
Intravenous Administration of Human Muse Cells Ameliorates Deficits in a Rat Model of Subacute Spinal Cord Injury
 , and
, and
Abstract
:1. Introduction
2. Results
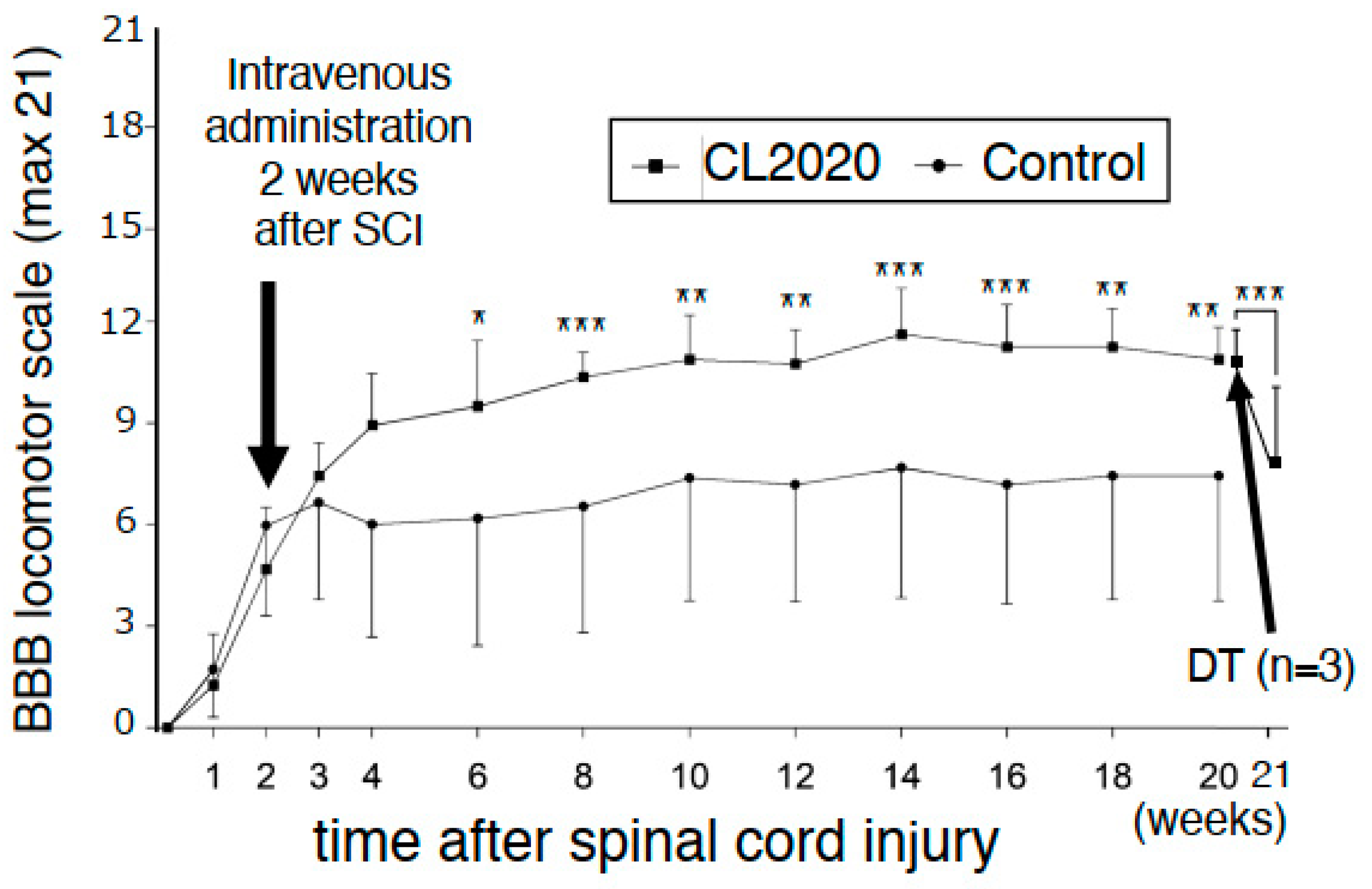
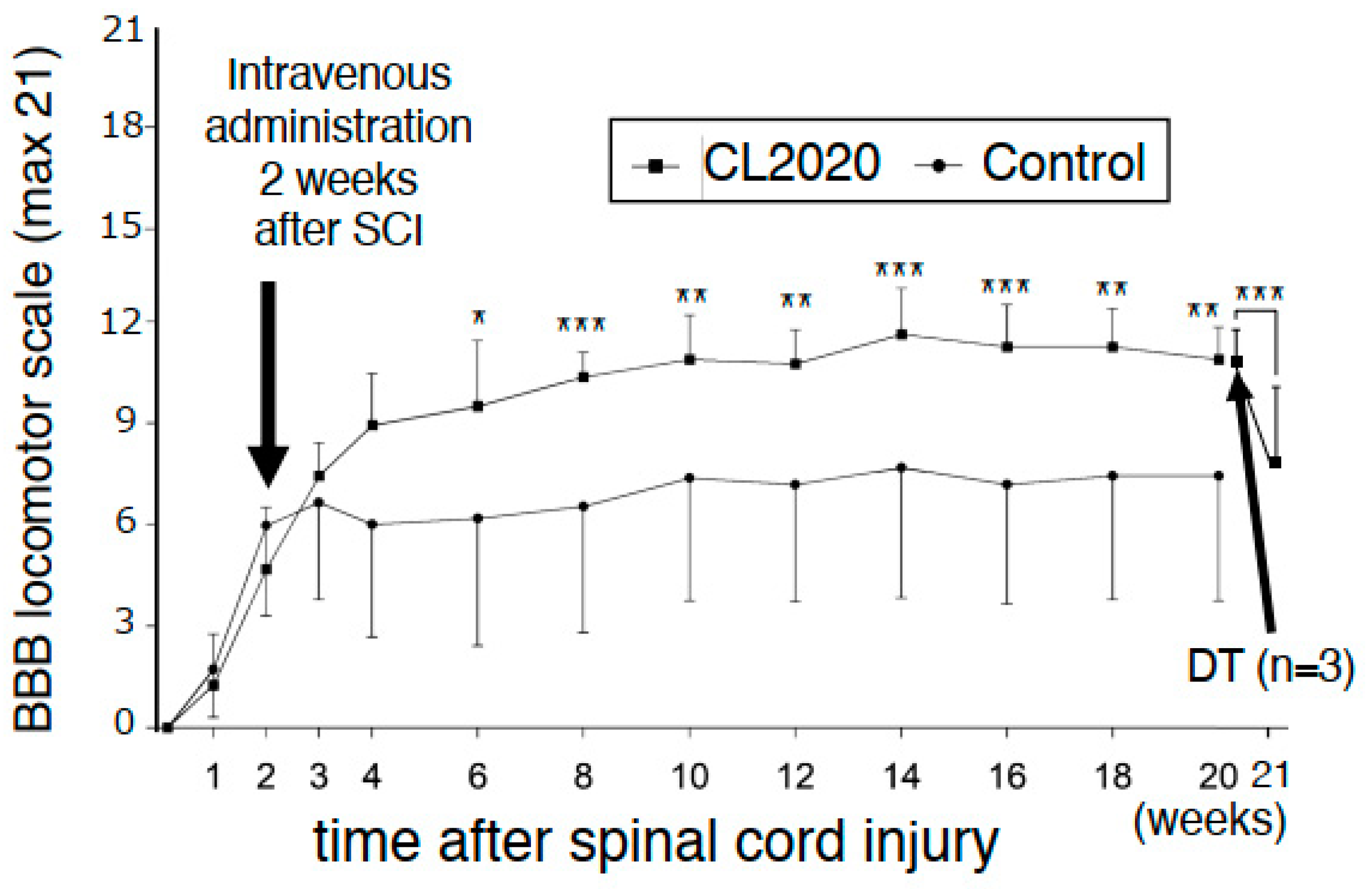
2.1. Intravenous CL2020 Improves Hindlimb Locomotor Functions after Subacute SCI
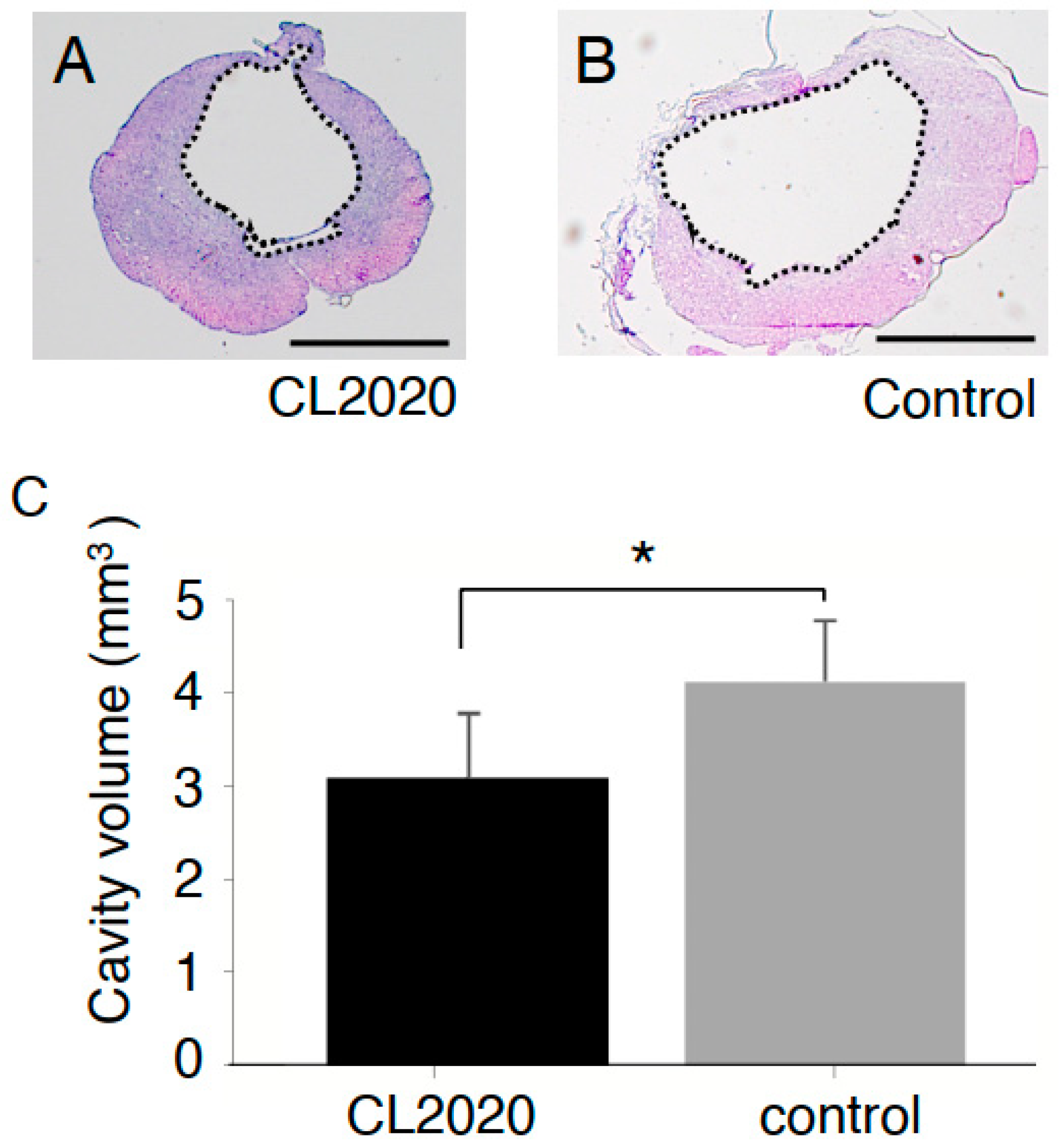
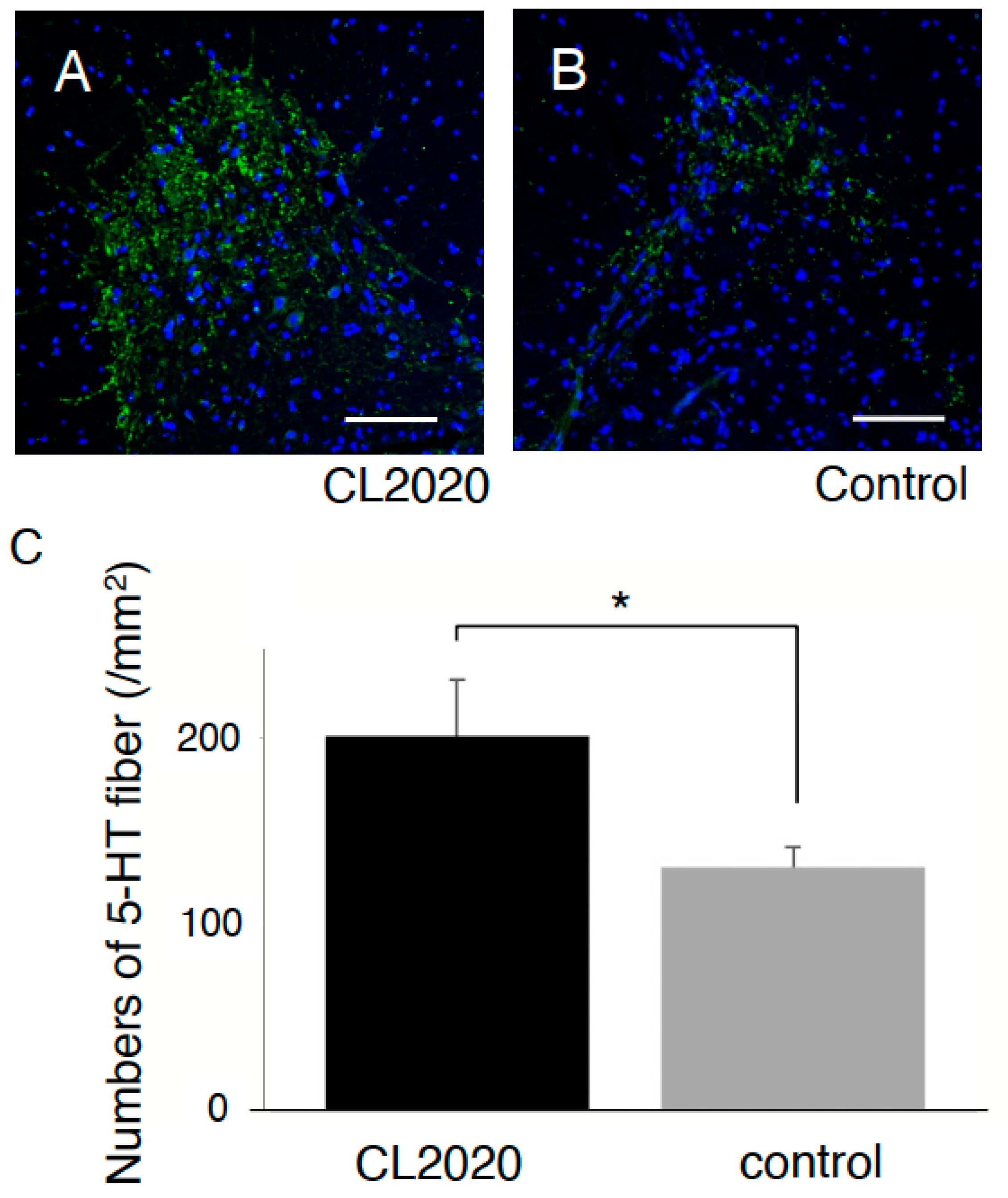
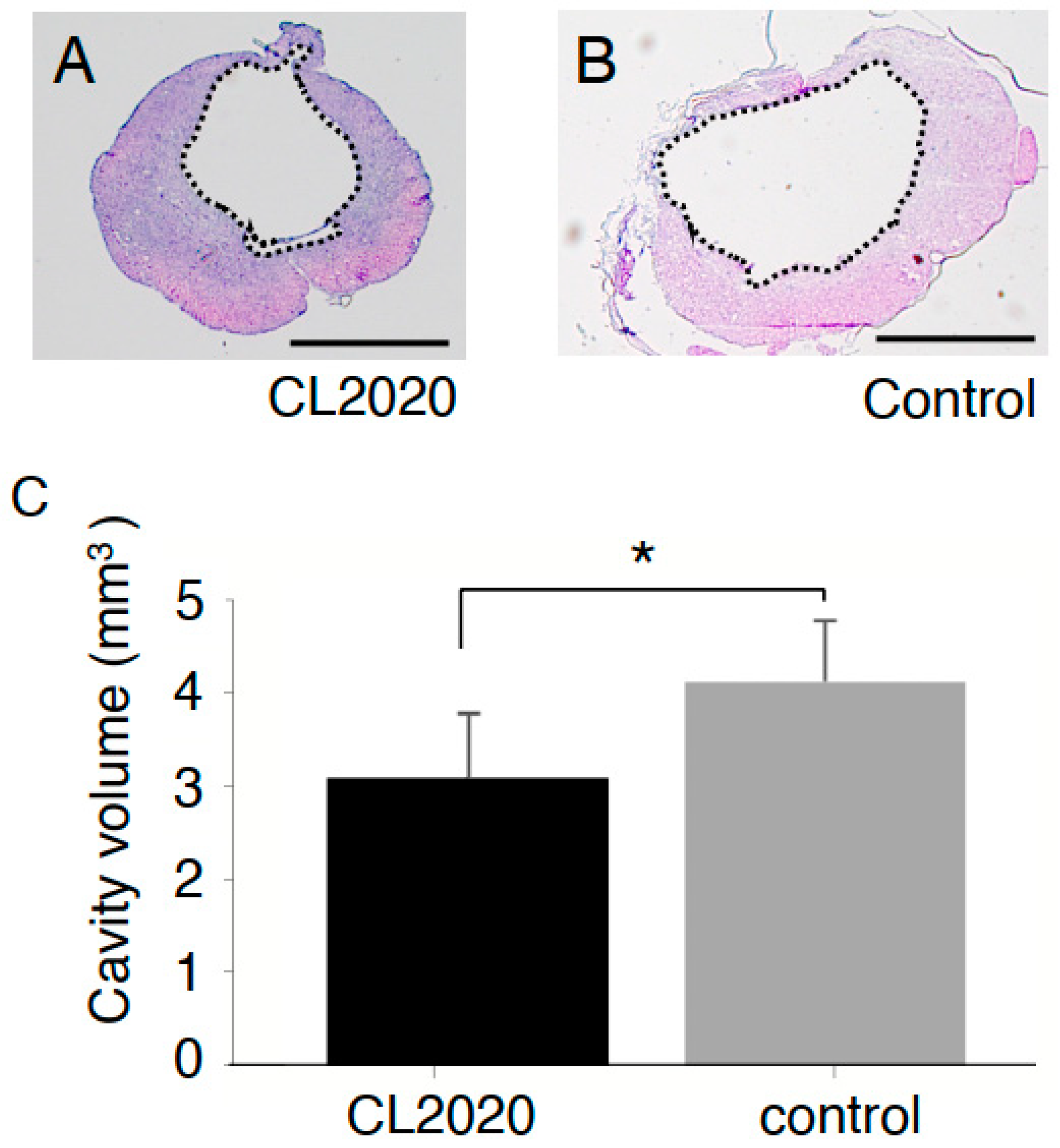
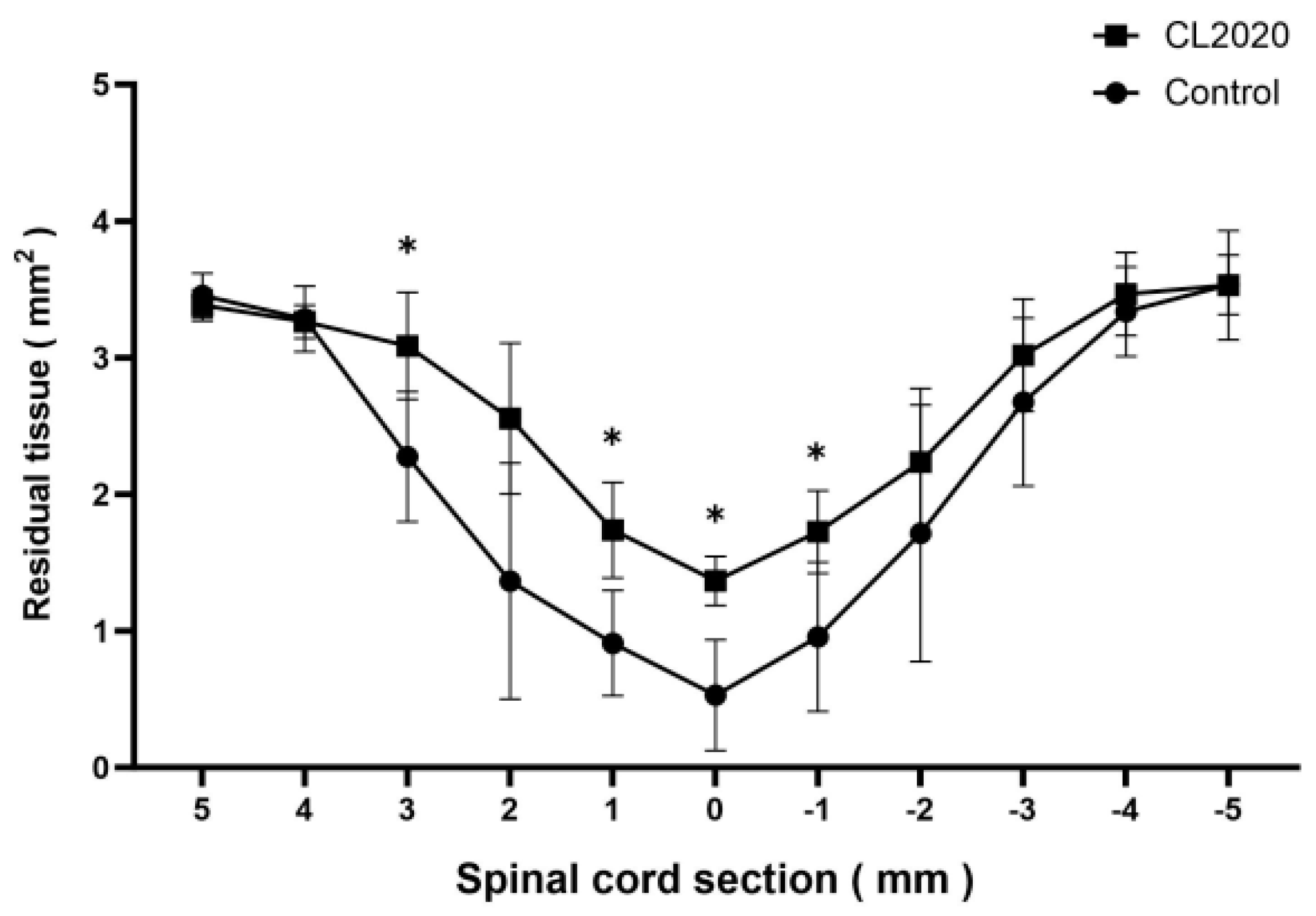
2.2. CL2020 Prevented Spinal Cord Damage and Contributed to Structural Preservation
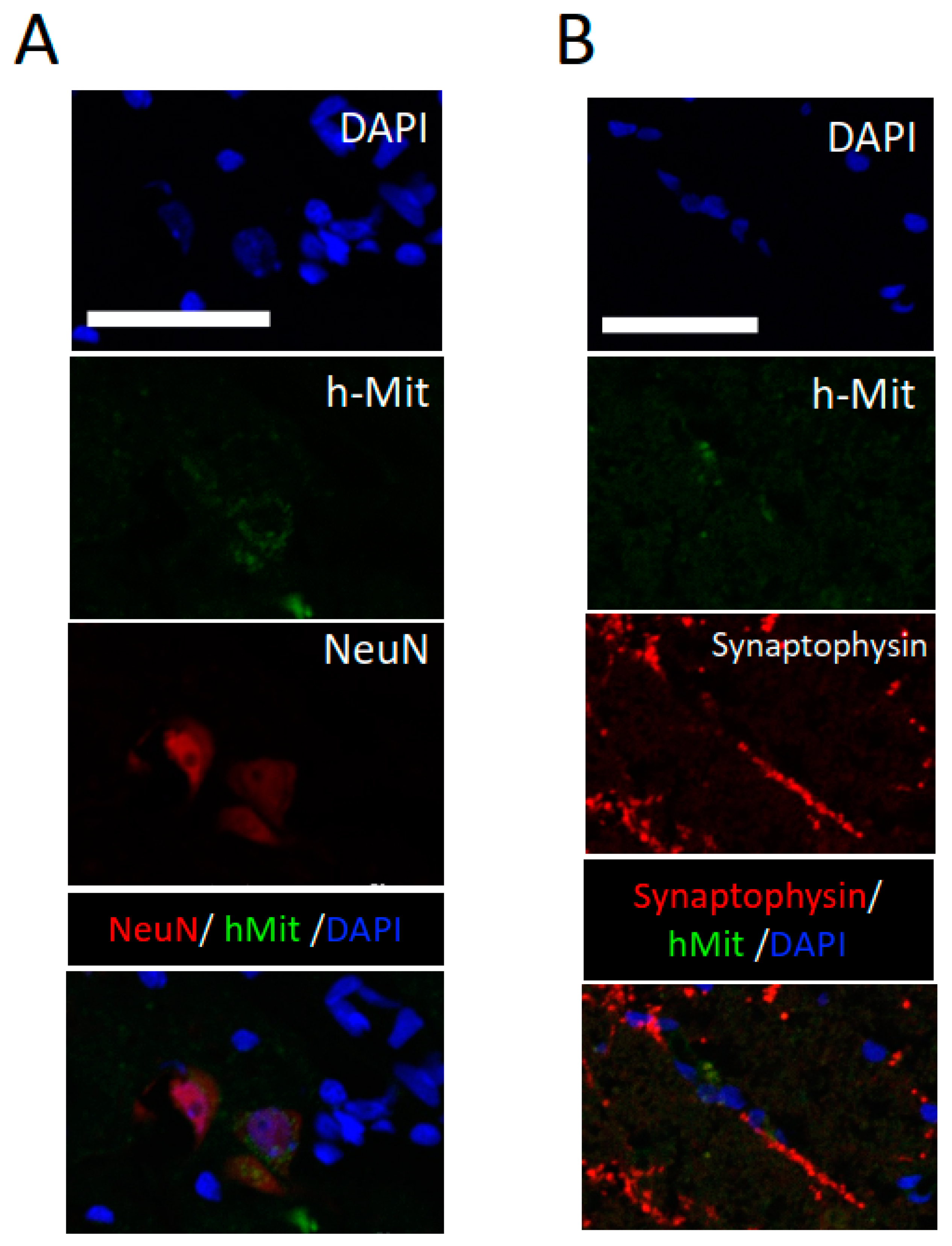
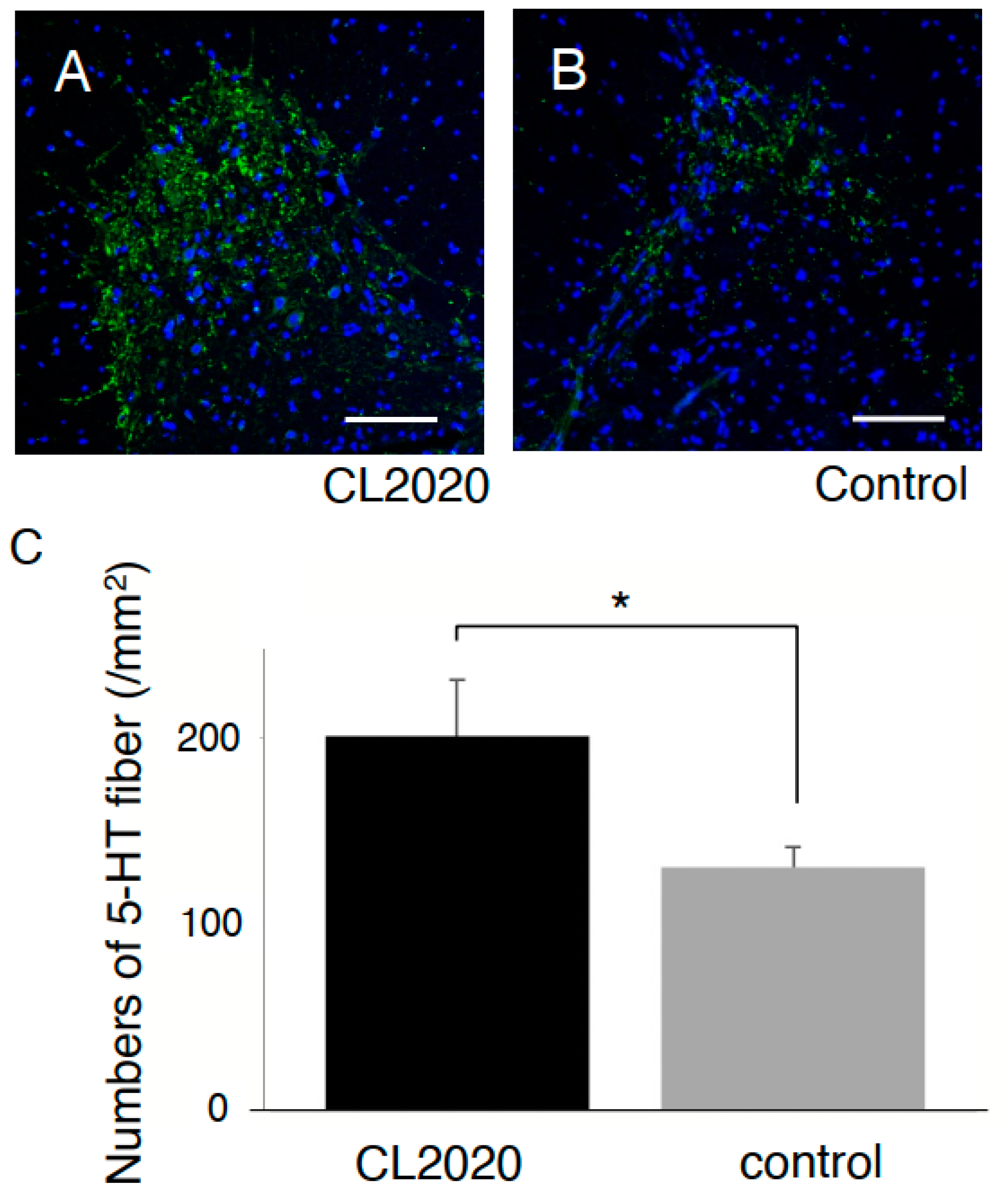
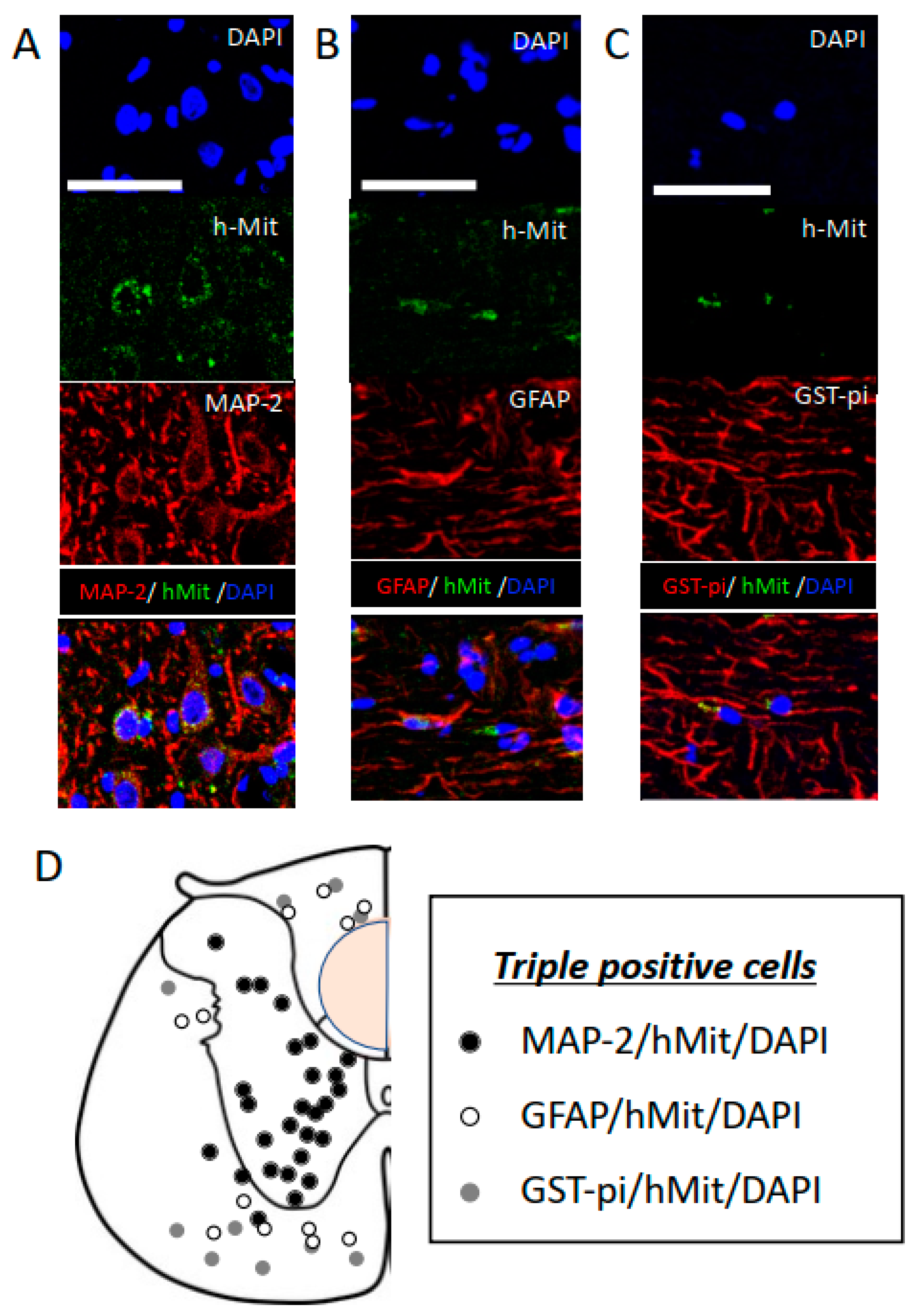
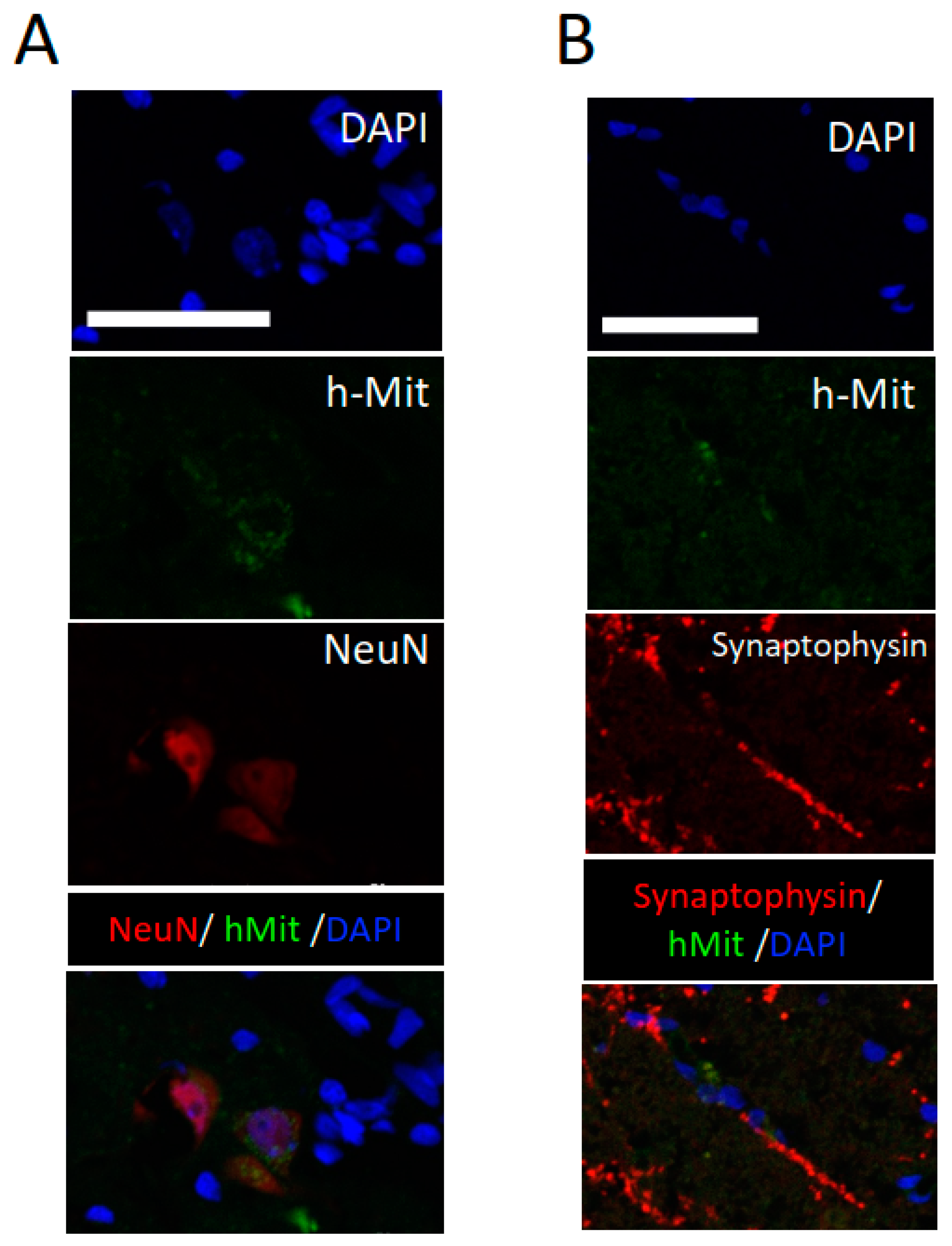
2.3. CL2020 Engrafts and Differentiates into Neuronal Cells in the Injured Spinal Cord
2.4. Deterioration of Hindlimb Functions after the Administration of Diphtheria Toxin
3. Discussion
3.1. Muse Cells Migrate to the Injured Spinal Cord
3.2. Mechanisms Underlying Muse Cell Migration and Homing
3.3. Muse Cells Differentiate into Neuronal Lineage Cells
3.4. Muse Cells Contribute to Functional Recovery
3.5. Limitations
4. Materials and Methods
4.1. Experimental Animals
4.2. SCI
4.3. Intravenous Administration of CL2020 (Muse Cells)
4.4. Behavioral Analysis
4.5. Immunohistochemical Analyses
