
Dynamic Reconfiguration of Switchgrass Proteomes in Response to Rust (Puccinia novopanici) Infection
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
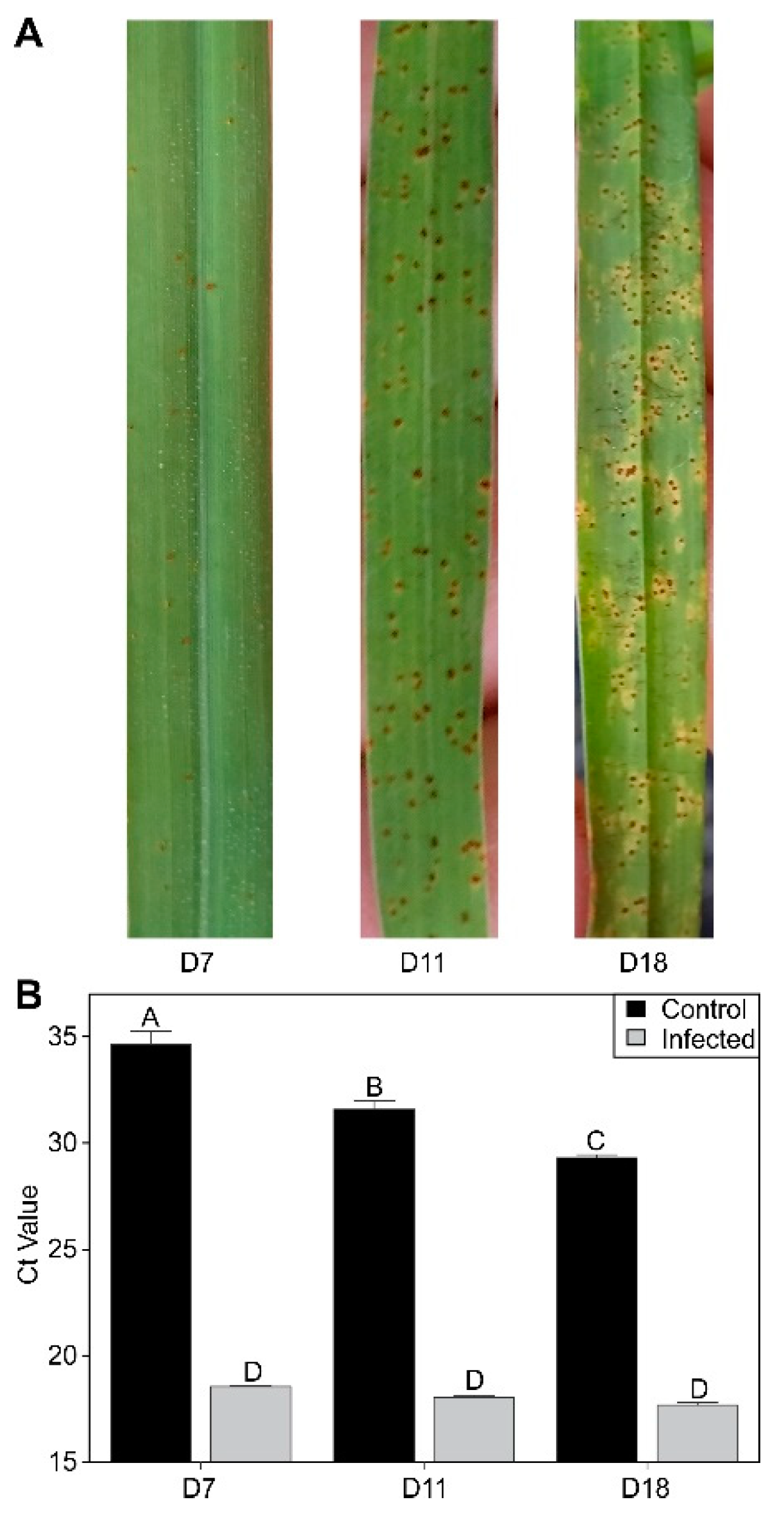
2.1. Disease Progression and Change in Fungal Loads
2.2. Total Protein Identified and Differentially Abundant
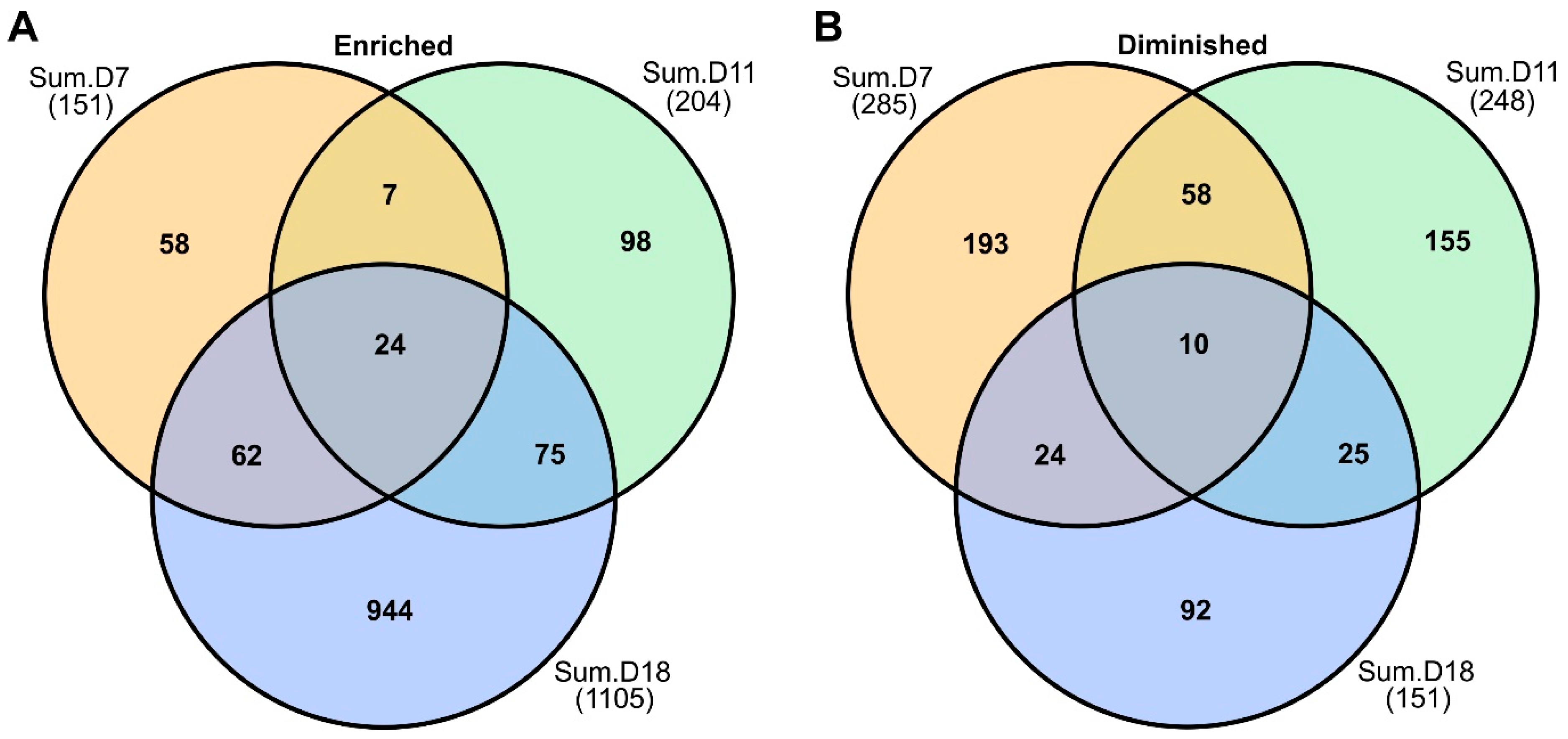
2.3. Rust Infection Substantially Altered Host Proteomes
2.4. STRING Analysis Using Arabidopsis Homologs and Annotations
2.5. Ribosomal Proteins and mRNA Splicing Were among EDAPs at 7 DAI
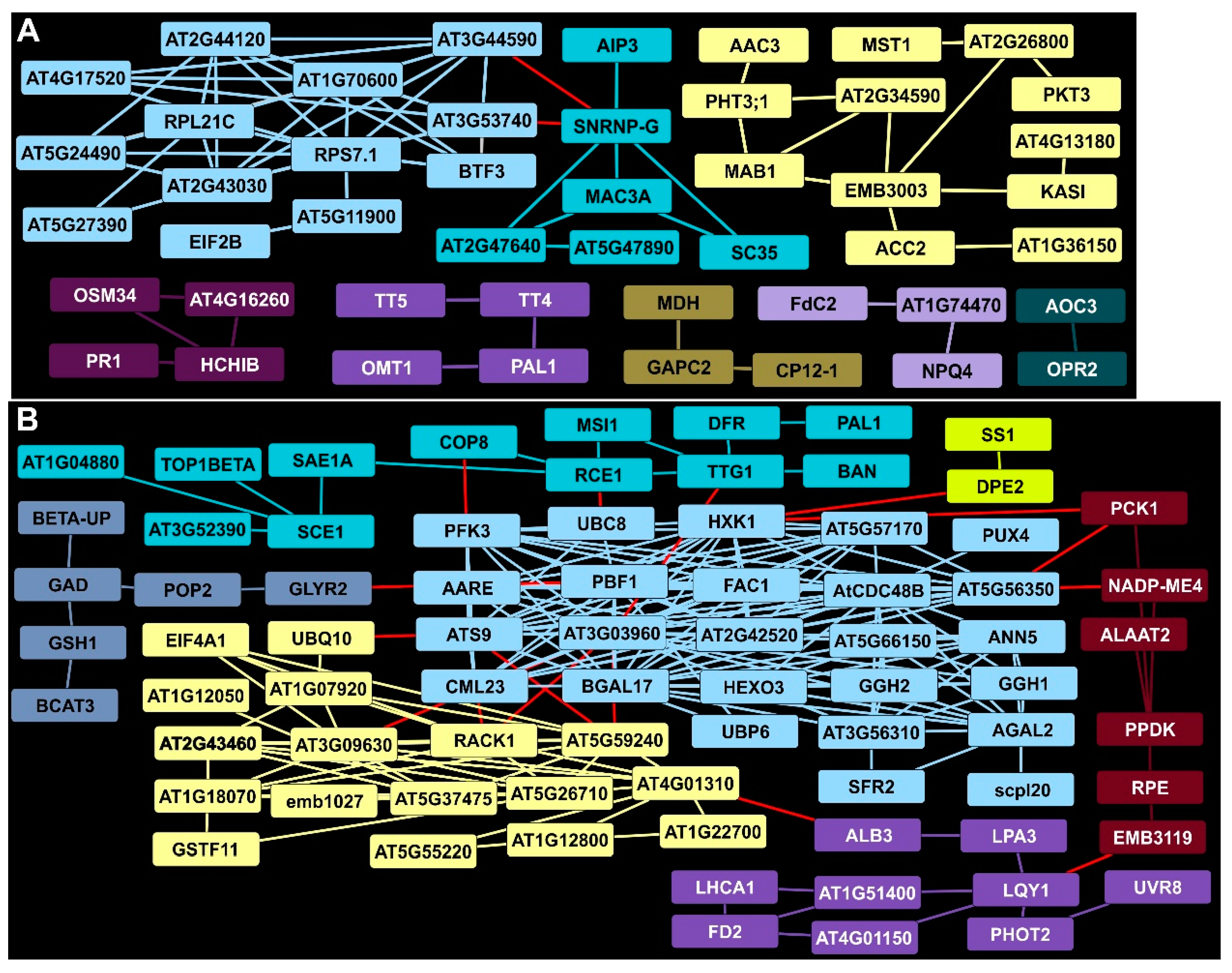
2.6. Identification of a Central Hub for DDAPs at 7 DAI in Rust-Infected Summer Plants
2.7. Plant Defense and Lignin Biosynthetic Clusters Were Enriched by 11 DAI
2.8. Protein Networks Indicated Continued Suppression of Primary Metabolism at 11 DAI
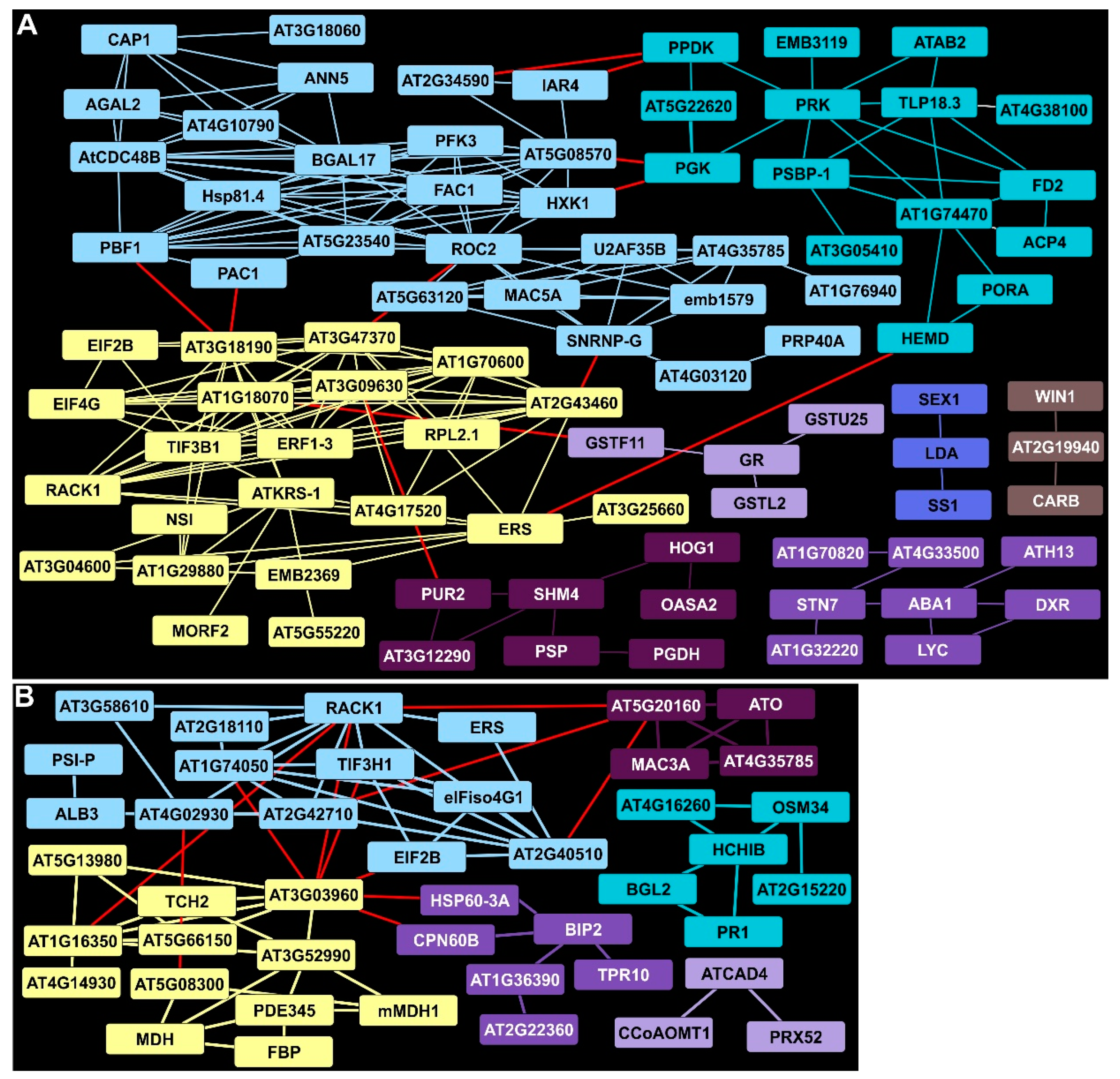
2.9. Extensive Remodeling of the Proteome Occurred at 18 DAI
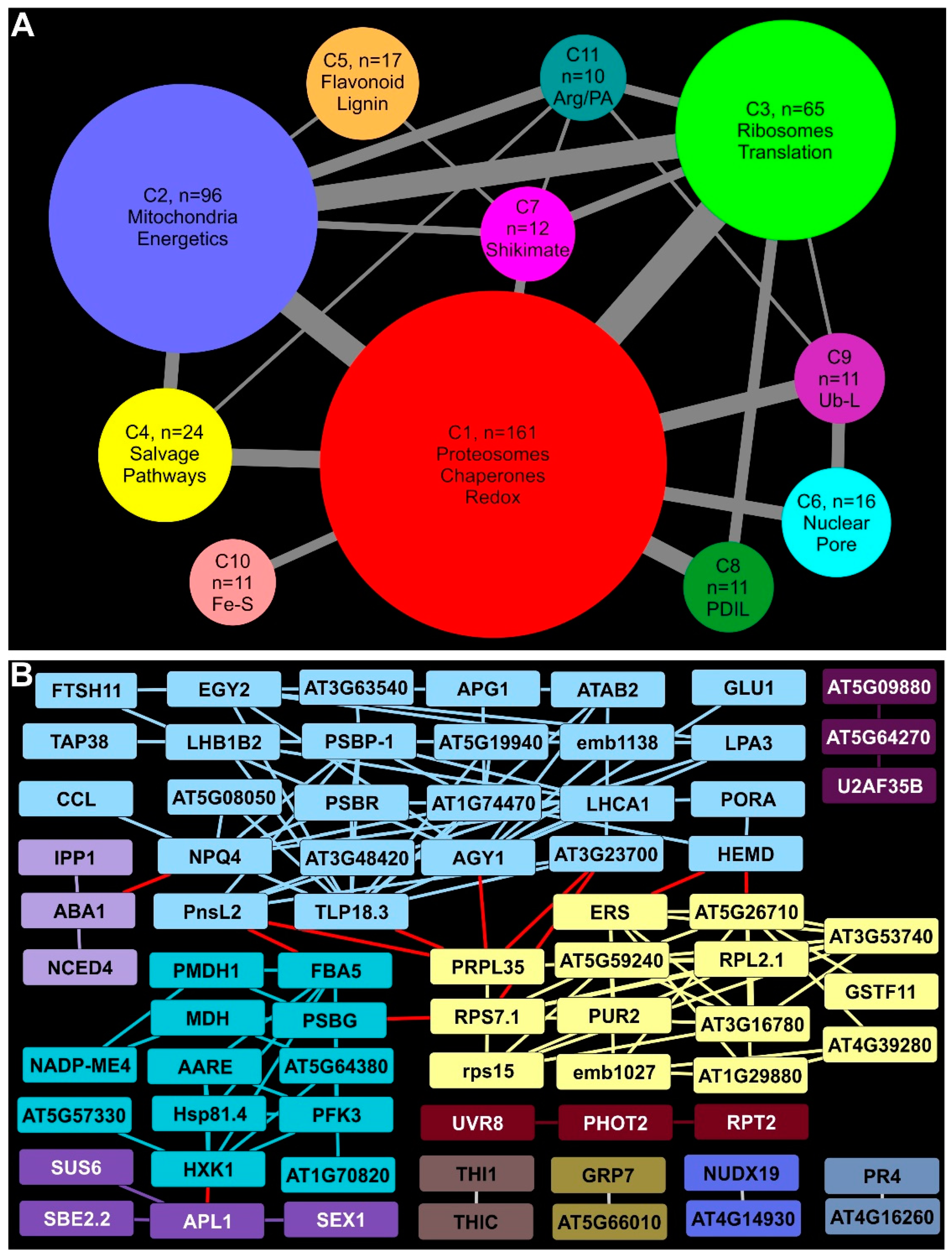
2.10. Proteosomal, Chaperone, and Redox Proteins Accumulated under Prolonged Rust Infection
2.11. Mitochondria and Energy Metabolism Were Distinctly Affected in Infected Switchgrass Plants
2.12. Ribosomal and Translation-Related Proteins Enrichment Indicated Substantial Protein Synthesis in Infected Plants
2.13. Proteins Needed for Salvage Pathways Were Enriched
2.14. Phenylpropanoid, Flavonoid, and Anthocyanin Biosynthetic Enzymes Were Enriched at 18 DAI
2.15. Other Notable Clusters
2.16. Plastid Biosynthetic Pathways Were Negatively Impacted by Rust Infection at 18 DAI
2.17. Changes in the Fungal Proteome
3. Discussion
4. Methods
4.1. Plant Materials
4.2. Rust Inoculation
4.3. Sample Collection and Processing for Proteomics
4.4. Experimental Design and Statistical Analyses
4.5. Functional Annotation and STRING Analysis
