
HS, an Ancient Molecular Recognition and Information Storage Glycosaminoglycan, Equips HS-Proteoglycans with Diverse Matrix and Cell-Interactive Properties Operative in Tissue Development and Tissue Function in Health and Disease

Abstract
1. Introduction
2. The Cell Instructive Properties of GAGs
2.1. GAG Interactions with Stem Cells
2.2. CS Sulfation Motifs Expressed by Stem Cells
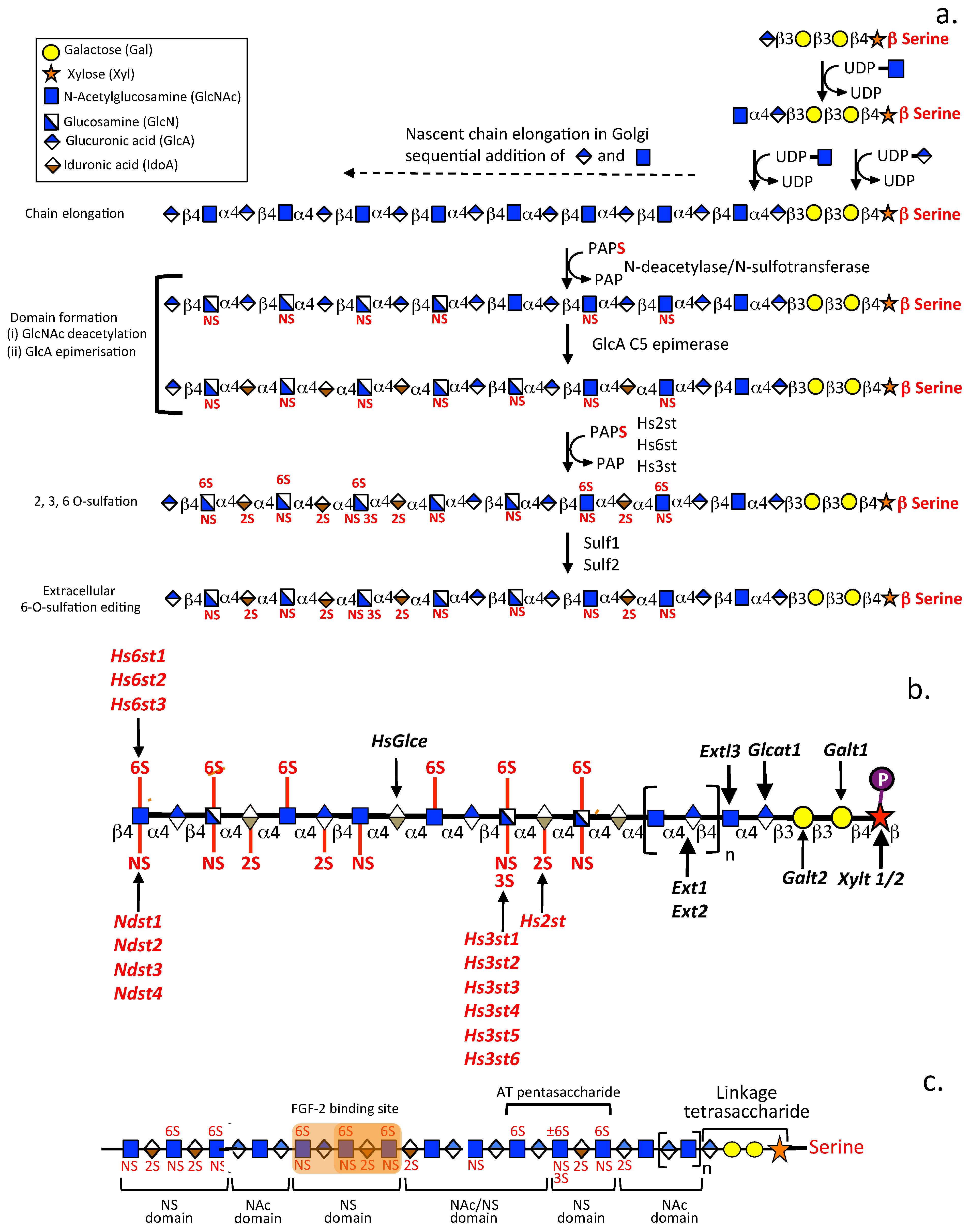
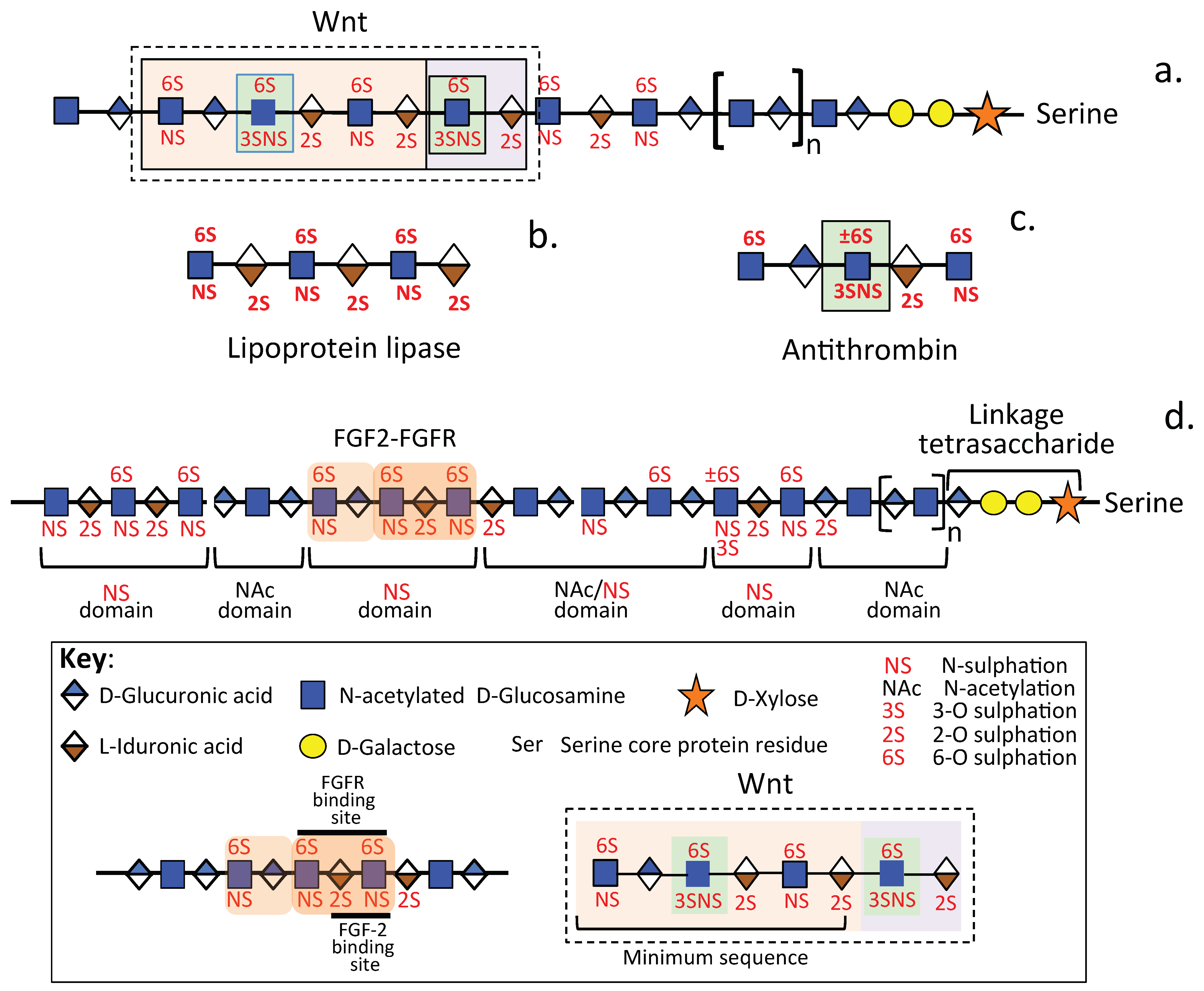
2.3. The Distribution of Sulfate Groups along GAG Chains Is Not Uniform
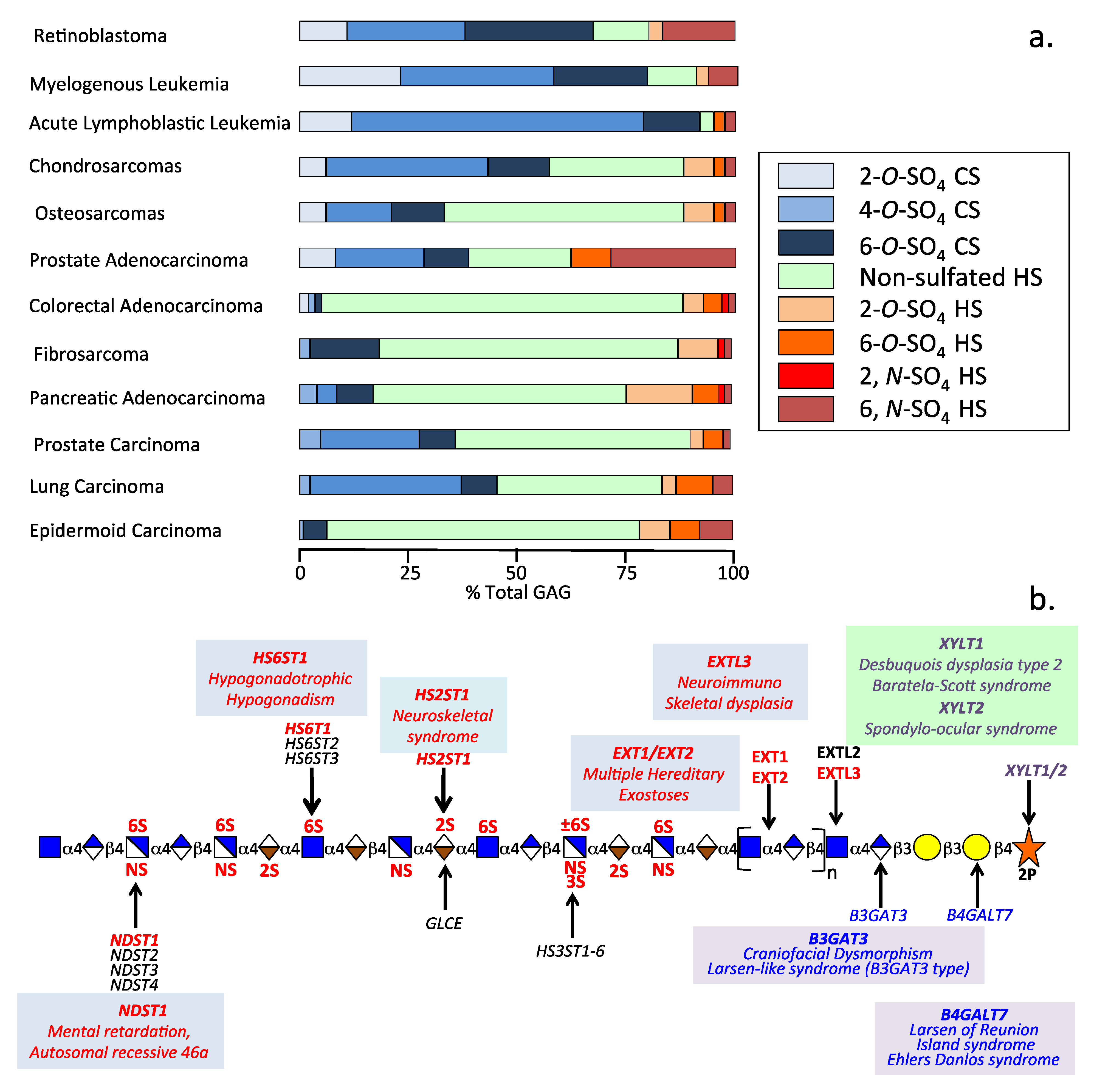
2.4. The Glyco-Code of HS and the Enigmatic 3-O Sulfated HS Sulfation Motif
3. Analysis of Glycan Structure and Function
3.1. Advanced Analytical Techniques for the Examination of GAG Structure and Function and the Identification of Their Molecular Targets
3.1.1. Neutron Scattering GAG Analysis
3.1.2. Electron Detachment Dissociation Analysis of GAGs
3.1.3. Ion Mobility Mass Spectrometry Methods for Analysis of HS Structure and Sulphation
3.1.4. X-ray Crystallography
3.1.5. In Silico Molecular Docking Simulations
3.1.6. Application of Artificial Intelligence (AI) in GAG Analysis
3.1.7. Analysis of GAG 3D Structure
3.1.8. Advanced High Precision Quantum Cellular Imaging Methodology for the Identification and Quantitation of Single Molecule Contributions to Cellular Phenotypes
3.1.9. GAGome Biomarkers of Disease
3.1.10. Transcriptomics and Its Application in the Analysis of Glycoproteomics
3.1.11. Antibodies That Detect HS Sulfation Motifs
3.1.12. Genome-Wide Analysis of Highly Specific CRISPR/Cas9 Gene Editing Sites in Musculoskeletal Disease and GAG and PG Systems

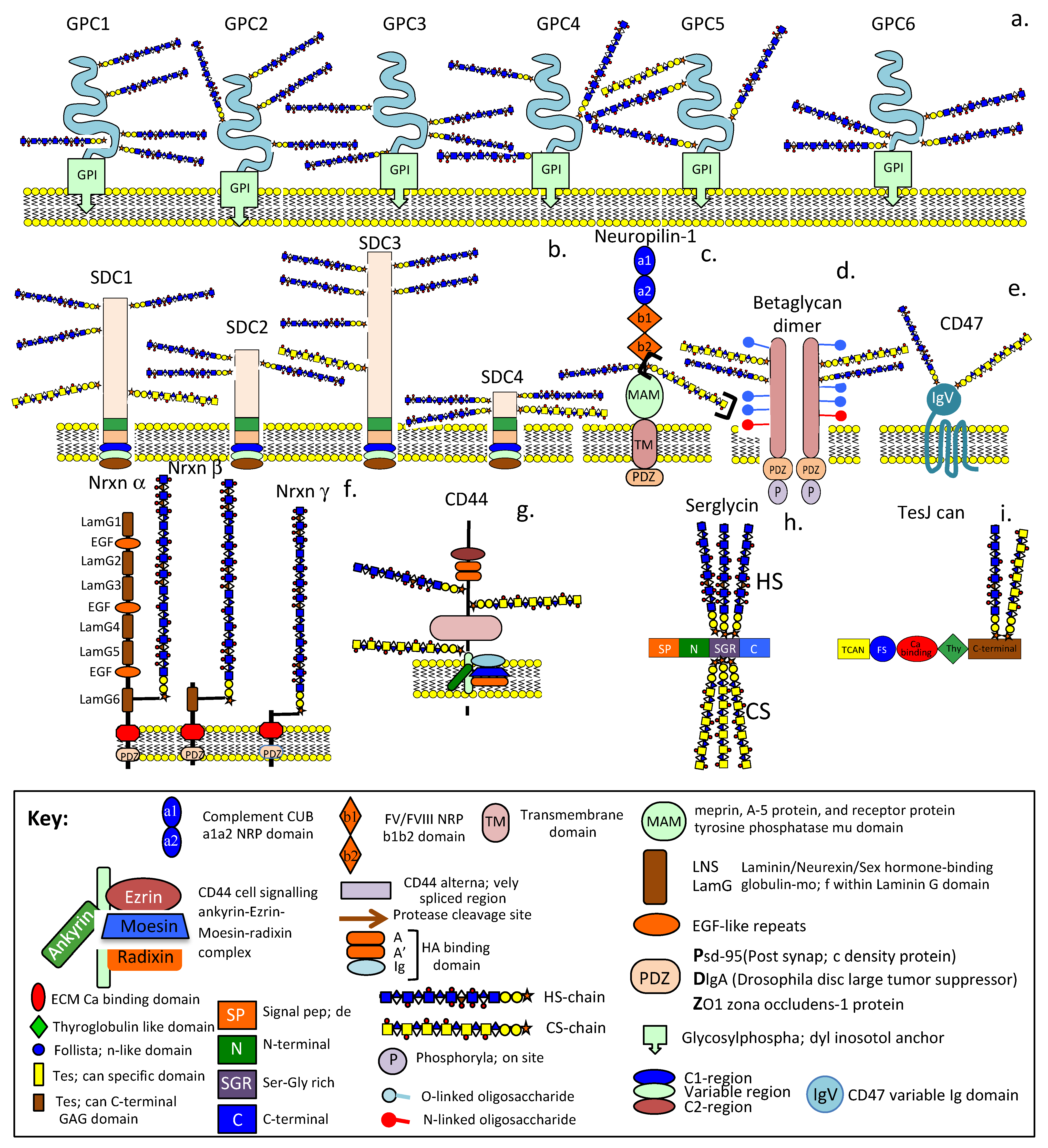
4. HS-Proteoglycans
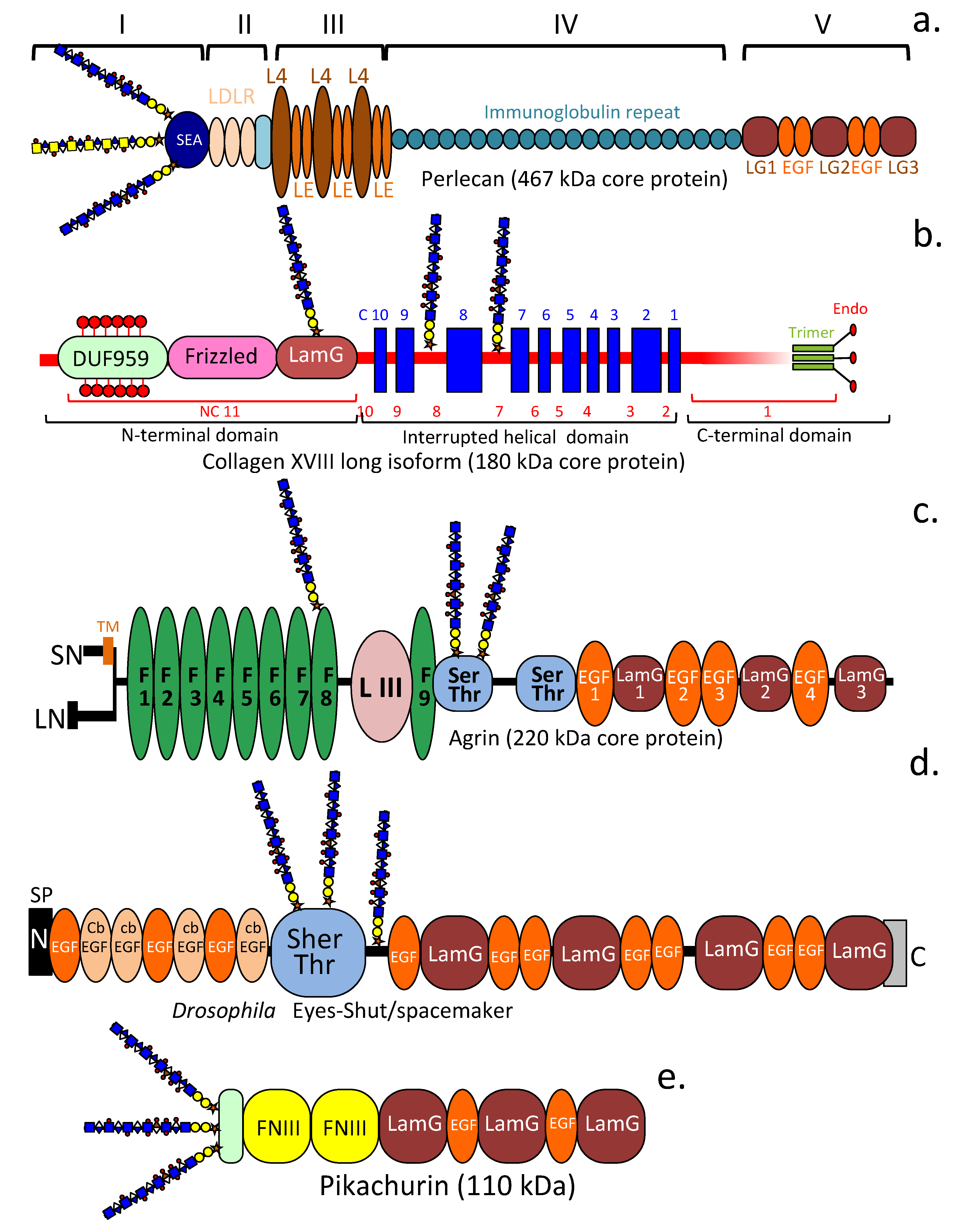
4.1. HS-PGs Are Broadly Distributed ECM, Cell Surface, Intracellular, and Nuclear Components (Table 1)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Localisation | Proteoglycan | Reference |
|---|---|---|
| Type XV Collagen | [215,216] | |
| Type XVIII Collagen | [217,218] | |
| ECM | Agrin | [219,220,221,222,223,224,225,226] |
| Perlecan | [77,227] | |
| SPOCK1, 2 (Testican-1, 2) | [228] | |
| Glypican 1–6 | [229,230,231,232] | |
| Syndecan 1–4 | [233,234,235,236,237,238] | |
| Cell surface | Betaglycan (TGF-β receptor III) | [239,240] |
| CD44E, Epican | [241,242,243,244,245,246] | |
| CD47 | [247,248] | |
| NRP-1 | [249,250] | |
| Intracellular | Serglycin | [251,252,253] |
| Neurexins α, β, γ | [104,106,254] | |
| Nuclear | Perlecan, Syndecan-1–3, Glypican-1 | [255,256,257,258,259] |
4.2. Betaglycan
4.3. CD47
4.4. Neuropilin
4.5. Neurexins
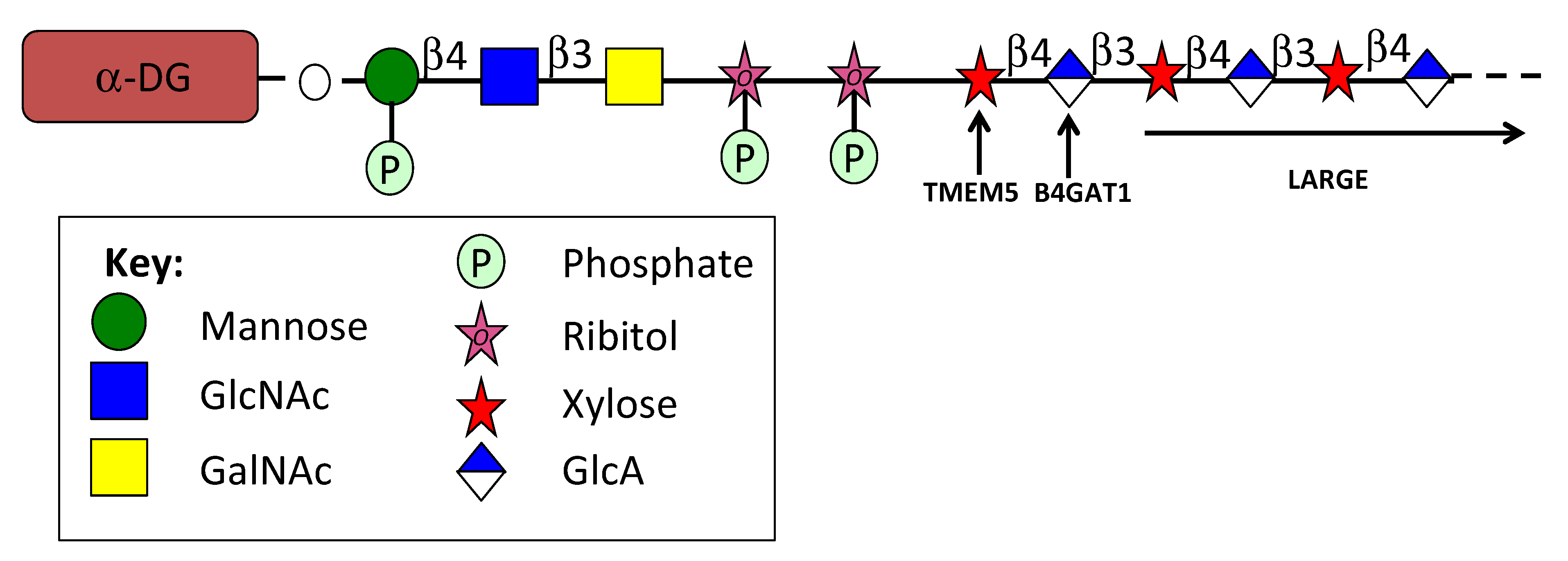
4.6. Pikachurin
4.7. Eyes Shut
5. Function Defining Properties of HS-PGs and CS-PGs in Neural Tissues
5.1. The Synapse and Perineuronal Nets (PNNs) Are Highly Sophisticated Sensory Glycocalyx-like Structures of Functional Significance in Neural Homeostasis and Neurodegenerative Pathophysiology
5.2. Roles for HSPGs in the Neuronal Synapse and PNNs
5.3. Cell Regulatory Roles of Dually Modified HS/CS PG Co-Receptors
5.4. The Roles of LamG and EGF-like Core Protein Motifs in HS-Proteoglycans
6. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Yamada, S.; Morimoto, H.; Fujisawa, T.; Sugahara, K. Glycosaminoglycans in Hydra magnipapillata (Hydrozoa, Cnidaria): Demonstration of chondroitin in the developing nematocyst, the sting organelle, and structural characterization of glycosaminoglycans. Glycobiology 2007, 17, 886–894. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Sugahara, K.; Ozbek, S. Evolution of glycosaminoglycans: Comparative biochemical study. Commun. Integr. Biol. 2011, 4, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Esko, J.; Lindahl, U. Molecular diversity of heparan sulfate. J. Clin. Investig. 2001, 108, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Esko, J.; Selleck, S.B. Order out of chaos: Assembly of ligand binding sites in heparan sulfate. Annu. Rev. Biochem. 2002, 71, 435–471. [Google Scholar] [CrossRef]
- Wisnovsky, S.; Bertozzi, C.R. Reading the glyco-code: New approaches to studying protein-carbohydrate interactions. Curr. Opin. Struct. Biol. 2022, 75, 102395. [Google Scholar] [CrossRef]
- Whitelock, J.; Iozzo, R.V. Heparan sulfate: A complex polymer charged with biological activity. Chem. Rev. 2005, 105, 2745–2764. [Google Scholar] [CrossRef] [PubMed]
- Caterson, B.; Melrose, J. Keratan sulfate, a complex glycosaminoglycan with unique functional capability. Glycobiology 2018, 28, 182–206. [Google Scholar] [CrossRef]
- Hayes, A.; Melrose, J. Immunolocalization of Keratan Sulfate in Rat Spinal Tissues Using the Keratanase Generated BKS-1(+) Neoepitope: Correlation of Expression Patterns with the Class II SLRPs, Lumican and Keratocan. Cells 2020, 9, 826. [Google Scholar] [CrossRef]
- Bertolotto, A.; Manzardo, E.; Iudicello, M.; Guglielmone, R.; Riccio, A. Keratan sulfate is a marker of differentiation of ramified microglia. Dev. Brain Res. 1995, 86, 233–241. [Google Scholar] [CrossRef]
- van den Born, J.; Salmivirta, K.; Henttinen, T.; Ostman, N.; Ishimaru, T.; Miyaura, S.; Yoshida, K.; Salmivirta, M. Novel heparan sulfate structures revealed by monoclonal antibodies. J. Biol. Chem. 2005, 280, 20516–20523. [Google Scholar] [CrossRef]
- Caterson, B.; Mahmoodian, F.; Sorrell JMHardingham, T.E.; Bayliss, M.T.; Carney, S.L.; Ratcliffe, A.; Muir, H. Modulation of native chondroitin sulfate structure in tissue development and in disease. J. Cell Sci. 1990, 97, 411–417. [Google Scholar] [CrossRef]
- David, G.; Bai, X.M.; Van der Schueren, B.; Cassiman, J.J.; Van den Berghe, H. Developmental changes in heparin sulfate expression: In situ detection with mAbs. J. Cell Biol. 1992, 119, 961–975. [Google Scholar] [CrossRef]
- Hayes, A.; Smith, S.M.; Caterson, B.; Melrose, J. Concise Review: Stem/Progenitor Cell Proteoglycans Decorated with 7-D-4, 4-C-3, and 3-B-3(-) Chondroitin Sulfate Motifs Are Morphogenetic Markers of Tissue Development. Stem Cells 2018, 36, 1475–1486. [Google Scholar] [CrossRef] [PubMed]
- Hayes, A.J.; Melrose, J. Keratan Sulfate in the Tumour Environment. Adv. Exp. Med. Biol. 2020, 1245, 39–66. [Google Scholar]
- Melrose, J. Keratan sulfate (KS)-proteoglycans and neuronal regulation in health and disease: The importance of KS-glycodynamics and interactive capability with neuroregulatory ligands. J. Neurochem. 2019, 149, 170–194. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Kusche-Gullberg, M. Heparan sulfate: Biosynthesis, Structure, and Function. Int. Rev. Cell Mol. Biol. 2016, 325, 215–273. [Google Scholar]
- Ori, A.; Wilkinson, M.C.; Fernig, D.G. A Systems Biology Approach for the Investigation of the Heparin/Heparan Sulfate Interactome. J. Biol. Chem. 2011, 286, 19892–19904. [Google Scholar] [CrossRef] [PubMed]
- Monneau, Y.; Arenzana-Seisdedos, F.; Lortat-Jacob, H. The sweet spot: How GAGs help chemokines guide migrating cells. J. Leukoc. Biol. 2016, 99, 935–953. [Google Scholar] [CrossRef]
- Nunes, Q.; Mournetas, V.; Lane, B.; Sutton, R.; Fernig, D.G.; Vasieva, O. The Heparin-Binding Protein Interactome in Pancreatic Diseases. Pancreatology 2013, 13, 598–604. [Google Scholar] [CrossRef]
- Nunes, Q.; Su, D.; Brownridge, P.J.; Simpson, D.M.; Sun, C.; Li, Y.; Bui, T.P.; Zhang, X.; Huang, W.; Rigden, D.J.; et al. The Heparin-Binding Proteome in Normal Pancreas and Murine Experimental Acute Pancreatitis. PLoS ONE 2019, 14, e0217633. [Google Scholar] [CrossRef]
- Meneghetti, M.; Hughes, A.J.; Rudd, T.R.; Nader, H.B.; Powell, A.K.; Yates, E.A.; Lima, M.A. Heparan sulfate and heparin interactions with proteins. J. R. Soc. Interface 2015, 12, 20150589. [Google Scholar] [CrossRef]
- Suflita, M.; Fu, L.; He, W.; Koffas, M.; Linhardt, R.J. Heparin and related polysaccharides: Synthesis using recombinant enzymes and metabolic engineering. Appl. Microbiol. Biotechnol. 2015, 99, 7465–7479. [Google Scholar] [CrossRef] [PubMed]
- Neelamegham, S.; Aoki-Kinoshita, K.; Bolton, E.; Frank, M.; Lisacek, F.; Lütteke, T.; O’Boyle, N.; Packer, N.H.; Stanley, P.; Toukach, P.; et al. Updates to the Symbol Nomenclature for Glycans guidelines. Glycobiology 2019, 29, 620–624. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Patel, N.G.; Nicholson, E.D.; Weiss, R.J. Spatiotemporal diversity and regulation of glycosaminoglycans in cell homeostasis and human disease. Am. J. Physiol. Cell Physiol. 2022, 322, C849–C864. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, U.; Li, J.P. Interactions between heparan sulfate and proteins-design and functional implications. Int. Rev. Cell Mol. Biol. 2009, 276, 105–159. [Google Scholar] [PubMed]
- Kjellén, L.; Lindahl, U. Specificity of glycosaminoglycan-protein interactions. Curr. Opin. Struct. Biol. 2018, 50, 101–108. [Google Scholar] [CrossRef]
- Xu, D.; Esko, J.D. Demystifying heparan sulfate-protein interactions. Annu. Rev. Biochem. 2014, 83, 129–157. [Google Scholar] [CrossRef]
- Fuerer, C.; Habib, S.J.; Nusse, R. A study on the interactions between heparan sulfate proteoglycans and Wnt proteins. Dev. Dyn. 2010, 239, 184–190. [Google Scholar] [CrossRef]
- Hayes, A.; Whitelock, J.; Melrose, J. Regulation of FGF-2, FGF-18 and Transcription Factor Activity by Perlecan in the Maturational Development of Transitional Rudiment and Growth Plate Cartilages and in the Maintenance of Permanent Cartilage Homeostasis. Int. J. Mol. Sci. 2022, 23, 1934. [Google Scholar] [CrossRef]
- Melrose, J. Perlecan, a modular instructive proteoglycan with diverse functional properties. Int. J. Biochem. Cell Biol. 2020, 128, 105849. [Google Scholar] [CrossRef] [PubMed]
- Coulson-Thomas, V. The role of heparan sulfate in development: The ectodermal story. Int. J. Exp. Pathol. 2016, 97, 213–229. [Google Scholar] [CrossRef]
- Fuster, M.; Wang, L. Endothelial heparan sulfate in angiogenesis. Prog. Mol. Biol. Transl. Sci. 2010, 93, 179–212. [Google Scholar]
- Li, J.; Partovian, C.; Li, J.; Hampton, T.G.; Metais, C.; Tkachenko, E.; Sellke, F.W.; Simons, M. Modulation of microvascular signaling by heparan sulfate matrix: Studies in syndecan-4 transgenic mice. Microvasc. Res. 2002, 64, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Bishop, J.R.; Schuksz, M.; Esko, J.D. Heparan sulfate proteoglycans fine-tune mammalian physiology. Nature 2007, 446, 1030–1037. [Google Scholar] [CrossRef] [PubMed]
- Sarrazin, S.; Lamanna, W.C.; Esko, J.D. Heparan sulfate proteoglycans. Cold Spring Harb Perspect. Biol. 2011, 3, a004952. [Google Scholar] [CrossRef]
- Chakraborty, S.; Njah, K.; Hong, W. Agrin mediates angiogenesis in the tumor microenvironment. Trends Cancer 2020, 6, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, S.; Moradi, M.; Khorshidahmad, T.; Motamedi, M. Anti-inflammatory effects of heparin and its derivatives: A systematic review. Adv. Pharmacol. Sci. 2015, 2015, 507151. [Google Scholar] [CrossRef]
- Van Kuppevelt, T.; Dennissen, M.A.B.A.; Van Venrooij, W.J.; Hoet, R.M.A.; Veerkamp, J.H. Generation and application of type-specific anti-heparan sulfate antibodies using phage display technology—Further evidence for heparin sulfate heterogeneity in the kidney. J. Biol. Chem. 1998, 273, 12960–12966. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Ha, S.; Kim, M.; Kim, S.-W.; Yun, J.; Ozcan, S.; Hwang, H.; Ji, I.J.; Yin, D.; Webster, M.J.; et al. Spatial and temporal diversity of glycome expression in mammalian brain. Proc. Natl. Acad. Sci. USA 2020, 117, 28743–28753. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, F.; Linhardt, R.J. Analysis of the glycosaminoglycan chains of proteoglycans. J. Histochem. Cytochem. 2021, 69, 121–135. [Google Scholar] [CrossRef]
- Karlsson, M.; Zhang, C.; Méar, L.; Zhong, W.; Digre, A.; Katona, B.; Sjöstedt, E.; Butler, L.; Odeberg, J.; Dusart, P.; et al. A single-cell type transcriptomics map of human tissues. Sci. Adv. 2021, 7, eabh2169. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Manabe, R.-I.; Tsutsui, K.; Yamada, T.; Kimura, M.; Nakano, I.; Shimono, C.; Sanzen, N.; Furutani, Y.; Fukuda, T.; Oguri, Y.; et al. Transcriptome-based systematic identification of extracellular matrix proteins. Proc. Natl. Acad. Sci. USA 2008, 105, 12849–12854. [Google Scholar] [CrossRef] [PubMed]
- Noborn, F.; Nilsson, J.; Larson, G. Site-specific glycosylation of proteoglycans: A revisited frontier in proteoglycan research. Matrix Biol. 2022, 12, 289–306. [Google Scholar] [CrossRef] [PubMed]