4.6. Cystic Cavity and Spared Spinal Cord Tissue Measurement
4.7. Quantification of Preserved 5-HT Fibers
4.8. Identification of Muse Cells in the Injured Spinal Cord
4.9. Administration of Diphtheria Toxin
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Filbin, M.T. Myelin-Associated Inhibitors of Axonal Regeneration in the Adult Mammalian CNS. Nat. Rev. Neurosci. 2003, 4, 703–713. [Google Scholar] [CrossRef]
- Yiu, G.; He, Z. Glial Inhibition of CNS Axon Regeneration. Nat. Rev. Neurosci. 2006, 7, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Hofstetter, C.P.; Holmström, N.A.; Lilja, J.A.; Schweinhardt, P.; Hao, J.; Spenger, C.; Wiesenfeld-Hallin, Z.; Kurpad, S.N.; Frisén, J.; Olson, L. Allodynia limits the usefulness of intraspinal neural stem cell grafts; directed differentiation improves outcome. Nat. Neurosci. 2005, 8, 346–353. [Google Scholar] [CrossRef]
- Kuroda, Y.; Kitada, M.; Wakao, S.; Nishikawa, K.; Tanimura, Y.; Makinoshima, H.; Goda, M.; Akashi, H.; Inutsuka, A.; Niwa, A.; et al. Unique Multipotent Cells in Adult Human Mesenchymal Cell Populations. Proc. Natl. Acad. Sci. USA 2010, 107, 8639–8643. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, Y.; Wakao, S.; Kitada, M.; Murakami, T.; Nojima, M.; Dezawa, M. Isolation, Culture and Evaluation of Multilineage-Differentiating Stress-Enduring (Muse) Cells. Nat. Protoc. 2013, 8, 1391–1415. [Google Scholar] [CrossRef] [PubMed]
