
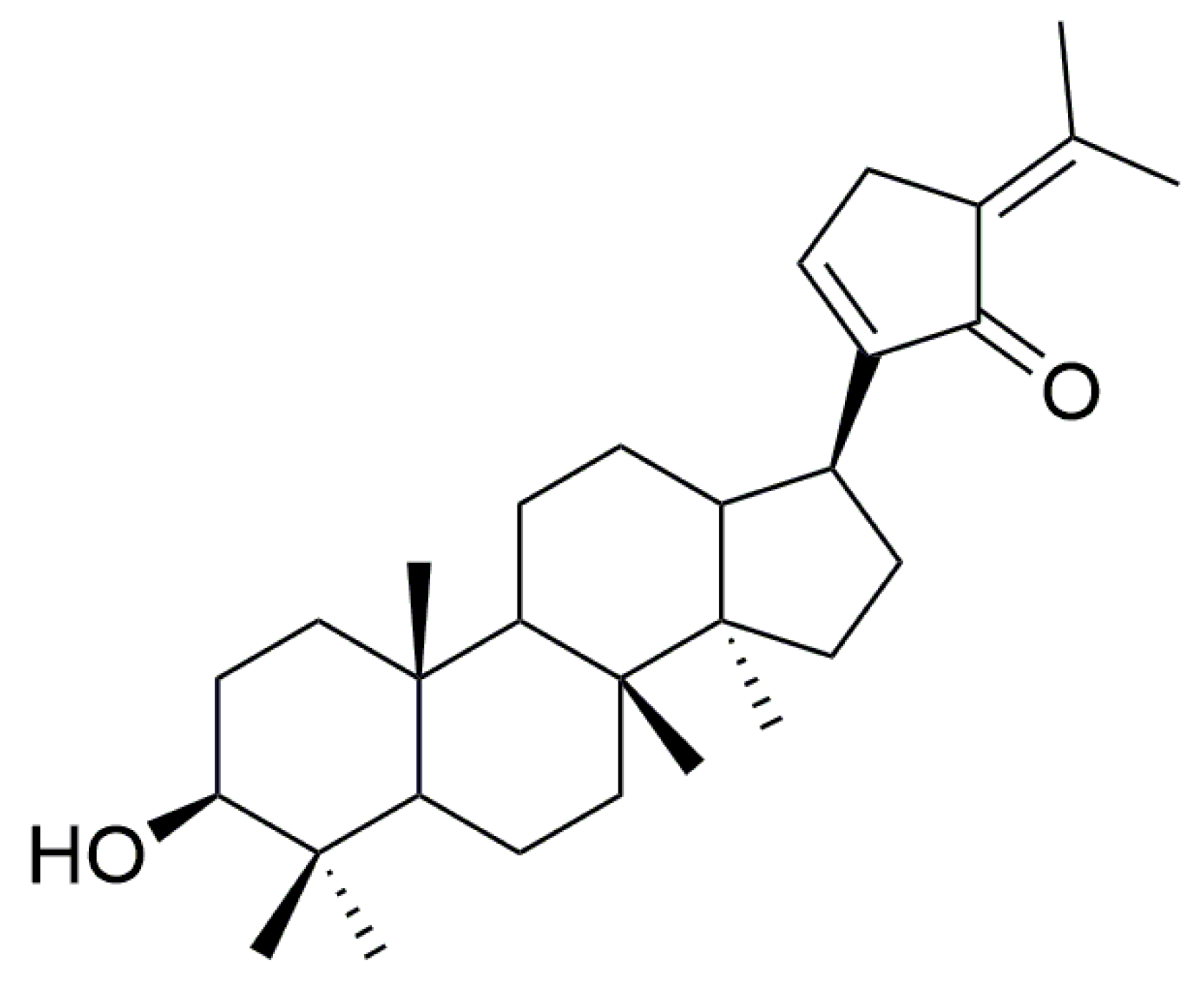
Identification of Human UDP-Glucuronosyltransferase Involved in Gypensapogenin C Glucuronidation and Species Differences
 ,
,
Abstract
1. Introduction
2. Results
2.1. Glucuronidation of GPC in Human Liver Microsomes
2.2. Kinetics of GPC Glucuronidation in HLMs
2.3. Chemical Inhibition Experiment
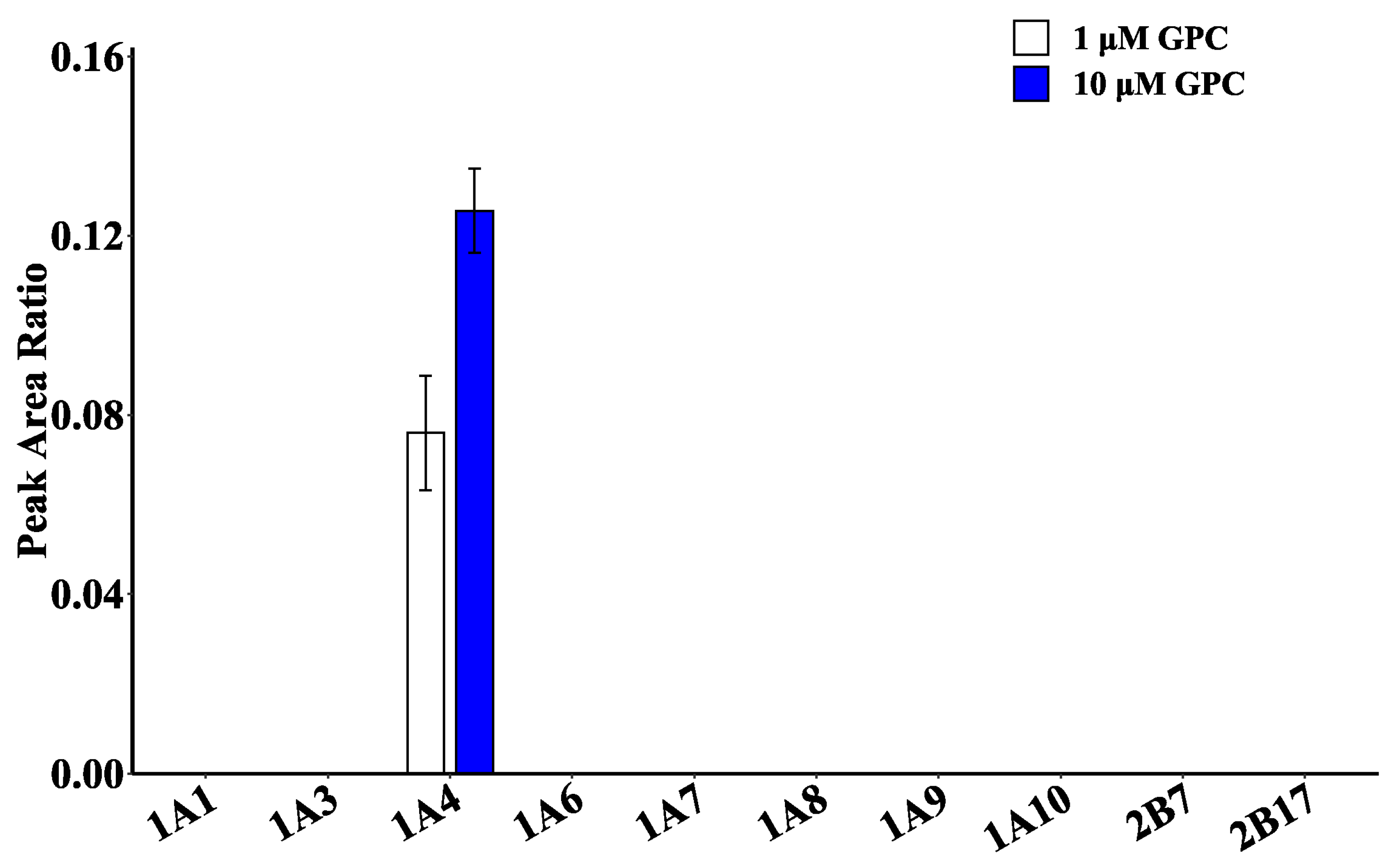
2.4. Glucuronidation Metabolism of GPC in Recombinant Human UGT Enzymes
2.5. Glucuronidation Metabolism of GPC among Different Species
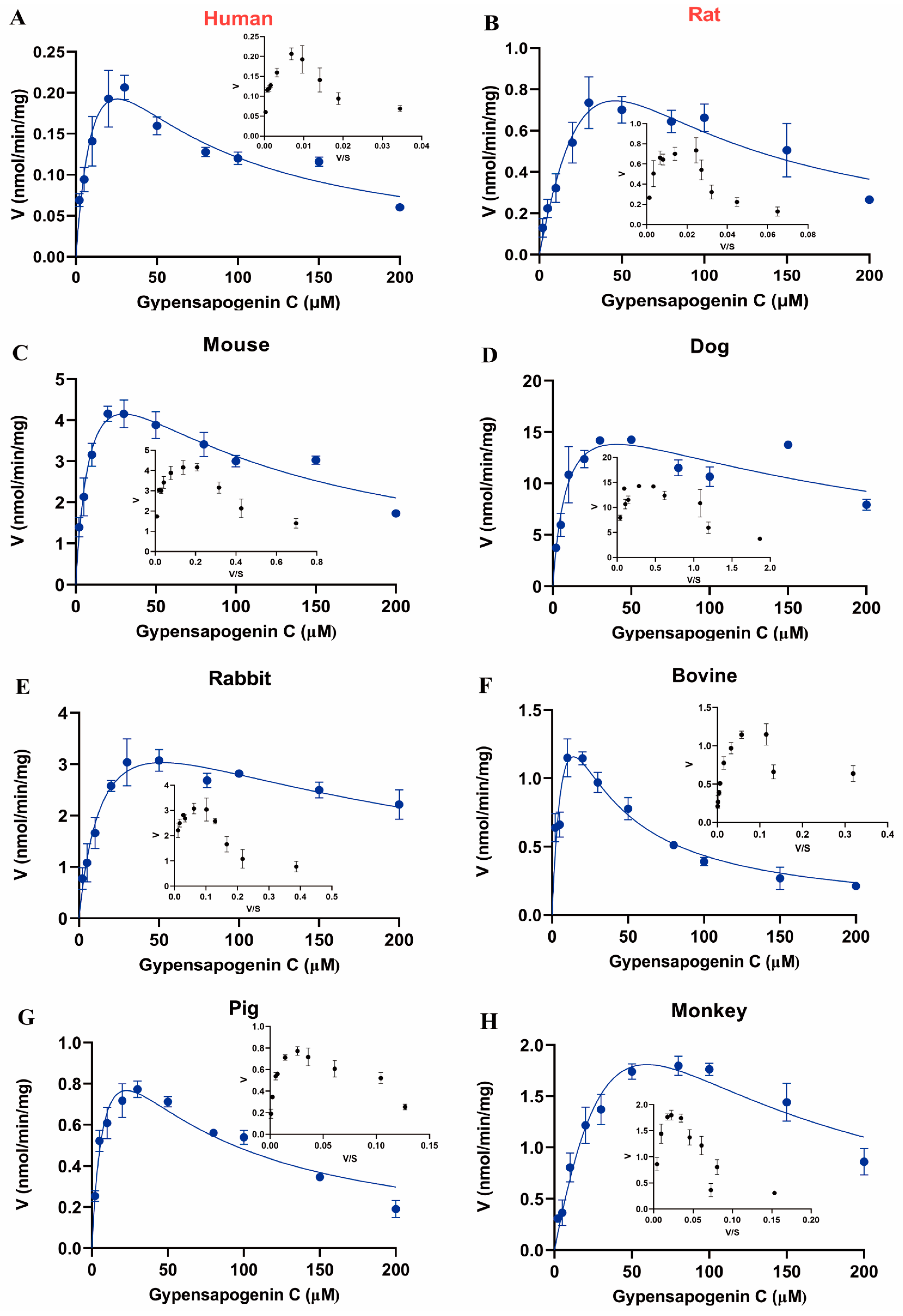
2.6. Enzymatic Kinetic Analysis of GPC in Liver Microsomes of Different Species
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Separation and Purification of GPC
4.3. Glucuronidation of GPC in Human Liver Microsomes
4.4. LC–MS/MS Analysis
4.5. Enzyme Kinetic Analysis
4.6. Chemical Inhibition Experiment
4.7. Recombinant Human UGT Enzyme Assay
4.8. Species Difference Analysis
4.9. Enzyme Kinetic Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, J. Flora of China. Harv. Pap. Bot. 2007, 13, 301–302. [Google Scholar] [CrossRef]
- Li, Y.; Lin, W.; Huang, J.; Xie, Y.; Ma, W. Anti-cancer effects of Gynostemma pentaphyllum (Thunb.) Makino (Jiaogulan). Chin. Med. 2016, 11, 43. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Ha, T.K.Q.; Yang, J.L.; Pham, H.T.T.; Oh, W.K. Triterpenoids from the genus Gynostemma: Chemistry and pharmacological activities. J. Ethnopharmacol. 2021, 268, 113574. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.; Lee, J.W.; Lee, C.; Jin, Q.; Lee, M.K.; Lee, C.K.; Lee, M.K.; Hwang, B.Y. Flavonol glycosides from the aerial parts of Gynostemma pentaphyllum and their antioxidant activity. Arch. Pharm. Res. 2016, 39, 1232–1236. [Google Scholar] [CrossRef]
