
Flavonoids as Modulators of Potassium Channels
 , and
, and 
Abstract
1. Introduction
2. Kv Channels
2.1. Kv1.3 Channel
2.2. Kv1.5 Channel
2.3. Kv2.1 Channel
2.4. Kv4 Channels
2.5. hERG Channels

{kind=link}
{kind=link}
| Flavonoid | Type of Cell | Effect | References | |
|---|---|---|---|---|
| Acacetin | HEK 293 | ↓ | Li et al. [42] (2008) | |
| Apigenin | Xenopus oocyte | → | Zitron et al. [82] (2005) | |
| Chrysin | Xenopus oocyte | → | Zitron et al. [82] (2005) | |
| Daidzein | HEK 293 | ↓ | Zhang et al. [85] (2008) | |
| 7,8-Dimethoxyflavone | Xenopus oocyte | ↓ | Du et al. [86] (2015) | |
| (−)-Epigallocatechin gallate | HEK 293 | ↓ | Kelemen et al. [87] (2007) | |
| Xenopus oocyte | ↓ | |||
| CHO | ↓ | Kang et al. [44] (2010) | ||
| Fisetin | Xenopus oocyte | → | Zitron et al. [82] (2005) | |
| HEK 293 | ↓ | Sun et al. [88] (2017) | ||
| Flavone | Xenopus oocyte | ↓ | Zitron et al. [82] (2005) | |
| Galangin | Xenopus oocyte | → | Zitron et al. [82] (2005) | |
| HEK 293 | ↓ | Sun et al. [88] (2017) | ||
| Genistein | HEK 293 | ↓ | Zhang et al. [85] (2008) | |
| Hesperetin | Xenopus oocyte | ↓ | Zitron et al. [82] (2005) | |
| ↓ | Scholz et al. [89] (2007) | |||
| Hesperidin | Xenopus oocyte | ↑ | Zitron et al. [82] (2005) | |
| Isorhamnetin | HEK 293 | ↓ | Sun et al. [88] (2017) | |
| Kaempferol | Xenopus oocyte | ↓ | Zitron et al. [82] (2005) | |
| HEK 293 | ↓ | Sun et al. [90] (2017) | ||
| Liquiritigenin | CHL | ↓ | Sweeney et al. [91] (2019) | |
| Luteolin | HEK 293 | ↓ | Sun et al. [88] (2017) | |
| 7,8-Methylenedioxyflavone | Xenopus oocyte | ↓ | Du et al. [86] (2015) | |
| Morin | Xenopus oocyte | ↓ | Zitron et al. [82] (2005) | |
| HEK 293 | ↓ | Sun et al. [88] (2017) | ||
| Myricetin | Xenopus oocyte | ↑ | Zitron et al. [82] (2005) | |
| HEK 293 | → | Sun et al. [88] (2017) | ||
| Naringenin | Xenopus oocyte | ↓ | Scholz et al. [83] (2005) | |
| ↓ | Lin et al. [84] (2008) | |||
| ↓ | Zitron et al. [82] (2005) | |||
| HEK 293 | ↓ | |||
| CHO | ↓ | Sanson et al.[45] (2022) | ||
| Naringin | Xenopus oocyte | → | Zitron et al. [82] (2005) | |
| Neohesperidin | Xenopus oocyte | → | Zitron et al. [82] (2005) | |
| Quercetin | Xenopus oocyte | ↓ | Zitron et al. [82] (2005) | |
| HEK 293 | ↓ | Sun et al. [88] (2017) | ||
| Rutin | Xenopus oocyte | → | Zitron et al. [82] (2005) | |
| HEK 293 | → | Sun et al. [88] (2017) | ||
| Taxifolin | HEK 293 | → | Sun et al. [88] (2017) | |
| Taxifolin 3-O--D-glucopyranoside | CHO | ↓ | Yun et al. [92] (2013) | |
| Trimethylapigenin | HEK 293 | ↓ | 18–32 | Liu et al. [38] (2012) |
2.6. Further Kv Channels
3. Calcium-Activated Channels (KCa)
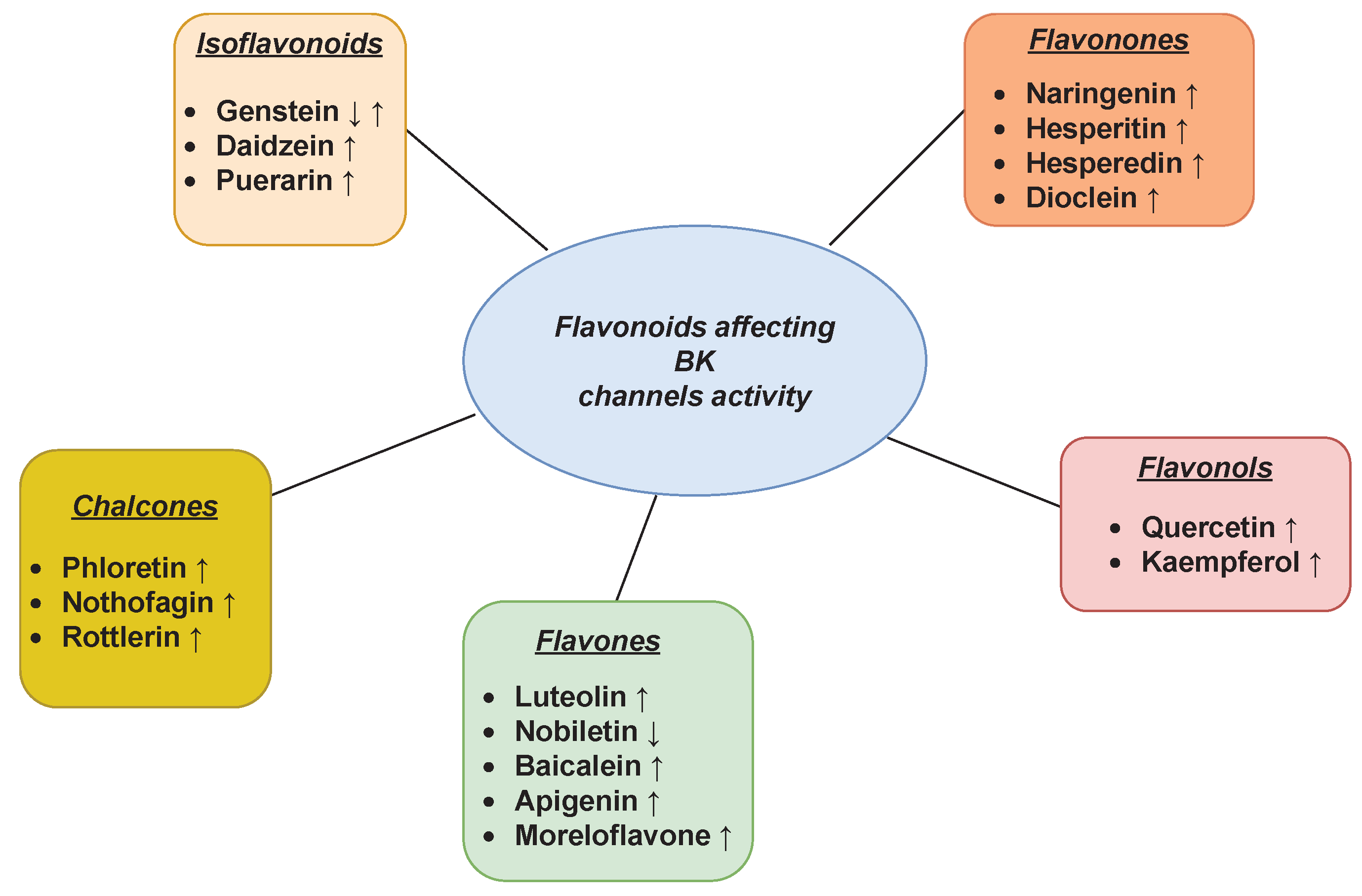
3.1. BK Channel
3.2. IK and SK Channels
4. Inward Rectifying Potassium Channels (Kir)
5. Two-Pore Domain Potassium Channels (K2P)
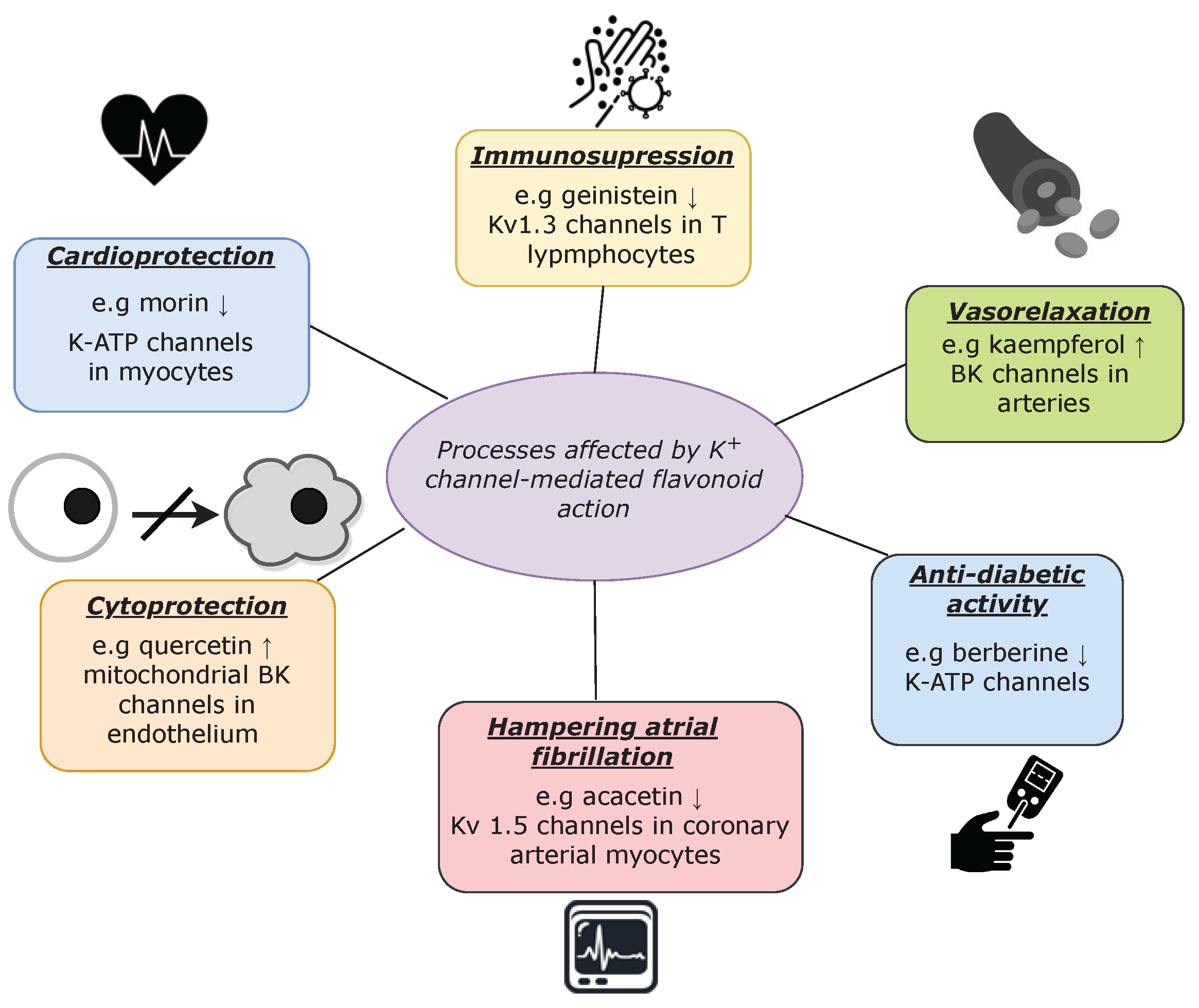
6. Discussion
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AF | atrial fibrillation |
| AP | action potential |
| BK | big-conductance (large-conductance) -dependent potassium channels |
| CNS | central nervous system |
| half maximal effective concentration | |
| EGCG | (−)-epigallocatechin-3-gallate |
| inhibitory concentration 50% | |
| IK | intermediate-conductance -dependent potassium channels |
| ultra-rapid delayed rectifier current | |
| K2P | two-pore domain potassium channels |
| KCa | -regulated potassium channels |
| Kv | voltage-regulated (voltage-gated) potassium channels |
| Kir | inward rectifier potassium channels |
| mitoKATP | mitochondrial ATP-regulated channels |
| mitoBK | mitochondrial BK channels |
| PH | pulmonary hypertension |
| SK | small-conductance -dependent potassium channels |
| T2D | type 2 diabetes |
| TMDs | transmembrane domains |
| TALK | TWIK-related alkaline pH-activated channels |
| TASK | TWIK-related acid-sensitive channel channels |
| THIK | tandem pore domain halothane-inhibited channels |
| TRAAK | TWIK-related arachidonic acid-activated potassium channel |
| TREK | TWIK-related channels |
| TRIK | TWIK-related spinal cord channel |
| TWIK | tandem of pore domains in a weak inward rectifying channels |
References
- Hille, B. Ionic channels in excitable membranes. Current problems and biophysical approaches. Biophys. J. 1978, 22, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Coetzee, W.A.; Amarillo, Y.; Chiu, J.; Chow, A.; Lau, D.; McCormack, T.; Morena, H.; Nadal, M.S.; Ozaita, A.; Pountney, D.; et al. Molecular diversity of K+ channels. Ann. N. Y. Acad. Sci. 1999, 868, 233–255. [Google Scholar] [CrossRef] [PubMed]
- Miller, C. An overview of the potassium channel family. Genome Biol. 2000, 1, reviews0004.1. [Google Scholar] [CrossRef] [PubMed]
- MacKinnon, R. Potassium channels. FEBS Lett. 2003, 555, 62–65. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Q.; Purhonen, P.; Hebert, H. Structure of potassium channels. Cell. Mol. Life Sci. 2015, 72, 3677–3693. [Google Scholar] [CrossRef]
- Wulff, H.; Castle, N.A.; Pardo, L.A. Voltage-gated potassium channels as therapeutic targets. Nat. Rev. Drug Discov. 2009, 8, 982–1001. [Google Scholar] [CrossRef]
- Hutchings, C.J.; Colussi, P.; Clark, T.G. Ion channels as therapeutic antibody targets. Proc. Mabs. Taylor Fr. 2019, 11, 265–296. [Google Scholar] [CrossRef]
- Mathie, A.; Veale, E.L.; Cunningham, K.P.; Holden, R.G.; Wright, P.D. Two-pore domain potassium channels as drug targets: Anesthesia and beyond. Annu. Rev. Pharmacol. Toxicol. 2021, 61, 401–420. [Google Scholar] [CrossRef]
- Cui, M.; Cantwell, L.; Zorn, A.; Logothetis, D.E. Kir Channel Molecular Physiology, Pharmacology, and Therapeutic Implications. Pharmacol. Potassium Channels 2021, 267, 277–356. [Google Scholar]
- Dudem, S.; Sergeant, G.P.; Thornbury, K.D.; Hollywood, M.A. Calcium-activated K+ channels (KCa) and therapeutic implications. In Pharmacology of Potassium Channels; Springer: Berlin, Germany, 2021; pp. 379–416. [Google Scholar]
- Zúñiga, L.; Cayo, A.; González, W.; Vilos, C.; Zúñiga, R. Potassium Channels as a Target for Cancer Therapy: Current Perspectives. Oncotargets Ther. 2022, 15, 783–797. [Google Scholar] [CrossRef]
- Wulff, H.; Christophersen, P.; Colussi, P.; Chandy, K.G.; Yarov-Yarovoy, V. Antibodies and venom peptides: New modalities for ion channels. Nat. Rev. Drug Discov. 2019, 18, 339–357. [Google Scholar] [CrossRef] [PubMed]
- Adorisio, S.; Fierabracci, A.; Rossetto, A.; Muscari, I.; Nardicchi, V.; Liberati, A.M.; Riccardi, C.; Van Sung, T.; Thuy, T.T.; Delfino, D.V. Integration of traditional and western medicine in Vietnamese populations: A review of health perceptions and therapies. Nat. Prod. Commun. 2016, 11, 1409–1416. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Ma, Q.; Ye, L.; Piao, G. The traditional medicine and modern medicine from natural products. Molecules 2016, 21, 559. [Google Scholar] [CrossRef]
- Georgiev, M.I. From plants to pharmacy shelf: Natural products revival. Phytochem. Rev. 2016, 15, 511–513. [Google Scholar] [CrossRef]
- Gechev, T.S.; Hille, J.; Woerdenbag, H.J.; Benina, M.; Mehterov, N.; Toneva, V.; Fernie, A.R.; Mueller-Roeber, B. Natural products from resurrection plants: Potential for medical applications. Biotechnol. Adv. 2014, 32, 1091–1101. [Google Scholar] [CrossRef] [PubMed]
- Monjotin, N.; Amiot, M.J.; Fleurentin, J.; Morel, J.M.; Raynal, S. Clinical evidence of the benefits of phytonutrients in human healthcare. Nutrients 2022, 14, 1712. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Guo, M.; Lim, S.S. Recent advances in recognition of bioactive phytonutrients for specific targets in plant foods. Front. Nutr. 2022, 9, 1018946. [Google Scholar] [CrossRef]
- Muscari, I.; Adorisio, S.; Thuy, T.T.; Van Sung, T.; Delfino, D.V. Recent Insights on the Role of Natural Medicines in Immunostimulation. In Natural Medicines; CRC Press: Boca Raton, FL, USA, 2019; pp. 349–360. [Google Scholar]
- Heinrich, M.; Barnes, J.; Prieto-Garcia, J.; Gibbons, S.; Williamson, E.M. Natural product chemistry. In Fundamentals of Pharmacognosy and Phytotherapy; Elsevier Health Sciences: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef]
- Ranjan, R.; Logette, E.; Marani, M.; Herzog, M.; Tâche, V.; Scantamburlo, E.; Buchillier, V.; Markram, H. A kinetic map of the homomeric voltage-gated potassium channel (Kv) family. Front. Cell Neurosci. 2019, 13, 358. [Google Scholar] [CrossRef] [PubMed]
- González, C.; Baez-Nieto, D.; Valencia, I.; Oyarzún, I.; Rojas, P.; Naranjo, D.; Latorre, R. K+ channels: Function-structural overview. Compr. Physiol. 2012, 2, 2087–2149. [Google Scholar] [PubMed]
- Teisseyre, A.; Michalak, K. Genistein inhibits the activity of Kv1. 3 potassium channels in human T lymphocytes. J. Membr. Biol. 2005, 205, 71–79. [Google Scholar] [CrossRef]