- Wakao, S.; Kitada, M.; Kuroda, Y.; Shigemoto, T.; Matsuse, D.; Akashi, H.; Tanimura, Y.; Tsuchiyama, K.; Kikuchi, T.; Goda, M.; et al. Multilineage-Differentiating Stress-Enduring (Muse) Cells Are a Primary Source of Induced Pluripotent Stem Cells in Human Fibroblasts. Proc. Natl. Acad. Sci. USA 2011, 108, 9875–9880. [Google Scholar] [CrossRef]
- Young, W. Future of Muse Cells. Adv. Exp. Med. Biol. 2018, 1103, 309–315. [Google Scholar] [CrossRef]
- Yamada, Y.; Wakao, S.; Kushida, Y.; Minatoguchi, S.; Mikami, A.; Higashi, K.; Baba, S.; Shigemoto, T.; Kuroda, Y.; Kanamori, H.; et al. S1P-S1PR2 Axis Mediates Homing of Muse Cells Into Damaged Heart for Long-Lasting Tissue Repair and Functional Recovery After Acute Myocardial Infarction. Circ. Res. 2018, 122, 1069–1083. [Google Scholar] [CrossRef]
- Uchida, H.; Niizuma, K.; Kushida, Y.; Wakao, S.; Tominaga, T.; Borlongan, C.V.; Dezawa, M. Human Muse Cells Reconstruct Neuronal Circuitry in Subacute Lacunar Stroke Model. Stroke 2017, 48, 428–435. [Google Scholar] [CrossRef]
- Kajitani, T.; Endo, T.; Iwabuchi, N.; Inoue, T.; Takahashi, Y.; Abe, T.; Niizuma, K.; Tominaga, T. Association of Intravenous Administration of Human Muse Cells with Deficit Amelioration in a Rat Model of Spinal Cord Injury. J. Neurosurg. Spine 2021, 34, 648–655. [Google Scholar] [CrossRef]