- Yan, W.; Niu, Y.; Lv, J.; Xie, Z.; Jin, L.; Yao, W.; Gao, X.; Yu, L.L. Characterization of a heteropolysaccharide isolated from diploid Gynostemma pentaphyllum Makino. Carbohydr. Polym. 2013, 92, 2111–2117. [Google Scholar] [CrossRef]
- Hu, Y.; Ip, F.C.; Fu, G.; Pang, H.; Ye, W.; Ip, N.Y. Dammarane saponins from Gynostemma pentaphyllum. Phytochemistry 2010, 71, 1149–1157. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Li, N.; Ren, R.; Wang, Y.; Su, X.; Lu, F.; Zong, R.; Yang, L.; Ma, X. Progress in the Medicinal Value, Bioactive Compounds, and Pharmacological Activities of Gynostemma pentaphyllum. Molecules 2021, 26, 6249. [Google Scholar] [CrossRef]
- Ling, Y.; Lei, Z.; Xinying, L.; Yan, X.; Siying, W.; Yang, W.; Nie, Q.; Zhang, Q.; Hai’ou, B.; Yu, J.; et al. Characterization and Identification of the Chemical Constituents of Gynostemma pentaphyllum Using High Performance Liquid Chromatography–Electrospray Ionization–Quadrupole Time-of-Flight Tandem Mass Spectrometry (HPLC-ESI-QTOF-MS/MS). Anal. Lett. 2020, 53, 760–773. [Google Scholar] [CrossRef]
- Shen, Z.; Wang, Z.; Hou, H.; Zhang, L.; Huang, J. Studies on the Anti-oxidant Activity and Anti-aging Effect of Compound Granules from Red Ginseng and Sea Cucumber. Ginseng Res. 2020, 32, 59–64. [Google Scholar]
- Pu, X.; Wu, Q.; Yang, J.; Liu, H. A549 Cell Inhibitory Activity from Heat-Processed Gynostemma pentaphyllum. J. Minzu Univ. China (Nat. Sci. Ed.) 2012, 21, 49–53. [Google Scholar]