4.6. Estimation of Pn DNA Levels in Leaves
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Jashni, M.K.; Mehrabi, R.; Collemare, J.; Mesarich, C.H.; de Wit, P.J. The battle in the apoplast: Further insights into the roles of proteases and their inhibitors in plant-pathogen interactions. Front. Plant Sci. 2015, 6, 584. [Google Scholar] [CrossRef] [PubMed]
- Dean, R.A. Signal pathways and appressorium morphogenesis. Annu. Rev. Phytopathol. 1997, 35, 211–234. [Google Scholar] [CrossRef]
- Kachroo, A.; Robin, G.P. Systemic signaling during plant defense. Curr. Opin. Plant Biol. 2013, 16, 527–533. [Google Scholar] [CrossRef]
- Muthamilarasan, M.; Prasad, M. Plant innate immunity: An updated insight into defense mechanism. J. Biosci. 2013, 38, 433–449. [Google Scholar] [CrossRef] [PubMed]
- Agrios, G. Plant Pathology, 5th ed.; Elsevier: Amsterdam, The Netherlands, 2004; Volume 9780080473789, pp. 1–922. [Google Scholar]
- Stewart, C.L.; Pyle, J.D.; Jochum, C.C.; Vogel, K.P.; Yuen, G.Y.; Scholthof, K.B.G. Multi-Year Pathogen Survey of Biofuel Switchgrass Breeding Plots Reveals High Prevalence of Infections by Panicum mosaic virus and Its Satellite Virus. Phytopathology 2015, 105, 1146–1154. [Google Scholar] [CrossRef]
- Stewart, C.L.; Jochum, C.; Yuen, G.Y.; Vogel, K.; Pyle, J.D.; Scholthof, K.B.G. Incidence and impact of dual infection by Panicum mosaic virus and its satellite virus in switchgrass breeding fields. Phytopathology 2014, 104, 114. [Google Scholar]
- Gustafson, D.M.; Boe, A.; Jin, Y. Genetic variation for Puccinia emaculata infection in switchgrass. Crop Sci. 2003, 43, 755–759. [Google Scholar] [CrossRef]
- Agindotan, B.; Okanu, N.; Oladeinde, A.; Voigt, T.; Long, S.; Gray, M.; Bradley, C. Detection of Switchgrass mosaic virus in Miscanthus and other grasses. Can. J. Plant Pathol. 2013, 35, 81–86. [Google Scholar] [CrossRef]