- Nakamura, M.; Okano, H. Cell Transplantation Therapies for Spinal Cord Injury Focusing on Induced Pluripotent Stem Cells. Cell Res. 2013, 23, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Iseki, M.; Kushida, Y.; Wakao, S.; Akimoto, T.; Mizuma, M.; Motoi, F.; Asada, R.; Shimizu, S.; Unno, M.; Chazenbalk, G.; et al. Muse Cells, Nontumorigenic Pluripotent-Like Stem Cells, Have Liver Regeneration Capacity Through Specific Homing and Cell Replacement in a Mouse Model of Liver Fibrosis. Cell Transplant. 2017, 26, 821–840. [Google Scholar] [CrossRef] [PubMed]
- Katagiri, H.; Kushida, Y.; Nojima, M.; Kuroda, Y.; Wakao, S.; Ishida, K.; Endo, F.; Kume, K.; Takahara, T.; Nitta, H.; et al. A Distinct Subpopulation of Bone Marrow Mesenchymal Stem Cells, Muse Cells, Directly Commit to the Replacement of Liver Components. Am. J. Transplant. 2016, 16, 468–483. [Google Scholar] [CrossRef] [PubMed]
- Kimura, A.; Ohmori, T.; Ohkawa, R.; Madoiwa, S.; Mimuro, J.; Murakami, T.; Kobayashi, E.; Hoshino, Y.; Yatomi, Y.; Sakata, Y. Essential Roles of Sphingosine 1-Phosphate/S1P1 Receptor Axis in the Migration of Neural Stem Cells toward a Site of Spinal Cord Injury. Stem Cells 2007, 25, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Kushida, Y.; Wakao, S.; Dezawa, M. Muse Cells Are Endogenous Reparative Stem Cells. Adv. Exp. Med. Biol. 2018, 1103, 43–68. [Google Scholar] [CrossRef]
