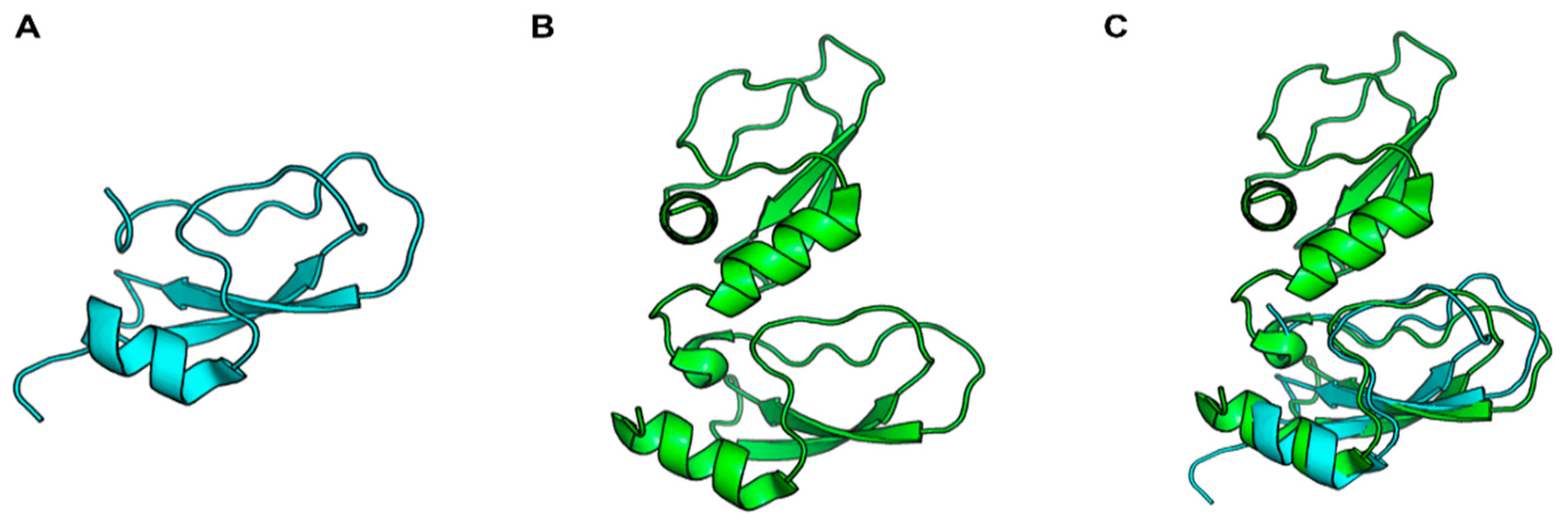
2. Kunitz-Type Inhibitors: Low Molecular Weight Serine Protease Inhibitors
3. Tick Saliva as a Source of Bioactive Kunitz-Type Inhibitors
4. Hemostasis Modulation by Tick Salivary Kunitz Inhibitors

{kind=link}
{kind=link}
| Kunitz Protein | Tick Species | Number of Kunitz Domains | Target Protease(s) | Biological Effect | Transcriptomic Induction/Elevation by Blood Feeding | Vaccine-Related Study (Observed Effect on Ticks) | Reference |
|---|---|---|---|---|---|---|---|
| HA11 | Hyalomma asiaticum | Monolaris | Anticoagulant (intrinsic pathway) | Yes | Yes (reduced engorged body weight) | [48] | |
| Rhipilin-1 | Rhipicephalus hemaphysaloides | Monolaris | Anticoagulant (intrinsic pathway) | Yes | No | [49] | |
| Rhipilin-2 | Rhipicephalus hemaphysaloides | Monolaris | Trypsin, elastase | Anticoagulant (intrinsic pathway) | Yes | No | [50] |
| AsKunitz | Amblyomma sculptum | Monolaris | Thrombin | Anticoagulant, anti-complement (classical and alternative pathways) | Yes | Yes (reduced egg hatching, increased mortality) | [29] |
| Amblyomin-X | Amblyomma sculptum | Monolaris | FXa | Anticoagulant, antithrombotic, antiangiogenic, antitumor (reduces tumor growth and metastasis, induces apoptosis in tumor cell lines) | No | No | [39] |
| Amblin | Amblyomma hebraeum | Bilaris | Thrombin | Anticoagulant | No | No | [51] |
| IrSPI | Ixodes ricinus | Monolaris | Elastase | Immunomodulatory (repression of proliferation of CD4+ T lymphocytes and proinflammatory cytokine secretion from both splenocytes and macrophages) | Yes | Yes (increased engorgement, decreased mortality, increased molting) | [52,53] |
| Ir-CPI | Ixodes ricinus | Monolaris | FXIa, FXIIa, kallikrein | Anticoagulant (intrinsic pathway), antifibrinolytic, antithrombotic | No | No | [44] |
| Ixolaris | Ixodes scapularis | Bilaris | FX(a) | Anticoagulant (extrinsic pathway), antithrombotic, antiangiogenic, antitumor | No | No | [47,54] |
| Penthalaris | Ixodes scapularis | Pentalaris | Fx(a) | Anticoagulant | No | No | [55] |
| Tryptogalinin | Ixodes scapularis | Monolaris | Human skin β-tryptase, matriptase, plasmin, elastase, α-chymotrypsin, trypsin | Not characterized yet | No | No | [56] |
| Ra-KLP | Rhipicephalus appendiculatus | Monolaris | No anti-protease activity | Activates maxiK channels | Yes | No | [57] |
| Boophilin | Rhipicephalus microplus | Bilaris | Thrombin, trypsin, plasmin, FXIa, kallikrein, elastase | Anticoagulant, platelet antiaggregant | No | No | [58,59] |
| rBmTI-A | Rhipicephalus microplus | Bilaris | Trypsin, kallikrein, elastase, plasmin | Anti-inflammatory, antiangiogenic; protective role in pulmonary disorders (emphysema and allergic inflammation) | No | No | [60,61,62,63,64] |
| rBmTI-6 | Rhipicephalus microplus | Trilaris | Trypsin, plasmin | Attenuates inflammation in elastase-induced emphysema | No | No | [65,66] |
| Haemangin | Haemaphysalis longicornis | Monolaris | Trypsin, chymotrypsin, plasmin | Anti-angiogenic (inhibits proliferation and induces apoptosis of endothelial cells), modulates wound healing | Yes | No | [67] |
| HlMKI | Haemaphysalis longicornis | Monolaris | Haemaphysalis longicornis trypsin-like serine proteinase (HlSP) | Not characterized yet | Yes | No | [68] |
| HlChI | Haemaphysalis longicornis | Monolaris | Chymotrypsin, trypsin | Not characterized yet | Yes | No | [69] |
| KPI | Dermacentor variabilis | Pentalaris | Trypsin | Anticoagulant (intrinsic pathway) | Yes (in midgut) | No | [70] |
| Ornithodorin | Ornithodoros moubata | Bilaris | Thrombin | Not characterized yet | No | No | [71] |
| Disagregin | Ornithodoros moubata | No anti-protease activity | Platelet antiaggregant | No | No | [72] | |
| Savignygrin | Ornithodoros savignyi | Monolaris | No anti-protease activity | Platelet antiaggregant | No | No | [73] |
5. Modulation of Host Inflammation and Immunity by Tick Salivary Kunitz Inhibitors