- Ghatak, S.; Maytin, E.V.; Mack, J.A.; Hascall, V.C.; Atanelishvili, I.; Moreno Rodriguez, R.; Markwald, R.R.; Misra, S. Roles of Proteoglycans and Glycosaminoglycans in Wound Healing and Fibrosis. Int. J. Cell Biol. 2015, 2015, 834893. [Google Scholar] [CrossRef] [PubMed]
- Melrose, J. Glycosaminoglycans in wound healing. Bone Tissue Regen. Insights 2016, 7, 29–50. [Google Scholar] [CrossRef]
- Hayes, A.J.; Melrose, J. Glycans and glycosaminoglycans in neurobiology: Key regulators of neuronal cell function and fate. Biochem. J. 2018, 475, 2511–2545. [Google Scholar] [CrossRef]
- Melrose, J. Functional Consequences of KS Sulfation In Electrosensory Tissues and in Neuronal Regulation. Adv. Biosyst. 2019, 3, e1800327. [Google Scholar] [CrossRef]
- Hayes, A.; Melrose, J. Aggrecan, the Primary Weight-Bearing Cartilage Proteoglycan, Has Context-Dependent, Cell-Directive Properties in Embryonic Development and Neurogenesis: Aggrecan Glycan Side Chain Modifications Convey Interactive Biodiversity. Biomolecules 2020, 10, 1244. [Google Scholar] [CrossRef]
- Roughley, P.J.; Mort, J.S. The role of aggrecan in normal and osteoarthritic cartilage. J. Exp. Orthop. 2014, 1, 8. [Google Scholar] [CrossRef]
- Roughley, P.; Melching, L.I.; Heathfield, T.F.; Pearce, R.H.; Mort, J.S. The structure and degradation of aggrecan in human intervertebral disc. Eur. Spine J. 2006, 15 (Suppl. S3), S326–S332. [Google Scholar] [CrossRef] [PubMed]
- Farrugia, B.; Hayes, A.J.; Melrose, J. Use of chondroitin sulfate to aid in-vitro stem cell differentiation in Proteoglycans. In Proteoglycans in Stem Cells; Götte, M., Forsberg-Nilsson, K., Eds.; Springer Nature Series Biology of The Extracellular Matrix; Springer: Cham, Switzerland, 2020. [Google Scholar]
- Hayes, A.; Sugahara, K.; Farrugia, B.; Whitelock, J.M.; Caterson, B.; Melrose, J. Biodiversity of CS-proteoglycan sulphation motifs: Chemical messenger recognition modules with roles in information transfer, control of cellular behaviour and tissue morphogenesis. Biochem. J. 2018, 475, 587–620. [Google Scholar] [CrossRef] [PubMed]
- Hayes, A.; Melrose, J. Neural Tissue Homeostasis and Repair Is Regulated via CS and DS Proteoglycan Motifs. Front. Cell Dev. Biol. 2021, 9, 696640. [Google Scholar] [CrossRef]
- Mizumoto, S.; Yamada, S. The Specific Role of Dermatan Sulfate as an Instructive Glycosaminoglycan in Tissue Development. Int. J. Mol. Sci. 2022, 23, 7485. [Google Scholar] [CrossRef]
- Ashworth, S.; Harrington, J.; Hammond, G.M.; Bains, K.K.; Koudouna, E.; Hayes, A.J.; Ralphs, J.R.; Regini, J.W.; Young, R.D.; Hayashi, R.; et al. Chondroitin Sulfate as a Potential Modulator of the Stem Cell Niche in Cornea. Front. Cell Dev. Biol. 2021, 8, 567358. [Google Scholar] [CrossRef] [PubMed]
- Farrugia, B.; Lord, M.S.; Whitelock, J.M.; Melrose, J. Harnessing chondroitin sulfate in composite scaffolds to direct progenitor and stem cell function for tissue repair. Biomater. Sci. 2018, 6, 947–957. [Google Scholar] [CrossRef]
- Izumikawa, T.; Sato, B.; Kitagawa, H. Chondroitin sulfate is indispensable for pluripotency and differentiation of mouse embryonic stem cells. Sci. Rep. 2014, 4, 3701. [Google Scholar] [CrossRef]
- Smith, R.; Meade, K.; Pickford, C.E.; Holley, R.J.; Merry, C.L. Glycosaminoglycans as regulators of stem cell differentiation. Biochem. Soc. Trans. 2011, 39, 383–387. [Google Scholar] [CrossRef]
- Kerever, A.; Schnack, J.; Vellinga, D.; Ichikawa, N.; Moon, C.; Arikawa-Hirasawa, E.; Efird, J.T.; Mercier, F. Novel extracellular matrix structures in the neural stem cell niche capture the neurogenic factor fibroblast growth factor 2 from the extracellular milieu. Stem Cells 2007, 25, 2146–2157. [Google Scholar] [CrossRef]
- Kerever, A.; Mercier, F.; Nonaka, R.; de Vega, S.; Oda, Y.; Zalc, B.; Okada, Y.; Hattori, N.; Yamada, Y.; Arikawa-Hirasawa, E. Perlecan is required for FGF-2 signaling in the neural stem cell niche. Stem Cell Res. 2014, 12, 492–505. [Google Scholar] [CrossRef]
- Melrose, J. Fractone Stem Cell Niche Components Provide Intuitive Clues in the Design of New Therapeutic Procedures/Biomatrices for Neural Repair. Int. J. Mol. Sci. 2022, 23, 5148. [Google Scholar] [CrossRef]
- Smith, S.; Melrose, J. Perlecan Delineates Stem Cell Niches in Human Foetal Hip, Knee and Elbow Cartilage Rudiments and Has Potential Roles in the Regulation of Stem Cell Differentiation. J. Stem Cells Res. Dev. Ther. 2016, 3, 9–16. [Google Scholar] [CrossRef]
- Archer, C.; Dowthwaite, G.P.; Francis-West, P. Development of synovial joints. Birth Defects Res. C Embryo Today 2003, 69, 144–155. [Google Scholar] [CrossRef]
- Decker, R.; Koyama, E.; Pacifici, M. Articular Cartilage: Structural and Developmental Intricacies and Questions. Curr. Osteoporos. Rep. 2015, 13, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Decker, R. Articular cartilage and joint development from embryogenesis to adulthood. Semin. Cell Dev. Biol. 2017, 62, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Dowthwaite, G.; Bishop, J.C.; Redman, S.N.; Khan, I.M.; Rooney, P.; Evans, D.J.; Haughton, L.; Bayram, Z.; Boyer, S.; Thomson, B.; et al. The surface of articular cartilage contains a progenitor cell population. J. Cell Sci. 2004, 117, 889–897. [Google Scholar] [CrossRef]
- Hayashida, K.; Aquino, R.S.; Park, P.W. Coreceptor functions of cell surface heparan sulfate proteoglycans. Am. J. Physiol. Cell Physiol. 2022, 322, C896–C912. [Google Scholar] [CrossRef]
- Hayes, A.; Tudor, D.; Nowell, M.A.; Caterson, B.; Hughes, C.E. Chondroitin sulfate sulfation motifs as putative biomarkers for isolation of articular cartilage progenitor cells. J. Histochem. Cytochem. 2008, 56, 125–138. [Google Scholar] [CrossRef]
- Otsuki, S.; Taniguchi, N.; Grogan, S.P.; D’Lima, D.; Kinoshita, M.; Lotz, M. Expression of novel extracellular sulfatases Sulf-1 and Sulf-2 in normal and osteoarthritic articular cartilage. Arthritis Res. Ther. 2008, 10, R61. [Google Scholar] [CrossRef] [PubMed]
- Hayes, A.; MacPherson, S.; Morrison, H.; Dowthwaite, G.; Archer, C.W. The development of articular cartilage: Evidence for an appositional growth mechanism. Anat. Embryol. 2001, 203, 469–479. [Google Scholar] [CrossRef]
- Forsberg, M.; Holmborn, K.; Kundu, S.; Dagälv, A.; Kjellén, L.; Forsberg-Nilsson, K. Undersulfation of heparan sulfate restricts differentiation potential of mouse embryonic stem cells. J. Biol. Chem. 2012, 287, 10853–10862. [Google Scholar] [CrossRef]
- Papy-Garcia, D.; Albanese, P. Heparan sulfate proteoglycans as key regulators of the mesenchymal niche of hematopoietic stem cells. Glycoconj. J. 2017, 34, 377–391. [Google Scholar] [CrossRef]
- Johnson, C.; Crawford, B.E.; Stavridis, M.; Ten Dam, G.; Wat, A.L.; Rushton, G.; Ward, C.M.; Wilson, V.; van Kuppevelt, T.H.; Esko, J.D.; et al. Essential alterations of heparan sulfate during the differentiation of embryonic stem cells to Sox1-enhanced green fluorescent protein-expressing neural progenitor cells. Stem Cells 2007, 25, 1913–1923. [Google Scholar] [CrossRef] [PubMed]
- Ravikumar, M.; Smith, R.A.A.; Nurcombe, V.; Cool, S.M. Heparan Sulfate Proteoglycans: Key Mediators of Stem Cell Function. Front. Cell Dev. Biol. 2020, 8, 581213. [Google Scholar] [CrossRef] [PubMed]
- Whitelock, J.; Melrose, J. Heparan sulfate proteoglycans in healthy and diseased systems. Wiley Interdiscip Rev. Syst. Biol. Med. 2011, 3, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Whitelock, J.M.; Melrose, J.; Iozzo, R.V. Diverse cell signaling events modulated by perlecan. Biochemistry 2008, 47, 11174–11183. [Google Scholar] [CrossRef]
- Shu, C.; Hughes, C.; Smith, S.M.; Smith, M.M.; Hayes, A.; Caterson, B.; Little, C.B.; Melrose, J. The ovine newborn and human foetal intervertebral disc contain perlecan and aggrecan variably substituted with native 7D4 CS sulphation motif: Spatiotemporal immunolocalisation and co-distribution with Notch-1 in the human foetal disc. Glycoconj. J. 2013, 30, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Hayes, A.; Hughes, C.E.; Smith, S.M.; Caterson, B.; Little, C.B.; Melrose, J. The CS Sulfation Motifs 4C3, 7D4, 3B3[-]; and Perlecan Identify Stem Cell Populations and Their Niches, Activated Progenitor Cells and Transitional Areas of Tissue Development in the Fetal Human Elbow. Stem Cells Dev. 2016, 25, 836–847. [Google Scholar] [CrossRef]
- Melrose, J.; Isaacs, M.D.; Smith, S.M.; Hughes, C.E.; Little, C.B.; Caterson, B.; Hayes, A.J. Chondroitin sulfate and heparan sulfate sulphation motifs and their proteoglycans are involved in articular cartilage formation during human foetal knee joint development. Histochem. Cell Biol. 2012, 138, 461–475. [Google Scholar] [CrossRef] [PubMed]
- Sorrell, J.; Mahmoodian, F.; Schafer, I.A.; Davis, B.; Caterson, B. Identification of monoclonal antibodies that recognize novel epitopes in native chondroitin/dermatan sulfate glycosaminoglycan chains: Their use in mapping functionally distinct domains of human skin. J. Histochem. Cytochem. 1990, 38, 393–402. [Google Scholar] [CrossRef]
- Persson, A.; Vorontsov, E.; Larson, G.; Nilsson, J. Glycosaminoglycan Domain Mapping of Cellular Chondroitin/Dermatan Sulfates. Sci. Rep. 2020, 10, 3506. [Google Scholar] [CrossRef] [PubMed]
- Persson, A.; Nikpour, M.; Vorontsov, E.; Nilsson, J.; Larson, G. Domain Mapping of Chondroitin/Dermatan Sulfate Glycosaminoglycans Enables Structural Characterization of Proteoglycans. Mol. Cell. Proteom. 2021, 20, 100074. [Google Scholar] [CrossRef] [PubMed]
- Holmes, S.; Nagarajan, B.; Desai, U.R. 3-O-Sulfation induces sequence-specific compact topologies in heparan sulfate that encode a dynamic sulfation code. Comput. Struct. Biotechnol. J. 2022, 20, 3884–3898. [Google Scholar] [CrossRef] [PubMed]
- Chittum, J.; Sankaranarayanan, N.V.; O’Hara, C.P.; Desai, U.R. On the selectivity of heparan sulfate recognition by SARS-CoV-2 spike glycoprotein. ACS Med. Chem. Lett. 2021, 12, 1710–1717. [Google Scholar] [CrossRef]
- Jin, L.; Abrahams, J.P.; Skinner, R.; Petitou, M.; Pike, R.N.; Carrell, R.W. The anticoagulant activation of antithrombin by heparin. Proc. Natl. Acad. Sci. USA 1997, 94, 14683–14688. [Google Scholar] [CrossRef]
- Zhao, J.; Zhu, Y.; Song, X.; Xiao, Y.; Su, G.; Liu, X.; Wang, Z.; Xu, Y.; Liu, J.; Eliezer, D.; et al. 3-O-Sulfation of heparan sulfate enhances tau interaction and cellular uptake. Angew. Chem. Int. Ed. Engl. 2020, 59, 1818–1827. [Google Scholar] [CrossRef]
- Borjigin, J.; Deng, J.; Sun, X.; De Jesus, M.; Liu, T.; Wang, M.M. Diurnal pineal 3-O-sulphotransferase 2 expression controlled by beta-adrenergic repression. J. Biol. Chem. 2003, 278, 16315–16319. [Google Scholar] [CrossRef]
- Kuberan, B.; Lech, M.; Borjigin, J.; Rosenberg, R.D. Light-induced 3-O-sulfotransferase expression alters pineal heparan sulfate fine structure. A surprising link to circadian rhythm. J. Biol. Chem. 2004, 279, 5053–5054. [Google Scholar] [CrossRef]
- Thacker, B.; Xu, D.; Lawrence, R.; Esko, J.D. Heparan sulfate 3-O-sulfation: A rare modification in search of a function. Matrix Biol. 2014, 35, 60–72. [Google Scholar] [CrossRef]
- de Agostini, A.; Dong, J.C.; de Vantéry, A.C.; Ramus, M.A.; Dentand-Quadri, I.; Thalmann, S.; Ventura, P.; Ibecheole, V.; Monge, F.; Fischer, A.-M.; et al. Human follicular fluid heparan sulfate contains abundant 3-O-sulfated chains with anticoagulant activity. J. Biol. Chem. 2008, 283, 28115–28124. [Google Scholar] [CrossRef]
- Li, J.; Su, G.; Xu, Y.; Arnold, K.; Pagadala, V.; Wang, C.; Liu, J. Synthesis of 3-O-Sulfated Heparan Sulfate Oligosaccharides Using 3-O-Sulfotransferase Isoform 4. ACS Chem. Biol. 2021, 16, 2026–2035. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Shworak, N.W.; Sinaÿ, P.; Schwartz, J.J.; Zhang, L.; Fritze, L.M.S.; Rosenberg, R.D. Expression of heparan sulfate D-glucosaminyl 3-O-sulfotransferase isoforms reveals novel substrate specificities. J. Biol. Chem. 1999, 274, 5185–5192. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Pedersen, L.C. Emerging chemical and biochemical tools for studying 3-O-sulfated heparan sulfate. Am. J. Physiol. Cell Physiol. 2022, 322, C1166–C1175. [Google Scholar] [CrossRef] [PubMed]