- Crouch, J.A.; Beirn, L.A.; Cortese, L.M.; Bonos, S.A.; Clarke, B.B. Anthracnose disease of switchgrass caused by the novel fungal species Colletotrichum navitas. Mycol. Res. 2009, 113, 1411–1421. [Google Scholar] [CrossRef]
- Demers, J.E.; Liu, M.; Hambleton, S.; Castlebury, L.A. Rust fungi on panicum. Mycologia 2017, 109, 1–17. [Google Scholar] [CrossRef]
- Sykes, V.R.; Allen, F.L.; Mielenz, J.R.; Stewart, C.N., Jr.; Windham, M.T.; Hamilton, C.Y.; Rodriguez, M., Jr.; Yee, K.L. Reduction of Ethanol Yield from Switchgrass Infected with Rust Caused by Puccinia emaculata. Bioenerg. Res. 2016, 9, 239–247. [Google Scholar] [CrossRef]
- Gill, U.S.; Sun, L.; Rustgi, S.; Tang, Y.; von Wettstein, D.; Mysore, K.S. Transcriptome-based analyses of phosphite-mediated suppression of rust pathogens Puccinia emaculata and Phakopsora pachyrhizi and functional characterization of selected fungal target genes. Plant J. 2018, 93, 894–904. [Google Scholar] [CrossRef] [PubMed]
- Bowen, K.L.; Hagan, A.K.; Miller, H.B. Rust (Puccinia emaculata) Management and Impact on Biomass Yield in Switchgrass. Plant Dis. 2022, 106, 390–394. [Google Scholar] [CrossRef] [PubMed]
- Vogel, K.P.; Mitchell, R.B.; Sarath, G.; Casler, M.D. Registration of ‘Liberty’ switchgrass. J. Plant Regist. 2014, 8, 242–247. [Google Scholar] [CrossRef]
- Uppalapati, S.R.; Serba, D.D.; Ishiga, Y.; Szabo, L.J.; Mittal, S.; Bhandari, H.S.; Bouton, J.H.; Mysore, K.S.; Saha, M.C. Characterization of the Rust Fungus, Puccinia emaculata, and Evaluation of Genetic Variability for Rust Resistance in Switchgrass Populations. Bioenergy Res. 2013, 6, 458–468. [Google Scholar] [CrossRef][Green Version]