- Liu, J.; Zhang, L.; Ren, Y.; Gao, Y.; Kang, L.; Qiao, Q. Anticancer and immunoregulatory activity of Gynostemma pentaphyllum polysaccharides in H22 tumor-bearing mice. Int. J. Biol. Macromol. 2014, 69, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Gou, S.H.; Huang, H.F.; Chen, X.Y.; Liu, J.; He, M.; Ma, Y.Y.; Zhao, X.N.; Zhang, Y.; Ni, J.M. Lipid-lowering, hepatoprotective, and atheroprotective effects of the mixture Hong-Qu and gypenosides in hyperlipidemia with NAFLD rats. J. Chin. Med. Assoc. 2016, 79, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Zhao, M.; Qi, X.; Liu, Y.; Li, N.; Liu, Z.; Bian, Y. Hypoglycemic effect of Gynostemma pentaphyllum saponins by enhancing the Nrf2 signaling pathway in STZ-inducing diabetic rats. Arch. Pharm. Res. 2016, 39, 221–230. [Google Scholar] [CrossRef]
- Lin, C.C.; Huang, P.C.; Lin, J.M. Antioxidant and hepatoprotective effects of Anoectochilus formosanus and Gynostemma pentaphyllum. Am. J. Chin. Med. 2000, 28, 87–96. [Google Scholar] [CrossRef]
- Wu, L. In Vivo and In Vitro Metabolism Studies and Pharmacodynamic Evaluation of Rotundic Acid; South China University of Technology: Guangzhou, China, 2020. [Google Scholar]
- Lee, H.; Heo, J.K.; Lee, G.H.; Park, S.Y.; Jang, S.N.; Kim, H.J.; Kwon, M.J.; Song, I.S.; Liu, K.H. Ginsenoside Rc Is a New Selective UGT1A9 Inhibitor in Human Liver Microsomes and Recombinant Human UGT Isoforms. Drug Metab. Dispos. 2019, 47, 1372–1379. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.F.; He, R.R.; Cao, J.; Chen, J.X.; Huang, T.; Liu, Y. Drug-Drug Interactions Potential of Icariin and Its Intestinal Metabolites via Inhibition of Intestinal UDP-Glucuronosyltransferases. Evid. Based Complement. Altern. Med. 2012, 2012, 395912. [Google Scholar] [CrossRef] [PubMed]
- Oda, S.; Fukami, T.; Yokoi, T.; Nakajima, M. A comprehensive review of UDP-glucuronosyltransferase and esterases for drug development. Drug Metab. Pharmacokinet. 2015, 30, 30–51. [Google Scholar] [CrossRef]
