

Brain Plasticity in Patients with Spinal Cord Injuries: A Systematic Review
 , , , and
, , , and 
Abstract
1. Introduction
2. Methods
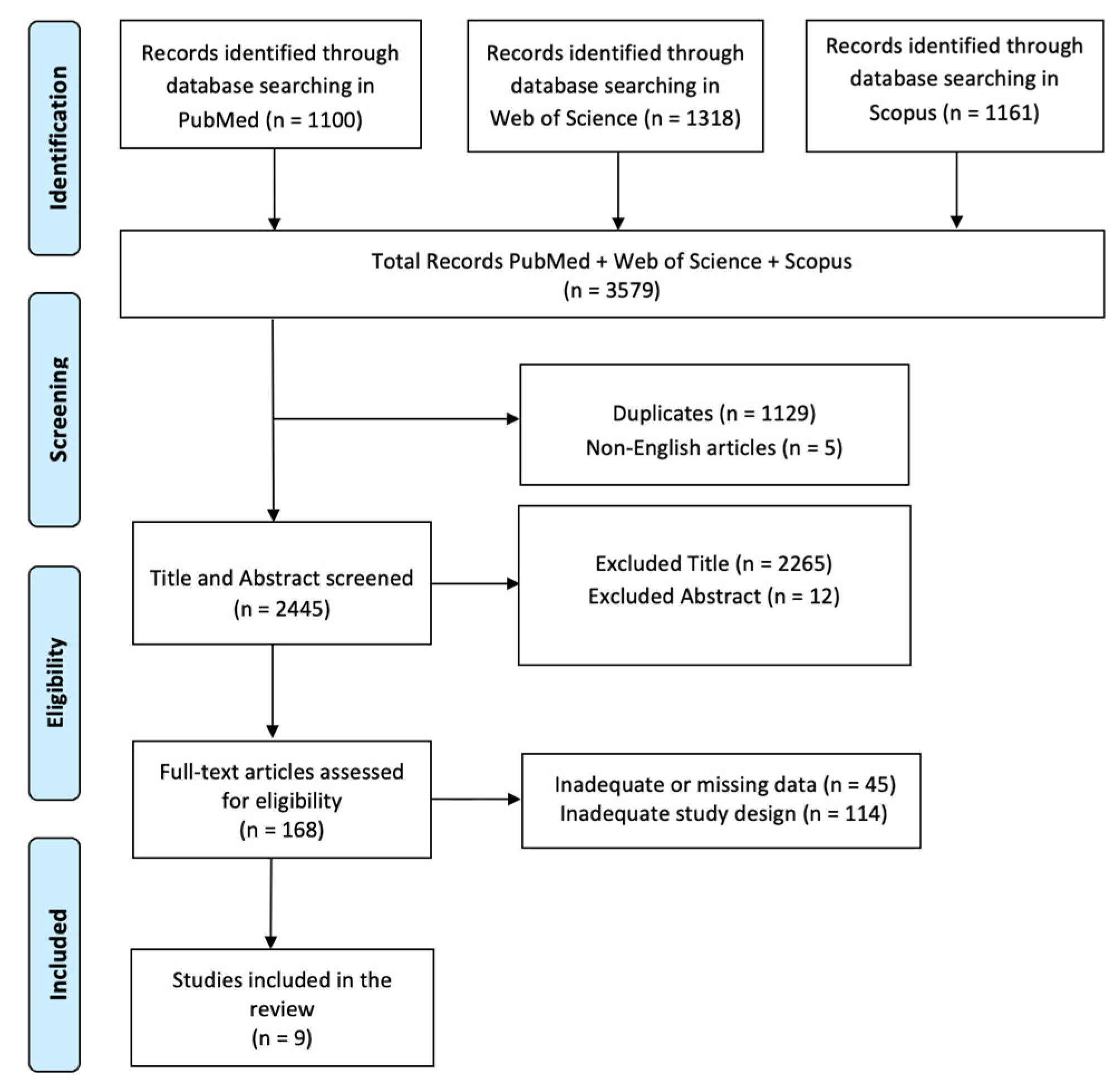
2.1. Search Strategy
2.2. PICO Evaluation
2.3. Inclusion Criteria
2.4. Exclusion Criteria
3. Results and Discussion
3.1. Cortical Sensory-Motor Plasticity in Patient with SCI
3.2. Rehabilitation, Neural Network, and Functional Recovery in SCI
3.3. Discussion
4. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Solstrand Dahlberg, L.; Becerra, L.; Borsook, D.; Linnman, C. Brain changes after spinal cord injury, a quantitative meta-analysis and review. Neurosci. Biobehav. Rev. 2018, 90, 272–293. [Google Scholar] [CrossRef]
- Fehlings, M.G.; Tetreault, L.A.; Wilson, J.R.; Kwon, B.K.; Burns, A.S.; Martin, A.R.; Hawryluk, G.; Harrop, J.S. A Clinical Practice Guideline for the Management of Acute Spinal Cord Injury: Introduction, Rationale, and Scope. Glob. Spine J. 2017, 7, 84S–94S. [Google Scholar] [CrossRef]
- Anderson, K.D. Targeting recovery: Priorities of the spinal cord-injured population. J. Neurotrauma 2004, 21, 1371–1383. [Google Scholar] [CrossRef]
- Fawcett, J.W.; Curt, A.; Steeves, J.D.; Coleman, W.P.; Tuszynski, M.H.; Lammertse, D.; Bartlett, P.F.; Blight, A.R.; Dietz, V.; Ditunno, J.; et al. Guidelines for the conduct of clinical trials for spinal cord injury as developed by the ICCP panel: Spontaneous recovery after spinal cord injury and statistical power needed for therapeutic clinical trials. Spinal Cord 2007, 45, 190–205. [Google Scholar] [CrossRef]
- Rabchevsky, A.G.; Kitzman, P.H. Latest approaches for the treatment of spasticity and autonomic dysreflexia in chronic spinal cord injury. Neurotherapeutics 2011, 8, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Hiersemenzel, L.-P.; Curt, A.; Dietz, V. From spinal shock to spasticity: Neuronal adaptations to a spinal cord injury. Neurology 2000, 54, 1574–1582. [Google Scholar] [CrossRef] [PubMed]