- Cheng, Q.; Windham, A.S.; Lamour, K.H.; Saxton, A.M.; Windham, M.T. Evaluation of variation in switchgrass (Panicum virgatum L.) cultivars for rust (Puccinia emaculata) resistance. J. Environ. Hortic. 2019, 37, 127–135. [Google Scholar] [CrossRef]
- VanWallendael, A.; Bonnette, J.; Juenger, T.E.; Fritschi, F.B.; Fay, P.A.; Mitchell, R.B.; Lloyd-Reilley, J.; Rouquette, F.M., Jr.; Bergstrom, G.C.; Lowry, D.B. Geographic variation in the genetic basis of resistance to leaf rust between locally adapted ecotypes of the biofuel crop switchgrass (Panicum virgatum). New Phytol. 2020, 227, 1696–1708. [Google Scholar] [CrossRef]
- Paudel, B.; Das, A.; Tran, M.; Boe, A.; Palmer, N.A.; Sarath, G.; Gonzalez-Hernandez, J.L.; Rushton, P.J.; Rohila, J.S. Proteomic Responses of Switchgrass and Prairie Cordgrass to Senescence. Front. Plant Sci. 2016, 7, 293. [Google Scholar] [CrossRef]
- Lao, J.; Sharma, M.K.; Sharma, R.; Fernandez-Nino, S.M.; Schmutz, J.; Ronald, P.C.; Heazlewood, J.L.; Schwessinger, B. Proteome profile of the endomembrane of developing coleoptiles from switchgrass (Panicum virgatum). Proteomics 2015, 15, 2286–2290. [Google Scholar] [CrossRef][Green Version]
- Ye, Z.; Sangireddy, S.; Okekeogbu, I.; Zhou, S.; Yu, C.L.; Hui, D.; Howe, K.J.; Fish, T.; Thannhauser, T.W. Drought-Induced Leaf Proteome Changes in Switchgrass Seedlings. Int. J. Mol. Sci. 2016, 17, 1251. [Google Scholar] [CrossRef]
- Ye, Z.J.; Sangireddy, S.R.; Yu, C.L.; Hui, D.F.; Howe, K.; Fish, T.; Thannhauser, T.W.; Zhou, S.P. Comparative Proteomics of Root Apex and Root Elongation Zones Provides Insights into Molecular Mechanisms for Drought Stress and Recovery Adjustment in Switchgrass. Proteomes 2020, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Zogli, P.; Alvarez, S.; Naldrett, M.J.; Palmer, N.A.; Koch, K.G.; Pingault, L.; Bradshaw, J.D.; Twigg, P.; Heng-Moss, T.M.; Louis, J.; et al. Greenbug (Schizaphis graminum) herbivory significantly impacts protein and phosphorylation abundance in switchgrass (Panicum virgatum). Sci. Rep. 2020, 10, 14842. [Google Scholar] [CrossRef] [PubMed]