6. Kunitz-Type Inhibitors Used as Vaccine Antigens against Ticks and Tick-Borne Diseases
7. Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Corral-Rodriguez, M.A.; Macedo-Ribeiro, S.; Barbosa Pereira, P.J.; Fuentes-Prior, P. Tick-derived Kunitz-type inhibitors as antihemostatic factors. Insect Biochem. Mol. Biol. 2009, 39, 579–595. [Google Scholar]
- Chmelar, J.; Calvo, E.; Pedra, J.H.; Francischetti, I.M.; Kotsyfakis, M. Tick salivary secretion as a source of antihemostatics. J. Proteom. 2012, 75, 3842–3854. [Google Scholar] [CrossRef]
- Vector-Borne Diseases: Biology of Vector Host Relationship. Available online: https://grantome.com/grant/NIH/ZIA-AI000810-15 (accessed on 21 June 2011).
- Karasuyama, H.; Miyake, K.; Yoshikawa, S. Immunobiology of Acquired Resistance to Ticks. Front. Immunol. 2020, 11, 601504. [Google Scholar] [CrossRef]
- Jmel, M.A.; Aounallah, H.; Bensaoud, C.; Mekki, I.; Chmelar, J.; Faria, F.; M’Ghirbi, Y.; Kotsyfakis, M. Insights into the Role of Tick Salivary Protease Inhibitors during Ectoparasite-Host Crosstalk. Int. J. Mol. Sci. 2021, 22, 892. [Google Scholar] [CrossRef]
- Penalver, E.; Arillo, A.; Delclos, X.; Peris, D.; Grimaldi, D.A.; Anderson, S.R.; Nascimbene, P.C.; Perez-de la Fuente, R. Parasitised feathered dinosaurs as revealed by Cretaceous amber assemblages. Nat. Commun. 2017, 8, 1924. [Google Scholar] [CrossRef]
- Sa-Nunes, A.; Oliveira, C.J.F.; Ribeiro, J.M. Mast Cells and Basophils: From Malevolent Design to Coevolutionary Arms Race. Trends Parasitol. 2020, 36, 655–659. [Google Scholar] [CrossRef]
- Simo, L.; Kazimirova, M.; Richardson, J.; Bonnet, S.I. The Essential Role of Tick Salivary Glands and Saliva in Tick Feeding and Pathogen Transmission. Front. Cell. Infect. Microbiol. 2017, 7, 281. [Google Scholar]
- Nuttall, P.A. Wonders of tick saliva. Ticks Tick-Borne Dis. 2019, 10, 470–481. [Google Scholar] [CrossRef]
- Francischetti, I.M.; Sa-Nunes, A.; Mans, B.J.; Santos, I.M.; Ribeiro, J.M. The role of saliva in tick feeding. Front. Biosci. 2009, 14, 2051–2088. [Google Scholar] [CrossRef]
- Chmelar, J.; Kotal, J.; Langhansova, H.; Kotsyfakis, M. Protease Inhibitors in Tick Saliva: The Role of Serpins and Cystatins in Tick-host-Pathogen Interaction. Front. Cell. Infect. Microbiol. 2017, 7, 216. [Google Scholar]
- Chmelar, J.; Kotal, J.; Karim, S.; Kopacek, P.; Francischetti, I.M.B.; Pedra, J.H.F.; Kotsyfakis, M. Sialomes and Mialomes: A Systems-Biology View of Tick Tissues and Tick-Host Interactions. Trends Parasitol. 2016, 32, 242–254. [Google Scholar]
- Nuttall, P.A.; Labuda, M. Tick-host interactions: Saliva-activated transmission. Parasitology 2004, 129 (Suppl. S1), 77–189. [Google Scholar] [CrossRef]
- Oliva, M.L.; Sampaio, M.U. Action of plant proteinase inhibitors on enzymes of physiopathological importance. An. Acad. Bras. Cienc. 2009, 81, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Stibraniova, I.; Bartikova, P.; Holikova, V.; Kazimirova, M. Deciphering Biological Processes at the Tick-Host Interface Opens New Strategies for Treatment of Human Diseases. Front. Physiol. 2019, 10, 830. [Google Scholar] [PubMed]
- Page, M.J.; Di Cera, E. Serine peptidases: Classification, structure and function. Cell. Mol. Life Sci. 2008, 65, 1220–1236. [Google Scholar] [CrossRef] [PubMed]
- Ranasinghe, S.; McManus, D.P. Structure and function of invertebrate Kunitz serine protease inhibitors. Dev. Comp. Immunol. 2013, 39, 219–227. [Google Scholar] [CrossRef]
- Francischetti, I.M.; Valenzuela, J.G.; Andersen, J.F.; Mather, T.N.; Ribeiro, J.M. Ixolaris, a novel recombinant tissue factor pathway inhibitor (TFPI) from the salivary gland of the tick, Ixodes scapularis: Identification of factor X and factor Xa as scaffolds for the inhibition of factor VIIa/tissue factor complex. Blood 2002, 99, 3602–3612. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, J.M.; Alarcon-Chaidez, F.; Francischetti, I.M.; Mans, B.J.; Mather, T.N.; Valenzuela, J.G.; Wikel, S.K. An annotated catalog of salivary gland transcripts from Ixodes scapularis ticks. Insect Biochem. Mol. Biol. 2006, 36, 111–129. [Google Scholar] [CrossRef]