- Teisseyre, A.; Palko-Labuz, A.; Uryga, A.; Michalak, K. The influence of 6-Prenylnaringenin and selected non-prenylated flavonoids on the activity of Kv1. 3 channels in human Jurkat T cells. J. Membr. Biol. 2018, 251, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Dong, Q.; Fu, X.X.; Du, L.L.; Cheng, X.; Du, Y.M.; Liao, Y.H. Acacetin blocks kv1.3 channels and inhibits human T cell activation. Cell. Physiol. Biochem. 2014, 34, 1359–1372. [Google Scholar] [CrossRef]
- Teisseyre, A.; Chmielarz, M.; Uryga, A.; Środa Pomianek, K.; Palko-Łabuz, A. Co-Application of Statin and Flavonoids as an Effective Strategy to Reduce the Activity of Voltage-Gated Potassium Channels Kv1.3 and Induce Apoptosis in Human Leukemic T Cell Line Jurkat. Molecules 2022, 27, 3227. [Google Scholar] [CrossRef]
- Teisseyre, A.; Michalak, K. Inhibition of the activity of human lymphocyte Kv1.3 potassium channels by resveratrol. J. Membr. Biol. 2006, 214, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Teisseyre, A.; Duarte, N.; Ferreira, M.J.U.; Michalak, K. Influence of the multidrug transporter inhibitors on the activity of Kv1.3 voltage-gated potassium channels. Acta Physiol. Pol. 2009, 60, 69. [Google Scholar]
- Gąsiorowska, J.; Teisseyre, A.; Uryga, A.; Michalak, K. Inhibition of Kv1.3 channels in human Jurkat T cells by xanthohumol and isoxanthohumol. J. Membr. Biol. 2015, 248, 705–711. [Google Scholar] [CrossRef]
- Gasiorowska, J.; Teisseyre, A.; Uryga, A.; Michalak, K. The influence of 8-prenylnaringenin on the activity of voltage-gated kv1.3 potassium channels in human jurkat t cells. Cell. Mol. Biol. Lett. 2012, 17, 559–570. [Google Scholar] [CrossRef]
- Phan, H.T.L.; Kim, H.J.; Jo, S.; Kim, W.K.; Namkung, W.; Nam, J.H. Anti-Inflammatory Effect of Licochalcone A via Regulation of ORAI1 and K+ Channels in T-Lymphocytes. Int. J. Mol. Sci. 2021, 22, 10847. [Google Scholar] [CrossRef] [PubMed]
- Ou, X.; Bin, X.; Wang, L.; Li, M.; Yang, Y.; Fan, X.; Zeng, X. Myricetin inhibits Kv1.5 channels in HEK293 cells. Mol. Med. Rep. 2016, 13, 1725–1731. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, H.f.; Wang, C.; Chen, Y.f.; Ma, R.; Xiang, J.z.; Du, X.l.; Tang, Q. Inhibitory effects of hesperetin on Kv1.5 potassium channels stably expressed in HEK 293 cells and ultra-rapid delayed rectifier K+ current in human atrial myocytes. Eur. J. Pharmacol. 2016, 789, 98–108. [Google Scholar] [CrossRef]
- Yang, L.; Ma, J.H.; Zhang, P.H.; Zou, A.R.; Tu, D.N. Quercetin activates human Kv1.5 channels by a residue I502 in the S6 segment. Clin. Exp. Pharmacol. Physiol. 2009, 36, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Morales-Cano, D.; Menendez, C.; Moreno, E.; Moral-Sanz, J.; Barreira, B.; Galindo, P.; Pandolfi, R.; Jimenez, R.; Moreno, L.; Cogolludo, A.; et al. The flavonoid quercetin reverses pulmonary hypertension in rats. PLoS ONE 2014, 9, e114492. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, X.H.; Liu, Z.; Du, X.L.; Chen, K.H.; Xin, X.; Jin, Z.D.; Shen, J.Z.; Hu, Y.; Li, G.R.; et al. Effects of the natural flavone trimethylapigenin on cardiac potassium currents. Biochem. Pharmacol. 2012, 84, 498–506. [Google Scholar] [CrossRef]
- Choi, B.H.; Choi, J.S.; Yoon, S.H.; Rhie, D.J.; Jo, Y.H.; Kim, M.S.; Hahn, S.J. Effects of (−)-epigallocatechin-3-gallate, the main component of green tea, on the cloned rat brain Kv1.5 potassium channels. Biochem. Pharmacol. 2001, 62, 527–535. [Google Scholar] [CrossRef]
- Noguchi, C.; Yang, J.; Sakamoto, K.; Maeda, R.; Takahashi, K.; Takasugi, H.; Ono, T.; Murakawa, M.; Kimura, J. Inhibitory effects of isoliquiritigenin and licorice extract on voltage-dependent K+ currents in H9c2 cells. J. Pharmacol. Sci. 2008, 108, 439–445. [Google Scholar] [CrossRef]
- Wu, H.J.; Wu, W.; Sun, H.Y.; Qin, G.W.; Wang, H.B.; Wang, P.; Yalamanchili, H.K.; Wang, J.; Tse, H.F.; Lau, C.P.; et al. Acacetin causes a frequency-and use-dependent blockade of hKv1.5 channels by binding to the S6 domain. J. Mol. Cell. Cardiol. 2011, 51, 966–973. [Google Scholar] [CrossRef]
- Li, G.R.; Wang, H.B.; Qin, G.W.; Jin, M.W.; Tang, Q.; Sun, H.Y.; Du, X.L.; Deng, X.L.; Zhang, X.H.; Chen, J.B.; et al. Acacetin, a natural flavone, selectively inhibits human atrial repolarization potassium currents and prevents atrial fibrillation in dogs. Circulation 2008, 117, 2449–2457. [Google Scholar] [CrossRef]
- Xu, H.; Zhao, M.; Liang, S.; Huang, Q.; Xiao, Y.; Ye, L.; Wang, Q.; He, L.; Ma, L.; Zhang, H.; et al. The effects of puerarin on rat ventricular myocytes and the potential mechanism. Sci. Rep. 2016, 6, 35475. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Cheng, H.; Ji, J.; Incardona, J.; Rampe, D. In vitro electrocardiographic and cardiac ion channel effects of (−)-epigallocatechin-3-gallate, the main catechin of green tea. J. Pharmacol. Exp. Ther. 2010, 334, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Sanson, C.; Boukaiba, R.; Houtmann, S.; Maizières, M.A.; Fouconnier, S.; Partiseti, M.; Bohme, G.A. The grapefruit polyphenol naringenin inhibits multiple cardiac ion channels. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2022, 395, 735–740. [Google Scholar] [CrossRef] [PubMed]
- Aréchiga-Figueroa, I.A.; Morán-Zendejas, R.; Delgado-Ramírez, M.; Rodríguez-Menchaca, A.A. Phytochemicals genistein and capsaicin modulate Kv2.1 channel gating. Pharmacol. Rep. 2017, 69, 1145–1153. [Google Scholar] [CrossRef] [PubMed]
- Gu, R.R.; Meng, X.H.; Zhang, Y.; Xu, H.Y.; Zhan, L.; Gao, Z.B.; Yang, J.L.; Zheng, Y.M. (−)-Naringenin 4′,7-dimethyl Ether Isolated from Nardostachys jatamansi Relieves Pain through Inhibition of Multiple Channels. Molecules 2022, 27, 1735. [Google Scholar] [PubMed]
- Kim, H.J.; Ahn, H.S.; Choi, B.H.; Hahn, S.J. Inhibition of Kv4.3 by genistein via a tyrosine phosphorylation-independent mechanism. Am. J. Physiol. Cell Physiol. 2011, 300, C567–C575. [Google Scholar] [CrossRef] [PubMed]
- Na, W.; Ma, B.; Shi, S.; Chen, Y.; Zhang, H.; Zhan, Y.; An, H. Procyanidin B1, a novel and specific inhibitor of Kv10.1 channel, suppresses the evolution of hepatoma. Biochem. Pharmacol. 2020, 178, 114089. [Google Scholar] [CrossRef]
- Cahalan, M.D.; Chandy, K.G. The functional network of ion channels in T lymphocytes. Immunol. Rev. 2009, 231, 59–87. [Google Scholar] [CrossRef]
- DeCoursey, T.E.; Chandy, K.G.; Gupta, S.; Cahalan, M.D. Voltage-gated K+ channels in human T lymphocytes: A role in mitogenesis? Nature 1984, 307, 465–468. [Google Scholar] [CrossRef]
- Panyi, G.; Possani, L.; De La Vega, R.R.; Gaspar, R.; Varga, Z. K+ channel blockers: Novel tools to inhibit T cell activation leading to specific immunosuppression. Curr. Pharm. Des. 2006, 12, 2199–2220. [Google Scholar] [CrossRef]
- Teisseyre, A.; Gąsiorowska, J.; Michalak, K. Voltage-Gated Potassium Channels Kv1.3–Potentially New Molecular Target in Cancer Diagnostics and Therapy. Adv. Clin. Exp. Med. Off. Organ Wroc. Med. Univ. 2015, 24, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Li, W.; Liu, F. Inhibition of proliferation and induction of apoptosis by genistein in colon cancer HT-29 cells. Cancer Lett. 2004, 215, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Teisseyre, A.; Uryga, A.; Michalak, K. Statins as inhibitors of voltage-gated potassium channels Kv1.3 in cancer cells. J. Mol. Struct. 2021, 1230, 129905. [Google Scholar] [CrossRef]
- Baell, J.B.; Gable, R.W.; Harvey, A.J.; Toovey, N.; Herzog, T.; Hänsel, W.; Wulff, H. Khellinone derivatives as blockers of the voltage-gated potassium channel Kv1.3: Synthesis and immunosuppressive activity. J. Med. Chem. 2004, 47, 2326–2336. [Google Scholar] [CrossRef] [PubMed]
- Cianci, J.; Baell, J.B.; Flynn, B.L.; Robert, W.G.; Mould, J.A.; Paul, D.; Harvey, A.J. Synthesis and biological evaluation of chalcones as inhibitors of the voltage-gated potassium channel Kv1.3. Bioorg. Med. Chem. Lett. 2008, 18, 2055–2061. [Google Scholar] [CrossRef] [PubMed]
- Tamkun, M.M.; Knoth, K.M.; Walbridge, J.A.; Kroemer, H.; Roden, D.M.; Glover, D.M. Molecular cloning and characterization of two voltage-gated K+ channel cDNAs from human ventricle. FASEB J. 1991, 5, 331–337. [Google Scholar] [CrossRef]
- Overturf, K.E.; Russell, S.N.; Carl, A.; Vogalis, F.; Hart, P.; Hume, J.; Sanders, K.; Horowitz, B. Cloning and characterization of a Kv1.5 delayed rectifier K+ channel from vascular and visceral smooth muscles. Am. J. Physiol. Cell Physiol. 1994, 267, C1231–C1238. [Google Scholar] [CrossRef]
- Wang, Z.; Fermini, B.; Nattel, S. Sustained depolarization-induced outward current in human atrial myocytes. Evidence for a novel delayed rectifier K+ current similar to Kv1.5 cloned channel currents. Circ. Res. 1993, 73, 1061–1076. [Google Scholar] [CrossRef]
- Fedida, D.; Wible, B.; Wang, Z.; Fermini, B.; Faust, F.; Nattel, S.; Brown, A. Identity of a novel delayed rectifier current from human heart with a cloned K+ channel current. Circ. Res. 1993, 73, 210–216. [Google Scholar] [CrossRef]