- Zhang, L.; Lin, Y.; Guan, H.; Hu, L.; Pan, G. Simultaneous determination of gypenoside LVI, gypenoside XLVI, 2α-OH-protopanaxadiol and their two metabolites in rat plasma by LC-MS/MS and its application to pharmacokinetic studies. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2015, 1005, 9–16. [Google Scholar] [CrossRef]
- Chen, D.J.; Hu, H.G.; Xing, S.F.; Liu, H.M.; Piao, X.L. Metabolite profiling of gypenoside LVI in rat after oral and intravenous administration. Arch. Pharm. Res. 2015, 38, 1157–1167. [Google Scholar] [CrossRef]
- He, M.; Jiang, J.; Qiu, F.; Liu, S.; Peng, P.; Gao, C.; Miao, P. Inhibitory effects of gypenosides on seven human cytochrome P450 enzymes in vitro. Food Chem. Toxicol. 2013, 57, 262–265. [Google Scholar] [CrossRef]
- Zhuo, S.; Qin, H.; Chen, J.; Yang, Z. Effects of Gynostemma pentaphyllum on the cytochrome P450 and GST, UGT activities in liver of rat. Guihaia 2014, 34, 194–197. [Google Scholar]
- He, Y.Q.; Liu, Y.; Zhang, B.F.; Liu, H.X.; Lu, Y.L.; Yang, L.; Xiong, A.Z.; Xu, L.L.; Wang, C.H.; Yang, L.; et al. Identification of the UDP-glucuronosyltransferase isozyme involved in senecionine glucuronidation in human liver microsomes. Drug Metab. Dispos. 2010, 38, 626–634. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; He, C.; Fang, L.; Yang, L.; Wang, Z. Identification of Human UDP-Glucuronosyltransferase 1A4 as the Major Isozyme Responsible for the Glucuronidation of 20(S)-Protopanaxadiol in Human Liver Microsomes. Int. J. Mol. Sci. 2016, 17, 205. [Google Scholar] [CrossRef] [PubMed]
- Omura, K.; Motoki, K.; Kobashi, S.; Miyata, K.; Yamano, K.; Iwanaga, T. Identification of Human UDP-Glucuronosyltransferase and Sulfotransferase as Responsible for the Metabolism of Dotinurad, a Novel Selective Urate Reabsorption Inhibitor. Drug Metab. Dispos. 2021, 49, 1016–1024. [Google Scholar] [CrossRef]