- Kunitz, M.; Northrop, J.H. Isolation from Beef Pancreas of Crystalline Trypsinogen, Trypsin, a Trypsin Inhibitor, and an Inhibitor-Trypsin Compound. J. Gen. Physiol. 1936, 19, 991–1007. [Google Scholar] [CrossRef]
- Ascenzi, P.; Bocedi, A.; Bolognesi, M.; Spallarossa, A.; Coletta, M.; De Cristofaro, R.; Menegatti, E. The bovine basic pancreatic trypsin inhibitor (Kunitz inhibitor): A milestone protein. Curr. Protein. Pept. Sci. 2003, 4, 231–251. [Google Scholar] [CrossRef]
- Mishra, M. Evolutionary Aspects of the Structural Convergence and Functional Diversification of Kunitz-Domain Inhibitors. J. Mol. Evol. 2020, 88, 537–548. [Google Scholar] [PubMed]
- Flo, M.; Margenat, M.; Pellizza, L.; Grana, M.; Duran, R.; Baez, A.; Salceda, E.; Soto, E.; Alvarez, B.; Fernandez, C. Functional diversity of secreted cestode Kunitz proteins: Inhibition of serine peptidases and blockade of cation channels. PLoS Pathog. 2017, 13, e1006169. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.A.; Kotal, J.; Bensaoud, C.; Chmelar, J.; Kotsyfakis, M. Small protease inhibitors in tick saliva and salivary glands and their role in tick-host-pathogen interactions. Biochim. Biophys. Acta Proteins Proteom. 2020, 1868, 140336. [Google Scholar] [CrossRef] [PubMed]
- de Magalhaes, M.T.Q.; Mambelli, F.S.; Santos, B.P.O.; Morais, S.B.; Oliveira, S.C. Serine protease inhibitors containing a Kunitz domain: Their role in modulation of host inflammatory responses and parasite survival. Microbes Infect. 2018, 20, 606–609. [Google Scholar] [CrossRef]
- Gomes, M.T.; Oliva, M.L.; Lopes, M.T.; Salas, C.E. Plant proteinases and inhibitors: An overview of biological function and pharmacological activity. Curr. Protein Pept. Sci. 2011, 12, 417–436. [Google Scholar] [CrossRef]
- Shamsi, T.N.; Parveen, R.; Fatima, S. Characterization, biomedical and agricultural applications of protease inhibitors: A review. Int. J. Biol. Macromol. 2016, 91, 1120–1133. [Google Scholar] [CrossRef]
- Ribeiro, J.M.C.; Mans, B.J. TickSialoFam (TSFam): A Database That Helps to Classify Tick Salivary Proteins, a Review on Tick Salivary Protein Function and Evolution, With Considerations on the Tick Sialome Switching Phenomenon. Front. Cell. Infect. Microbiol. 2020, 10, 374. [Google Scholar] [CrossRef]
- Costa, G.C.A.; Ribeiro, I.C.T.; Melo-Junior, O.; Gontijo, N.F.; Sant’Anna, M.R.V.; Pereira, M.H.; Pessoa, G.C.D.; Koerich, L.B.; Oliveira, F.; Valenzuela, J.G.; et al. Amblyomma sculptum Salivary Protease Inhibitors as Potential Anti-Tick Vaccines. Front. Immunol. 2020, 11, 611104. [Google Scholar] [CrossRef]
- Reck, J.; Webster, A.; Dall’Agnol, B.; Pienaar, R.; de Castro, M.H.; Featherston, J.; Mans, B.J. Transcriptomic Analysis of Salivary Glands of Ornithodoros brasiliensis Aragao, 1923, the Agent of a Neotropical Tick-Toxicosis Syndrome in Humans. Front. Physiol. 2021, 12, 725635. [Google Scholar] [CrossRef]
- Perez-Sanchez, R.; Carnero-Moran, A.; Soriano, B.; Llorens, C.; Oleaga, A. RNA-seq analysis and gene expression dynamics in the salivary glands of the argasid tick Ornithodoros erraticus along the trophogonic cycle. Parasites Vectors 2021, 14, 170. [Google Scholar]
- Oleaga, A.; Soriano, B.; Llorens, C.; Perez-Sanchez, R. Sialotranscriptomics of the argasid tick Ornithodoros moubata along the trophogonic cycle. PLoS Negl. Trop. Dis. 2021, 15, e0009105. [Google Scholar] [CrossRef]
- Garcia, G.R.; Gardinassi, L.G.; Ribeiro, J.M.; Anatriello, E.; Ferreira, B.R.; Moreira, H.N.; Mafra, C.; Martins, M.M.; Szabo, M.P.; de Miranda-Santos, I.K.; et al. The sialotranscriptome of Amblyomma triste, Amblyomma parvum and Amblyomma cajennense ticks, uncovered by 454-based RNA-seq. Parasit Vectors 2014, 7, 430. [Google Scholar] [CrossRef] [PubMed]