- Raghuraman, A.; Mosier, P.D.; Desai, U.R. Finding a needle in a haystack: Development of a combinatorial virtual screening approach for identifying high specificity heparin/heparan sulfate sequence(s). J. Med. Chem. 2006, 49, 3553–3562. [Google Scholar] [CrossRef] [PubMed]
- McKeehan, W.; Wu, X.; Kan, M. Requirement for anticoagulant heparan sulfate in the fibroblast growth factor receptor complex. J. Biol. Chem. 1999, 274, 21511–21514. [Google Scholar] [CrossRef]
- Liu, J.; Shriver, Z.; Pope, R.M.; Thorp, S.C.; Duncan, M.B.; Copeland, R.J.; Raska, C.S.; Yoshida, K.; Eisenberg, R.J.; Cohen, G.; et al. Characterization of a heparan sulfate octasaccharide that binds to herpes simplex virus type 1 glycoprotein D. J. Biol. Chem. 2002, 277, 33456–33467. [Google Scholar] [CrossRef] [PubMed]
- Deligny, A.; Denys, A.; Marcant, A.; Melchior, A.; Mazurier, J.; van Kuppevelt, T.H.; Allain, F. Synthesis of heparan sulfate with cyclophilin B-binding properties is determined by cell type-specific expression of sulfotransferases. J. Biol. Chem. 2010, 285, 1701–1715. [Google Scholar] [CrossRef] [PubMed]
- Thacker, B.; Seamen, E.; Lawrence, R.; Parker, M.W.; Xu, Y.; Liu, J.; Vander Kooi, C.W.; Esko, J.D. Expanding the 3-O-Sulfate Proteome--Enhanced Binding of Neuropilin-1 to 3-O-Sulfated Heparan Sulfate Modulates Its Activity. ACS Chem. Biol. 2016, 11, 971–980. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, J.; Powell, A.; Guimond, S. Heparan sulfate: Decoding a dynamic multifunctional cell regulator. Trends Cell Biol. 2001, 11, 75–82. [Google Scholar] [CrossRef]
- Sankarayanarayanan, N.; Strebel, T.R.; Boothello, R.S.; Sheerin, K.; Raghuraman, A.; Sallas, F.; Mosier, P.D.; Watermeyer, N.D.; Oscarson, S.; Desai, U.R. A Hexasaccharide Containing Rare 2-O-Sulfate-Glucuronic Acid Residues Selectively Activates Heparin Cofactor II. Angew Chem. Int. Ed. Engl. 2017, 56, 2312–2317. [Google Scholar] [CrossRef]
- Wishart, T.; Lovicu, F.J. Heparan sulfate proteoglycans (HSPGs) of the ocular lens. Prog. Retin. Eye Res. 2022, 3, 101118. [Google Scholar] [CrossRef]
- Fawcett, J.; Kwok, J.C.F. Proteoglycan Sulphation in the Function of the Mature Central Nervous System. Front. Integr. Neurosci. 2022, 16, 895493. [Google Scholar] [CrossRef]
- Noborn, F.; Sterky, F.H. Role of neurexin heparan sulfate in the molecular assembly of synapses-Expanding the neurexin code? FEBS J. 2021. [Google Scholar] [CrossRef]
- Maïza, A.; Sidahmed-Adrar, N.; Michel, P.P.; Carpentier, G.; Habert, D.; Dalle, C.; Redouane, W.; Hamza, M.; van Kuppevelt, T.H.; Ouidja, M.O.; et al. 3-O-sulfated heparan sulfate interactors target synaptic adhesion molecules from neonatal mouse brain and inhibit neural activity and synaptogenesis in vitro. Sci. Rep. 2020, 10, 19114. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Lu, H.; Peixoto, R.T.; Pines, M.K.; Ge, Y.; Oku, S.; Siddiqui, T.J.; Xie, Y.; Wu, W.; Archer-Hartmann, S.; et al. Heparan Sulfate Organizes Neuronal Synapses through Neurexin Partnerships. Cell 2018, 174, 1450–1464e23.1. [Google Scholar] [CrossRef]
- Omori, Y.; Araki, F.; Chaya, T.; Kajimura, N.; Irie, S.; Terada, K.; Muranishi, Y.; Tsujii, T.; Ueno, S.; Koyasu, T.; et al. Presynaptic dystroglycan-pikachurin complex regulates the proper synaptic connection between retinal photoreceptor and bipolar cells. J. Neurosci. 2012, 32, 6126–6137. [Google Scholar] [CrossRef]
- Orlandi, C.; Omori, Y.; Wang, Y.; Cao, Y.; Ueno, A.; Roux, M.J.; Condomitti, G.; de Wit, J.; Kanagawa, M.; Furukawa, T.; et al. Transsynaptic Binding of Orphan Receptor GPR179 to Dystroglycan-Pikachurin Complex Is Essential for the Synaptic Organization of Photoreceptors. Cell Rep. 2018, 25, 130–145.e5. [Google Scholar] [CrossRef]
- Sato, S.; Omori, Y.; Katoh, K.; Kondo, M.; Kanagawa, M.; Miyata, K.; Funabiki, K.; Koyasu, T.; Kajimura, N.; Miyoshi, T.; et al. Pikachurin, a dystroglycan ligand, is essential for photoreceptor ribbon synapse formation. Nat. Neurosci. 2008, 11, 923–931. [Google Scholar] [CrossRef] [PubMed]
- Alfano, G.; Kruczek, P.M.; Shah, A.Z.; Kramarz, B.; Jeffery, G.; Zelhof, A.C.; Bhattacharya, S.S. EYS Is a Protein Associated with the Ciliary Axoneme in Rods and Cones. PLoS ONE 2016, 11, e0166397. [Google Scholar] [CrossRef]
- Husain, N.; Pellikka, M.; Hong, H.; Klimentova, T.; Choe, K.M.; Clandinin, T.R.; Tepass, U. The agrin/perlecan-related protein eyes shut is essential for epithelial lumen formation in the Drosophila retina. Dev. Cell 2006, 11, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Schjoldager, K.; Narimatsu, Y.; Joshi, H.J.; Clausen, H. Global view of human protein glycosylation pathways and functions. Nat. Rev. Mol. Cell Biol. 2020, 21, 729–749. [Google Scholar] [CrossRef]
- Dworkin, L.; Clausen, H.; Joshi, H.J. Applying transcriptomics to study glycosylation at the cell type level. iScience 2022, 25, 104419. [Google Scholar] [CrossRef]
- Alvarez, R.; Blixt, O. Identification of ligand specificities for glycan-binding proteins using glycan arrays. Methods Enzym. 2006, 415, 292–310. [Google Scholar]
- Rillahan, C.; Paulson, J.C. Glycan microarrays for decoding the glycome. Annu. Rev. Biochem. 2011, 80, 797–823. [Google Scholar] [CrossRef]
- Smith, D.; Song, X.; Cummings, R.D. Use of glycan microarrays to explore specificity of glycan-binding proteins. Methods Enzym. 2010, 480, 417–444. [Google Scholar]
- Bojar, D.; Lisacek, F. Glycoinformatics in the Artificial Intelligence Era. Chem. Rev. 2022, 122, 15971–15988. [Google Scholar] [CrossRef]
- Chautard, E.; Ballut, L.; Thierry-Mieg, N.; Ricard-Blum, S. MatrixDB, a database focused on extracellular protein-protein and protein-carbohydrate interactions. Bioinformatics 2009, 25, 690–691. [Google Scholar] [CrossRef]
- Clerc, O.; Deniaud, M.; Vallet, S.D.; Naba, A.; Rivet, A.; Perez, S.; Thierry-Mieg, N.; Ricard-Blum, S. MatrixDB: Integration of new data with a focus on glycosaminoglycan interactions. Nucleic Acids Res. 2019, 47, D376–D381. [Google Scholar] [CrossRef] [PubMed]
- Kellman, B.; Lewis, N.E. Big-Data Glycomics: Tools to Connect Glycan Biosynthesis to Extracellular Communication. Trends Biochem. Sci. 2021, 46, 284–300. [Google Scholar] [CrossRef] [PubMed]
- Pérez, S.; Bonnardel, F.; Lisacek, F.; Imberty, A.; Ricard Blum, S.; Makshakova, O. GAG-DB, the New Interface of the Three-Dimensional Landscape of Glycosaminoglycans. Biomolecules 2020, 10, 1660. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Montgomery, D.; Xue, X.; Foley, B.L.; Woods, R.J. GAG Builder: A web-tool for modeling 3D structures of glycosaminoglycans. Glycobiology 2019, 29, 515–518. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Nakaya, A. The KEGG databases at GenomeNet. Nucleic Acids Res. 2002, 30, 42–46. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S.; Sato, Y.; Furumichi, M.; Tanabe, M. KEGG for integration and interpretation of large-scale molecular data sets. Nucleic Acids Res. 2012, 40, D109–D114. [Google Scholar] [CrossRef]
- Hashimoto, K.; Goto, S.; Kawano, S.; Aoki-Kinoshita, K.F.; Ueda, N.; Hamajima, M.; Kawasaki, T.; Kanehisa, M. KEGG as a glycome informatics resource. Glycobiology 2006, 16, 63R–70R. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- The Gene Ontology Consortium. The Gene Ontology resource: Enriching a GOld mine. Nucleic. Acids Res. 2021, 49, D325–D334. [Google Scholar]
- Jasnin, M. Use of neutrons reveals the dynamics of cell surface glycosaminoglycans. Methods Mol. Biol. 2012, 836, 161–169. [Google Scholar] [PubMed]
- Leite, W.; Weiss, K.L.; Phillips, G.; Zhang, Q.; Qian, S.; Tsutakawa, S.E.; Coates, L.; O’Neill, H. Conformational Dynamics in the Interaction of SARS-CoV-2 Papain-like Protease with Human Interferon-Stimulated Gene 15 Protein. J. Phys. Chem. Lett. 2021, 12, 5608–5615. [Google Scholar] [CrossRef] [PubMed]
- Pepi, L.; Sasiene, Z.J.; Mendis, P.M.; Jackson, G.P.; Amster, I.J. Structural Characterization of Sulfated Glycosaminoglycans Using Charge-Transfer Dissociation. J. Am. Soc. Mass Spectrom. 2020, 31, 2143–2153. [Google Scholar] [CrossRef]
- Leach Fr Arungundram, S.; Al-Mafraji, K.; Venot, A.; Boons, G.J.; Amster, I.J. Electron Detachment Dissociation of Synthetic Heparan Sulfate Glycosaminoglycan Tetrasaccharides Varying in Degree of Sulfation and Hexuronic Acid Stereochemistry. Int. J. Mass Spectrom. 2012, 330–332, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Compagnon, I.; Schindler, B.; Renois-Predelus, G.; Daniel, R. Lasers and ion mobility: New additions to the glycosaminoglycanomics toolkit. Curr. Opin. Struct. Biol. 2018, 50, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, R.; Chopra, P.; Joshi, A.; Yang, Z.; Vakhrushev, S.Y.; Clausen, T.M.; Painter, C.D.; Szekeres, G.P.; Chen, Y.H.; Sandoval, D.R.; et al. Dissecting structure-function of 3-O-sulfated heparin and engineered heparan sulfates. Sci. Adv. 2021, 7, eabl6026. [Google Scholar] [CrossRef] [PubMed]
- Lettow, M.; Grabarics, M.; Greis, K.; Mucha, E.; Thomas, D.A.; Chopra, P.; Boons, G.J.; Karlsson, R.; Turnbull, J.E.; Meijer, G.; et al. Cryogenic Infrared Spectroscopy Reveals Structural Modular-ity in the Vibrational Fingerprints of Heparan Sulfate Diastereomers. Anal. Chem. 2020, 92, 10228–10232. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Yang, J.Y.; Moremen, K.W.; Flanagan, J.G.; Prestegard, J.H. Structural Characterization of a Heparan Sulfate Pentamer Interacting with LAR-Ig1-2. Biochemistry 2018, 57, 2189–2199. [Google Scholar] [CrossRef]
- Clausen, T.; Sandoval, D.R.; Spliid, C.B.; Pihl, J.; Perrett, H.R.; Painter, C.D.; Narayanan, A.; Majowicz, S.A.; Kwong, E.M.; McVicar, R.N.; et al. SARS-CoV-2 Infection Depends on Cellular Heparan Sulfate and ACE2. Cell 2020, 183, 1043–1057.e15. [Google Scholar] [CrossRef]
- Ypsilanti, A.; Chedotal, A. Roundabout receptors. Adv. Neurobiol. 2014, 8, 133–164. [Google Scholar]
- Gao, Q.; Chen, C.Y.; Zong, C.; Wang, S.; Ramiah, A.; Prabhakar, P.; Morris, L.C.; Boons, G.J.; Moremen, K.W.; Prestegard, J.H. Structural Aspects of Heparan Sulfate Binding to Robo1-Ig1-2. ACS Chem. Biol. 2016, 11, 3106–3113. [Google Scholar] [CrossRef]
- Carpentier, M.; Denys, A.; Allain, F.; Vergoten, G. Molecular docking of heparin oligosaccharides with Hep-II heparin-binding domain of fibronectin reveals an interplay between the different positions of sulfate groups. Glycoconj. J. 2014, 31, 161–169. [Google Scholar] [CrossRef]
- Ballut, L.; Sapay, N.; Chautard, E.; Imberty, A.; Ricard-Blum, S. Mapping of heparin/heparan sulfate binding sites on αvβ3 integrin by molecular docking. J. Mol. Recognit. 2013, 26, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Sapay, N.; Cabannes, E.; Petitou, M.; Imberty, A. Molecular modeling of the interaction between heparan sulfate and cellular growth factors: Bringing pieces together. Glycobiology 2011, 21, 1181–1193. [Google Scholar] [CrossRef]
- Frey, L. Informatics tools to advance the biology of glycosaminoglycans and proteoglycans. Methods Mol. Biol. 2015, 1229, 271–287. [Google Scholar]
- Aoki-Kinoshita, K. Databases and artificial intelligence. What paths do databases and AI open for the future of glycan research? Glycoforum 2020, 23, A11. [Google Scholar]
- Gupta, R.; Srivastava, D.; Sahu, M.; Tiwari, S.; Ambasta, R.K.; Kumar, P. Artificial intelligence to deep learning: Machine intelligence approach for drug discovery. Mol. Divers. 2021, 25, 1315–1360. [Google Scholar] [CrossRef]
- Nayarisseri, A.; Khandelwal, R.; Tanwar, P.; Madhavi, M.; Sharma, D.; Thakur, G.; Speck-Planche, A.; Singh, S.K. Artificial Intelligence, Big Data and Machine Learning Approaches in Precision Medicine & Drug Discovery. Curr. Drug Targets 2021, 22, 631–655. [Google Scholar] [PubMed]
- Tripathi, M.; Nath, A.; Singh, T.P.; Ethayathulla, A.S.; Kaur, P. Evolving scenario of big data and Artificial Intelligence (AI) in drug discovery. Mol. Divers. 2021, 25, 1439–1460. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, N.; Goshisht, M.K.; Sahu, S.K.; Arora, C. Applications of artificial intelligence to drug design and discovery in the big data era: A comprehensive review. Mol. Divers. 2021, 25, 1643–1664. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.; Chatterjee, S.; Seeberger, P.H.; Gilmore, K. Predicting glycosylation stereoselectivity using machine learning. Chem. Sci. 2020, 12, 2931–2939. [Google Scholar] [CrossRef]
- Pesciullesi, G.; Schwaller, P.; Laino, T.; Reymond, J.L. Transfer learning enables the molecular transformer to predict regio- and stereoselective reactions on carbohydrates. Nat. Commun. 2020, 11, 4874. [Google Scholar] [CrossRef]