- Muhle, A.A.; Palmer, N.A.; Edme, S.J.; Sarath, G.; Yuen, G.; Mitchell, R.B.; Tatineni, S. Effect of cultivar and temperature on the synergistic interaction between panicum mosaic virus and satellite panicum mosaic virus in switchgrass. Arch. Virol. 2022, 167, 1247–1256. [Google Scholar] [CrossRef] [PubMed]
- Pingault, L.; Palmer, N.A.; Koch, K.G.; Heng-Moss, T.; Bradshaw, J.D.; Seravalli, J.; Twigg, P.; Louis, J.; Sarath, G. Differential Defense Responses of Upland and Lowland Switchgrass Cultivars to a Cereal Aphid Pest. Int. J. Mol. Sci. 2020, 21, 7966. [Google Scholar] [CrossRef] [PubMed]
- Koch, K.G.; Palmer, N.A.; Donze-Reiner, T.; Scully, E.D.; Seravalli, J.; Amundsen, K.; Twigg, P.; Louis, J.; Bradshaw, J.D.; Heng-Moss, T.M.; et al. Aphid-Responsive Defense Networks in Hybrid Switchgrass. Front. Plant Sci. 2020, 11, 1145. [Google Scholar] [CrossRef] [PubMed]
- Palmer, N.A.; Chowda-Reddy, R.V.; Muhle, A.A.; Tatineni, S.; Yuen, G.; Edme, S.J.; Mitchell, R.B.; Sarath, G. Transcriptome divergence during leaf development in two contrasting switchgrass (Panicum virgatum L.) cultivars. PLoS ONE 2019, 14, e0222080. [Google Scholar] [CrossRef]
- Serba, D.D.; Uppalapati, S.R.; Mukherjee, S.; Krom, N.; Tang, Y.H.; Mysore, K.S.; Saha, M.C. Transcriptome Profiling of Rust Resistance in Switchgrass Using RNA-Seq Analysis. Plant Genome-Us 2015, 8. [Google Scholar] [CrossRef]
- Serba, D.D.; Uppalapati, S.R.; Krom, N.; Mukherjee, S.; Tang, Y.H.; Mysore, K.S.; Saha, M.C. Transcriptome analysis in switchgrass discloses ecotype difference in photosynthetic efficiency. BMC Genom. 2016, 17, 1040. [Google Scholar] [CrossRef]