- Karim, S.; Kumar, D.; Adamson, S.; Ennen, J.R.; Qualls, C.P.; Ribeiro, J.M.C. The sialotranscriptome of the gopher-tortoise tick, Amblyomma tuberculatum. Ticks Tick-Borne Dis. 2021, 12, 101560. [Google Scholar] [CrossRef] [PubMed]
- Karim, S.; Ribeiro, J.M. An Insight into the Sialome of the Lone Star Tick, Amblyomma americanum, with a Glimpse on Its Time Dependent Gene Expression. PLoS ONE 2015, 10, e0131292. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, S.R.; Garcia, G.R.; Teixeira, F.R.; Brandao, L.G.; Anderson, J.M.; Ribeiro, J.M.C.; Valenzuela, J.G.; Horackova, J.; Verissimo, C.J.; Katiki, L.M.; et al. Mining a differential sialotranscriptome of Rhipicephalus microplus guides antigen discovery to formulate a vaccine that reduces tick infestations. Parasite Vector 2017, 10. [Google Scholar] [CrossRef] [PubMed]
- Araujo, R.N.; Silva, N.C.S.; Mendes-Sousa, A.; Paim, R.; Costa, G.C.A.; Dias, L.R.; Oliveira, K.; Sant’Anna, M.R.V.; Gontijo, N.F.; Pereira, M.H.; et al. RNA-seq analysis of the salivary glands and midgut of the Argasid tick Ornithodoros rostratus. Sci. Rep. 2019, 9, 6764. [Google Scholar] [CrossRef]
- Chmelar, J.; Oliveira, C.J.; Rezacova, P.; Francischetti, I.M.; Kovarova, Z.; Pejler, G.; Kopacek, P.; Ribeiro, J.M.; Mares, M.; Kopecky, J.; et al. A tick salivary protein targets cathepsin G and chymase and inhibits host inflammation and platelet aggregation. Blood 2011, 117, 736–744. [Google Scholar] [CrossRef]
- Branco, V.G.; Iqbal, A.; Alvarez-Flores, M.P.; Sciani, J.M.; de Andrade, S.A.; Iwai, L.K.; Serrano, S.M.; Chudzinski-Tavassi, A.M. Amblyomin-X having a Kunitz-type homologous domain, is a noncompetitive inhibitor of FXa and induces anticoagulation in vitro and in vivo. Biochim Biophys Acta Proteins Proteom. 2016, 1864, 1428–1435. [Google Scholar] [CrossRef]
- Kolte, D.; Shariat-Madar, Z. Plasma Kallikrein Inhibitors in Cardiovascular Disease: An Innovative Therapeutic Approach. Cardiol. Rev. 2016, 24, 99–109. [Google Scholar] [CrossRef]
- Waxman, L.; Smith, D.E.; Arcuri, K.E.; Vlasuk, G.P. Tick anticoagulant peptide (TAP) is a novel inhibitor of blood coagulation factor Xa. Science 1990, 248, 593–596. [Google Scholar] [CrossRef]
- Schaffer, L.W.; Davidson, J.T.; Vlasuk, G.P.; Siegl, P.K. Antithrombotic efficacy of recombinant tick anticoagulant peptide. A potent inhibitor of coagulation factor Xa in a primate model of arterial thrombosis. Circulation 1991, 84, 1741–1748. [Google Scholar] [CrossRef] [PubMed]
- Karczewski, J.; Connolly, T.M. The interaction of disagregin with the platelet fibrinogen receptor, glycoprotein IIb-IIIa. Blood 1996, 88, 85. [Google Scholar] [CrossRef]
- Decrem, Y.; Rath, G.; Blasioli, V.; Cauchie, P.; Robert, S.; Beaufays, J.; Frere, J.M.; Feron, O.; Dogne, J.M.; Dessy, C.; et al. Ir-CPI, a coagulation contact phase inhibitor from the tick Ixodes ricinus, inhibits thrombus formation without impairing hemostasis. J. Exp. Med. 2009, 206, 2381–2395. [Google Scholar] [CrossRef] [PubMed]
- Akagi, E.M.; de Sa Junior, P.L.; Simons, S.M.; Bellini, M.H.; Barreto, S.A.; Chudzinski-Tavassi, A.M. Corrigendum to “Pro-apoptotic effects of Amblyomin-X in murine renal cell carcinoma “in vitro” [Biomed. Pharmacother. 66 (2012) 64-69]. Biomed. Pharmacother. 2019, 118, 109108. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, R.Q.; Rezaie, A.R.; Bae, J.S.; Calvo, E.; Andersen, J.F.; Francischetti, I.M. Ixolaris binding to factor X reveals a precursor state of factor Xa heparin-binding exosite. Protein Sci. 2008, 17, 146–153. [Google Scholar] [CrossRef]
- Nazareth, R.A.; Tomaz, L.S.; Ortiz-Costa, S.; Atella, G.C.; Ribeiro, J.M.; Francischetti, I.M.; Monteiro, R.Q. Antithrombotic properties of Ixolaris, a potent inhibitor of the extrinsic pathway of the coagulation cascade. Thromb. Haemost. 2006, 96, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Qiao, R.; Gong, H.; Cao, J.; Zhou, Y.; Zhou, J. Identification and anticoagulant activity of a novel Kunitz-type protein HA11 from the salivary gland of the tick Hyalomma asiaticum. Exp. Appl. Acarol. 2017, 71, 71–85. [Google Scholar] [CrossRef]