- Li, H.; Chiang, A.W.T.; Lewis, N.E. Artificial intelligence in the analysis of glycosylation data. Biotechnol. Adv. 2022, 60, 108008. [Google Scholar] [CrossRef] [PubMed]
- Paiardi, G.; Milanesi, M.; Wade, R.C.; D’Ursi, P.; Rusnati, M. A Bittersweet Computational Journey among Glycosaminoglycans. Biomolecules 2021, 11, 739. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Qin, C.; Chen, R.; Liu, Y.; Zhou, W.; Zhang, G.; Gao, Y.; Xiao, L.; Jia, S. Quantum Coherent Modulation-Enhanced Single-Molecule Imaging Microscopy. J. Phys. Chem. Lett. 2019, 10, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Coelho, S.; Baek, J.; Walsh, J.; Justin Gooding, J.; Gaus, K. Direct-laser writing for subnanometer focusing and single-molecule imaging. Nat. Commun. 2022, 13, 647. [Google Scholar] [CrossRef] [PubMed]
- Fischer, L.; Klingner, C.; Schlichthaerle, T.; Strauss, M.T.; Böttcher, R.; Fässler, R.; Jungmann, R.; Grashoff, C. Quantitative single-protein imaging reveals molecular complex formation of integrin, talin, and kindlin during cell adhesion. Nat. Commun. 2021, 12, 919. [Google Scholar] [CrossRef]
- Xu, H.; Xu, J.; Wang, X.; Wu, D.; Chen, Z.G.; Wang, A.Y. Quantum dot-based, quantitative, and multiplexed assay for tissue staining. ACS Appl. Mater. Interfaces 2013, 5, 2901–2907. [Google Scholar] [CrossRef]
- Kairdolf, B.; Smith, A.M.; Stokes, T.H.; Wang, M.D.; Young, A.N.; Nie, S. Semiconductor quantum dots for bioimaging and biodiagnostic applications. Annu. Rev. Anal. Chem. 2013, 6, 143–162. [Google Scholar] [CrossRef]
- Fischer, L.; Schlichthaerle, T.; Chrostek-Grashoff, A.; Grashoff, C. Peptide-PAINT Enables Investigation of Endogenous Talin with Molecular Scale Resolution in Cells and Tissues. Chembiochem 2021, 22, 2872–2879. [Google Scholar] [CrossRef]
- Bray, M.-A.; Singh, S.; Han, H.; Davis, C.T.; Borgeson, B.; Hartland, C.; Kost-Alimova, M.; Gustafsdottir, S.M.; Gibson, C.C.; E Carpenter, A. Cell Painting, a high-content image-based assay for morphological profiling using multiplexed fluorescent dyes. Nat. Protoc. 2016, 11, 1757–1774. [Google Scholar] [CrossRef]
- Lee, K. Quantum dots: A quantum jump for molecular imaging? J. Nucl. Med. 2007, 48, 1408–1410. [Google Scholar] [CrossRef]
- Bentolila, L.; Michalet, X.; Pinaud, F.F.; Tsay, J.M.; Doose, S.; Li, J.J.; Sundaresan, G.; Wu, A.M.; Gambhir, S.S.; Weiss, S. Quantum dots for molecular imaging and cancer medicine. Discov. Med. 2005, 5, 213–218. [Google Scholar]
- Yasui, M.; Hiroshima, M.; Kozuka, J.; Sako, Y.; Ueda, M. Automated single-molecule imaging in living cells. Nat. Commun. 2018, 9, 3061. [Google Scholar] [CrossRef]
- Basu, S.; Needham, L.M.; Lando, D.; Taylor, E.J.R.; Wohlfahrt, K.J.; Shah, D.; Boucher, W.; Tan, Y.L.; Bates, L.E.; Tkachenko, O.; et al. FRET-enhanced photostability allows improved single-molecule tracking of proteins and protein complexes in live mammalian cells. Nat. Commun. 2018, 9, 2520. [Google Scholar] [CrossRef] [PubMed]
- Attreed, M.; Bülow, H.E. A transgenic approach to live imaging of heparan sulfate modification patterns. Methods Mol. Biol. 2015, 1229, 253–268. [Google Scholar] [PubMed]
- Bülow, H. Imaging Glycosaminoglycan Modification Patterns In Vivo. Methods Mol. Biol. 2022, 2303, 539–557. [Google Scholar] [PubMed]
- Brown, D.; Eames, B.F. Emerging tools to study proteoglycan function during skeletal development. In Methods in Cell Biology; Academic Press: Cambridge, MA, USA, 2016; Volume 134, pp. 485–530. [Google Scholar]
- Kalita, M.; Chua JSBoothello, R.S.; Joice, A.; Antelope, O.; Roy, A.; Anandh Babu, P.V.; Saijoh, Y.; Desai, U.R.; Kuberan, B. Visualizing antithrombin-binding 3-O-sulfated heparan sulfate motifs on cell surfaces. Chem. Commun. 2020, 56, 14423–14426. [Google Scholar] [CrossRef]
- Schiff, L.; Migliori, B.; Chen, Y.; Carter, D.; Bonilla, C.; Hall, J.; Fan, M.; Tam, E.; Ahadi, S.; Fischbacher, B.; et al. Integrating deep learning and unbiased automated high-content screening to identify complex disease signatures in human fibroblasts. Nat. Commun. 2022, 13, 1590. [Google Scholar] [CrossRef]
- Grobe, K.; Ledin, J.; Ringvall, M.; Holmborn, K.; Forsberg, E.; Esko, J.D.; Kjellén, L. Heparan sulfate and development: Differential roles of the N-acetylglucosamine N-deacetylase/N-sulfotransferase isozymes. Biochim. Biophys. Acta 2002, 1573, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, J.; Miller, R.L.; Ahmed, Y.; Puvirajesinghe, T.M.; Guimond, S.E. Glycomics profiling of heparan sulfate structure and activity. Methods Enzymol. 2010, 480, 65–85. [Google Scholar]
- Gómez Toledo, A.; Sorrentino, J.T.; Sandoval, D.R.; Malmström, J.; Lewis, N.E.; Esko, J.D. A Systems View of the Heparan Sulfate Interactome. J. Histochem. Cytochem. 2021, 69, 105–119. [Google Scholar] [CrossRef]
- Abrahams, J.; Taherzadeh, G.; Jarvas, G.; Guttman, A.; Zhou, Y.; Campbell, M.P. Recent advances in glycoinformatic platforms for glycomics and glycoproteomics. Curr. Opin. Struct. Biol. 2020, 62, 56–69. [Google Scholar] [CrossRef]
- Danielsson, A.; Kogut, M.M.; Zieleniak, M.M.; Chopra, P.; Boons, G.-J.; Samsonov, S.A. Molecular Dynamics-based descriptors of 3-O-Sulfated Heparan Sulfate as Contributors of Protein Binding Specificity. Comput. Biol. Chem. 2022, 99, 107716. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Narimatsu, Y.; Clausen, T.M.; Gomes, C.; Karlsson, R.; Steentoft, C.; Spliid, C.B.; Gustavsson, T.; Salanti, A.; Persson, A.; et al. The GAGOme: A cell-based library of displayed glycosaminoglycans. Nat. Methods 2018, 15, 881–888. [Google Scholar] [CrossRef]
- Bratulic, S.; Limeta, A.; Dabestani, S.; Birgisson, H.; Enblad, G.; Stålberg, K.; Hesselager, G.; Häggman, M.; Höglund, M.; Simonson, O.E.; et al. Noninvasive detection of any-stage cancer using free glycosaminoglycans. Proc. Natl. Acad. Sci. USA 2022, 119, e2115328119. [Google Scholar] [CrossRef]
- Narimatsu, Y.; Joshi, H.J.; Nason, R.; Van Coillie, J.; Karlsson, R.; Sun, L.; Ye, Z.; Chen, Y.H.; Schjoldager, K.T.; Steentoft, C.; et al. An Atlas of Human Glycosylation Pathways Enables Display of the Human Glycome by Gene Engineered Cells. Mol. Cell 2019, 75, 394–407.e5. [Google Scholar] [CrossRef] [PubMed]
- Narimatsu, Y.; Büll, C.; Chen, Y.H.; Wandall, H.H.; Yang, Z.; Clausen, H. Genetic glycoengineering in mammalian cells. J. Biol. Chem. 2021, 296, 100448. [Google Scholar] [CrossRef] [PubMed]
- Antonopoulos, A.; Broome, S.; Sharov, V.; Ziegenfuss, C.; Easton, R.L.; Panico, M.; Dell, A.; Morris, H.R.; Haslam, S.M. Site-specific characterization of SARS-CoV-2 spike glycoprotein receptor-binding domain. Glycobiology 2021, 31, 181–187. [Google Scholar] [CrossRef]
- Byrnes, A.P.; Griffin, D.E. Binding of Sindbis virus to cell surface heparan sulfate. J. Virol. 1998, 72, 7349–7356. [Google Scholar] [CrossRef]
- Kori, M.; Aydin, B.; Gulfidan, G.; Beklen, H.; Kelesoglu, N.; Caliskan Iscan, A.; Turanli, B.; Erzik, C.; Karademir, B.; Arga, K.Y. The Repertoire of Glycan Alterations and Glycoproteins in Human Cancers. OMICS 2021, 25, 139–168. [Google Scholar] [CrossRef]
- Rosenbalm, K.; Tiemeyer, M.; Wells, L.; Aoki, K.; Zhao, P. Glycomics-informed glycoproteomic analysis of site-specific glycosylation for SARS-CoV-2 spike protein. STAR Protoc. 2020, 1, 100214. [Google Scholar] [CrossRef]
- Zhu, B.; Pardeshi, L.; Chen, Y.; Ge, W. Transcriptomic Analysis for Differentially Expressed Genes in Ovarian Follicle Activation in the Zebrafish. Front. Endocrinol. 2018, 9, 593. [Google Scholar] [CrossRef]
- Bix, G.; Iozzo, R.V. Novel interactions of perlecan: Unravelling perlecan’s roe in angiogenesis. Microsc. Res. Tech. 2008, 71, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Smits, N.; Kurup, S.; Rops, A.L.; ten Dam, G.B.; Massuger, L.F.; Hafmans, T.; Turnbull, J.E.; Spillmann, D.; Li, J.P.; Kennel, S.J.; et al. The heparan sulfate motif (GlcNS6S-IdoA2S)3, common in heparin, has a strict topography and is involved in cell behavior and disease. J. Biol. Chem. 2010, 285, 41143–41151. [Google Scholar] [CrossRef]
- Thompson, S.; Fernig, D.G.; Jesudason, E.C.; Losty, P.D.; van de Westerlo, E.M.; van Kuppevelt, T.H.; Turnbull, J.E. Heparan sulfate phage display antibodies identify distinct epitopes with complex binding characteristics: Insights into protein binding specificities. J. Biol. Chem. 2009, 284, 35621–35631. [Google Scholar] [CrossRef] [PubMed]
- Hirano, K.; Sasaki, N.; Ichimiya, T.; Miura, T.; Van Kuppevelt, T.H.; Nishihara, S. 3-O-sulfated heparan sulfate recognized by the antibody HS4C3 contributes to the differentiation of mouse embryonic stem cells via fas signaling. PLoS ONE 2012, 7, e43440. [Google Scholar] [CrossRef]
- Gallagher, J. Fell-Muir Lecture: Heparan sulfate and the art of cell regulation: A polymer chain conducts the protein orchestra. Int. J. Exp. Pathol. 2015, 96, 203–231. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Xu, Y.; Liu, J.; Ho, M. Epitope mapping by a Wnt-blocking antibody: Evidence of the Wnt binding domain in heparan sulfate. Sci. Rep. 2016, 6, 26245. [Google Scholar] [CrossRef] [PubMed]
- Lamanna, W.; Baldwin, R.J.; Padva, M.; Kalus, I.; Ten Dam, G.; van Kuppevelt, T.H.; Gallagher, J.T.; von Figura, K.; Dierks, T.; Merry, C.L. Heparan sulfate 6-O-endosulfatases: Discrete in vivo activities and functional co-operativity. Biochem. J. 2006, 400, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.; Shi, S.; Yue, J.; Xin, M.; Nairn, A.V.; Lin, L.; Liu, X.; Li, G.; Archer-Hartmann, S.A.; Dela Rosa, M.; et al. A mutant-cell library for systematic analysis of heparan sulfate structure-function relationships. Nat. Methods 2018, 15, 889–899. [Google Scholar] [CrossRef]
- van Gemst, J.; Kouwenberg, M.; Rops, A.L.W.M.M.; van Kuppevelt, T.H.; Berden, J.H.; Rabelink, T.J.; Loeven, M.A.; van der Vlag, J. Differential binding of chemokines CXCL1, CXCL2 and CCL2 to mouse glomerular endothelial cells reveals specificity for distinct heparan sulfate domains. PLoS ONE 2018, 13, e0201560. [Google Scholar] [CrossRef]
- Alhasan, A.; Spielhofer, J.; Kusche-Gullberg, M.; Kirby, J.A.; Ali, S. Role of 6-O-sulfated heparan sulfate in chronic renal fibrosis. J. Biol. Chem. 2014, 289, 20295–20306. [Google Scholar] [CrossRef]
- Caterson, B. Fell-Muir Lecture: Chondroitin sulfate glycosaminoglycans: Fun for some and confusion for others. Int. J. Exp. Pathol. 2012, 93, 1–10. [Google Scholar] [CrossRef]
- Farrugia, B.; Smith, S.M.; Shu, C.C.; Melrose, J. Spatiotemporal Expression of 3-B-3(-) and 7-D-4 Chondroitin Sulfation, Tissue Remodeling, and Attempted Repair in an Ovine Model of Intervertebral Disc Degeneration. Cartilage 2020, 11, 234–250. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, J. Heparan sulfate: Growth control with a restricted sequence menu. J. Clin. Investig. 2001, 108, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Li, S.Y.; Dang, S.S.; Cheng, Y.A.; Zhang, X.; Wang, W.J.; Hughes, C.E.; Caterson, B. Increased expression of chondroitin sulfate proteoglycans in rat hepatocellular carcinoma tissues. World J. Gastroenterol. 2012, 18, 3962–3976. [Google Scholar] [CrossRef]
- Schwend, T.; Deaton, R.J.; Zhang, Y.; Caterson, B.; Conrad, G.W. Corneal sulfated glycosaminoglycans and their effects on trigeminal nerve growth cone behavior in vitro: Roles for ECM in cornea innervation. Investig. Ophthalmol. Vis. Sci. 2012, 53, 8118–8137. [Google Scholar] [CrossRef] [PubMed]
- ten Dam, G.; van de Westerlo, E.M.; Purushothaman, A.; Stan, R.V.; Bulten, J.; Sweep, F.C.; Massuger, L.F.; Sugahara, K.; van Kuppevelt, T.H. Antibody GD3G7 selected against embryonic glycosaminoglycans defines chondroitin sulfate-E domains highly up-regulated in ovarian cancer and involved in vascular endothelial growth factor binding. Am. J. Pathol. 2007, 171, 1324–1333. [Google Scholar] [CrossRef] [PubMed]
- Thompson, S.; Connell, M.G.; van Kuppevelt, T.H.; Xu, R.; Turnbull, J.E.; Losty, P.D.; Fernig, D.G.; Jesudason, E.C. Structure and epitope distribution of heparan sulfate is disrupted in experimental lung hypoplasia: A glycobiological epigenetic cause for malformation? BMC Dev. Biol. 2011, 11, 38. [Google Scholar] [CrossRef]