- Hori, Y.; Kitani, T.; Yanishi, K.; Suga, T.; Kogure, M.; Kusaba, T.; Kushida, Y.; Dezawa, M.; Matoba, S. Intravenous Administration of Human Muse Cells Recovers Blood Flow in a Mouse Model of Hindlimb Ischemia. Front. Cardiovasc. Med. 2022, 9, 981088. [Google Scholar] [CrossRef]
- Uchida, N.; Kushida, Y.; Kitada, M.; Wakao, S.; Kumagai, N.; Kuroda, Y.; Kondo, Y.; Hirohara, Y.; Kure, S.; Chazenbalk, G.; et al. Beneficial Effects of Systemically Administered Human Muse Cells in Adriamycin Nephropathy. J. Am. Soc. Nephrol. 2017, 28, 2946–2960. [Google Scholar] [CrossRef]
- Rizzo, R.; Bortolotti, D.; Bolzani, S.; Fainardi, E. HLA-G Molecules in Autoimmune Diseases and Infections. Front. Immunol. 2014, 5, 592. [Google Scholar] [CrossRef]
- Hwu, P.; Du, M.X.; Lapointe, R.; Do, M.; Taylor, M.W.; Young, H.A. Indoleamine 2,3-Dioxygenase Production by Human Dendritic Cells Results in the Inhibition of T Cell Proliferation. J. Immunol. 2000, 164, 3596–3599. [Google Scholar] [CrossRef]
- Olson, L. Combinatory Treatments Needed for Spinal Cord Injury. Exp. Neurol. 2013, 248, 309–315. [Google Scholar] [CrossRef]