- Wettwer, E.; Hála, O.; Christ, T.; Heubach, J.F.; Dobrev, D.; Knaut, M.; Varró, A.; Ravens, U. Role of I Kur in controlling action potential shape and contractility in the human atrium: Influence of chronic atrial fibrillation. Circulation 2004, 110, 2299–2306. [Google Scholar] [CrossRef]
- Christophersen, I.E.; Olesen, M.S.; Liang, B.; Andersen, M.N.; Larsen, A.P.; Nielsen, J.B.; Haunsø, S.; Olesen, S.P.; Tveit, A.; Svendsen, J.H.; et al. Genetic variation in KCNA5: Impact on the atrial-specific potassium current I Kur in patients with lone atrial fibrillation. Eur. Heart J. 2013, 34, 1517–1525. [Google Scholar] [CrossRef] [PubMed]
- Brendel, J.; Peukert, S. Blockers of the Kv1.5 channel for the treatment of atrial arrhythmias. Expert Opin. Ther. Patents 2002, 12, 1589–1598. [Google Scholar] [CrossRef]
- Tamargo, J.; Caballero, R.; Gómez, R.; Delpón, E. IKur/Kv1.5 channel blockers for the treatment of atrial fibrillation. Expert Opin. Investig. Drugs 2009, 18, 399–416. [Google Scholar] [CrossRef]
- Ford, J.W.; Milnes, J.T. New drugs targeting the cardiac ultra-rapid delayed-rectifier current (IKur): Rationale, pharmacology and evidence for potential therapeutic value. J. Cardiovasc. Pharmacol. 2008, 52, 105–120. [Google Scholar] [CrossRef] [PubMed]
- Van Wagoner, D.R.; Pond, A.L.; McCarthy, P.M.; Trimmer, J.S.; Nerbonne, J.M. Outward K+ current densities and Kv1.5 expression are reduced in chronic human atrial fibrillation. Circ. Res. 1997, 80, 772–781. [Google Scholar] [CrossRef]
- He, Y.; Fang, X.; Shi, J.; Li, X.; Xie, M.; Liu, X. Apigenin attenuates pulmonary hypertension by inducing mitochondria-dependent apoptosis of PASMCs via inhibiting the hypoxia inducible factor 1α–KV1.5 channel pathway. Chem. Biol. Interact. 2020, 317, 108942. [Google Scholar] [CrossRef]
- Murakoshi, H.; Trimmer, J.S. Identification of the Kv2.1 K+ channel as a major component of the delayed rectifier K+ current in rat hippocampal neurons. J. Neurosci. 1999, 19, 1728–1735. [Google Scholar] [CrossRef]
- Malin, S.A.; Nerbonne, J.M. Delayed rectifier K+ currents, IK, are encoded by Kv2 α-subunits and regulate tonic firing in mammalian sympathetic neurons. J. Neurosci. 2002, 22, 10094–10105. [Google Scholar] [CrossRef]
- Trimmer, J.S. Immunological identification and characterization of a delayed rectifier K+ channel polypeptide in rat brain. Proc. Natl. Acad. Sci. USA 1991, 88, 10764–10768. [Google Scholar] [CrossRef]
- Misonou, H.; Mohapatra, D.P.; Trimmer, J.S. Kv2.1: A voltage-gated k+ channel critical to dynamic control of neuronal excitability. Neurotoxicology 2005, 26, 743–752. [Google Scholar] [CrossRef]
- Roe, M.W.; Worley, J.F.; Mittal, A.A.; Kuznetsov, A.; DasGupta, S.; Mertz, R.J.; Witherspoon, S.M.; Blair, N.; Lancaster, M.E.; McIntyre, M.S.; et al. Expression and function of pancreatic β-cell delayed rectifier K+ channels: Role in stimulus-secretion coupling. J. Biol. Chem. 1996, 271, 32241–32246. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, T.J.; Tsaur, M.L.; Lopez, G.A.; Jan, Y.N.; Jan, L.Y. Characterization of a mammalian cDNA for an inactivating voltage-sensitive K+ channel. Neuron 1991, 7, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Jerng, H.H.; Pfaffinger, P.J.; Covarrubias, M. Molecular physiology and modulation of somatodendritic A-type potassium channels. Mol. Cell. Neurosci. 2004, 27, 343–369. [Google Scholar] [CrossRef] [PubMed]
- Pak, M.D.; Baker, K.; Covarrubias, M.; Butler, A.; Ratcliffe, A.; Salkoff, L. mShal, a subfamily of A-type K+ channel cloned from mammalian brain. Proc. Natl. Acad. Sci. USA 1991, 88, 4386–4390. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zhang, C.; Ye, T.; Chen, X.; Liu, X.; Chen, X.; Sun, Y.; Qu, C.; Liang, J.; Shi, S.; et al. Pinocembrin ameliorates arrhythmias in rats with chronic ischaemic heart failure. Ann. Med. 2021, 53, 830–840. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wan, W.; Ran, Q.; Ye, T.; Sun, Y.; Liu, Z.; Liu, X.; Shi, S.; Qu, C.; Zhang, C.; et al. Pinocembrin mediates antiarrhythmic effects in rats with isoproterenol-induced cardiac remodeling. Eur. J. Pharmacol. 2022, 920, 174799. [Google Scholar] [CrossRef] [PubMed]
- Haverkamp, W.; Breithardt, G.; Camm, A.J.; Janse, M.J.; Rosen, M.R.; Antzelevitch, C.; Escande, D.; Franz, M.; Malik, M.; Moss, A.; et al. The potential for QT prolongation and pro-arrhythmia by non-anti-arrhythmic drugs: Clinical and regulatory implications: Report on a Policy Conference of the European Society of Cardiology. Cardiovasc. Res. 2000, 47, 219–233. [Google Scholar] [CrossRef]
- Taglialatela, M.; Castaldo, P.; Pannaccione, A.; Giorgio, G.; Annunziato, L. Human ether-a-gogo related gene (HERG) K channels as pharmacological targets: Present and future implications. Biochem. Pharmacol. 1998, 55, 1741–1746. [Google Scholar] [CrossRef]
- Saponara, S.; Fusi, F.; Iovinelli, D.; Ahmed, A.; Trezza, A.; Spiga, O.; Sgaragli, G.; Valoti, M. Flavonoids and hERG channels: Friends or foes? Eur. J. Pharmacol. 2021, 899, 174030. [Google Scholar] [CrossRef]
- Zitron, E.; Scholz, E.; Owen, R.W.; Luück, S.; Kiesecker, C.; Thomas, D.; Kathoöfer, S.; Niroomand, F.; Kiehn, J.; Kreye, V.A.; et al. QTc prolongation by grapefruit juice and its potential pharmacological basis: HERG channel blockade by flavonoids. Circulation 2005, 111, 835–838. [Google Scholar] [CrossRef]
- Scholz, E.P.; Zitron, E.; Kiesecker, C.; Lück, S.; Thomas, D.; Kathöfer, S.; Kreye, V.A.; Katus, H.A.; Kiehn, J.; Schoels, W.; et al. Inhibition of cardiac HERG channels by grapefruit flavonoid naringenin: Implications for the influence of dietary compounds on cardiac repolarisation. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2005, 371, 516–525. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Ke, X.; Ranade, V.; Somberg, J. The additive effects of the active component of grapefruit juice (naringenin) and antiarrhythmic drugs on HERG inhibition. Cardiology 2008, 110, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.Y.; Wang, Y.; Lau, C.P.; Tse, H.F.; Li, G.R. Both EGFR kinase and Src-related tyrosine kinases regulate human ether-a-go-go-related gene potassium channels. Cell. Signal. 2008, 20, 1815–1821. [Google Scholar] [CrossRef] [PubMed]
- Du, K.; De Mieri, M.; Saxena, P.; Phungula, K.V.; Wilhelm, A.; Hrubaru, M.M.; van Rensburg, E.; Zietsman, P.C.; Hering, S.; van der Westhuizen, J.H.; et al. HPLC-Based activity profiling for Herg channel inhibitors in the South African medicinal plant Galenia africana. Planta Med. 2015, 81, 1154–1162. [Google Scholar] [CrossRef] [PubMed]
- Kelemen, K.; Kiesecker, C.; Zitron, E.; Bauer, A.; Scholz, E.; Bloehs, R.; Thomas, D.; Greten, J.; Remppis, A.; Schoels, W.; et al. Green tea flavonoid epigallocatechin-3-gallate (EGCG) inhibits cardiac hERG potassium channels. Biochem. Biophys. Res. Commun. 2007, 364, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Xu, B.; Xue, Y.; Li, H.; Zhang, H.; Zhang, Y.; Kang, L.; Zhang, X.; Zhang, J.; Jia, Z.; et al. Characterization and structure-activity relationship of natural flavonoids as hERG K+ channel modulators. Int. Immunopharmacol. 2017, 45, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Scholz, E.P.; Zitron, E.; Kiesecker, C.; Thomas, D.; Kathöfer, S.; Kreuzer, J.; Bauer, A.; Katus, H.A.; Remppis, A.; Karle, C.A.; et al. Orange flavonoid hesperetin modulates cardiac hERG potassium channel via binding to amino acid F656. Nutr. Metab. Cardiovasc. Dis. 2007, 17, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.H.; Ding, J.P.; Li, H.; Pan, N.; Gan, L.; Yang, X.L.; Xu, H.B. Activation of large-conductance calcium-activated potassium channels by puerarin: The underlying mechanism of puerarin-mediated vasodilation. J. Pharmacol. Exp. Ther. 2007, 323, 391–397. [Google Scholar] [CrossRef]
- Sweeney, O.; Wang, T.; Ellory, C.; Wilkins, R.; Ma, Y. The effects of liquiritigenin on the activity of the hERG potassium channel. Br. J. Pharmacol. 2019, 176, 3067–3068. [Google Scholar]
- Yun, J.; Bae, H.; Choi, S.E.; Kim, J.H.; Choi, Y.W.; Lim, I.; Lee, C.S.; Lee, M.W.; Ko, J.H.; Seo, S.J.; et al. Taxifolin Glycoside Blocks Human ether-a-go-go Related Gene K+ Channels. Korean J. Physiol. Pharmacol. Off. J. Korean Physiol. Soc. Korean Soc. Pharmacol. 2013, 17, 37. [Google Scholar] [CrossRef]
- Sanguinetti, M.C.; Jurkiewicz, N.K. Two components of cardiac delayed rectifier K+ current. Differential sensitivity to block by class III antiarrhythmic agents. J. Gen. Physiol. 1990, 96, 195–215. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, L.; Dong, L.; Song, Q.; Guo, P.; Wang, Y.; Chen, Z.; Zhang, M. Hesperetin improves diabetic coronary arterial vasomotor responsiveness by upregulating myocyte voltage-gated K+ channels. Exp. Ther. Med. 2020, 20, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, S.; Wang, S.; Gao, P.; Dai, L. A comprehensive review on Pueraria: Insights on its chemistry and medicinal value. Biomed. Pharmacother. 2020, 131, 110734. [Google Scholar] [CrossRef] [PubMed]