- Yamada, K.; Young, R.D.; Lewis, P.N.; Shinomiya, K.; Meek, K.M.; Kinoshita, S.; Caterson, B.; Quantock, A.J. Mesenchymal-epithelial cell interactions and proteoglycan matrix composition in the presumptive stem cell niche of the rabbit corneal limbus. Mol. Vis. 2015, 21, 1328–1339. [Google Scholar]
- Buresh-Stiemke, R.; Malinowski, R.L.; Keil, K.P.; Vezina, C.M.; Oosterhof, A.; Van Kuppevelt, T.H.; Marker, P.C. Distinct expression patterns of Sulf1 and Hs6st1 spatially regulate heparan sulfate sulfation during prostate development. Dev. Dyn. 2012, 241, 2005–2013. [Google Scholar] [CrossRef]
- Hossain, M.; Hosono-Fukao, T.; Tang, R.; Sugaya, N.; van Kuppevelt, T.H.; Jenniskens, G.J.; Kimata, K.; Rosen, S.D.; Uchimura, K. Direct detection of HSulf-1 and HSulf-2 activities on extracellular heparan sulfate and their inhibition by PI-88. Glycobiology 2010, 20, 175–186. [Google Scholar] [CrossRef]
- McCarthy, K.; Wassenhove-McCarthy, D.J. The glomerular basement membrane as a model system to study the bioactivity of heparan sulfate glycosaminoglycans. Microsc. Microanal. 2012, 18, 3–21. [Google Scholar] [CrossRef] [PubMed]
- Nagamine, S.; Tamba, M.; Ishimine, H.; Araki, K.; Shiomi, K.; Okada, T.; Ohto, T.; Kunita, S.; Takahashi, S.; Wismans, R.G.; et al. Organ-specific sulfation patterns of heparan sulfate generated by extracellular sulfatases Sulf1 and Sulf2 in mice. J. Biol. Chem. 2012, 287, 9579–9590. [Google Scholar] [CrossRef] [PubMed]
- Rogers, C.; Clark, P.M.; Tully, S.E.; Abrol, R.; Garcia, K.C.; Goddard, W.A., III; Hsieh-Wilson, L.C. Elucidating glycosaminoglycan-protein-protein interactions using carbohydrate microarray and computational approaches. Proc. Nat. Acad. Sci. USA 2011, 108, 9747–9752. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Zhang, L.; Stevens, J.; Gibson, G. CRISPR/Cas9 mediated generation of stable chondrocyte cell lines with targeted gene knockouts; analysis of an aggrecan knockout cell line. Bone 2014, 69, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Schuh, R.; Gonzalez, E.A.; Tavares, A.M.V.; Seolin, B.G.; Elias, L.S.; Vera, L.N.P.; Kubaski, F.; Poletto, E.; Giugliani, R.; Teixeira, H.F.; et al. Neonatal nonviral gene editing with the CRISPR/Cas9 system improves some cardiovascular, respiratory, and bone disease features of the mucopolysaccharido-sis I phenotype in mice. Gene Ther. 2020, 27, 74–84. [Google Scholar] [CrossRef]
- Feng, S.; Gao, Y.; Yin, D.; Lv, L.; Wen, Y.; Li, Z.; Wang, B.; Wu, M.; Liu, B. Identification of Lumican and Fibromodulin as Hub Genes Associated with Accumulation of Extracellular Matrix in Diabetic Nephropathy. Kidney Blood Press. Res. 2021, 46, 275–285. [Google Scholar] [CrossRef]
- Maciej-Hulme, M.; Melrose, J.; Farrugia, B.L. Arthritis and Duchenne Muscular Dystrophy: The role of chondroitin sulfate and its associated proteoglycans in disease pathology and as a diagnostic marker. Am. J. Physiol. Cell Physiol. 2022. [Google Scholar] [CrossRef]
- Cummings, R. The repertoire of glycan determinants in the human glycome. Mol. Biosyst. 2009, 5, 1087–1104. [Google Scholar] [CrossRef]
- Gomes, A.; Stelling, M.P.; Pavao, M.S. Heparan sulfate and heparanase as modulators of breast cancer progression. Biomed Res. Int. 2013, 2013, 852093. [Google Scholar] [CrossRef]
- Wang, A.; Sankaranarayanan, N.V.; Yanagashita, M.; Templeton, D.M.; Desai, U.R.; Sugahara, K.; Wang, C.P.; Hascall, V.C. Heparin interaction with a receptor on hyperglycemic dividing cells prevents intracellular hyaluronan synthesis and autophagy responses in models of type I diabetes. Matrix Biol. 2015, 48, 36–41. [Google Scholar] [CrossRef]
- Guglier, S.; Hricovíni, M.; Raman, R.; Polito, L.; Torri, G.; Casu, B.; Sasisekharan, R.; Guerrini, M. Minimum FGF2 binding structural requirements of heparin and heparan sulfate oligosaccharides as determined by NMR spectroscopy. Biochemistry 2008, 47, 13862–13869. [Google Scholar] [CrossRef] [PubMed]
- Ori, A.; Wilkinson, M.C.; Fernig, D.G. The heparanome and regulation of cell function: Structures, functions and challenges. Front. Biosci. 2008, 13, 4309–4338. [Google Scholar] [CrossRef] [PubMed]
- Clementz, A.; Harris, A. Collagen XV: Exploring its structure and role within the tumor microenvironment. Mol. Cancer Res. 2013, 11, 1481–1486. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.C.; Amenta, P.S.; Dion, A.S.; Sciancalepore, J.P.; Nagaswami, C.; Weisel, J.W.; Yurchenco, P.D. The molecular structure of human tissue type XV presents a unique conformation among the collagens. Biochem. J. 2007, 404, 535–544. [Google Scholar] [CrossRef]
- Kawashima, H.; Watanabe, N.; Hirose, M.; Sun, X.; Atarashi, K.; Kimura, T.; Shikata, K.; Matsuda, M.; Ogawa, D.; Heljasvaara, R.; et al. Collagen XVIII, a basement membrane heparan sulfate proteoglycan, interacts with L-selectin and monocyte chemoattractant protein-1. J. Biol. Chem. 2003, 278, 13069–13076. [Google Scholar] [CrossRef]
- Marneros, A.G.; Olsen, B.R. Physiological role of collagen XVIII and endostatin. FASEB J. 2005, 19, 716–728. [Google Scholar] [CrossRef]
- Cole, G.J.; Halfter, W. Agrin: An extracellular matrix heparan sulfate proteoglycan involved in cell interactions and synaptogenesis. Perspect. Dev. Neurobiol. 1996, 3, 359–371. [Google Scholar]
- Groffen, A.J.; Buskens, C.A.; van Kuppevelt, T.H.; Veerkamp, J.H.; Monnens, L.A.; van den Heuvel, L.P. Primary structure and high expression of human agrin in basement membranes of adult lung and kidney. Eur. J. Biochem. 1998, 254, 123–128. [Google Scholar] [CrossRef]
- Groffen, A.J.; Ruegg, M.A.; Dijkman, H.; van de Velden, T.J.; Buskens, C.A.; van den Born, J.; Assmann, K.J.; Monnens, L.A.; Veerkamp, J.H.; van den Heuvel, L.P. Agrin is a major heparan sulfate proteoglycan in the human glomerular basement membrane. J. Histochem. Cytochem. 1998, 46, 19–27. [Google Scholar] [CrossRef]
- Halfter, W.; Dong, S.; Schurer, B.; Cole, G.J. Collagen XVIII is a basement membrane heparan sulfate proteoglycan. J. Biol. Chem. 1998, 273, 25404–25412. [Google Scholar] [CrossRef]
- Halfter, W.; Schurer, B.; Yip, J.; Yip, L.; Tsen, G.; Lee, J.A.; Cole, G.J. Distribution and substrate properties of agrin, a heparan sulfate proteoglycan of developing axonal pathways. J. Comp. Neurol. 1997, 383, 1–17. [Google Scholar] [CrossRef]
- Kirn-Safran, C.; Farach-Carson, M.C.; Carson, D.D. Multifunctionality of extracellular and cell surface heparan sulfate proteoglycans. Cell. Mol. Life Sci. 2009, 66, 3421–3434. [Google Scholar] [CrossRef] [PubMed]
- Tsen, G.; Halfter, W.; Kroger, S.; Cole, G.J. Agrin is a heparan sulfate proteoglycan. J. Biol. Chem. 1995, 270, 3392–3399. [Google Scholar] [CrossRef] [PubMed]
- Verbeek, M.M.; Otte-Holler, I.; van den Born, J.; van den Heuvel, L.P.; David, G.; Wesseling, P.; de Waal, R.M. Agrin is a major heparan sulfate proteoglycan accumulating in Alzheimer’s disease brain. Am. J. Pathol. 1999, 155, 2115–2125. [Google Scholar] [CrossRef] [PubMed]
- Melrose, J.; Roughley, P.; Knox, S.; Smith, S.; Lord, M.; Whitelock, J. The structure, location, and function of perlecan, a prominent pericellular proteoglycan of fetal, postnatal, and mature hyaline cartilages. J. Biol. Chem. 2006, 281, 36905–36914. [Google Scholar] [CrossRef]
- Iozzo, R.V.; Schaefer, L. Proteoglycan form and function: A comprehensive nomenclature of proteoglycans. Matrix Biol. 2015, 42, 11–55. [Google Scholar] [CrossRef]
- De Cat, B.; David, G. Developmental roles of the glypicans. In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2001; Volume 12, pp. 117–125. [Google Scholar]
- Filmus, J.; Capurro, M.; Rast, J. Glypicans. Genome Biol. 2008, 9, 224. [Google Scholar] [CrossRef]
- Fransson, L. Glypicans. Int. J. Biochem. Cell Biol. 2003, 35, 125–129. [Google Scholar] [CrossRef]
- Song, H.; Filmus, J. The role of glypicans in mammalian development. Biochim. Biophys. Acta 2002, 1573, 241–246. [Google Scholar] [CrossRef]
- Cheng, B.; Montmasson, M.; Terradot, L.; Rousselle, P. Syndecans as Cell Surface Receptors in Cancer Biology. A Focus on their Interaction with PDZ Domain Proteins. Front. Pharmacol. 2016, 7, 10. [Google Scholar] [CrossRef]
- Choi, S.; Kang, D.H.; Oh, E.S. Targeting syndecans: A promising strategy for the treatment of cancer. Expert Opin. Ther. Targets 2013, 17, 695–705. [Google Scholar] [CrossRef]
- Couchman, J.R.; Gopal, S.; Lim, H.C.; Norgaard, S.; Multhaupt, H.A. Fell-Muir Lecture: Syndecans: From peripheral coreceptors to mainstream regulators of cell behaviour. Int. J. Exp. Pathol. 2015, 96, 1–10. [Google Scholar] [CrossRef]
- Fears, C.Y.; Woods, A. The role of syndecans in disease and wound healing. Matrix Biol. 2006, 25, 443–456. [Google Scholar] [CrossRef]
- Afratis, N.; Nikitovic, D.; Multhaupt, H.A.; Theocharis, A.D.; Couchman, J.R.; Karamanos, N.K. Syndecans-key regulators of cell signaling and biological functions. FEBS J. 2017, 284, 27–41. [Google Scholar] [CrossRef]
- Xian, X.; Gopal, S.; Couchman, J.R. Syndecans as receptors and organizers of the extracellular matrix. Cell Tissue Res. 2010, 339, 31–46. [Google Scholar] [CrossRef]
- Bilandzic, M.; Stenvers, K.L. Betaglycan: A multifunctional accessory. Mol. Cell. Endocrinol. 2011, 339, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Gray, P.C.; Bilezikjian, L.M.; Vale, W. Antagonism of activin by inhibin and inhibin receptors: A functional role for betaglycan-glycan. Mol. Cell. Endocrinol. 2001, 180, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Grimme, H.U.; Termeer, C.C.; Bennett, K.L.; Weiss, J.M.; Schopf, E.; Aruffo, A.; Simon, J.C. Colocalization of basic fibroblast growth factor and CD44 isoforms containing the variably spliced exon v3 (CD44v3) in normal skin and in epidermal skin cancers. Br. J. Dermatol. 1999, 141, 824–832. [Google Scholar] [CrossRef] [PubMed]
- Suga, N.; Sugimura, M.; Koshiishi, T.; Yorifuji, T.; Makino, S.; Takeda, S. Heparin/heparan sulfate/CD44-v3 enhances cell migration in term placenta-derived immortalized human trophoblast cells. Biol. Reprod. 2012, 86, 134–138. [Google Scholar] [CrossRef]
- Brown, T.; Bouchard, T.; St John, T.; Wayner, E.; Carter, W.G. Human keratinocytes express a new CD44 core protein (CD44E) as a heparan-sulfate intrinsic membrane proteoglycan with additional exons. J. Cell Biol. 1991, 113, 207–221. [Google Scholar] [CrossRef]
- Kugelman, L.; Ganguly, S.; Haggerty, J.G.; Weissman, S.M.; Milstone, L.M. The core protein of epican, a heparan sulfate proteoglycan on keratinocytes, is an alternative form of CD44. J. Investig. Dermatol. 1992, 99, 886–891. [Google Scholar] [CrossRef] [PubMed]
- Milstone, L.; Hough-Monroe, L.; Kugelman, L.C.; Bender, J.R.; Haggerty, J.G. Epican, a heparan/chondroitin sulfate proteoglycan form of CD44, mediates cell-cell adhesion. J. Cell Sci. 1994, 107, 3183–3190. [Google Scholar] [CrossRef]
- Zhou, J.; Haggerty, J.G.; Milstone, L.M. Growth and differentiation regulate CD44 expression on human keratinocytes. Vitro Cell. Dev. Biol. Anim. 1999, 35, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Ahrens, T.; Bang-Christensen, S.R.; Jørgensen, A.M.; Løppke, C.; Spliid, C.B.; Sand, N.T.; Clausen, T.M.; Salanti, A.; Agerbæk, M.Ø. The Role of Proteoglycans in Cancer Metastasis and Circulating Tumor Cell Analysis. Front. Cell Dev. Biol. 2020, 8, 749. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Kuznetsova, S.A.; Pendrak, M.L.; Sipes, J.M.; Romeo, M.J.; Li, Z.; Zhang, L.; Roberts, D.D. Heparan sulfate modification of the transmembrane receptor CD47 is necessary for inhibition of T cell receptor signaling by thrombospondin-1. J. Biol. Chem. 2011, 286, 14991–15002. [Google Scholar] [CrossRef]
- Frankel, P.; Pellet-Many, C.; Lehtolainen, P.; D’Abaco, G.M.; Tickner, M.L.; Cheng, L.; Zachary, I.C. Chondroitin sulfate-modified neuropilin 1 is expressed in human tumour cells and modulates 3D invasion in the U87MG human glioblastoma cell line through a p130Cas-mediated pathway. EMBO Rep. 2008, 9, 983–989. [Google Scholar] [CrossRef] [PubMed]
- Shintani, Y.; Takashima, S.; Asano, Y.; Kato, H.; Liao, Y.; Yamazaki, S.; Tsukamoto, O.; Seguchi, O.; Yamamoto, H.; Fukushima, T.; et al. Glycosaminoglycan modification of neuropilin-1 modulates VEGFR2 signaling. EMBO J. 2006, 25, 3045–3055. [Google Scholar] [CrossRef]
- Kolset, S.O.; Tveit, H. Serglycin--structure and biology. Cell. Mol. Life Sci. 2008, 65, 1073–1085. [Google Scholar] [CrossRef] [PubMed]
- Li, X.J.; Qian, C.N. Serglycin in human cancers. Chin. J. Cancer 2011, 30, 585–589. [Google Scholar] [CrossRef][Green Version]