- Uchaipichat, V.; Mackenzie, P.I.; Elliot, D.J.; Miners, J.O. Selectivity of substrate (trifluoperazine) and inhibitor (amitriptyline, androsterone, canrenoic acid, hecogenin, phenylbutazone, quinidine, quinine, and sulfinpyrazone) “probes” for human udp-glucuronosyltransferases. Drug Metab. Dispos. 2006, 34, 449–456. [Google Scholar] [CrossRef]
- Xin, H.; Xia, Y.L.; Hou, J.; Wang, P.; He, W.; Yang, L.; Ge, G.B.; Xu, W. Identification and characterization of human UDP-glucuronosyltransferases responsible for the in-vitro glucuronidation of arctigenin. J. Pharm. Pharmacol. 2015, 67, 1673–1681. [Google Scholar] [CrossRef]
- Lu, D.; Xie, Q.; Wu, B. N-glucuronidation catalyzed by UGT1A4 and UGT2B10 in human liver microsomes: Assay optimization and substrate identification. J. Pharm. Biomed. Anal. 2017, 145, 692–703. [Google Scholar] [CrossRef]
- Kaivosaari, S.; Finel, M.; Koskinen, M. N-glucuronidation of drugs and other xenobiotics by human and animal UDP-glucuronosyltransferases. Xenobiotica 2011, 41, 652–669. [Google Scholar] [CrossRef]
- Huang, K.; Que, L.; Ding, Y.; Chu, N.; Qian, Z.; Qin, W.; Chen, Y.; Zhang, J.; He, Q. Identification of human uridine diphosphate-glucuronosyltransferase isoforms responsible for the glucuronidation of 10,11-dihydro-10-hydroxy-carbazepine. J. Pharm. Pharmacol. 2021, 73, 388–397. [Google Scholar] [CrossRef]
- Ding, J.; Chen, X.; Gao, Z.; Dai, X.; Li, L.; Xie, C.; Jiang, H.; Zhang, L.; Zhong, D. Metabolism and pharmacokinetics of novel selective vascular endothelial growth factor receptor-2 inhibitor apatinib in humans. Drug Metab. Dispos. 2013, 41, 1195–1210. [Google Scholar] [CrossRef]