- Gao, X.; Shi, L.; Zhou, Y.; Cao, J.; Zhang, H.; Zhou, J. Characterization of the anticoagulant protein Rhipilin-1 from the Rhipicephalus haemaphysaloides tick. J. Insect. Physiol. 2011, 57, 339–343. [Google Scholar] [CrossRef]
- Cao, J.; Shi, L.; Zhou, Y.; Gao, X.; Zhang, H.; Gong, H.; Zhou, J. Characterization of a new Kunitz-type serine protease inhibitor from the hard tick Rhipicephalus hemaphysaloides. Arch. Insect. Biochem. Physiol. 2013, 84, 104–113. [Google Scholar] [CrossRef]
- Lai, R.; Takeuchi, H.; Jonczy, J.; Rees, H.H.; Turner, P.C. A thrombin inhibitor from the ixodid tick, Amblyomma hebraeum. Gene 2004, 342, 243–249. [Google Scholar] [CrossRef]
- Blisnick, A.A.; Simo, L.; Grillon, C.; Fasani, F.; Brule, S.; Le Bonniec, B.; Prina, E.; Marsot, M.; Relmy, A.; Blaise-Boisseau, S.; et al. The Immunomodulatory Effect of IrSPI, a Tick Salivary Gland Serine Protease Inhibitor Involved in Ixodes ricinus Tick Feeding. Vaccines 2019, 7, 148. [Google Scholar] [CrossRef] [PubMed]
- Almazan, C.; Fourniol, L.; Rakotobe, S.; Simo, L.; Borneres, J.; Cote, M.; Peltier, S.; Maye, J.; Versille, N.; Richardson, J.; et al. Failed Disruption of Tick Feeding, Viability, and Molting after Immunization of Mice and Sheep with Recombinant Ixodes ricinus Salivary Proteins IrSPI and IrLip1. Vaccines 2020, 8, 475. [Google Scholar] [CrossRef]
- Carneiro-Lobo, T.C.; Konig, S.; Machado, D.E.; Nasciutti, L.E.; Forni, M.F.; Francischetti, I.M.; Sogayar, M.C.; Monteiro, R.Q. Ixolaris, a tissue factor inhibitor, blocks primary tumor growth and angiogenesis in a glioblastoma model. J. Thromb. Haemost. 2009, 7, 1855–1864. [Google Scholar] [CrossRef] [PubMed]
- Francischetti, I.M.; Mather, T.N.; Ribeiro, J.M. Penthalaris, a novel recombinant five-Kunitz tissue factor pathway inhibitor (TFPI) from the salivary gland of the tick vector of Lyme disease, Ixodes scapularis. Thromb. Haemost. 2004, 91, 886–898. [Google Scholar]
- Valdes, J.J.; Schwarz, A.; Cabeza de Vaca, I.; Calvo, E.; Pedra, J.H.; Guallar, V.; Kotsyfakis, M. Tryptogalinin is a tick Kunitz serine protease inhibitor with a unique intrinsic disorder. PLoS ONE 2013, 8, e62562. [Google Scholar] [CrossRef] [PubMed]
- Paesen, G.C.; Siebold, C.; Dallas, M.L.; Peers, C.; Harlos, K.; Nuttall, P.A.; Nunn, M.A.; Stuart, D.I.; Esnouf, R.M. An ion-channel modulator from the saliva of the brown ear tick has a highly modified Kunitz/BPTI structure. J. Mol. Biol. 2009, 389, 734–747. [Google Scholar] [CrossRef]
- Soares, T.S.; Watanabe, R.M.; Tanaka-Azevedo, A.M.; Torquato, R.J.; Lu, S.; Figueiredo, A.C.; Pereira, P.J.; Tanaka, A.S. Expression and functional characterization of boophilin, a thrombin inhibitor from Rhipicephalus (Boophilus) microplus midgut. Vet. Parasitol. 2012, 187, 521–528. [Google Scholar] [CrossRef]
- Assumpcao, T.C.; Ma, D.; Mizurini, D.M.; Kini, R.M.; Ribeiro, J.M.; Kotsyfakis, M.; Monteiro, R.Q.; Francischetti, I.M. In Vitro Mode of Action and Anti-thrombotic Activity of Boophilin, a Multifunctional Kunitz Protease Inhibitor from the Midgut of a Tick Vector of Babesiosis, Rhipicephalus microplus. PLoS Negl. Trop. Dis. 2016, 10, e0004298. [Google Scholar] [CrossRef]
- Sasaki, S.D.; Azzolini, S.S.; Hirata, I.Y.; Andreotti, R.; Tanaka, A.S. Boophilus microplus tick larvae, a rich source of Kunitz type serine proteinase inhibitors. Biochimie 2004, 86, 643–649. [Google Scholar] [CrossRef]
- Soares, T.S.; Oliveira, F.; Torquato, R.J.; Sasaki, S.D.; Araujo, M.S.; Paschoalin, T.; Tanaka, A.S. BmTI-A, a Kunitz type inhibitor from Rhipicephalus microplus able to interfere in vessel formation. Vet. Parasitol. 2016, 219, 44–52. [Google Scholar] [CrossRef]