- Scully, O.J.; Chua, P.J.; Harve, K.S.; Bay, B.H.; Yip, G.W. Serglycin in health and diseases. Anat. Rec. 2012, 295, 1415–1420. [Google Scholar] [CrossRef]
- Suzuki, K.; Yuzaki, M. La Dolce Vita of Neurexin: Synaptic Partnerships through Glycosaminoglycans. Cell 2018, 174, 1337–1338. [Google Scholar] [CrossRef]
- Farfán, N.; Orellana-Serradell, O.; Herrera, D.; Chrzanowsky, D.; Cubillos, P.; Marín, G.; Antonio García De Herreros, A.; Castellón, E.A.; Contreras, H.R. SNAIL expression correlates with the translocation of syndecan-1 intracellular domain into the nucleus in prostate cancer cell lines. Int. J. Mol. Med. 2020, 45, 1073–1080. [Google Scholar] [PubMed]
- Hayes, A.; Melrose, J. 3D distribution of perlecan within intervertebral disc chondrons suggests novel regulatory roles for this multifunctional modular heparan sulfate proteoglycan. Eur. Cells Mater. 2021, 41, 73–89. [Google Scholar] [CrossRef]
- Hayes, A.; Melrose, J. What Are the Potential Roles of Nuclear Perlecan and Other Heparan Sulfate Proteoglycans in the Normal and Malignant Phenotype. Int. J. Mol. Sci. 2021, 22, 4415. [Google Scholar] [CrossRef]
- Kumar-Singh, A.; Shrinet, J.; Parniewska, M.M.; Fuxe, J.; Dobra, K.; Hjerpe, A. Mapping the Interactome of the Nuclear Heparan Sulfate Proteoglycan Syndecan-1 in Mesothelioma Cells. Biomolecules 2020, 10, 1034. [Google Scholar] [CrossRef] [PubMed]
- Stewart, M.D.; Sanderson, R.D. Heparan sulfate in the nucleus and its control of cellular functions. Matrix Biol. 2014, 35, 56–59. [Google Scholar] [CrossRef] [PubMed]
- Mythreye, K.; Blobe, G.C. Proteoglycan signaling co-receptors: Roles in cell adhesion, migration and invasion. Cell. Signal. 2009, 21, 1548–1558. [Google Scholar] [CrossRef] [PubMed]
- Bernard, D.; Smith, C.L.; Brûlé, E. A Tale of Two Proteins: Betaglycan, IGSF1, and the Continuing Search for the Inhibin B Receptor. Trends Endocrinol. Metab. 2020, 31, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Boyd, F.; Cheifetz, S.; Andres, J.; Laiho, M.; Massagué, J. Transforming growth factor-beta receptors and binding proteoglycans. J. Cell Sci. 1990, 13, 131–138. [Google Scholar] [CrossRef]
- Kim, S.; Henen, M.A.; Hinck, A.P. Structural biology of betaglycan and endoglin, membrane-bound co-receptors of the TGF-beta family. Exp. Biol. Med. 2019, 244, 1547–1558. [Google Scholar] [CrossRef]
- Lewis, K.; Gray, P.C.; Blount, A.L.; MacConell, L.A.; Wiater, E.; Bilezikjian, L.M.; Vale, W. Betaglycan binds inhibin and can mediate functional antagonism of activin signalling. Nature 2000, 404, 411–414. [Google Scholar] [CrossRef]
- Massagué, J.; Andres, J.; Attisano, L.; Cheifetz, S.; López-Casillas, F.; Ohtsuki, M.; Wrana, J.L. TGF-beta receptors. Mol. Reprod. Dev. 1992, 32, 99–104. [Google Scholar] [CrossRef]
- Miyazono, K. TGF-beta receptors and signal transduction. Int. J. Hematol. 1997, 65, 97–104. [Google Scholar] [CrossRef]
- Sandbrink, R.; Masters, C.L.; Beyreuther, K. APP gene family. Alternative splicing generates functionally related isoforms. Ann. New York Acad. Sci. 1996, 777, 281–287. [Google Scholar] [CrossRef]
- Segarini, P. TGF-beta receptors. Ciba. Found Symp. 1991, 157, 29–50. [Google Scholar]
- Jenkins, L.; Horst, B.; Lancaster, C.L.; Mythreye, K. Dually modified transmembrane proteoglycans in development and disease. Cytokine Growth Factor Rev. 2018, 39, 124–136. [Google Scholar] [CrossRef]
- Jenkins, L.; Singh, P.; Varadaraj, A.; Lee, N.Y.; Shah, S.; Flores, H.V.; O’Connell, K.; Mythreye, K. Altering the Proteoglycan State of Transforming Growth Factor β Type III Receptor (TβRIII)/Betaglycan Modulates Canonical Wnt/β-Catenin Signaling. J. Biol. Chem. 2016, 291, 25716–25728. [Google Scholar] [CrossRef]
- Pantazaka, E.; Papadimitriou, E. Chondroitin sulfate-cell membrane effectors as regulators of growth factor-mediated vascular and cancer cell migration. Biochim. Biophys. Acta 2014, 1840, 2643–2650. [Google Scholar] [CrossRef]
- Makanji, Y.; Harrison, C.A.; Stanton, P.G.; Krishna, R.; Robertson, D.M. Inhibin A and B in vitro bioactivities are modified by their degree of glycosylation and their affinities to betaglycan. Endocrinology 2007, 148, 2309–2316. [Google Scholar] [CrossRef]
- Miller, M.; Lambert-Messerlian, G.M.; Eklund, E.E.; Heath, N.L.; Donahue, J.E.; Stopa, E.G. Expression of inhibin/activin proteins and receptors in the human hypothalamus and basal forebrain. J. Neuroendocrinol. 2012, 24, 962–972. [Google Scholar] [CrossRef]
- MacConell, L.; Leal, A.M.; Vale, W.W. The distribution of betaglycan protein and mRNA in rat brain, pituitary, and gonads: Implications for a role for betaglycan in inhibin-mediated reproductive functions. Endocrinology 2002, 143, 1066–1075. [Google Scholar] [CrossRef][Green Version]
- Knelson, E.; Gaviglio, A.L.; Tewari, A.K.; Armstrong, M.B.; Mythreye, K.; Blobe, G.C. Type III TGF-β receptor promotes FGF2-mediated neuronal differentiation in neuroblastoma. J. Clin. Investig. 2013, 123, 4786–4798. [Google Scholar] [CrossRef]
- Battista, D.; Ferrari, C.C.; Gage, F.H.; Pitossi, F.J. Neurogenic niche modulation by activated microglia: Transforming growth factor beta increases neurogenesis in the adult dentate gyrus. Eur. J. Neurosci. 2006, 23, 83–93. [Google Scholar] [CrossRef]
- Mathieu, P.; Piantanida, A.P.; Pitossi, F. Chronic expression of transforming growth factor-beta enhances adult neurogenesis. Neuroimmunomodulation 2010, 17, 200–201. [Google Scholar] [CrossRef]
- Vale, W.; Wiater, E.; Gray, P.; Harrison, C.; Bilezikjian, L.; Choe, S. Activins and inhibins and their signaling. Ann. N. Y. Acad. Sci. 2004, 1038, 142–147. [Google Scholar] [CrossRef]
- Florio, P.; Gazzolo, D.; Luisi, S.; Petraglia, F. Activin A in brain injury. Adv. Clin. Chem. 2007, 43, 117–130. [Google Scholar]
- Woodruff, T. Regulation of cellular and system function by activin. Biochem. Pharmacol. 1998, 55, 953–963. [Google Scholar] [CrossRef]
- Alzheimer, C.; Werner, S. Fibroblast growth factors and neuroprotection. Adv. Exp. Med. Biol. 2002, 513, 335–351. [Google Scholar]
- Barclay, A.; Van den Berg, T.K. The interaction between signal regulatory protein alpha (SIRPα) and CD47: Structure, function, and therapeutic target. Annu. Rev. Immunol. 2014, 32, 25–50. [Google Scholar] [CrossRef]
- Murata, Y.; Kotani, T.; Ohnishi, H.; Matozaki, T. The CD47-SIRPα signalling system: Its physiological roles and therapeutic application. J. Biochem. 2014, 155, 335–344. [Google Scholar] [CrossRef]
- Bauer, E.; Qin, Y.; Miller, T.W.; Bandle, R.W.; Csanyi, G.; Pagano, P.J.; Bauer, P.M.; Schnermann, J.; Roberts, D.D.; Isenberg, J.S. Thrombospondin-1 supports blood pressure by limiting eNOS activation and endothelial-dependent vasorelaxation. Cardiovasc. Res. 2010, 88, 471–481. [Google Scholar] [CrossRef]
- Matlung, H.; Szilagyi, K.; Barclay, N.A.; van den Berg, T.K. The CD47-SIRPα signaling axis as an innate immune checkpoint in cancer. Immunol. Rev. 2017, 276, 145–164. [Google Scholar] [CrossRef]
- Weiskopf, K. Cancer immunotherapy targeting the CD47/SIRPα axis. Eur. J. Cancer 2017, 76, 100–109. [Google Scholar] [CrossRef]
- Zhang, H.; Li, F.; Yang, Y.; Chen, J.; Hu, X. SIRP/CD47 signaling in neurological disorders. Brain Res. 2015, 1623, 74–80. [Google Scholar] [CrossRef]
- Li, Z.; Li, Y.; Gao, J.; Fu, Y.; Hua, P.; Jing, Y.; Cai, M.; Wang, H.; Tong, T. The role of CD47-SIRPα immune checkpoint in tumor immune evasion and innate immunotherapy. Life Sci. 2021, 273, 119150. [Google Scholar] [CrossRef]
- Gholamin, S.; Mitra, S.S.; Feroze, A.H.; Liu, J.; Kahn, S.A.; Zhang, M.; Esparza, R.; Richard, C.; Ramaswamy, V.; Remke, M.; et al. Disrupting the CD47-SIRPα anti-phagocytic axis by a humanized anti-CD47 antibody is an efficacious treatment for malignant pediatric brain tumors. Sci. Transl. Med. 2017, 9, eaaf2968. [Google Scholar] [CrossRef]
- Hu, J.; Xiao, Q.; Dong, M.; Guo, D.; Wu, X.; Wang, B. Glioblastoma Immunotherapy Targeting the Innate Immune Checkpoint CD47-SIRPα Axis. Front. Immunol. 2020, 11, 593219. [Google Scholar] [CrossRef]
- Hutter, G.; Theruvath, J.; Graef, C.M.; Zhang, M.; Schoen, M.K.; Manz, E.M.; Bennett, M.L.; Olson, A.; Azad, T.D.; Sinha, R.; et al. Microglia are effector cells of CD47-SIRPα antiphagocytic axis disruption against glioblastoma. Proc. Natl. Acad. Sci. USA 2019, 116, 997–1006. [Google Scholar] [CrossRef]
- Kuo, T.; Chen, A.; Harrabi, O.; Sockolosky, J.T.; Zhang, A.; Sangalang, E.; Doyle, L.V.; Kauder, S.E.; Fontaine, D.; Bollini, S.; et al. Targeting the myeloid checkpoint receptor SIRPα potentiates innate and adaptive immune responses to promote anti-tumor activity. J. Hematol. Oncol. 2020, 13, 160. [Google Scholar] [CrossRef]
- Zhang, W.; Huang, Q.; Xiao, W.; Zhao, Y.; Pi, J.; Xu, H.; Zhao, H.; Xu, J.; Evans, C.E.; Jin, H. Advances in Anti-Tumor Treatments Targeting the CD47/SIRPα Axis. Front. Immunol. 2020, 11, 18. [Google Scholar] [CrossRef]
- Jeanne, A.; Sarazin, T.; Charlé, M.; Kawecki, C.; Kauskot, A.; Hedtke, T.; Schmelzer, C.E.H.; Martiny, L.; Maurice, P.; Dedieu, S. Towards the Therapeutic Use of Thrombospondin 1/CD47 Targeting TAX2 Peptide as an Antithrombotic Agent. Arterioscler. Thromb. Vasc. Biol. 2021, 41, e1–e17. [Google Scholar] [CrossRef]
- Painter, C.; Clausen, T.M.; Park, J.; Vasquez, G.J.; Jian Liu, J.; Gordts, P.L.S.M.; Esko, J.D. Molecular Interaction of Neuropilin-1 and 3-O-sulfated Heparan Sulfate Modulates Angiogenesis-dependent Tumor Growth. FASEB J. 2020, 34, 1. [Google Scholar] [CrossRef]
- Mamluk, R.; Gechtman, Z.; Kutcher, M.E.; Gasiunas, N.; Gallagher, J.; Klagsbrun, M. Neuropilin-1 binds vascular endothelial growth factor 165, placenta growth factor-2, and heparin via its b1b2 domain. J. Biol. Chem. 2002, 277, 24818–24825. [Google Scholar] [CrossRef]
- West, D.; Rees, C.G.; Duchesne, L.; Patey, S.J.; Terry, C.J.; Turnbull, J.E.; Delehedde, M.; Heegaard, C.W.; Allain, F.; Vanpouille, C.; et al. Interactions of multiple heparin binding growth factors with neuropilin-1 and potentiation of the activity of fibroblast growth factor-2. J. Biol. Chem. 2005, 280, 13457–13464. [Google Scholar] [CrossRef]
- Tollefsen, D. Vascular dermatan sulfate and heparin cofactor II. Prog. Mol. Biol. Transl. Sci. 2010, 93, 351–372. [Google Scholar]
- Brose, N. Synaptogenic proteins and synaptic organizers: “many hands make light work”. Neuron 2009, 61, 650–652. [Google Scholar] [CrossRef]
- Varoqueaux, F.; Aramuni, G.; Rawson, R.L.; Mohrmann, R.; Missler, M.; Gottmann, K.; Zhang, W.; Südhof, T.C.; Brose, N. Neuroligins determine synapse maturation and function. Neuron 2006, 51, 741–754. [Google Scholar] [CrossRef]
- Kim, H.; Um, J.W.; Ko, J. Proper synaptic adhesion signaling in the control of neural circuit architecture and brain function. Prog. Neurobiol. 2021, 200, 101983. [Google Scholar] [CrossRef]
- Sakers, K.; Eroglu, C. Control of neural development and function by glial neuroligins. Curr. Opin. Neurobiol. 2019, 57, 163–170. [Google Scholar] [CrossRef]
- Roppongi, R.; Dhume, S.H.; Padmanabhan, N.; Silwal, P.; Zahra, N.; Karimi, B.; Bomkamp, C.; Patil, C.S.; Champagne-Jorgensen, K.; Twilley, R.E.; et al. LRRTMs Organize Synapses through Differential Engagement of Neurexin and PTPσ. Neuron 2020, 106, 108–125.e12. [Google Scholar] [CrossRef]
- Südhof, T. Synaptic Neurexin Complexes: A Molecular Code for the Logic of Neural Circuits. Cell 2017, 171, 745–769. [Google Scholar] [CrossRef] [PubMed]
- Lisé, M.; El-Husseini, A. The neuroligin and neurexin families: From structure to function at the synapse. Cell. Mol. Life Sci. 2006, 63, 1833–1849. [Google Scholar] [CrossRef] [PubMed]
- Cuttler, K.; Hassan, M.; Carr, J.; Cloete, R.; Bardien, S. Emerging evidence implicating a role for neurexins in neurodegenerative and neuropsychiatric disorders. Open Biol. 2021, 11, 210091. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Tabuchi, K. Functions of synapse adhesion molecules neurexin/neuroligins and neurodevelopmental disorders. Neurosci. Res. 2017, 116, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Südhof, T. Neuroligins and neurexins link synaptic function to cognitive disease. Nature 2008, 455, 903–911. [Google Scholar] [CrossRef]
- Gauthier, J.; Siddiqui, T.J.; Huashan, P.; Yokomaku, D.; Hamdan, F.F.; Champagne, N.; Lapointe, M.; Spiegelman, D.; Noreau, A.; Lafrenière, R.G.; et al. Truncating mutations in NRXN2 and NRXN1 in autism spectrum disorders and schizophrenia. Hum. Genet. 2011, 130, 563–573. [Google Scholar] [CrossRef]