- Frazier, T.P.; Palmer, N.A.; Xie, F.; Tobias, C.M.; Donze-Reiner, T.J.; Bombarely, A.; Childs, K.L.; Shu, S.; Jenkins, J.W.; Schmutz, J.; et al. Identification, characterization, and gene expression analysis of nucleotide binding site (NB)-type resistance gene homologues in switchgrass. BMC Genom. 2016, 17, 892. [Google Scholar] [CrossRef]
- Gill, U.S.; Nandety, R.S.; Krom, N.; Dai, X.; Zhuang, Z.; Tang, Y.; Zhao, P.X.; Mysore, K.S. Draft genome sequence resource of switchgrass rust pathogen, puccinia novopanici isolate ard-01. Phytopathology 2019, 109, 1513–1515. [Google Scholar] [CrossRef]
- Nandety, R.S.; Gill, U.S.; Krom, N.; Dai, X.; Dong, Y.; Zhao, P.X.; Mysore, K.S. Comparative Genome Analyses of Plant Rust Pathogen Genomes Reveal a Confluence of Pathogenicity Factors to Quell Host Plant Defense Responses. Plants 2022, 11, 1962. [Google Scholar] [CrossRef] [PubMed]
- Kuzniak, E.; Kopczewski, T. The Chloroplast Reactive Oxygen Species-Redox System in Plant Immunity and Disease. Front. Plant Sci. 2020, 11, 572686. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhai, T.; Zhang, X.; Tang, C.; Zhuang, R.; Zhao, H.; Xu, Q.; Cheng, Y.; Wang, J.; Duplessis, S.; et al. Two stripe rust effectors impair wheat resistance by suppressing import of host Fe-S protein into chloroplasts. Plant Physiol. 2021, 187, 2530–2543. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Tang, C.; Wang, X.; Sun, S.; Zhao, J.; Kang, Z.; Wang, X. An effector protein of the wheat stripe rust fungus targets chloroplasts and suppresses chloroplast function. Nat. Commun. 2019, 10, 5571. [Google Scholar] [CrossRef]
- Petre, B.; Lorrain, C.; Saunders, D.G.; Win, J.; Sklenar, J.; Duplessis, S.; Kamoun, S. Rust fungal effectors mimic host transit peptides to translocate into chloroplasts. Cell. Microbiol. 2016, 18, 453–465. [Google Scholar] [CrossRef]