- Florencio, A.C.; de Almeida, R.S.; Arantes-Costa, F.M.; Saraiva-Romanholo, B.M.; Duran, A.F.; Sasaki, S.D.; Martins, M.A.; Lopes, F.; Tiberio, I.; Leick, E.A. Effects of the serine protease inhibitor rBmTI-A in an experimental mouse model of chronic allergic pulmonary inflammation. Sci. Rep. 2019, 9, 12624. [Google Scholar] [CrossRef] [PubMed]
- Lourenco, J.D.; Ito, J.T.; Cervilha, D.A.B.; Sales, D.S.; Riani, A.; Suehiro, C.L.; Genaro, I.S.; Duran, A.; Puzer, L.; Martins, M.A.; et al. The tick-derived rBmTI-A protease inhibitor attenuates the histological and functional changes induced by cigarette smoke exposure. Histol Histopathol 2018, 33, 289–298. [Google Scholar] [PubMed]
- Lourenco, J.D.; Neves, L.P.; Olivo, C.R.; Duran, A.; Almeida, F.M.; Arantes, P.M.; Prado, C.M.; Leick, E.A.; Tanaka, A.S.; Martins, M.A.; et al. A treatment with a protease inhibitor recombinant from the cattle tick (Rhipicephalus Boophilus microplus) ameliorates emphysema in mice. PLoS ONE 2014, 9, e98216. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, S.D.; Tanaka, A.S. rBmTI-6, a Kunitz-BPTI domain protease inhibitor from the tick Boophilus microplus, its cloning, expression and biochemical characterization. Vet. Parasitol. 2008, 155, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Duran, A.F.A.; Neves, L.P.; da Silva, F.R.S.; Machado, G.C.; Ferreira, G.C.; Lourenco, J.D.; Tanaka, A.S.; Martins, M.A.; Lopes, F.; Sasaki, S.D. rBmTI-6 attenuates pathophysiological and inflammatory parameters of induced emphysema in mice. Int. J. Biol. Macromol. 2018, 111, 1214–1221. [Google Scholar] [CrossRef]
- Islam, M.K.; Tsuji, N.; Miyoshi, T.; Alim, M.A.; Huang, X.; Hatta, T.; Fujisaki, K. The Kunitz-like modulatory protein haemangin is vital for hard tick blood-feeding success. PLoS Pathog. 2009, 5, e1000497. [Google Scholar] [CrossRef]
- Miyoshi, T.; Tsuji, N.; Islam, M.K.; Alim, M.A.; Hatta, T.; Yamaji, K.; Anisuzzaman Fujisaki, K. A Kunitz-type proteinase inhibitor from the midgut of the ixodid tick, Haemaphysalis longicornis, and its endogenous target serine proteinase. Mol. Biochem. Parasitol. 2010, 170, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Alim, M.A.; Islam, M.K.; Anisuzzaman Miyoshi, T.; Hatta, T.; Yamaji, K.; Matsubayashi, M.; Fujisaki, K.; Tsuji, N. A hemocyte-derived Kunitz-BPTI-type chymotrypsin inhibitor, HlChI, from the ixodid tick Haemaphysalis longicornis, plays regulatory functions in tick blood-feeding processes. Insect Biochem. Mol. Biol. 2012, 42, 925–934. [Google Scholar] [CrossRef]
- Ceraul, S.M.; Dreher-Lesnick, S.M.; Mulenga, A.; Rahman, M.S.; Azad, A.F. Functional characterization and novel rickettsiostatic effects of a Kunitz-type serine protease inhibitor from the tick Dermacentor variabilis. Infect. Immun. 2008, 76, 5429–5435. [Google Scholar] [CrossRef]
- van de Locht, A.; Stubbs, M.T.; Bode, W.; Friedrich, T.; Bollschweiler, C.; Höffken, W.; Huber, R. The ornithodorin-thrombin crystal structure, a key to the TAP enigma? EMBO J. 1996, 15, 6011–6017. [Google Scholar] [CrossRef]
- Karczewski, J.; Endris, R.; Connolly, T.M. Disagregin Is a Fibrinogen Receptor Antagonist Lacking the Arg-Gly-Asp Sequence from the Tick, Ornithodoros-Moubata. J. Biol. Chem. 1994, 269, 6702–6708. [Google Scholar] [CrossRef] [PubMed]
- Mans, B.J.; Louw, A.I.; Neitz, A.W. Savignygrin, a platelet aggregation inhibitor from the soft tick Ornithodoros savignyi, presents the RGD integrin recognition motif on the Kunitz-BPTI fold. J. Biol. Chem. 2002, 277, 21371–21378. [Google Scholar] [CrossRef] [PubMed]
- Ceraul, S.M.; Chung, A.; Sears, K.T.; Popov, V.L.; Beier-Sexton, M.; Rahman, M.S.; Azad, A.F. A Kunitz protease inhibitor from Dermacentor variabilis, a vector for spotted fever group rickettsiae, limits Rickettsia montanensis invasion. Infect. Immun. 2011, 79, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Manen, J.F.; Simon, P.; Van Slooten, J.C.; Osteras, M.; Frutiger, S.; Hughes, G.J. A nodulin specifically expressed in senescent nodules of winged bean is a protease inhibitor. Plant Cell 1991, 3, 259–270. [Google Scholar]