- Ouadid-Ahidouch, H.; Ahidouch, A.; Pardo, L.A. Kv10.1 K+ channel: From physiology to cancer. PflÜGers-Arch. Eur. J. Physiol. 2016, 468, 751–762. [Google Scholar] [CrossRef]
- Marty, A. Ca-dependent K channels with large unitary conductance in chromaffin cell membranes. Nature 1981, 291, 497–500. [Google Scholar] [CrossRef]
- Cui, J.; Yang, H.; Lee, U.S. Molecular mechanisms of BK channel activation. Cell. Mol. Life Sci. 2009, 66, 852–875. [Google Scholar] [CrossRef]
- Kulawiak, B.; Kudin, A.P.; Szewczyk, A.; Kunz, W.S. BK channel openers inhibit ROS production of isolated rat brain mitochondria. Exp. Neurol. 2008, 212, 543–547. [Google Scholar] [CrossRef]
- Szabo, I.; Zoratti, M. Mitochondrial channels: Ion fluxes and more. Physiol. Rev. 2014, 94, 519–608. [Google Scholar] [CrossRef]
- Krabbendam, I.E.; Honrath, B.; Culmsee, C.; Dolga, A.M. Mitochondrial Ca2+-activated K+ channels and their role in cell life and death pathways. Cell Calcium 2018, 69, 101–111. [Google Scholar] [CrossRef]
- Saponara, S.; Testai, L.; Iozzi, D.; Martinotti, E.; Martelli, A.; Chericoni, S.; Sgaragli, G.; Fusi, F.; Calderone, V. (+/−)-Naringenin as large conductance Ca2+-activated K+ (BKCa) channel opener in vascular smooth muscle cells. Br. J. Pharmacol. 2006, 149, 1013–1021. [Google Scholar] [CrossRef]
- Hsu, H.T.; Tseng, Y.T.; Lo, Y.C.; Wu, S.N. Ability of naringenin, a bioflavonoid, to activate M-type potassium current in motor neuron-like cells and to increase BKCa-channel activity in HEK293T cells transfected with α-hSlo subunit. BMC Neurosci. 2014, 15, 135. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Pan, A.; Zuo, W.; Guo, J.; Zhou, W. Relaxant effect of flavonoid naringenin on contractile activity of rat colonic smooth muscle. J. Ethnopharmacol. 2014, 155, 1177–1183. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Xu, J.W.; Xiao, Z.T.; Chen, R.F.; Zhang, Y.L.; Lin, J.B.; Cheng, K.L.; Wei, G.Y.; Li, P.B.; Zhou, W.L.; et al. Naringin and naringenin relax rat tracheal smooth by regulating BKCa activation. J. Med. Food 2019, 22, 963–970. [Google Scholar] [CrossRef] [PubMed]
- Testai, L.; Martelli, A.; Marino, A.; D’antongiovanni, V.; Ciregia, F.; Giusti, L.; Lucacchini, A.; Chericoni, S.; Breschi, M.C.; Calderone, V. The activation of mitochondrial BK potassium channels contributes to the protective effects of naringenin against myocardial ischemia/reperfusion injury. Biochem. Pharmacol. 2013, 85, 1634–1643. [Google Scholar] [CrossRef] [PubMed]
- Testai, L.; Da Pozzo, E.; Piano, I.; Pistelli, L.; Gargini, C.; Breschi, M.C.; Braca, A.; Martini, C.; Martelli, A.; Calderone, V. The citrus flavanone naringenin produces cardioprotective effects in hearts from 1 year old rat, through activation of mitoBK channels. Front. Pharmacol. 2017, 8, 71. [Google Scholar] [CrossRef]
- Kampa, R.P.; Kicinska, A.; Jarmuszkiewicz, W.; Pasikowska-Piwko, M.; Dolegowska, B.; Debowska, R.; Szewczyk, A.; Bednarczyk, P. Naringenin as an opener of mitochondrial potassium channels in dermal fibroblasts. Exp. Dermatol. 2019, 28, 543–550. [Google Scholar] [CrossRef]
- Kicinska, A.; Kampa, R.P.; Daniluk, J.; Sek, A.; Jarmuszkiewicz, W.; Szewczyk, A.; Bednarczyk, P. Regulation of the mitochondrial BKCa channel by the citrus flavonoid naringenin as a potential means of preventing cell damage. Molecules 2020, 25, 3010. [Google Scholar] [CrossRef]
- Da Pozzo, E.; Costa, B.; Cavallini, C.; Testai, L.; Martelli, A.; Calderone, V.; Martini, C. The citrus flavanone naringenin protects myocardial cells against age-associated damage. Oxidative Med. Cell. Longev. 2017, 2017, 9536148. [Google Scholar] [CrossRef]
- Richter-Laskowska, M.; Trybek, P.; Bednarczyk, P.; Wawrzkiewicz-Jałowiecka, A. To what extent naringenin binding and membrane depolarization shape mitoBK channel gating—A machine learning approach. PLoS Comput. Biol. 2022, 18, e1010315. [Google Scholar] [CrossRef]
- Côrtes, S.F.; Rezende, B.A.; Corriu, C.; Medeiros, I.A.; Teixeira, M.M.; Lopes, M.J.; Lemos, V.S. Pharmacological evidence for the activation of potassium channels as the mechanism involved in the hypotensive and vasorelaxant effect of dioclein in rat small resistance arteries. Br. J. Pharmacol. 2001, 133, 849. [Google Scholar] [CrossRef]
- Dimpfel, W. Different anticonvulsive effects of hesperidin and its aglycone hesperetin on electrical activity in the rat hippocampus in-vitro. J. Pharm. Pharmacol. 2006, 58, 375–379. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kim, W.J.; Cha, E.J. Quercetin-induced growth inhibition in human bladder cancer cells is associated with an increase in Ca2+-activated K+ channels. Korean J. Physiol. Pharmacol. 2011, 15, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Melnyk, M.I.; Dryn, D.O.; Al Kury, L.T.; Zholos, A.V.; Soloviev, A.I. Liposomal quercetin potentiates maxi-K channel openings in smooth muscles and restores its activity after oxidative stress. J. Liposome Res. 2019, 29, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.M.; Zhang, Z.Y.; Wang, R.X. Protective mechanisms of quercetin against myocardial ischemia reperfusion injury. Front. Physiol. 2020, 11, 956. [Google Scholar] [CrossRef] [PubMed]
- Kampa, R.P.; Sek, A.; Szewczyk, A.; Bednarczyk, P. Cytoprotective effects of the flavonoid quercetin by activating mitochondrial BKCa channels in endothelial cells. Biomed. Pharmacother. 2021, 142, 112039. [Google Scholar] [CrossRef]
- Kampa, R.P.; Gliździńska, A.; Szewczyk, A.; Bednarczyk, P.; Filipek, S. Flavonoid quercetin abolish paxilline inhibition of the mitochondrial BKCa channel. Mitochondrion 2022, 65, 23–32. [Google Scholar] [CrossRef]
- Li, Y.; Starrett, J.E.; Meanwell, N.A.; Johnson, G.; Harte, W.E.; Dworetzky, S.I.; Boissard, C.G.; Gribkoff, V.K. The discovery of novel openers of Ca2+-dependent large-conductance potassium channels: Pharmacophore search and physiological evaluation of flavonoids. Bioorg. Med. Chem. Lett. 1997, 7, 759–762. [Google Scholar] [CrossRef]
- Mahobiya, A.; Singh, T.U.; Rungsung, S.; Kumar, T.; Chandrasekaran, G.; Parida, S.; Kumar, D. Kaempferol-induces vasorelaxation via endothelium-independent pathways in rat isolated pulmonary artery. Pharmacol. Rep. 2018, 70, 863–874. [Google Scholar] [CrossRef]
- Xu, Y.; Leung, G.; Wong, P.; Vanhoutte, P.; Man, R. Kaempferol stimulates large conductance Ca2+-activated K+ (BKCa) channels in human umbilical vein endothelial cells via a cAMP/PKA-dependent pathway. Br. J. Pharmacol. 2008, 154, 1247–1253. [Google Scholar] [CrossRef]
- Xu, Y.; Leung, S.; Leung, G.; Man, R. Kaempferol enhances endothelium-dependent relaxation in the porcine coronary artery through activation of large-conductance Ca 2+-activated K+ channels. Br. J. Pharmacol. 2015, 172, 3003–3014. [Google Scholar] [CrossRef]
- Kampa, R.P.; Flori, L.; Sęk, A.; Spezzini, J.; Brogi, S.; Szewczyk, A.; Calderone, V.; Bednarczyk, P.; Testai, L. Luteolin-Induced Activation of Mitochondrial BKCa Channels: Undisclosed Mechanism of Cytoprotection. Antioxidants 2022, 11, 1892. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Gonzalez, L.A.; Horrigan, F.T. Nobiletin Inhibition of BK Channels. Biophys. J. 2019, 116, 104a. [Google Scholar] [CrossRef]
- Saadat, S.; Boskabadi, J.; Boskabady, M.H. Contribution of potassium channels, beta2-adrenergic and histamine H1 receptors in the relaxant effect of baicalein on rat tracheal smooth muscle. Iran. J. Basic Med. Sci. 2019, 22, 1347. [Google Scholar] [PubMed]
- Lamai, J.; Mahabusarakam, W.; Ratithammatorn, T.; Hiranyachattada, S. Effects of morelloflavone from Garcinia dulcis on vasorelaxation of isolated rat thoracic aorta. J. Physiol. Biomed. Sci. 2013, 26, 13–17. [Google Scholar]
- Bai, B.; Lu, N.; Zhang, W.; Lin, J.; Zhao, T.; Zhou, S.; Khasanova, E.; Zhang, L. Inhibitory Effects of Genistein on Vascular Smooth Muscle Cell Proliferation Induced by Ox-LDL: Role of BKCa Channels. Anal. Cell. Pathol. 2020, 2020, 8895449. [Google Scholar] [CrossRef]
- Sun, L.; Zhao, T.; Ju, T.; Wang, X.; Li, X.; Wang, L.; Zhang, L.; Yu, G. A combination of intravenous genistein plus Mg2+ enhances antihypertensive effects in SHR by endothelial protection and BKCa channel inhibition. Am. J. Hypertens. 2015, 28, 1114–1120. [Google Scholar] [CrossRef]
- Zhou, R.; Liu, L.; Hu, D. Involvement of BKCa α subunit tyrosine phosphorylation in vascular hyporesponsiveness of superior mesenteric artery following hemorrhagic shock in rats. Cardiovasc. Res. 2005, 68, 327–335. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, H.Y.; Liu, Y.G.; Song, Z.; She, G.; Xiao, G.S.; Wang, Y.; Li, G.R.; Deng, X.L. Tyrphostin AG 556 increases the activity of large conductance Ca2+-activated K+ channels by inhibiting epidermal growth factor receptor tyrosine kinase. J. Cell. Mol. Med. 2017, 21, 1826–1834. [Google Scholar] [CrossRef]