- Dominguez, F.; Cejudo, F.J. Chloroplast dismantling in leaf senescence. J. Exp. Bot. 2021, 72, 5905–5918. [Google Scholar] [CrossRef]
- Balakireva, A.V.; Deviatkin, A.A.; Zgoda, V.G.; Kartashov, M.I.; Zhemchuzhina, N.S.; Dzhavakhiya, V.G.; Golovin, A.V.; Zamyatnin, A.A. Proteomics Analysis Reveals That Caspase-Like and Metacaspase-Like Activities Are Dispensable for Activation of Proteases Involved in Early Response to Biotic Stress in Triticum aestivum L. Int. J. Mol. Sci. 2018, 19, 3991. [Google Scholar] [CrossRef]
- Della Coletta, R.; Lavell, A.A.; Garvin, D.F. A Homolog of the Arabidopsis TIME FOR COFFEE Gene Is Involved in Nonhost Resistance to Wheat Stem Rust in Brachypodium distachyon. Mol. Plant Microbe Interact. 2021, 34, 1298–1306. [Google Scholar] [CrossRef]
- Sekiya, A.; Marques, F.G.; Leite, T.F.; Cataldi, T.R.; de Moraes, F.E.; Pinheiro, A.L.M.; Labate, M.T.V.; Labate, C.A. Network Analysis Combining Proteomics and Metabolomics Reveals New Insights Into Early Responses of Eucalyptus grandis During Rust Infection. Front. Plant Sci. 2021, 11, 604849. [Google Scholar] [CrossRef]
- Rampitsch, C.; Huang, M.; Djuric-Cignaovic, S.; Wang, X.B.; Fernando, U. Temporal Quantitative Changes in the Resistant and Susceptible Wheat Leaf Apoplastic Proteome During Infection by Wheat Leaf Rust (Puccinia triticina). Front. Plant Sci. 2019, 10, 1291. [Google Scholar] [CrossRef]
- Yang, Y.; Yu, Y.; Bi, C.; Kang, Z. Quantitative Proteomics Reveals the Defense Response of Wheat against Puccinia striiformis f. sp. tritici. Sci. Rep. 2016, 6, 34261. [Google Scholar] [CrossRef] [PubMed]