- Levi, M.; van der Poll, T.; Buller, H.R. Bidirectional relation between inflammation and coagulation. Circulation 2004, 109, 2698–2704. [Google Scholar] [CrossRef]
- Francischetti, I.M.; Seydel, K.B.; Monteiro, R.Q. Blood coagulation, inflammation, and malaria. Microcirculation 2008, 15, 81–107. [Google Scholar] [CrossRef]
- Schechter, M.E.; Andrade, B.B.; He, T.; Richter, G.H.; Tosh, K.W.; Policicchio, B.B.; Singh, A.; Raehtz, K.D.; Sheikh, V.; Ma, D.; et al. Inflammatory monocytes expressing tissue factor drive SIV and HIV coagulopathy. Sci. Transl. Med. 2017, 9, eaam5441. [Google Scholar] [CrossRef]
- Paesen, G.C.; Siebold, C.; Harlos, K.; Peacey, M.F.; Nuttall, P.A.; Stuart, D.I. A tick protein with a modified Kunitz fold inhibits human tryptase. J. Mol. Biol. 2007, 368, 1172–1186. [Google Scholar] [CrossRef]
- Hellman, L.; Akula, S.; Fu, Z.; Wernersson, S. Mast Cell and Basophil Granule Proteases—In Vivo Targets and Function. Front. Immunol. 2022, 13, 918305. [Google Scholar] [CrossRef]
- Valdes, J.J.; Moal, I.H. Prediction of Kunitz ion channel effectors and protease inhibitors from the Ixodes ricinus sialome. Ticks Tick-Borne Dis. 2014, 5, 947–950. [Google Scholar] [CrossRef]
- Kettritz, R. Neutral serine proteases of neutrophils. Immunol. Rev. 2016, 273, 232–248. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, P.A. The potential of neutrophil elastase inhibitors as anti-inflammatory therapies. Curr. Opin. Hematol. 2014, 21, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, G.C.; Bomediano Camillo, L.M.; Sasaki, S.D. Structural and functional properties of rBmTI-A. A Kunitz-BPTI serine protease inhibitor with therapeutical potential. Biochimie, 2022; in press. [Google Scholar] [CrossRef] [PubMed]
- Batista, I.F.; Ramos, O.H.; Ventura, J.S.; Junqueira-de-Azevedo, I.L.; Ho, P.L.; Chudzinski-Tavassi, A.M. A new Factor Xa inhibitor from Amblyomma cajennense with a unique domain composition. Arch. Biochem. Biophys. 2010, 493, 151–156. [Google Scholar] [CrossRef]
- Pasqualoto, K.F.; Balan, A.; Barreto, S.A.; Simons, S.M.; Chudzinski-Tavassi, A.M. Structural findings and molecular modeling approach of a TFPI-like inhibitor. Protein Pept. Lett. 2014, 21, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Drewes, C.C.; Dias, R.Y.; Hebeda, C.B.; Simons, S.M.; Barreto, S.A.; Ferreira, J.M., Jr.; Chudzinski-Tavassi, A.M.; Farsky, S.H. Actions of the Kunitz-type serine protease inhibitor Amblyomin-X on VEGF-A-induced angiogenesis. Toxicon 2012, 60, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Chudzinski-Tavassi, A.M.; De-Sa-Junior, P.L.; Simons, S.M.; Maria, D.A.; de Souza Ventura, J.; Batista, I.F.; Faria, F.; Duraes, E.; Reis, E.M.; Demasi, M. A new tick Kunitz type inhibitor, Amblyomin-X, induces tumor cell death by modulating genes related to the cell cycle and targeting the ubiquitin-proteasome system. Toxicon 2010, 56, 1145–1154. [Google Scholar] [CrossRef]
- Ventura, J.S.; Faria, F.; Batista, I.F.; Simons, S.M.; Oliveira, D.G.; Morais, K.L.; Chudzinski-Tavassi, A.M. A Kunitz-type FXa inhibitor affects tumor progression, hypercoagulable state and triggers apoptosis. Biomed. Pharmacother. 2013, 67, 192–196. [Google Scholar] [CrossRef]
- Maria, D.A.; de Souza, J.G.; Morais, K.L.; Berra, C.M.; Zampolli Hde, C.; Demasi, M.; Simons, S.M.; de Freitas Saito, R.; Chammas, R.; Chudzinski-Tavassi, A.M. A novel proteasome inhibitor acting in mitochondrial dysfunction, ER stress and ROS production. Invest. New Drugs 2013, 31, 493–505. [Google Scholar] [CrossRef]
- Ali, A.; Zeb, I.; Alouffi, A.; Zahid, H.; Almutairi, M.M.; Ayed Alshammari, F.; Alrouji, M.; Termignoni, C.; Vaz, I.D.S., Jr.; Tanaka, T. Host Immune Responses to Salivary Components—A Critical Facet of Tick-Host Interactions. Front. Cell. Infect. Microbiol. 2022, 12, 809052. [Google Scholar] [CrossRef]