- Abe, T.; Aburakawa, D.; Niizuma, K.; Iwabuchi, N.; Kajitani, T.; Wakao, S.; Kushida, Y.; Dezawa, M.; Borlongan, C.V.; Tominaga, T. Intravenously Transplanted Human Multilineage-Differentiating Stress-Enduring Cells Afford Brain Repair in a Mouse Lacunar Stroke Model. Stroke 2020, 51, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Abematsu, M.; Tsujimura, K.; Yamano, M.; Saito, M.; Kohno, K.; Kohyama, J.; Namihira, M.; Komiya, S.; Nakashima, K. Neurons Derived from Transplanted Neural Stem Cells Restore Disrupted Neuronal Circuitry in a Mouse Model of Spinal Cord Injury. J. Clin. Investig. 2010, 120, 3255–3266. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yin, X.-Y.; Zhao, Y.-Y.; Wang, C.-C.; Du, P.; Lu, Y.-C.; Jin, H.-B.; Yang, C.-C.; Ying, J.-L. Human Muse Cells-Derived Neural Precursor Cells as the Novel Seed Cells for the Repair of Spinal Cord Injury. Biochem. Biophys. Res. Commun. 2021, 568, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Nistor, G.I.; Totoiu, M.O.; Haque, N.; Carpenter, M.K.; Keirstead, H.S. Human Embryonic Stem Cells Differentiate into Oligodendrocytes in High Purity and Myelinate after Spinal Cord Transplantation. Glia 2005, 49, 385–396. [Google Scholar] [CrossRef]
- Allard, J.; Li, K.; Lopez, X.M.; Blanchard, S.; Barbot, P.; Rorive, S.; Decaestecker, C.; Pochet, R.; Bohl, D.; Lepore, A.C.; et al. Immunohistochemical Toolkit for Tracking and Quantifying Xenotransplanted Human Stem Cells. Regen. Med. 2014, 9, 437–452. [Google Scholar] [CrossRef]