- Stumpff, F.; Que, Y.; Boxberger, M.; Strauss, O.; Wiederholt, M. Stimulation of maxi-K channels in trabecular meshwork by tyrosine kinase inhibitors. Investig. Ophthalmol. Vis. Sci. 1999, 40, 1404–1417. [Google Scholar]
- Zhang, H.T.; Wang, Y.; Deng, X.L.; Dong, M.Q.; Zhao, L.M.; Wang, Y.W. Daidzein relaxes rat cerebral basilar artery via activation of large-conductance Ca2+-activated K+ channels in vascular smooth muscle cells. Eur. J. Pharmacol. 2010, 630, 100–106. [Google Scholar] [CrossRef]
- Nevala, R.; Paukku, K.; Korpela, R.; Vapaatalo, H. Calcium-sensitive potassium channel inhibitors antagonize genistein-and daidzein-induced arterial relaxation in vitro. Life Sci. 2001, 69, 1407–1417. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Gao, Q.; Yao, H.; Xia, Q. Mitochondrial mechanism of cardioprotective effect of puerarin against H2O2-stress in rats. Zhongguo Ying Yong Sheng Xue Zhi = Zhongguo Yingyong Shenglixue Zazhi = Chin. J. Appl. Physiol. 2008, 24, 399–404. [Google Scholar]
- Yao, H.; Gao, Q.; Xia, Q. The role of mitochondrial K+ channels in the cardioprotection of puerarin against hypoxia/reoxygenation injury in rats. Zhongguo Ying Yong Sheng Xue Zhi = Zhongguo Yingyong Shenglixue Zazhi = Chin. J. Appl. Physiol. 2010, 26, 459–462. [Google Scholar]
- Koh, D.S.; Reid, G.; Vogel, W. Effect of the flavoid phloretin on Ca2+-activated K+ channels in myelinated nerve fibres of Xenopus laevis. Neurosci. Lett. 1994, 165, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, L.A.; Ma, Z.; Horrigan, F.T. The BK Channel Opener Phloretin Influences Voltage-and Calcium-Dependent Gating. Biophys. J. 2012, 102, 683a–684a. [Google Scholar] [CrossRef]
- Gonzalez, L.A.; Ma, Z.; Horrigan, F.T. Potential Sites of Action for Phloretin on BK Channels. Biophys. J. 2013, 104, 463a. [Google Scholar] [CrossRef]
- Marques, A.A.M.; da Silva, C.H.F.; de Souza, P.; de Almeida, C.L.; Cechinel-Filho, V.; Lourenço, E.L.; Junior, A.G. Nitric oxide and Ca2+-activated high-conductance K+ channels mediate nothofagin-induced endothelium-dependent vasodilation in the perfused rat kidney. Chem. Biol. Interact. 2020, 327, 109182. [Google Scholar] [CrossRef]
- Teodoro, J.S.; Duarte, F.V.; Rolo, A.P.; Palmeira, C.M. Chapter 28—Mitochondria as a Target for Safety and Toxicity Evaluation of Nutraceuticals. In Nutraceuticals; Gupta, R.C., Ed.; Academic Press: Boston, MA, USA, 2016; pp. 387–400. [Google Scholar] [CrossRef]
- Ma, Y.G.; Liang, L.; Zhang, Y.B.; Wang, B.F.; Bai, Y.G.; Dai, Z.J.; Xie, M.J.; Wang, Z.W. Berberine reduced blood pressure and improved vasodilation in diabetic rats. J. Mol. Endocrinol. 2017, 59, 191–204. [Google Scholar] [CrossRef]
- Wu, J.; Nakashima, S.; Shigyo, M.; Yamasaki, M.; Ikuno, S.; Morikawa, A.; Takegami, S.; Nakamura, S.; Konishi, A.; Kitade, T.; et al. Antihypertensive constituents in Sanoshashinto. J. Nat. Med. 2020, 74, 421–433. [Google Scholar] [CrossRef]
- Cordeiro, B.; Shinn, C.; Sellke, F.W.; Clements, R.T. Rottlerin-induced BKCa channel activation impairs specific contractile responses and promotes vasodilation. Ann. Thorac. Surg. 2015, 99, 626–634. [Google Scholar] [CrossRef]
- Goldklang, M.P.; Perez-Zoghbi, J.F.; Trischler, J.; Nkyimbeng, T.; Zakharov, S.I.; Shiomi, T.; Zelonina, T.; Marks, A.R.; D’Armiento, J.M.; Marx, S.O. Treatment of experimental asthma using a single small molecule with anti-inflammatory and BK channel-activating properties. FASEB J. 2013, 27, 4975. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Han, B.; Zhang, M.; Wang, Y.H.; Tao, K.; Zhu, M.X.; He, K.; Zhang, Z.G.; Hou, S. Activation of BK channels prevents hepatic stellate cell activation and liver fibrosis through the suppression of TGFβ1/SMAD3 and JAK/STAT3 profibrotic signaling pathways. Front. Pharmacol. 2020, 11, 165. [Google Scholar] [CrossRef] [PubMed]
- Nishida, S.; Satoh, H. Possible involvement of Ca2+ activated K+ channels, SK channel, in the quercetin-induced vasodilatation. Korean J. Physiol. Pharmacol. 2009, 13, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Maaliki, D.; Shaito, A.A.; Pintus, G.; El-Yazbi, A.; Eid, A.H. Flavonoids in hypertension: A brief review of the underlying mechanisms. Curr. Opin. Pharmacol. 2019, 45, 57–65. [Google Scholar] [CrossRef]
- Calderone, V.; Chericoni, S.; Martinelli, C.; Testai, L.; Nardi, A.; Morelli, I.; Breschi, M.C.; Martinotti, E. Vasorelaxing effects of flavonoids: Investigation on the possible involvement of potassium channels. Naunyn-Schmiedeberg’S Arch. Pharmacol. 2004, 370, 290–298. [Google Scholar] [CrossRef]
- Lopes, K.S.; Marques, A.A.M.; Moreno, K.G.T.; Lorençone, B.R.; Leite, P.R.T.; da Silva, G.P.; Dos Santos, A.C.; Souza, R.I.C.; Gasparotto, F.M.; Cassemiro, N.S.; et al. Small conductance calcium-activated potassium channels and nitric oxide/cGMP pathway mediate cardioprotective effects of Croton urucurana Baill. In hypertensive rats. J. Ethnopharmacol. 2022, 293, 115255. [Google Scholar] [CrossRef]
- Oliani, J.; Ferreira, M.J.P.; Salatino, A.; Salatino, M.L.F. Leaf flavonoids from Croton urucurana and C. floribundus (Euphorbiaceae). Biochem. Syst. Ecol. 2021, 94, 104217. [Google Scholar] [CrossRef]
- da Rocha Lapa, F.; Soares, K.C.; Rattmann, Y.D.; Crestani, S.; Missau, F.C.; Pizzolatti, M.G.; Marques, M.C.A.; Rieck, L.; Santos, A.R.S. Vasorelaxant and hypotensive effects of the extract and the isolated flavonoid rutin obtained from Polygala paniculata L. J. Pharm. Pharmacol. 2011, 63, 875–881. [Google Scholar] [CrossRef]
- Chen, K.H.; Liu, H.; Sun, H.Y.; Jin, M.W.; Xiao, G.S.; Wang, Y.; Li, G.R. The natural flavone acacetin blocks small conductance Ca2+-activated K+ channels stably expressed in HEK 293 cells. Front. Pharmacol. 2017, 8, 716. [Google Scholar] [CrossRef]
- Hsueh, C.H.; Chang, P.C.; Hsieh, Y.C.; Reher, T.; Chen, P.S.; Lin, S.F. Proarrhythmic effect of blocking the small conductance calcium activated potassium channel in isolated canine left atrium. Heart Rhythm. 2013, 10, 891–898. [Google Scholar] [CrossRef]
- Tirloni, C.A.S.; Palozi, R.A.C.; Schaedler, M.I.; Guarnier, L.P.; Silva, A.O.; Marques, M.A.; Gasparotto, F.M.; Lourenço, E.L.B.; de Souza, L.M.; Junior, A.G. Influence of Luehea divaricata Mart. extracts on peripheral vascular resistance and the role of nitric oxide and both Ca+2-sensitive and Kir6. 1 ATP-sensitive K+ channels in the vasodilatory effects of isovitexin on isolated perfused mesenteric beds. Phytomedicine 2019, 56, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Reimann, F.; Ashcroft, F.M. Inwardly rectifying potassium channels. Curr. Opin. Cell Biol. 1999, 11, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Hibino, H.; Inanobe, A.; Furutani, K.; Murakami, S.; Findlay, I.; Kurachi, Y. Inwardly rectifying potassium channels: Their structure, function, and physiological roles. Physiol. Rev. 2010, 90, 291–366. [Google Scholar] [CrossRef]
- Jiao, Y.; Fan, Y.F.; Wang, Y.L.; Zhang, J.Y.; Chen, S.; Chen, Z.W. Protective effect and mechanism of total flavones from Rhododendron simsii planch flower on cultured rat cardiomyocytes with anoxia and reoxygenation. Evid. Based Complement. Altern. Med. 2015, 2015, 863531. [Google Scholar] [CrossRef]
- Li, W.; Dong, M.; Guo, P.; Liu, Y.; Jing, Y.; Chen, R.; Zhang, M. Luteolin-induced coronary arterial relaxation involves activation of the myocyte voltage-gated K+ channels and inward rectifier K+ channels. Life Sci. 2019, 221, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Trezza, A.; Cicaloni, V.; Porciatti, P.; Langella, A.; Fusi, F.; Saponara, S.; Spiga, O. From in silico to in vitro: A trip to reveal flavonoid binding on the Rattus norvegicus Kir6.1 ATP-sensitive inward rectifier potassium channel. PeerJ 2018, 6, e4680. [Google Scholar] [CrossRef]
- Rameshrad, M.; Omidkhoda, S.F.; Razavi, B.M.; Hosseinzadeh, H. Evaluating the possible role of mitochondrial ATP-sensitive potassium channels in the cardioprotective effects of morin in the isolated rat heart. Life Sci. 2021, 264, 118659. [Google Scholar] [CrossRef]
- Khan, A.u.; Gilani, A.H. Selective bronchodilatory effect of Rooibos tea (Aspalathus linearis) and its flavonoid, chrysoeriol. Eur. J. Nutr. 2006, 45, 463–469. [Google Scholar] [CrossRef]
- Jin, J.Y.; Park, S.H.; Bae, J.H.; Cho, H.C.; Lim, J.G.; Park, W.S.; Han, J.; Lee, J.H.; Song, D.K. Uncoupling by (−)-epigallocatechin-3-gallate of ATP-sensitive potassium channels from phosphatidylinositol polyphosphates and ATP. Pharmacol. Res. 2007, 56, 237–247. [Google Scholar] [CrossRef]
- Pinho-Ribeiro, F.A.; Zarpelon, A.C.; Fattori, V.; Manchope, M.F.; Mizokami, S.S.; Casagrande, R.; Verri, W.A., Jr. Naringenin reduces inflammatory pain in mice. Neuropharmacology 2016, 105, 508–519. [Google Scholar] [CrossRef]