- Li, Y.; Xu, C.; Xu, J.; Qin, Z.; Li, S.; Hu, L.; Yao, Z.; Gonzalez, F.J.; Yao, X. Characterization of metabolic activity, isozyme contribution and species differences of bavachin, and identification of efflux transporters for bavachin-O-glucuronide in HeLa1A1 cells. J. Pharm. Pharmacol. 2020, 72, 1771–1786. [Google Scholar] [CrossRef] [PubMed]
- Ling, L.; Zeng, Y.; Lu, Y.; Qin, L.; Tan, D.; Du, Y.; Zhou, X.; He, Y. Species difference in chenodeoxycholic acid glucuronidation in liver microsomes. Chin. J. New Drugs 2019, 28, 1751–1758. [Google Scholar]
- Isobe, T.; Ohkawara, S.; Ochi, S.; Tanaka-Kagawa, T.; Jinno, H.; Hanioka, N. Naringenin glucuronidation in liver and intestine microsomes of humans, monkeys, rats, and mice. Food Chem. Toxicol. 2018, 111, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Gan, L.; Ma, J.; You, G.; Mai, J.; Wang, Z.; Yang, R.; Xie, C.; Fei, J.; Tang, L.; Zhao, J.; et al. Glucuronidation and its effect on the bioactivity of amentoflavone, a biflavonoid from Ginkgo biloba leaves. J. Pharm. Pharmacol. 2020, 72, 1840–1853. [Google Scholar] [CrossRef]
- Li, N.; Wu, C.F.; Xu, X.Y.; Liu, Z.Y.; Li, X.; Zhao, Y.Q. Triterpenes possessing an unprecedented skeleton isolated from hydrolyzate of total saponins from Gynostemma pentaphyllum. Eur. J. Med. Chem. 2012, 50, 173–178. [Google Scholar] [CrossRef]
- Nardone-White, D.T.; Bissada, J.E.; Abouda, A.A.; Jackson, K.D. Detoxication versus Bioactivation Pathways of Lapatinib In Vitro: UGT1A1 Catalyzes the Hepatic Glucuronidation of Debenzylated Lapatinib. Drug Metab. Dispos. 2021, 49, 233–244. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme Sources | Km (μM) | Vmax (nmol/min/mg) | Ki (μM) | CLint (μL/min/mg) |