- Wakao, S.; Oguma, Y.; Kushida, Y.; Kuroda, Y.; Tatsumi, K.; Dezawa, M. Phagocytosing Differentiated Cell-Fragments Is a Novel Mechanism for Controlling Somatic Stem Cell Differentiation within a Short Time Frame. Cell Mol. Life Sci. 2022, 79, 542. [Google Scholar] [CrossRef]
- Uchida, H.; Morita, T.; Niizuma, K.; Kushida, Y.; Kuroda, Y.; Wakao, S.; Sakata, H.; Matsuzaka, Y.; Mushiake, H.; Tominaga, T.; et al. Transplantation of Unique Subpopulation of Fibroblasts, Muse Cells, Ameliorates Experimental Stroke Possibly via Robust Neuronal Differentiation. Stem Cells 2016, 34, 160–173. [Google Scholar] [CrossRef]
- Basso, D.M.; Beattie, M.S.; Bresnahan, J.C. A Sensitive and Reliable Locomotor Rating Scale for Open Field Testing in Rats. J. Neurotrauma 1995, 12, 1–21. [Google Scholar] [CrossRef]
- Pappenheimer, A.M.; Harper, A.A.; Moynihan, M.; Brockes, J.P. Diphtheria Toxin and Related Proteins: Effect of Route of Injection on Toxicity and the Determination of Cytotoxicity for Various Cultured Cells. J. Infect Dis. 1982, 145, 94–102. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MAP-2 | GFAP | GST-pi | |
|---|---|---|---|
| Rostral to the injury center | 54.0 ± 3.5% | 24.0 ± 2.5% | 22.0 ± 1.9% |
| Injury center | 48.4 ± 2.8% | 26.6 ± 6.0% | 25.0 ± 5.0% |