- Manchope, M.F.; Calixto-Campos, C.; Coelho-Silva, L.; Zarpelon, A.C.; Pinho-Ribeiro, F.A.; Georgetti, S.R.; Baracat, M.M.; Casagrande, R.; Verri, W.A., Jr. Naringenin inhibits superoxide anion-induced inflammatory pain: Role of oxidative stress, cytokines, Nrf-2 and the NO- cGMP- PKG- KATPChannel signaling pathway. PLoS ONE 2016, 11, e0153015. [Google Scholar] [CrossRef] [PubMed]
- Moradi, N.; Fakhri, S.; Farzaei, M.H.; Abbaszadeh, F. The anti-nociceptive activity of naringenin passes through L-arginine/NO/cGMP/KATP channel pathway and opioid receptors. Behav. Pharmacol. 2021, 32, 590–598. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.M.; Ma, H.J.; Guo, H.; Kong, Q.Q.; Zhang, Y. The cardioprotective effect of naringenin against ischemia–Reperfusion injury through activation of ATP-sensitive potassium channel in rat. Can. J. Physiol. Pharmacol. 2016, 94, 973–978. [Google Scholar] [CrossRef] [PubMed]
- Hua, Z.; Wang, X. Inhibitory effect of berberine on potassium channels in guinea pig ventricular myocytes. Yao Xue Xue Bao = Acta Pharm. Sin. 1994, 29, 576–580. [Google Scholar]
- Wang, Y.X.; Zheng, Y.M.; Zhou, X.B. Inhibitory effects of berberine on ATP-sensitive K+ channels in cardiac myocytes. Eur. J. Pharmacol. 1996, 316, 307–315. [Google Scholar] [CrossRef]
- Suantawee, T.; Elazab, S.T.; Hsu, W.H.; Yao, S.; Cheng, H.; Adisakwattana, S. Cyanidin stimulates insulin secretion and pancreatic β-cell gene expression through activation of L-type voltage-dependent Ca2+ channels. Nutrients 2017, 9, 814. [Google Scholar] [CrossRef]
- Ribeiro, A.R.S.; do Nascimento Valença, J.D.; da Silva Santos, J.; Boeing, T.; da Silva, L.M.; de Andrade, S.F.; Albuquerque-Júnior, R.L.; Thomazzi, S.M. The effects of baicalein on gastric mucosal ulcerations in mice: Protective pathways and anti-secretory mechanisms. Chem. Biol. Interact. 2016, 260, 33–41. [Google Scholar] [CrossRef]
- Zhao, Z.; Liu, B.; Zhang, G.; Jia, Z.; Jia, Q.; Geng, X.; Zhang, H. Molecular basis for genistein-induced inhibition of Kir2.3 currents. PflÜGers-Arch. Eur. J. Physiol. 2008, 456, 413–423. [Google Scholar] [CrossRef]
- Ko, E.A.; Park, W.S.; Son, Y.K.; Kim, D.H.; Kim, N.; Kim, H.K.; Choi, T.H.; Jung, I.D.; Park, Y.M.; Han, J. The effect of tyrosine kinase inhibitor genistein on voltage-dependent K+ channels in rabbit coronary arterial smooth muscle cells. Vasc. Pharmacol. 2009, 50, 51–56. [Google Scholar] [CrossRef]
- Okamoto, F.; Okabe, K.; Kajiya, H. Genistein, a soybean isoflavone, inhibits inward rectifier K+ channels in rat osteoclasts. Jpn. J. Physiol. 2001, 51, 501–509. [Google Scholar] [CrossRef]
- Ogata, R.; Kitamura, K.; Ito, Y.; Nakano, H. Inhibitory effects of genistein on ATP-sensitive K+ channels in rabbit portal vein smooth muscle. Br. J. Pharmacol. 1997, 122, 1395–1404. [Google Scholar] [CrossRef] [PubMed]
- Kampa, R.P.; Sęk, A.; Bednarczyk, P.; Szewczyk, A.; Calderone, V.; Testai, L. Flavonoids as new regulators of mitochondrial potassium channels: Ccontribution to cardioprotection. J. Pharm. Pharmacol. 2022, rgac093. [Google Scholar] [CrossRef] [PubMed]
- Song, D.K.; Jang, Y.; Kim, J.H.; Chun, K.J.; Lee, D.; Xu, Z. Polyphenol (−)-epigallocatechin gallate during ischemia limits infarct size via mitochondrial KATP channel activation in isolated rat hearts. J. Korean Med. Sci. 2010, 25, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Huang, X.; Li, Q.; Guan, Y.; Yuan, F.; Zhang, Y. ATP-dependent potassium channels and mitochondrial permeability transition pores play roles in the cardioprotection of theaflavin in young rat. J. Physiol. Sci. 2011, 61, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Li, L.; Yin, W.; Shen, L.; You, B.; Gao, H. Protective effect of proanthocyanidins on anoxia-reoxygenation injury of myocardial cells mediated by the PI3K/Akt/GSK-3β pathway and mitochondrial ATP-sensitive potassium channel. Mol. Med. Rep. 2014, 10, 2051–2058. [Google Scholar] [CrossRef] [PubMed]
- Couvreur, N.; Tissier, R.; Pons, S.; Chenoune, M.; Waintraub, X.; Berdeaux, A.; Ghaleh, B. The Ceiling Effect of Pharmacological Postconditioning with the Phytoestrogen Genistein Is Reversed by the GSK3β Inhibitor SB 216763 [3-(2, 4-Dichlorophenyl)-4 (1-methyl-1H-indol-3-yl)-1H-pyrrole-2, 5-dione] through Mitochondrial ATP-Dependent Potassium Channel Opening. J. Pharmacol. Exp. Ther. 2009, 329, 1134–1141. [Google Scholar]
- Tu, I.H.; Yen, H.T.D.; Cheng, H.W.; Chiu, J.H. Baicalein protects chicken embryonic cardiomyocyte against hypoxia–reoxygenation injury via μ-and δ-but not κ-opioid receptor signaling. Eur. J. Pharmacol. 2008, 588, 251–258. [Google Scholar] [CrossRef]
- Protić, D.; Beleslin-Čokić, B.; Spremović-Rađenović, S.; Radunović, N.; Heinle, H.; Šćepanović, R.; Gojković Bukarica, L. The Different Effects of Resveratrol and Naringenin on Isolated Human Umbilical Vein: The Role of ATP-Sensitive K+ Channels. Phytother. Res. 2014, 28, 1412–1418. [Google Scholar] [CrossRef]
- Yow, T.T.; Pera, E.; Absalom, N.; Heblinski, M.; Johnston, G.A.; Hanrahan, J.R.; Chebib, M. Naringin directly activates inwardly rectifying potassium channels at an overlapping binding site to tertiapin-Q. Br. J. Pharmacol. 2011, 163, 1017–1033. [Google Scholar] [CrossRef]
- Loscalzo, L.M.; Yow, T.T.; Wasowski, C.; Chebib, M.; Marder, M. Hesperidin induces antinociceptive effect in mice and its aglicone, hesperetin, binds to μ-opioid receptor and inhibits GIRK1/2 currents. Pharmacol. Biochem. Behav. 2011, 99, 333–341. [Google Scholar] [CrossRef]
- Hammadi, R.; Kúsz, N.; Mwangi, P.W.; Kulmány, Á.; Zupkó, I.; Orvos, P.; Tálosi, L.; Hohmann, J.; Vasas, A. Isolation and pharmacological investigation of compounds from Euphorbia matabelensis. Nat. Prod. Commun. 2019, 14, 1934578X19863509. [Google Scholar] [CrossRef]
- Lesage, F.; Lazdunski, M. Molecular and functional properties of two-pore-domain potassium channels. Am. J. Physiol. Ren. Physiol. 2000, 279, F793–F801. [Google Scholar] [CrossRef] [PubMed]
- Feliciangeli, S.; Chatelain, F.C.; Bichet, D.; Lesage, F. The family of K2P channels: Salient structural and functional properties. J. Physiol. 2015, 593, 2587–2603. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Pérez, S.; Campos-Ríos, A.; Rueda-Ruzafa, L.; Lamas, J.A. Contribution of K2P potassium channels to cardiac physiology and pathophysiology. Int. J. Mol. Sci. 2021, 22, 6635. [Google Scholar] [CrossRef]
- Wiedmann, F.; Frey, N.; Schmidt, C. Two-pore-domain potassium (K2P-) channels: Cardiac expression patterns and disease-specific remodelling processes. Cells 2021, 10, 2914. [Google Scholar] [CrossRef]
- Andres-Bilbe, A.; Castellanos, A.; Pujol-Coma, A.; Callejo, G.; Comes, N.; Gasull, X. The background K+ channel TRESK in sensory physiology and pain. Int. J. Mol. Sci. 2020, 21, 5206. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Huang, L.; Liao, P.; Jiang, R. Contribution of Neuronal and Glial Two-Pore-Domain Potassium Channels in Health and Neurological Disorders. Neural Plast. 2021, 2021, 8643129. [Google Scholar] [CrossRef]
- Djillani, A.; Mazella, J.; Heurteaux, C.; Borsotto, M. Role of TREK-1 in health and disease, focus on the central nervous system. Front. Pharmacol. 2019, 10, 379. [Google Scholar] [CrossRef]
- Medhurst, A.D.; Rennie, G.; Chapman, C.G.; Meadows, H.; Duckworth, M.D.; Kelsell, R.E.; Gloger, I.I.; Pangalos, M.N. Distribution analysis of human two pore domain potassium channels in tissues of the central nervous system and periphery. Mol. Brain Res. 2001, 86, 101–114. [Google Scholar] [CrossRef]
- Ren, K.; Liu, H.; Guo, B.; Li, R.; Mao, H.; Xue, Q.; Yao, H.; Wu, S.; Bai, Z.; Wang, W. Quercetin relieves D-amphetamine-induced manic-like behaviour through activating TREK-1 potassium channels in mice. Br. J. Pharmacol. 2021, 178, 3682–3695. [Google Scholar] [CrossRef]
- Kim, E.J.; Kang, D.; Han, J. Baicalein and wogonin are activators of rat TREK-2 two-pore domain K+ channel. Acta Physiol. 2011, 202, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Gierten, J.; Ficker, E.; Bloehs, R.; Schlömer, K.; Kathöfer, S.; Scholz, E.; Zitron, E.; Katus, H.; Karle, C.; Thomas, D. Inhibition of hK2P3. 1 (TASK-1) Potassium Channels by the Tyrosine Kinase Inhibitor Genistein. Biophys. J. 2009, 96, 563a. [Google Scholar] [CrossRef]
- Gierten, J.; Ficker, E.; Bloehs, R.; Schlömer, K.; Kathöfer, S.; Scholz, E.; Zitron, E.; Kiesecker, C.; Bauer, A.; Becker, R.; et al. Regulation of two-pore-domain (K2P) potassium leak channels by the tyrosine kinase inhibitor genistein. Br. J. Pharmacol. 2008, 154, 1680–1690. [Google Scholar] [CrossRef] [PubMed]
- Kirkegaard, S.S.; Lambert, I.H.; Gammeltoft, S.; Hoffmann, E.K. Activation of the TASK-2 channel after cell swelling is dependent on tyrosine phosphorylation. Am. J. Physiol. Cell Physiol. 2010, 299, C844–C853. [Google Scholar] [CrossRef] [PubMed]