|---|---|---|---|---|
| HLMs | 15.36 ± 5.26 | 0.42 ± 0.09 | 42.68 ± 13.63 | 27 |
| RLMs | 94.07 ± 89.75 | 3.80 ± 3.01 | 22.44 ± 21.19 | 40 |
| MLMs | 10.04 ± 2.10 | 6.93 ± 0.69 | 89.46 ± 17.97 | 690 |
| DLMs | 9.56 ± 2.83 | 20.28 ± 2.57 | 173.90 ± 57.45 | 2121 |
| RaLMs | 16.19 ± 3.77 | 4.92 ± 0.57 | 167.50 ± 45.87 | 304 |
| BLMs | 12.24 ± 5.16 | 3.17± 0.89 | 16.21 ± 6.24 | 259 |
| PLMs | 9.58 ± 2.32 | 1.41 ± 0.18 | 54.32 ± 11.96 | 147 |
| MkLMs | 107.10 ± 73.18 | 8.29 ± 4.69 | 33.34 ± 23.44 | 77 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Qin, L.; Wu, X.; Tan, D.; Lu, Y.; Du, Y.; Wu, D.; He, Y. Identification of Human UDP-Glucuronosyltransferase Involved in Gypensapogenin C Glucuronidation and Species Differences. Int. J. Mol. Sci. 2023, 24, 1454. https://doi.org/10.3390/ijms24021454
Chen J, Qin L, Wu X, Tan D, Lu Y, Du Y, Wu D, He Y. Identification of Human UDP-Glucuronosyltransferase Involved in Gypensapogenin C Glucuronidation and Species Differences. International Journal of Molecular Sciences. 2023; 24(2):1454. https://doi.org/10.3390/ijms24021454
Chicago/Turabian StyleChen, Juan, Lin Qin, Xingdong Wu, Daopeng Tan, Yanliu Lu, Yimei Du, Di Wu, and Yuqi He. 2023. "Identification of Human UDP-Glucuronosyltransferase Involved in Gypensapogenin C Glucuronidation and Species Differences" International Journal of Molecular Sciences 24, no. 2: 1454. https://doi.org/10.3390/ijms24021454
APA StyleChen, J., Qin, L., Wu, X., Tan, D., Lu, Y., Du, Y., Wu, D., & He, Y. (2023). Identification of Human UDP-Glucuronosyltransferase Involved in Gypensapogenin C Glucuronidation and Species Differences. International Journal of Molecular Sciences, 24(2), 1454. https://doi.org/10.3390/ijms24021454