- Skare, J.T.; Garcia, B.L. Complement Evasion by Lyme Disease Spirochetes. Trends Microbiol. 2020, 28, 889–899. [Google Scholar] [CrossRef] [PubMed]
- Torina, A.; Villari, S.; Blanda, V.; Vullo, S.; La Manna, M.P.; Shekarkar Azgomi, M.; Di Liberto, D.; de la Fuente, J.; Sireci, G. Innate Immune Response to Tick-Borne Pathogens: Cellular and Molecular Mechanisms Induced in the Hosts. Int. J. Mol. Sci. 2020, 21, 5437. [Google Scholar] [CrossRef] [PubMed]
- Willadsen, P. Anti-tick vaccines. Parasitology 2004, 129 (Suppl. S3), 67–387. [Google Scholar] [CrossRef] [PubMed]
- Andreotti, R.; Gomes, A.; Malavazi-Piza, K.C.; Sasaki, S.D.; Sampaio, C.A.; Tanaka, A.S. BmTI antigens induce a bovine protective immune response against Boophilus microplus tick. Int. Immunopharmacol. 2002, 2, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Andreotti, R. A synthetic bmti n-terminal fragment as antigen in bovine immunoprotection against the tick Boophilus microplus in a pen trial. Exp. Parasitol. 2007, 116, 66–70. [Google Scholar] [CrossRef]
- Andreotti, R.; Cunha, R.C.; Soares, M.A.; Guerrero, F.D.; Leite, F.P.; de Leon, A.A. Protective immunity against tick infestation in cattle vaccinated with recombinant trypsin inhibitor of Rhipicephalus microplus. Vaccine 2012, 30, 6678–6685. [Google Scholar] [CrossRef]
- de la Fuente, J.; Kocan, K.M. The Impact of RNA Interference in Tick Research. Pathogens 2022, 11, 827. [Google Scholar] [CrossRef]
- Liao, M.; Zhou, J.; Gong, H.; Boldbaatar, D.; Shirafuji, R.; Battur, B.; Nishikawa, Y.; Fujisaki, K. Hemalin, a thrombin inhibitor isolated from a midgut cDNA library from the hard tick Haemaphysalis longicornis. J. Insect. Physiol. 2009, 55, 164–173. [Google Scholar] [CrossRef]
- Macedo-Ribeiro, S.; Almeida, C.; Calisto, B.M.; Friedrich, T.; Mentele, R.; Sturzebecher, J.; Fuentes-Prior, P.; Pereira, P.J. Isolation, cloning and structural characterisation of boophilin, a multifunctional Kunitz-type proteinase inhibitor from the cattle tick. PLoS ONE 2008, 3, e1624. [Google Scholar] [CrossRef]
- Valenzuela, J.G.; Francischetti, I.M.; Pham, V.M.; Garfield, M.K.; Mather, T.N.; Ribeiro, J.M. Exploring the sialome of the tick Ixodes scapularis. J. Exp. Biol. 2002, 205, 2843–2864. [Google Scholar] [CrossRef]
- Lobba, A.R.M.; Alvarez-Flores, M.P.; Fessel, M.R.; Buri, M.V.; Oliveira, D.S.; Gomes, R.N.; Cunegundes, P.S.; DeOcesano-Pereira, C.; Cinel, V.D.; Chudzinski-Tavassi, A.M. A Kunitz-type inhibitor from tick salivary glands: A promising novel antitumor drug candidate. Front. Mol. Biosci. 2022, 9, 936107. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jmel, M.A.; Voet, H.; Araújo, R.N.; Tirloni, L.; Sá-Nunes, A.; Kotsyfakis, M. Tick Salivary Kunitz-Type Inhibitors: Targeting Host Hemostasis and Immunity to Mediate Successful Blood Feeding. Int. J. Mol. Sci. 2023, 24, 1556. https://doi.org/10.3390/ijms24021556
Jmel MA, Voet H, Araújo RN, Tirloni L, Sá-Nunes A, Kotsyfakis M. Tick Salivary Kunitz-Type Inhibitors: Targeting Host Hemostasis and Immunity to Mediate Successful Blood Feeding. International Journal of Molecular Sciences. 2023; 24(2):1556. https://doi.org/10.3390/ijms24021556
Chicago/Turabian StyleJmel, Mohamed Amine, Hanne Voet, Ricardo N. Araújo, Lucas Tirloni, Anderson Sá-Nunes, and Michail Kotsyfakis. 2023. "Tick Salivary Kunitz-Type Inhibitors: Targeting Host Hemostasis and Immunity to Mediate Successful Blood Feeding" International Journal of Molecular Sciences 24, no. 2: 1556. https://doi.org/10.3390/ijms24021556
APA StyleJmel, M. A., Voet, H., Araújo, R. N., Tirloni, L., Sá-Nunes, A., & Kotsyfakis, M. (2023). Tick Salivary Kunitz-Type Inhibitors: Targeting Host Hemostasis and Immunity to Mediate Successful Blood Feeding. International Journal of Molecular Sciences, 24(2), 1556. https://doi.org/10.3390/ijms24021556