- Fusi, F.; Trezza, A.; Tramaglino, M.; Sgaragli, G.; Saponara, S.; Spiga, O. The beneficial health effects of flavonoids on the cardiovascular system: Focus on K+ channels. Pharmacol. Res. 2020, 152, 104625. [Google Scholar] [CrossRef]
- He, J.; Li, S.; Ding, Y.; Tong, Y.; Li, X. Research Progress on Natural Products’ Therapeutic Effects on Atrial Fibrillation by Regulating Ion Channels. Cardiovasc. Ther. 2022, 2022, 4559809. [Google Scholar] [CrossRef]
- Scholz, E.P.; Zitron, E.; Katus, H.A.; Karle, C.A. Cardiovascular ion channels as a molecular target of flavonoids. Cardiovasc. Ther. 2010, 28, e46–e52. [Google Scholar] [CrossRef]
- Van Dijk, C.; Driessen, A.J.; Recourt, K. The uncoupling efficiency and affinity of flavonoids for vesicles. Biochem. Pharmacol. 2000, 60, 1593–1600. [Google Scholar] [CrossRef]
- Ulrih, N.P.; Ota, A.; Šentjurc, M.; Kure, S.; Abram, V. Flavonoids and cell membrane fluidity. Food Chem. 2010, 121, 78–84. [Google Scholar] [CrossRef]
- Selvaraj, S.; Krishnaswamy, S.; Devashya, V.; Sethuraman, S.; Krishnan, U.M. Influence of membrane lipid composition on flavonoid–membrane interactions: Implications on their biological activity. Prog. Lipid Res. 2015, 58, 1–13. [Google Scholar] [CrossRef]
- Erlejman, A.; Verstraeten, S.; Fraga, C.; Oteiza, P. The interaction of flavonoids with membranes: Potential determinant of flavonoid antioxidant effects. Free Radic. Res. 2004, 38, 1311–1320. [Google Scholar] [CrossRef] [PubMed]
- Hendrich, A.B. Flavonoid-membrane interactions: Possible consequences for biological effects of some polyphenolic compounds. Acta Pharmacol. Sin. 2006, 27, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Verstraeten, S.V.; Fraga, C.G.; Oteiza, P.I. Flavonoid–Membrane interactions: Consequences for biological actions. In Plant Phenolics and Human Health: Biochemistry, Nutrition, and Pharmacology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2009; pp. 107–135. [Google Scholar]
- Veiko, A.G.; Sekowski, S.; Lapshina, E.A.; Wilczewska, A.Z.; Markiewicz, K.H.; Zamaraeva, M.; Zhao, H.c.; Zavodnik, I.B. Flavonoids modulate liposomal membrane structure, regulate mitochondrial membrane permeability and prevent erythrocyte oxidative damage. Biochim. Biophys. Acta (BBA) Biomembr. 2020, 1862, 183442. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.G. How lipids affect the activities of integral membrane proteins. Biochim. Biophys. Acta (BBA) Biomembr. 2004, 1666, 62–87. [Google Scholar] [CrossRef] [PubMed]
- Hedger, G.; Sansom, M.S. Lipid interaction sites on channels, transporters and receptors: Recent insights from molecular dynamics simulations. Biochim. Biophys. Acta (BBA) Biomembr. 2016, 1858, 2390–2400. [Google Scholar] [CrossRef]
- Corradi, V.; Sejdiu, B.I.; Mesa-Galloso, H.; Abdizadeh, H.; Noskov, S.Y.; Marrink, S.J.; Tieleman, D.P. Emerging diversity in lipid–protein interactions. Chem. Rev. 2019, 119, 5775–5848. [Google Scholar] [CrossRef]
- Levental, I.; Lyman, E. Regulation of membrane protein structure and function by their lipid nano-environment. Nat. Rev. Mol. Cell Biol. 2022, 1–16. [Google Scholar] [CrossRef]
- Yam, M.F.; Tan, C.S.; Shibao, R. Vasorelaxant effect of sinensetin via the NO/sGC/cGMP pathway and potassium and calcium channels. Hypertens. Res. 2018, 41, 787–797. [Google Scholar] [CrossRef]
- Jiang, H.; Xia, Q.; Wang, X.; Song, J.; Bruce, I. Luteolin induces vasorelaxion in rat thoracic aorta via calcium and potassium channels. Die-Pharm. Int. J. Pharm. Sci. 2005, 60, 444–447. [Google Scholar]
- Lee, W.; Woo, E.R.; Lee, D.G. Effect of apigenin isolated from Aster yomena against Candida albicans: Apigenin-triggered apoptotic pathway regulated by mitochondrial calcium signaling. J. Ethnopharmacol. 2019, 231, 19–28. [Google Scholar] [CrossRef]
- Lee, J.H.; Amarsanaa, K.; Wu, J.; Jeon, S.C.; Cui, Y.; Jung, S.C.; Park, D.B.; Kim, S.J.; Han, S.H.; Kim, H.W.; et al. Nobiletin attenuates neurotoxic mitochondrial calcium overload through K+ influx and ΔΨm across mitochondrial inner membrane. Korean J. Physiol. Pharmacol. Off. J. Korean Physiol. Soc. Korean Soc. Pharmacol. 2018, 22, 311. [Google Scholar] [CrossRef] [PubMed]
- Donato, F.; Filho, C.B.; Giacomeli, R.; Alvater, E.E.T.; Fabbro, L.D.; Antunes, M.d.S.; de Gomes, M.G.; Goes, A.T.R.; Souza, L.C.; Boeira, S.P.; et al. Evidence for the involvement of potassium channel inhibition in the antidepressant-like effects of hesperidin in the tail suspension test in mice. J. Med. Food 2015, 18, 818–823. [Google Scholar] [CrossRef] [PubMed]
- Roohbakhsh, A.; Parhiz, H.; Soltani, F.; Rezaee, R.; Iranshahi, M. Molecular mechanisms behind the biological effects of hesperidin and hesperetin for the prevention of cancer and cardiovascular diseases. Life Sci. 2015, 124, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Ngo, Q.A.; Thi, T.H.N.; Pham, M.Q.; Delfino, D.; Do, T.T. Antiproliferative and antiinflammatory coxib–Combretastatin hybrids suppress cell cycle progression and induce apoptosis of MCF7 breast cancer cells. Mol. Divers. 2021, 25, 2307–2319. [Google Scholar] [CrossRef]
- Al-Ishaq, R.K.; Abotaleb, M.; Kubatka, P.; Kajo, K.; Büsselberg, D. Flavonoids and their anti-diabetic effects: Cellular mechanisms and effects to improve blood sugar levels. Biomolecules 2019, 9, 430. [Google Scholar] [CrossRef] [PubMed]
- Den Hartogh, D.J.; Tsiani, E. Antidiabetic properties of naringenin: A citrus fruit polyphenol. Biomolecules 2019, 9, 99. [Google Scholar] [CrossRef]
- Li, S.; Zhang, Y.; Sun, Y.; Zhang, G.; Bai, J.; Guo, J.; Su, X.; Du, H.; Cao, X.; Yang, J.; et al. Naringenin improves insulin sensitivity in gestational diabetes mellitus mice through AMPK. Nutr. Diabetes 2019, 9, 28. [Google Scholar] [CrossRef]
- Multhoff, G.; Molls, M.; Radons, J. Chronic inflammation in cancer development. Front. Immunol. 2012, 2, 98. [Google Scholar] [CrossRef]
- Michels, N.; van Aart, C.; Morisse, J.; Mullee, A.; Huybrechts, I. Chronic inflammation towards cancer incidence: A systematic review and meta-analysis of epidemiological studies. Crit. Rev. Oncol. 2021, 157, 103177. [Google Scholar] [CrossRef]
- Iervolino, M.; Lepore, E.; Forte, G.; Laganà, A.S.; Buzzaccarini, G.; Unfer, V. Natural molecules in the management of polycystic ovary syndrome (PCOS): An analytical review. Nutrients 2021, 13, 1677. [Google Scholar] [CrossRef]
- Wawrzkiewicz-Jałowiecka, A.; Kowalczyk, K.; Trybek, P.; Jarosz, T.; Radosz, P.; Setlak, M.; Madej, P. In Search of New Therapeutics—Molecular Aspects of the PCOS Pathophysiology: Genetics, Hormones, Metabolism and Beyond. Int. J. Mol. Sci. 2020, 21, 7054. [Google Scholar] [CrossRef] [PubMed]
- Wawrzkiewicz-Jałowiecka, A.; Lalik, A.; Soveral, G. Recent Update on the molecular mechanisms of gonadal steroids action in adipose tissue. Int. J. Mol. Sci. 2021, 22, 5226. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, I.; Bioletti, L.; Peparini, S.; Solomita, E.; Ricci, C.; Casini, I.; Miceli, E.; Aloisi, A.M. Estrogens and phytoestrogens in body functions. Neurosci. Biobehav. Rev. 2021, 132, 648–663. [Google Scholar] [CrossRef]
- Gorzkiewicz, J.; Bartosz, G.; Sadowska-Bartosz, I. The potential effects of phytoestrogens: The role in neuroprotection. Molecules 2021, 26, 2954. [Google Scholar] [CrossRef] [PubMed]
- Torrens-Mas, M.; Roca, P. Phytoestrogens for cancer prevention and treatment. Biology 2020, 9, 427. [Google Scholar] [CrossRef] [PubMed]


| Kv Channel Subtype | Flavonoid | Type of Cell | Effect | References | |
|---|---|---|---|---|---|
| Kv1.3 | Genistein | HTL | ↓ | 30–60 | Teisseyre et al. [25] (2005) |
| Daidzein | HTL | → | Teisseyre et al. [25] (2005) | ||
| 6-Prenylnaringenin | HLJT | ↓ | Teisseyre et al. [26] (2018) | ||
| Acacetin | HLJT | ↓ | Teisseyre et al. [26] (2018) | ||
| ↓ | Zhao et al. [27] (2014) | ||||
| Chrysin | HLJT | ↓ | 26 | Teisseyre et al. [26] (2018) | |
| Chrysin + mevastatin | HLTJ | ↓ | 8 | Teisseyere et al. [28] (2022) | |
| Chrysin + simvastatin | HLTJ | ↓ | Teisseyere et al. [28] (2022) | ||
| Baicalein | HLJT | → | Teisseyre et al. [26] (2018) | ||
| Wogonin | HLJT | → | Teisseyre et al. [26] (2018) | ||
| Luteolin | HLJT | → | Teisseyre et al. [26] (2018) | ||
| Resveratrol | HTL | ↓ | Teisseyre et al. [29] (2006) | ||
| Naringenin | HTL | → | Teisseyre et al. [30] (2009) | ||
| Naringenin-4,7-dimethylether | HTL | ↓ | Teisseyre et al. [30] (2009) | ||
| HLTJ | ↓ | Gąsiorowska et al. [31] (2015) | |||
| Naringenin-7-methylether | HTL | ↓ | Teisseyre et al. [30] (2009) | ||
| HLJT | ↓ | Gąsiorowska et al. [31] (2009) | |||
| Aromadendrin | HTL | → | Teisseyre et al. [30] (2009) | ||
| Isoxanthohumol | HLJT | ↓ | Gąsiorowska et al. [31] (2015) | ||
| Xanthohumol | HLJT | ↓ | Gąsiorowska et al. [31] (2015) | ||