- Uchigashima, M.; Cheung, A.; Futai, K. Neuroligin-3: A Circuit-Specific Synapse Organizer That Shapes Normal Function and Autism Spectrum Disorder-Associated Dysfunction. Front. Mol. Neurosci. 2021, 14, 749164. [Google Scholar] [CrossRef]
- Oguro, K.; Shimazaki, K.; Yokota, H.; Onuki, Y.; Murashima, Y.; Kawai, K.; Muramatsu, S.I. Global brain delivery of neuroligin 2 gene ameliorates seizures in a mouse model of epilepsy. J. Gene Med. 2022, 24, e3402. [Google Scholar] [CrossRef]
- Blundell, J.; Tabuchi, K.; Bolliger, M.F.; Blaiss, C.A.; Brose, N.; Liu, X.; Südhof, T.C.; Powell, C.M. Increased anxiety-like behavior in mice lacking the inhibitory synapse cell adhesion molecule neuroligin 2. Genes Brain Behav. 2009, 8, 114–126. [Google Scholar] [CrossRef]
- Maćkowiak, M.; Mordalska, P.; Wędzony, K. Neuroligins, synapse balance and neuropsychiatric disorders. Pharmacol. Rep. 2014, 66, 830–835. [Google Scholar] [CrossRef]
- Trobiani, L.; Meringolo, M.; Diamanti, T.; Bourne, Y.; Marchot, P.; Martella, G.; Dini, L.; Pisani, A.; De Jaco, A.; Bonsi, P. The neuroligins and the synaptic pathway in Autism Spectrum Disorder. Neurosci. Biobehav. Rev. 2020, 119, 37–51. [Google Scholar] [CrossRef]
- Lai, E.; Nakayama, H.; Miyazaki, T.; Nakazawa, T.; Tabuchi, K.; Hashimoto, K.; Watanabe, M.; Kano, M. An Autism-Associated Neuroligin-3 Mutation Affects Developmental Synapse Elimination in the Cerebellum. Front. Neural Circuits 2021, 15, 676891. [Google Scholar] [CrossRef]
- Wang, J.; Gong, J.; Li, L.; Chen, Y.; Liu, L.; Gu, H.; Luo, X.; Hou, F.; Zhang, J.; Song, R. Neurexin gene family variants as risk factors for autism spectrum disorder. Autism. Res. 2018, 11, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Snow, A.; Cummings, J.A.; Lake, T. The Unifying Hypothesis of Alzheimer’s Disease: Heparan Sulfate Proteoglycans/Glycosaminoglycans Are Key as First Hypothesized Over 30 Years Ago. Front. Aging Neurosci. 2021, 13, 710683. [Google Scholar] [CrossRef] [PubMed]
- Kanagawa, M.; Omori, Y.; Sato, S.; Kobayashi, K.; Miyagoe-Suzuki, Y.; Takeda, S.; Endo, T.; Furukawa, T.; Toda, T. Post-translational maturation of dystroglycan is necessary for pikachurin binding and ribbon synaptic localization. J. Biol. Chem. 2010, 285, 31208–31216. [Google Scholar] [CrossRef]
- Hu, H.; Li, J.; Zhang, Z.; Yu, M. Pikachurin interaction with dystroglycan is diminished by defective O-mannosyl glycosylation in congenital muscular dystrophy models and rescued by LARGE overexpression. Neurosci. Lett. 2011, 489, 10–15. [Google Scholar] [CrossRef]
- Han, J.; Townes-Anderson, E. Cell specific post-translational processing of pikachurin, a protein involved in retinal synaptogenesis. PLoS ONE 2012, 7, e50552. [Google Scholar] [CrossRef] [PubMed]
- Wheway, G.; Parry, D.A.; Johnson, C.A. The role of primary cilia in the development and disease of the retina. Organogenesis 2014, 10, 69–85. [Google Scholar] [CrossRef]
- McGuigan, D.; Heon, E.; Cideciyan, A.V.; Ratnapriya, R.; Lu, M.; Sumaroka, A.; Roman, A.J.; Batmanabane, V.; Garafalo, A.V.; Stone, E.M.; et al. EYS Mutations Causing Autosomal Recessive Retinitis Pigmentosa: Changes of Retinal Structure and Function with Disease Progression. Genes 2017, 8, 178. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Delgado, A.; Valdes-Sanchez, L.; Morillo-Sanchez, M.J.; Ponte-Zuñiga, B.; Diaz-Corrales, F.J.; de la Cerda, B. Dissecting the role of EYS in retinal degeneration: Clinical and molecular aspects and its implications for future therapy. Orphanet J. Rare Dis. 2021, 16, 222. [Google Scholar] [CrossRef] [PubMed]
- Couchman, J. Syndecans: Proteoglycan regulators of cell-surface microdomains? Nat. Rev. Mol. Cell Biol. 2003, 4, 926–937. [Google Scholar] [CrossRef]
- Couchman, J. Syndecan-1 (CD138), Carcinomas and EMT. Int. J. Mol. Sci. 2021, 22, 4227. [Google Scholar] [CrossRef]
- Farach-Carson, M.; Hecht, J.T.; Carson, D.D. Heparan sulfate proteoglycans: Key players in cartilage biology. Crit. Rev. Eukaryot. Gene Expr. 2005, 15, 29–48. [Google Scholar] [CrossRef]
- Farach-Carson, M.; Carson, D.D. Perlecan--a multifunctional extracellular proteoglycan scaffold. Glycobiology 2007, 17, 897–905. [Google Scholar] [CrossRef]
- Gopal, S.; Arokiasamy, S.; Pataki, C.; Whiteford, J.R.; Couchman, J.R. Syndecan receptors: Pericellular regulators in development and inflammatory disease. Open Biol. 2021, 11, 200377. [Google Scholar] [CrossRef]
- Hayes, A.; Farrugia, B.L.; Biose, I.J.; Bix, G.J.; Melrose, J. Perlecan, A Multi-Functional, Cell-Instructive, Matrix-Stabilizing Proteoglycan With Roles in Tissue Development Has Relevance to Connective Tissue Repair and Regeneration. Front. Cell Dev. Biol. 2022, 10, 856261. [Google Scholar] [CrossRef]
- Multhaupt, H.; Yoneda, A.; Whiteford, J.R.; Oh, E.S.; Lee, W.; Couchman, J.R. Syndecan signaling: When, where and why? J. Physiol. Pharmacol. 2009, 60 (Suppl. S4), 31–38. [Google Scholar]
- Pan, J.; Ho, M. Role of glypican-1 in regulating multiple cellular signaling pathways. Am. J. Physiol. Cell Physiol. 2021, 321, C846–C858. [Google Scholar] [CrossRef]
- Pataki, C.; Couchman, J.R.; Brábek, J. Wnt Signaling Cascades and the Roles of Syndecan Proteoglycans. J. Histochem. Cytochem. 2015, 63, 465–480. [Google Scholar] [CrossRef]
- Karamanos, N.; Piperigkou, Z.; Theocharis, A.D.; Watanabe, H.; Franchi, M.; Baud, S.; Brézillon, S.; Götte, M.; Passi, A.; Vigetti, D.; et al. Proteoglycan Chemical Diversity Drives Multifunctional Cell Regulation and Therapeutics. Chem. Rev. 2018, 118, 9152–9232. [Google Scholar] [CrossRef]
- Kamimura, K.; Maeda, N. Glypicans and Heparan Sulfate in Synaptic Development, Neural Plasticity, and Neurological Disorders. Front. Neural Circuits 2021, 15, 595596. [Google Scholar] [CrossRef] [PubMed]
- Ivins, J.; Litwack, E.D.; Kumbasar, A.; Stipp, C.S.; Lander, A.D. Cerebroglycan, a developmentally regulated cell-surface heparan sulfate proteoglycan, is expressed on developing axons and growth cones. Dev. Biol. 1997, 184, 320–332. [Google Scholar] [CrossRef] [PubMed]
- Stipp, C.; Litwack, E.D.; Lander, A.D. Cerebroglycan: An integral membrane heparan sulfate proteoglycan that is unique to the developing nervous system and expressed specifically during neuronal differentiation. J. Cell Biol. 1994, 124, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Lander, A.; Stipp, C.S.; Ivins, J.K. The glypican family of heparan sulfate proteoglycans: Major cell-surface proteoglycans of the developing nervous system. Perspect. Dev. Neurobiol. 1996, 3, 347–358. [Google Scholar]
- Bisiak, F.; McCarthy, A.A. Structure and Function of Roundabout Receptors. In: Harris, J., Marles-Wright, J. (eds) Macromolecular Protein Complexes II: Structure and Function. Subcell. Biochem. 2019, 93, 291–319. [Google Scholar] [CrossRef]
- Blockus, H.; Chédotal, A. Slit-Robo signaling. Development 2016, 143, 3037–3044. [Google Scholar] [CrossRef]
- Kastenhuber, E.; Kern, U.; Bonkowsky, J.L.; Chien, C.B.; Driever, W.; Schweitzer, J. Netrin-DCC, Robo-Slit, and heparan sulfate proteoglycans coordinate lateral positioning of longitudinal dopaminergic diencephalospinal axons. J. Neurosci. 2009, 29, 8914–8926. [Google Scholar] [CrossRef]
- Liang, Y.; Annan, R.S.; Carr, S.A.; Popp, S.; Mevissen, M.; Margolis, R.K.; Margolis, R.U. Mammalian homologues of the Drosophila slit protein are ligands of the heparan sulfate proteoglycan glypican-1 in brain. J. Biol. Chem. 1999, 274, 17885–17892. [Google Scholar] [CrossRef]
- Ronca, F.; Andersen, J.S.; Paech, V.; Margolis, R.U. Characterization of Slit protein interactions with glypican-1. J. Biol. Chem. 2001, 276, 29141–29147. [Google Scholar] [CrossRef]
- Xiong, A.; Kundu, S.; Forsberg-Nilsson, K. Heparan sulfate in the regulation of neural differentiation and glioma development. FEBS J. 2014, 281, 4993–5008. [Google Scholar] [CrossRef]
- Condomitti, G.; de Wit, J. Heparan Sulfate Proteoglycans as Emerging Players in Synaptic Specificity. Front. Mol. Neurosci. 2018, 11, 14. [Google Scholar] [CrossRef]
- Maeda, N.; Fukazawa, N.; Ishii, M. Chondroitin sulfate proteoglycans in neural development and plasticity. Front. Biosci. 2010, 15, 626–644. [Google Scholar]
- Miyata, S.; Kitagawa, H. Mechanisms for modulation of neural plasticity and axon regeneration by chondroitin sulfate. J. Biochem. 2015, 157, 13–22. [Google Scholar] [CrossRef]
- Sakamoto, K.; Ozaki, T.; Suzuki, Y.; Kadomatsu, K. Type IIa RPTPs and Glycans: Roles in Axon Regeneration and Synaptogenesis. Int. J. Mol. Sci. 2021, 22, 5524. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, N.; Domowicz, M.S. Chemistry and Function of Glycosaminoglycans in the Nervous System. Adv. Neurobiol. 2023, 29, 117–162. [Google Scholar] [PubMed]
- Gallagher, J. Multiprotein signalling complexes: Regional assembly on heparan sulfate. Biochem. Soc. Trans. 2006, 34, 438–441. [Google Scholar] [CrossRef]
- Sorg, B.; Berretta, S.; Blacktop, J.M.; Fawcett, J.W.; Kitagawa, H.; Kwok, J.C.; Miquel, M. Casting a Wide Net: Role of Perineuronal Nets in Neural Plasticity. J. Neurosci. 2016, 36, 11459–11468. [Google Scholar] [CrossRef]
- Fawcett, J.; Fyhn, M.; Jendelova, P.; Kwok, J.C.F.; Ruzicka, J.; Sorg, B.A. The extracellular matrix and perineuronal nets in memory. Mol. Psychiatry 2022, 27, 3192–3203. [Google Scholar] [CrossRef]
- Hensch, T. Critical period plasticity in local cortical circuits. Nat. Rev. Neurosci. 2005, 6, 877–888. [Google Scholar] [CrossRef]
- Pintér, P.; Alpár, A. The Role of Extracellular Matrix in Human Neurodegenerative Diseases. Int. J. Mol. Sci. 2022, 23, 11085. [Google Scholar]
- Sethi, M.; Zaia, J. Extracellular matrix proteomics in schizophrenia and Alzheimer’s disease. Anal. Bioanal. Chem. 2017, 409, 379–394. [Google Scholar] [CrossRef]
- Gomez, A.; Traunmüller, L.; Scheiffele, P. Neurexins: Molecular codes for shaping neuronal synapses. Nat. Rev. Neurosci. 2021, 22, 137–151. [Google Scholar] [CrossRef]
- Reissner, C.; Runkel, F.; Missler, M. Neurexins. Genome Biol. 2013, 14, 213. [Google Scholar] [CrossRef] [PubMed]
- Kasem, E.; Kurihara, T.; Tabuchi, K. Neurexins and neuropsychiatric disorders. Neurosci. Res. 2018, 127, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Logsdon, A.; Francis, K.L.; Richardson, N.E.; Hu, S.J.; Faber, C.L.; Phan, B.A.; Nguyen, V.; Setthavongsack, N.; Banks, W.A.; Woltjer, R.L.; et al. Decoding perineuronal net glycan sulfation patterns in the Alzheimer’s disease brain. Alzheimers Dement. 2022, 18, 942–954. [Google Scholar] [CrossRef]
- Carulli, D.; Verhaagen, J. An Extracellular Perspective on CNS Maturation: Perineuronal Nets and the Control of Plasticity. Int. J. Mol. Sci. 2021, 22, 2434. [Google Scholar] [CrossRef]
- Pantazopoulos, H.; Berretta, S. In Sickness and in Health: Perineuronal Nets and Synaptic Plasticity in Psychiatric Disorders. Neural Plast. 2016, 2016, 9847696. [Google Scholar] [CrossRef]
- Morawski, M.; Brückner, G.; Jäger, C.; Seeger, G.; Matthews, R.T.; Arendt, T. Involvement of perineuronal and perisynaptic extracellular matrix in Alzheimer’s disease neuropathology. Brain Pathol. 2012, 22, 547–561. [Google Scholar] [CrossRef]
- Reichelt, A. Is loss of perineuronal nets a critical pathological event in Alzheimer’s disease? EBioMedicine 2020, 59, 102946. [Google Scholar] [CrossRef]
- Scarlett, J.; Hu, S.J.; Alonge, K.M. The “Loss” of Perineuronal Nets in Alzheimer’s Disease: Missing or Hiding in Plain Sight? Front. Integr. Neurosci. 2022, 16, 896400. [Google Scholar] [CrossRef]
- Wen, T.; Binder, D.K.; Ethell, I.M.; Razak, K.A. The Perineuronal ‘Safety’ Net? Perineuronal Net Abnormalities in Neurological Disorders. Front. Mol. Neurosci. 2018, 11, 270. [Google Scholar] [CrossRef]
- Li, Y.; Li, Z.X.; Jin, T.; Wang, Z.Y.; Zhao, P. Tau Pathology Promotes the Reorganization of the Extracellular Matrix and Inhibits the Formation of Perineuronal Nets by Regulating the Expression and the Distribution of Hyaluronic Acid Synthases. J. Alzheimers Dis. 2017, 57, 395–409. [Google Scholar] [CrossRef] [PubMed]
- Crapser, J.; Spangenberg, E.E.; Barahona, R.A.; Arreola, M.A.; Hohsfield, L.A.; Green, K.N. Microglia facilitate loss of perineuronal nets in the Alzheimer’s disease brain. EBioMedicine 2020, 58, 102919. [Google Scholar] [CrossRef]