| Caudal to the injury center | 52.3 ± 4.6% | 25.6 ± 1.7% | 22.1 ± 3.1% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takahashi, Y.; Kajitani, T.; Endo, T.; Nakayashiki, A.; Inoue, T.; Niizuma, K.; Tominaga, T. Intravenous Administration of Human Muse Cells Ameliorates Deficits in a Rat Model of Subacute Spinal Cord Injury. Int. J. Mol. Sci. 2023, 24, 14603. https://doi.org/10.3390/ijms241914603
Takahashi Y, Kajitani T, Endo T, Nakayashiki A, Inoue T, Niizuma K, Tominaga T. Intravenous Administration of Human Muse Cells Ameliorates Deficits in a Rat Model of Subacute Spinal Cord Injury. International Journal of Molecular Sciences. 2023; 24(19):14603. https://doi.org/10.3390/ijms241914603
Chicago/Turabian StyleTakahashi, Yoshiharu, Takumi Kajitani, Toshiki Endo, Atsushi Nakayashiki, Tomoo Inoue, Kuniyasu Niizuma, and Teiji Tominaga. 2023. "Intravenous Administration of Human Muse Cells Ameliorates Deficits in a Rat Model of Subacute Spinal Cord Injury" International Journal of Molecular Sciences 24, no. 19: 14603. https://doi.org/10.3390/ijms241914603
APA StyleTakahashi, Y., Kajitani, T., Endo, T., Nakayashiki, A., Inoue, T., Niizuma, K., & Tominaga, T. (2023). Intravenous Administration of Human Muse Cells Ameliorates Deficits in a Rat Model of Subacute Spinal Cord Injury. International Journal of Molecular Sciences, 24(19), 14603. https://doi.org/10.3390/ijms241914603