| 8-prenylnaringenin | HLJT | ↓ | Gasiorowska et al. [32] (2012) | ||
| Licochalcone A | HLJT | ↓ | Phan et al. [33] (2021) | ||
| 8-prenylnaringenin+mevastatin | HLTJ | ↓ | Teisseyre et al. [28] (2022) | ||
| Kv1.5 | Myricetin | HEK 293 | ↓ | Ou et al. [34] (2016) | |
| Hesperetin | HEK 293 | ↓ | Wang et al. [35] (2016) | ||
| Quercetin | Xenopus oocytes | ↑ | Yang et al. [36] (2009) | ||
| rats (in vivo) | ↑ | Morales-Cano et al. [37] (2014) | |||
| HEK 293 | ↓ | Liu et al. [38] (2012) | |||
| 3,7,3,4-tetramethylquecertin | HEK 293 | ↓ | Liu et al. [38] (2012) | ||
| 3,5,7,3,4-pentamethylquecertin | HEK 293 | ↓ | Liu et al. [38] (2012) | ||
| Apigenin | HEK 293 | ↓ | Liu et al. [38] (2012) | ||
| 7,4-dimethylapigenin | HEK 293 | ↓ | Liu et al. [38] (2012) | ||
| 5,7,4-trimethylapigenin | HEK 293 | ↓ | Liu et al. [38] (2012) | ||
| EGCG | CHO | ↓ | Choi et al. [39] (2001) | ||
| Isoliquiritigenin | H9c2 | ↓ | Noguchi et al. [40] (2008) | ||
| Acacetin | HEK 293 | ↓ | Wu et.al [41] (2011) | ||
| atrial myocytes | ↓ | Li et al. [42] (2008) | |||
| Kv1.7 | Puerarin | HEK 293 | ↓ | Xu et al. [43] (2016) | |
| (−)-Epigallocatechin-3-gallate | CHO | ↓ | Kang et al. [44] | ||
| Naringenin | CHO | ↓ | Sanson et al. [45] (2022) | ||
| Kv2.1 | Isoliquiritigenin | H9c2 | ↓ | Noguchi et al. [40] (2008) | |
| Genistein | HEK 293 | ↓ | Aréchiga-Figueroa et al. [46] (2017) | ||
| Naringenin-4,7-dimethylether | CHO | ↓ | Gu et.al [47] (2022) | ||
| Kv4.3 | Genistein | CHO | ↓ | Kim et al. [48] (2011) | |
| Daidzein | CHO | ↓ | Kim et al. [48] (2011) | ||
| Genistin | CHO | → | Kim et al. [48] (2011) | ||
| Epigallocatechin-3-gallate | CHO | ↓ | Kang et al. [44] (2010) | ||
| Naringenin | CHO | ↓ | Sanson et al. [45] (2022) | ||
| 5,7,4-trimethylapigenin | human atrial myocytes | ↓ | Liu et al. [38] (2012) | ||
| Kv10.1 | Procyanidin B1 | HEK | ↓ | Na et al. [49] (2020) |
| Flavonoid | Material | Effect | References |
|---|---|---|---|
| Naringenin | rat aortic rings | ↑ | Saponara et al. [102] (2006) |
| HEK 293T | ↑ | Hsu et al. [103] (2014) | |
| colonic smooth muscle cells | ↑ | Yang et al. [104] (2014) | |
| rat tracheal smooth muscle cells | ↑ | Shi et al. [105] (2019) | |
| mitoplasts from rat heart (left ventricular tissue) | ↑ | Tesai et al. [106,107] (2013, 2017) | |
| mitoplasts from primary human dermal fibroblasts | ↑ | Kampa et al. [108] (2019) | |
| mitoplasts from human endothelial cells EA.hy926 | ↑ | Kicinska et al. [109] (2020) | |
| Naringin | rat tracheal smooth muscle cells | ↑ | Shi et al. [105] (2019) |
| Dioclein | rat small mesenteric arteries | ↑ | Cortes et al. [112] (2001) |
| Hesperidin | rat hippocampal cells | ↑ | Dimpfel et al. [113] (2006) |
| Hesperetin | rat hippocampal cells | ↑ | Dimpfel et al. [113] (2006) |
| Quercetin | human bladder cancer cells | ↑ | Kim et al. [114] (2011) |
| murine smooth muscles (ileal myocytes) | ↑ | Melnyk et al. [115] (2019) | |
| rat coronary smooth muscle cells | ↑ | Zhang et al. [116] (2020) | |
| mitoplasts from human endothelial cells EA.hy926 | ↑ | Kampa et al. [117,118] (2021, 2022) | |
| Kaempferol | Xenopus oocytes | ↑ | Li et al. [119] (1997) |
| human umbilical vein endothelial cells | ↑ | Xu et al. [121] (2008) | |
| porcine coronary artery | ↑ | Xu et al. [122] (2015) | |
| rat pulmonary artery | ↑ | Mahobiya et al. [120] (2018) | |
| Luteolin | mitoplasts from rat cardiomyocytes, mitoplasts from human endothelial cells EA.hy926 | ↑ | Kampa et al. [123] (2022) |
| Baicalein | rat tracheal smooth muscle | ↑ | Saadat et al. [125] (2019) |
| Apigenin | Xenopus oocytes | ↑ | Li et al. [119] (1997) |
| Morelloflavone | rat thoracic aorta | ↑ | Lamai et al. [126] (2013) |
| Genistein | rat vascular smooth muscle cells | ↓ | Bai et al. [127] (2020) |
| vascular smooth muscle cells | ↓ | Sun et al. [128] (2015) | |
| rat superior mesenteric artery | ↓ | Zhou et al. [129] (2005) | |
| HEK 293 cells | ↑ | Wang et al. [130] (2017) | |
| rat mesenteric artery rings | ↑ | Nevala et al. [133] (2001) | |
| bovine trabecular meshwork cells | ↑ | Stumpff et al. [131] (1999) | |
| Daidzein | rat cerebral basilar artery smooth muscle cells | ↑ | Zhang et al. [132] (2010) |
| Xenopus oocytes | ↑ | Sun et al. [90] (2007) | |
| rat mesenteric artery rings | ↑ | Nevala et al. [133] (2001) | |
| Puerarin | Xenopus oocytes | ↑ | Sun et al. [90] (2007) |
| mitochondria of rat cardiomyocytes | ↑ | Yang et al. [134] (2008) | |
| mitochondria of rat cardiomyocytes | ↑ | Yao et al. [135] (2010) | |
| Phloretin | myelinated nerve fibres of Xenopus laevis | ↑ | Koh et al. [136] (1994) |
| heterologous expression models (unspecified in the cited work) | ↑ | Gonzalez et al. [137,138] (2012, 2013) | |
| Nothofagin | rat kidney cells | ↑ | Marques et al. [139] (2020) |
| Berberine | cerebral vascular smooth muscle cells | ↑ | Ma et al. [141] (2017) |
| Rottlerin | murine tracheal smooth muscle | ↑ | Goldklang et al. [144] (2013) |
| human hepatic stellate cells | ↑ | Yang et al. [145] (2020) |
| KATP Channels | Flavonoid | Cell Type | Effect | References |
|---|---|---|---|---|
| Kir6.1 | Quercetin | Rat norvegicus aorta/MD | ↓ | Trezza et al. [159] (2018) |
| 5–Hydroxyflavone | Rat norvegicus aorta/MD | ↓ | Trezza et al. [159] (2018) | |
| isovitexin | rat isolated mesenteric beds | ↑ | Tirloni et al. [154] (2019) | |
| Kir6.2 | Cyanidin | Rat Pancreatic -cells INS-1 | ↑ | Suantawee et al. [169] (2017) |
| (−)-Epigallocatechin-3-gallate | Xenopus oocytes | ↓ | Jin et al. [162] (2007) | |
| (−)-Epicatechin-3-gallate | Xenopus oocytes | ↓ | Jin et al. [162] (2007) | |
| (−)-Epicatechine | Xenopus oocytes | → | Jin et al. [162] (2007) | |
| (−)-Epigallocatechin | Xenopus oocytes | → | Jin et al. [162] (2007) | |
| Kir6.x | Berberine | Guinea pig ventricular myocytes | ↓ | Hua Z et al. [167] (1994) |
| Kir6.x | Berberine | Guinea pig ventricular myocytes | ↓ | Wang et al. [168] (1996) |
| Kir6.x | Naringenin | myocardial cells of Sprague-Dawley rats | ↑ | Meng et al. [166] (2016) |
| Kir6.x | Naringenin | Human Umbilical Vein | ↑ | Protic et al. [181] (2014) |
| Kir6.x | Baicelin | Rat tracheal smooth muscle | ↑ | Saadat et al. [125] (2019) |
| Kir6.x | TFR | Gat cardiomyocytes | ↑ | Jiao Li et.al. [157] (2015) |
| Kir6.x | Genistein | Rabbit portal vein smooth muscle | ↓ | Ogata et.al. [174] (1997) |
| Kir6.x | Baicalein | Mice gastric mucosal ulcerations | ↑ | Ribeiro et.al. [170] (2016) |
| Kir6.x | Morin | Mitoplasts from rat myocardial cells | ↑ | Rameshrad et.al. [160] (2021) |
| Kir6.x | Chrysoeriol | rabbit jejunum and aortic rings, guinea-pig trachea | ↑ | Khan et.al. [161] (2006) |
| Kir6.x | Vitexin | rabbit jejunum, guinea-pig trachea | ↑ | Khan et.al. [161] (2006) |
| Kir6.x | Orientin | rabbit jejunum | ↑ | Khan et.al. [161] (2006) |
| mitoKATP | Puerarin | Rat cardiomyocytes | ↑ | Yao et al. [135] (2012) |
| mitoKATP | Naringenin | Rat cardiomyocytes | ↑ | Meng et al. [166] (2016) |
| mitoKATP | Baicalein | Chicken embryonic cardiomyocyte | ↑ | Tu et al. [180] (2008) |
| mitoKATP | (−)-Epigallocatechin-3-gallate | Rat cardiomyocytes | ↑ | Song et al. [176] (2010) |
| mitoKATP | Theaflavin | Rat cardiomyocytes | ↑ | Ma et al. [177] (2011) |
| mitoKATP | Proanthocyanidins | Rat cardiomyocytes | ↑ | Hu et al. [178] (2014) |
| mitoKATP | Genistein | Rabbit cardiomyocytes | ↑ | Yao et al. [179] (2009) |
| mitoKATP | Morin | Rat cardiomyocytes | ↑ | Yao et al. [160] (2021) |
| GIRK Channels | Flavonoid | Cell Type | Effect | References |
|---|---|---|---|---|
| Kir3.1/Kir3.2 | Hesperidin | Xenopus laevis oocytes | ↓ | Loscalzo et al. [183] (2011) |
| Kir3.1/Kir3.4 | Eriodictyol | HEK-293 (human embryonic kidney) | ↓ | Hammadi et al. [184] (2019) |
| Kir3 | Naringin | Xenopus laevis oocytes | ↑ | Yow et al. [182] (2011) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Richter-Laskowska, M.; Trybek, P.; Delfino, D.V.; Wawrzkiewicz-Jałowiecka, A. Flavonoids as Modulators of Potassium Channels. Int. J. Mol. Sci. 2023, 24, 1311. https://doi.org/10.3390/ijms24021311
Richter-Laskowska M, Trybek P, Delfino DV, Wawrzkiewicz-Jałowiecka A. Flavonoids as Modulators of Potassium Channels. International Journal of Molecular Sciences. 2023; 24(2):1311. https://doi.org/10.3390/ijms24021311
Chicago/Turabian StyleRichter-Laskowska, Monika, Paulina Trybek, Domenico Vittorio Delfino, and Agata Wawrzkiewicz-Jałowiecka. 2023. "Flavonoids as Modulators of Potassium Channels" International Journal of Molecular Sciences 24, no. 2: 1311. https://doi.org/10.3390/ijms24021311
APA StyleRichter-Laskowska, M., Trybek, P., Delfino, D. V., & Wawrzkiewicz-Jałowiecka, A. (2023). Flavonoids as Modulators of Potassium Channels. International Journal of Molecular Sciences, 24(2), 1311. https://doi.org/10.3390/ijms24021311