- Acosta, C.; Anderson, H.D.; Anderson, C.M. Astrocyte dysfunction in Alzheimer disease. J. Neurosci. Res. 2017, 95, 2430–2447. [Google Scholar] [CrossRef] [PubMed]
- Nanclares, C.; Baraibar, A.M.; Araque, A.; Kofuji, P. Dysregulation of Astrocyte-Neuronal Communication in Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 7887. [Google Scholar] [CrossRef]
- Farhy Tselnicker, I.; Boisvert, M.M.; Allen, N.J. The role of neuronal versus astrocyte-derived heparan sulfate proteoglycans in brain development and injury. Biochem. Soc. Trans. 2014, 42, 1263–1269. [Google Scholar] [CrossRef]
- Arnst, N.; Kuznetsova, S.; Lipachev, N.; Shaikhutdinov, N.; Melnikova, A.; Mavlikeev, M.; Uvarov, P.; Baltina, T.V.; Rauvala, H.; Osin, Y.N.; et al. Spatial patterns and cell surface clusters in perineuronal nets. Brain Res. 2016, 1648, 214–223. [Google Scholar] [CrossRef]
- Testa, D.; Prochiantz, A.; Di Nardo, A.A. Perineuronal nets in brain physiology and disease. In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2019; Volume 89, pp. 125–135. [Google Scholar]
- Miyata, S.; Nishimura, Y.; Nakashima, T. Perineuronal nets protect against amyloid beta-protein neurotoxicity in cultured cortical neurons. Brain Res. 2007, 1150, 200–206. [Google Scholar] [CrossRef]
- Dow, K.; Riopelle, R.J.; Kisilevsky, R. Domains of neuronal heparan sulfate proteoglycans involved in neurite growth on laminin. Cell Tissue Res. 1991, 265, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Riopelle, R.; Dow, K.E. Functional interactions of neuronal heparan sulfate proteoglycans with laminin. Brain Res. 1990, 525, 92–100. [Google Scholar] [CrossRef]
- Andres, J.; DeFalcis, D.; Noda, M.; Massagué, J. Binding of two growth factor families to separate domains of the proteoglycan betaglycan. J. Biol. Chem. 1992, 267, 5927–5930. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Buck, M. Neuropilin-1 assists SARS-CoV-2 infection by stimulating the separation of Spike protein S1 and S2. Biophys. J. 2021, 120, 2828–2837. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, R.; Newton, G.; Carman, C.V.; Greenwood, J.; Luscinskas, F.W. Novel role of CD47 in rat microvascular endothelium: Signaling and regulation of T-cell transendothelial migration. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 2566–2576. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Yu, Z.; Liu, Y.; Bai, Y.; Jiang, Y.; van Leyen, K.; Yang, Y.G.; Lok, J.M.; Whalen, M.J.; Lo, E.H.; et al. CD47 deficiency improves neurological outcomes of traumatic brain injury in mice. Neurosci. Lett. 2017, 643, 125–130. [Google Scholar] [CrossRef]
- Lehrman, E.; Wilton, D.K.; Litvina, E.Y.; Welsh, C.A.; Chang, S.T.; Frouin, A.; Walker, A.J.; Heller, M.D.; Umemori, H.; Chen, C.; et al. CD47 Protects Synapses from Excess Microglia-Mediated Pruning during Development. Neuron 2018, 100, 120–134. [Google Scholar] [CrossRef] [PubMed]
- Gheibihayat, S.; Cabezas, R.; Nikiforov, N.G.; Jamialahmadi, T.; Johnston, T.P.; Sahebkar, A. CD47 in the Brain and Neurodegeneration: An Update on the Role in Neuroinflammatory Pathways. Molecules 2021, 26, 3943. [Google Scholar] [CrossRef]
- Griffiths, M.; Neal, J.W.; Gasque, P. Innate immunity and protective neuroinflammation: New emphasis on the role of neuroimmune regulatory proteins. Int. Rev. Neurobiol. 2007, 82, 29–55. [Google Scholar]
- Kvist, A.; Johnson, A.E.; Mörgelin, M.; Gustafsson, E.; Bengtsson, E.; Lindblom, K.; Aszódi, A.; Fässler, R.; Sasaki, T.; Timpl, R.; et al. Chondroitin sulfate perlecan enhances collagen fibril formation. Implications for perlecan chondrodysplasias. J. Biol. Chem. 2006, 281, 33127–33139. [Google Scholar] [CrossRef]
- Smith, S.; West, L.A.; Govindraj, P.; Zhang, X.; Ornitz, D.M.; Hassell, J.R. Heparan and chondroitin sulfate on growth plate perlecan mediate binding and delivery of FGF-2 to FGF receptors. Matrix Biol. 2007, 26, 175–184. [Google Scholar] [CrossRef]
- Yamada, S.; Mizumoto, S. Characterization of Hyaluronidase 4 Involved in the Catabolism of Chondroitin Sulfate. Molecules 2022, 27, 6103. [Google Scholar] [CrossRef]
- Backen, A.; Cole, C.L.; Lau, S.C.; Clamp, A.R.; McVey, R.; Gallagher, J.T.; Jayson, G.C. Heparan sulfate synthetic and editing enzymes in ovarian cancer. Br. J. Cancer 2007, 96, 1544–1548. [Google Scholar] [CrossRef]
- Cole, C.; Rushton, G.; Jayson, G.C.; Avizienyte, E. Ovarian cancer cell heparan sulfate 6-O-sulfotransferases regulate an angiogenic program induced by heparin-binding epidermal growth factor (EGF)-like growth factor/EGF receptor signaling. J. Biol. Chem. 2014, 289, 10488–10501. [Google Scholar] [CrossRef]
- Ferreras, C.; Rushton, G.; Cole, C.L.; Babur, M.; Telfer, B.A.; van Kuppevelt, T.H.; Gardiner, J.M.; Williams, K.J.; Jayson, G.C.; Avizienyte, E. Endothelial heparan sulfate 6-O-sulfation levels regulate angiogenic responses of endothelial cells to fibroblast growth factor 2 and vascular endothelial growth factor. J. Biol. Chem. 2012, 287, 36132–36146. [Google Scholar] [CrossRef] [PubMed]
- Deepa, S.; Yamada, S.; Zako, M.; Goldberger, O.; Sugahara, K. Chondroitin sulfate chains on syndecan-1 and syndecan-4 from normal murine mammary gland epithelial cells are structurally and functionally distinct and cooperate with heparan sulfate chains to bind growth factors. A novel function to control binding of midkine, pleiotrophin, and basic fibroblast growth factor. J. Biol. Chem. 2004, 279, 37368–37376. [Google Scholar] [PubMed]
- Ervasti, J.; Campbell, K.P. A role for the dystrophin glycoprotein complex as a transmembrane linker between laminin and actin. J. Cell Biol. 1993, 122, 809–823. [Google Scholar] [CrossRef] [PubMed]
- Gee, S.; Montanaro, F.; Lindenbaum, M.H.; Carbonetto, S. Dystroglycan α, a dystrophin associated glycoprotein, is a functional agrin receptor. Cell 1994, 77, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Ali, A.; Daggett, D.; Rauvala, H.; Hassell, J.; Smalheiser, N. The relationship between perlecan and dystroglycan and its implication in the formation of the neuromuscular junction. Cell Adhes. Commun. 1998, 5, 475–489. [Google Scholar] [CrossRef]
- Liu, Y.; Yu, M.; Shang, X.; Nguyen, M.H.H.; Balakrishnan, S.; Sager, R.; Hu, H. Eyes shut homolog (EYS) interacts with matriglycan of O mannosyl glycans whose deficiency results in EYS mislocalization and degeneration of photoreceptors. Sci. Rep. 2020, 10, 7795. [Google Scholar] [CrossRef]
- Yamaguchi, Y. Lecticans: Organizers of the brain extracellular matrix. Cell. Mol. Life Sci. 2000, 57, 276–289. [Google Scholar] [CrossRef]
- Du, W.; Fang, L.; Yang, W.; Sheng, W.; Zhang, Y.; Seth, A.; Yang, B.B.; Yee, A.J. The role of versican G3 domain in regulating breast cancer cell motility including effects on osteoblast cell growth and differentiation in vitro-evaluation towards understanding breast cancer cell bone metastasis. BMC Cancer 2012, 12, 341. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Dong, H.; Wan, Y.; Li, J.; Yee, A.; Yang, B.B.; Lu, W.Y. Versican G3 domain regulates neurite growth and synaptic transmission of hippocampal neurons by activation of epidermal growth factor receptor. J. Biol. Chem. 2006, 281, 19358–19368. [Google Scholar] [CrossRef]
- Reissner, C.; Stahn, J.; Breuer, D.; Klose, M.; Pohlentz, G.; Mormann, M.; Missler, M. Dystroglycan binding to α-neurexin competes with neurexophilin-1 and neuroligin in the brain. J. Biol. Chem. 2014, 289, 27585–27603. [Google Scholar] [CrossRef] [PubMed]
- Born, G.; Breuer, D.; Wang, S.; Rohlmann, A.; Coulon, P.; Vakili, P.; Reissner, C.; Kiefer, F.; Heine, M.; Pape, H.C.; et al. Modulation of synaptic function through the α-neurexin-specific ligand neurexophilin-1. Proc. Natl. Acad. Sci. USA 2014, 111, E1274–E1283. [Google Scholar] [CrossRef] [PubMed]
- Craig, A.; Kang, Y. Neurexin-neuroligin signaling in synapse development. Curr. Opin. Neurobiol. 2007, 17, 43–52. [Google Scholar] [CrossRef]
- Liu, X.; Hua, F.; Yang, D.; Lin, Y.; Zhang, L.; Ying, J.; Sheng, H.; Wang, X. Roles of neuroligins in central nervous system development: Focus on glial neuroligins and neuron neuroligins. J. Transl. Med. 2022, 20, 418. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.; White, K.I.; Zhou, Q.; Pfuetzner, R.A.; Choi, U.B.; Südhof, T.C.; Brunger, A.T. Structures of neurexophilin-neurexin complexes reveal a regulatory mechanism of alternative splicing. EMBO J. 2019, 38, e101603. [Google Scholar] [CrossRef] [PubMed]
- Dean, C.; Dresbach, T. Neuroligins and neurexins: Linking cell adhesion, synapse formation and cognitive function. Trends Neurosci. 2006, 29, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Graf, E.; Zhang, X.; Jin, S.X.; Linhoff, M.W.; Craig, A.M. Neurexins induce differentiation of GABA and glutamate postsynaptic specializations via neuroligins. Cell 2004, 119, 1013–1026. [Google Scholar] [CrossRef]
- Yoshida-Moriguchi, T.; Campbell, K.P. Matriglycan: A novel polysaccharide that links dystroglycan to the basement membrane. Glycobiology 2015, 25, 702–713. [Google Scholar] [CrossRef]
- Kanagawa, M.; Saito, F.; Kunz, S.; Yoshida-Moriguchi, T.; Barresi, R.; Kobayashi, Y.M.; Muschler, J.; Dumanski, J.P.; Michele, D.E.; Oldstone, M.B.; et al. Molecular recognition by LARGE is essential for expression of functional dystroglycan. Cell 2004, 117, 953–964. [Google Scholar] [CrossRef]
- Manya, H.; Yamaguchi, Y.; Kanagawa, M.; Kobayashi, K.; Tajiri, M.; Akasaka-Manya, K.; Kawakami, H.; Mizuno, M.; Wada, Y.; Toda, T.; et al. The Muscular Dystrophy Gene TMEM5 Encodes a Ribitol β1,4-Xylosyltransferase Required for the Functional Glycosylation of Dystroglycan. J. Biol. Chem. 2016, 291, 24618–24627. [Google Scholar] [CrossRef]
- Riemersma, M.; Froese, D.S.; van Tol, W.; Engelke, U.F.; Kopec, J.; van Scherpenzeel, M.; Ashikov, A.; Krojer, T.; von Delft, F.; Tessari, M.; et al. Human ISPD is a Cytidyltransferase Required for Dystroglycan O-Mannosylation. Chem. Biol. 2015, 22, 1643–1652. [Google Scholar] [CrossRef]
- Praissman, J.; Willer, T.; Sheikh, M.O.; Toi, A.; Chitayat, D.; Lin, Y.Y.; Lee, H.; Stalnaker, S.H.; Wang, S.; Prabhakar, P.K.; et al. The functional O-mannose glycan on α-dystroglycan contains a phospho-ribitol primed for matriglycan addition. eLife 2016, 5, e14473. [Google Scholar] [CrossRef] [PubMed]
- Willer, T.; Lee, H.; Lommel, M.; Yoshida-Moriguchi, T.; de Bernabe, D.B.; Venzke, D.; Cirak, S.; Schachter, H.; Vajsar, J.; Voit, T.; et al. ISPD loss-of-function mutations disrupt dystroglycan O-mannosylation and cause Walker-Warburg syndrome. Nat. Genet. 2012, 44, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Walimbe, A.; Okuma, H.; Joseph, S.; Yang, T.; Yonekawa, T.; Hord, J.M.; Venzke, D.; Anderson, M.E.; Torelli, S.; Manzur, A.; et al. POMK regulates dystroglycan function via LARGE1-mediated elongation of matriglycan. Elife 2020, 9, e61388. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hayes, A.J.; Melrose, J. HS, an Ancient Molecular Recognition and Information Storage Glycosaminoglycan, Equips HS-Proteoglycans with Diverse Matrix and Cell-Interactive Properties Operative in Tissue Development and Tissue Function in Health and Disease. Int. J. Mol. Sci. 2023, 24, 1148. https://doi.org/10.3390/ijms24021148
Hayes AJ, Melrose J. HS, an Ancient Molecular Recognition and Information Storage Glycosaminoglycan, Equips HS-Proteoglycans with Diverse Matrix and Cell-Interactive Properties Operative in Tissue Development and Tissue Function in Health and Disease. International Journal of Molecular Sciences. 2023; 24(2):1148. https://doi.org/10.3390/ijms24021148
Chicago/Turabian StyleHayes, Anthony J., and James Melrose. 2023. "HS, an Ancient Molecular Recognition and Information Storage Glycosaminoglycan, Equips HS-Proteoglycans with Diverse Matrix and Cell-Interactive Properties Operative in Tissue Development and Tissue Function in Health and Disease" International Journal of Molecular Sciences 24, no. 2: 1148. https://doi.org/10.3390/ijms24021148
APA StyleHayes, A. J., & Melrose, J. (2023). HS, an Ancient Molecular Recognition and Information Storage Glycosaminoglycan, Equips HS-Proteoglycans with Diverse Matrix and Cell-Interactive Properties Operative in Tissue Development and Tissue Function in Health and Disease. International Journal of Molecular Sciences, 24(2), 1148. https://doi.org/10.3390/ijms24021148