- Possa, K.F.; Silva, J.A.G.; Resende, M.L.V.; Tenente, R.; Pinheiro, C.; Chaves, I.; Planchon, S.; Monteiro, A.C.A.; Renaut, J.; Carvalho, M.A.F.; et al. Primary Metabolism Is Distinctly Modulated by Plant Resistance Inducers in Coffea arabica Leaves Infected by Hemileia vastatrix. Front. Plant Sci. 2020, 11, 309. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.X.; Chen, Z.; Tian, L.; Ding, Y.Z.; Zhang, J.; Zhou, J.L.; Liu, P.; Chen, Y.H.; Wu, L.J. Comparative proteomics combined with analyses of transgenic plants reveal ZmREM1.3 mediates maize resistance to southern corn rust. Plant Biotechnol. J. 2019, 17, 2153–2168. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palmer, N.A.; Alvarez, S.; Naldrett, M.J.; Muhle, A.; Sarath, G.; Edmé, S.J.; Tatineni, S.; Mitchell, R.B.; Yuen, G. Dynamic Reconfiguration of Switchgrass Proteomes in Response to Rust (Puccinia novopanici) Infection. Int. J. Mol. Sci. 2023, 24, 14630. https://doi.org/10.3390/ijms241914630
Palmer NA, Alvarez S, Naldrett MJ, Muhle A, Sarath G, Edmé SJ, Tatineni S, Mitchell RB, Yuen G. Dynamic Reconfiguration of Switchgrass Proteomes in Response to Rust (Puccinia novopanici) Infection. International Journal of Molecular Sciences. 2023; 24(19):14630. https://doi.org/10.3390/ijms241914630
Chicago/Turabian StylePalmer, Nathan A., Sophie Alvarez, Michael J. Naldrett, Anthony Muhle, Gautam Sarath, Serge J. Edmé, Satyanarayana Tatineni, Robert B. Mitchell, and Gary Yuen. 2023. "Dynamic Reconfiguration of Switchgrass Proteomes in Response to Rust (Puccinia novopanici) Infection" International Journal of Molecular Sciences 24, no. 19: 14630. https://doi.org/10.3390/ijms241914630
APA StylePalmer, N. A., Alvarez, S., Naldrett, M. J., Muhle, A., Sarath, G., Edmé, S. J., Tatineni, S., Mitchell, R. B., & Yuen, G. (2023). Dynamic Reconfiguration of Switchgrass Proteomes in Response to Rust (Puccinia novopanici) Infection. International Journal of Molecular Sciences, 24(19), 14630. https://doi.org/10.3390/ijms241914630