- Basbaum, A.; Wall, P. Chronic changes in the response of cells in adult cat dorsal horn following partial deafferentation: The appearance of responding cells in a previously nonresponsive region. Brain Res. 1976, 116, 181–204. [Google Scholar] [CrossRef] [PubMed]
- Fawcett, J. Repair of spinal cord injuries: Where are we, where are we going? Spinal Cord 2002, 40, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Fouad, K.; Tse, A. Adaptive changes in the injured spinal cord and their role in promoting functional recovery. Neurol. Res. 2008, 30, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.N.; Chambers, W.W. Intraspinal sprouting of dorsal root axons; development of new collaterals and preterminals following partial denervation of the spinal cord in the cat. AMA Arch. Neurol. Psychiatry 1958, 79, 46–61. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.; Goldberger, M.E. Restitution of function and collateral sprouting in the cat spinal cord: The partially hemisected animal. J. Comp. Neurol. 1974, 158, 19–36. [Google Scholar] [CrossRef]
- Bruehlmeier, M.; Dietz, V.; Leenders, K.L.; Roelcke, U.; Missimer, J.; Curt, A. How does the human brain deal with a spinal cord injury? Eur. J. Neurosci. 1998, 10, 3918–3922. [Google Scholar] [CrossRef] [PubMed]
- Endo, T.; Spenger, C.; Tominaga, T.; Brené, S.; Olson, L. Cortical sensory map rearrangement after spinal cord injury: fMRI responses linked to Nogo signalling. Brain 2007, 130, 2951–2961. [Google Scholar] [CrossRef] [PubMed]
- Kaas, J.H.; Qi, H.-X.; Burish, M.J.; Gharbawie, O.A.; Onifer, S.M.; Massey, J.M. Cortical and subcortical plasticity in the brains of humans, primates, and rats after damage to sensory afferents in the dorsal columns of the spinal cord. Exp. Neurol. 2008, 209, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Darian-Smith, C.; Gilbert, C.D. Axonal sprouting accompanies functional reorganization in adult cat striate cortex. Nature 1994, 368, 737–740. [Google Scholar] [CrossRef]
- Fouad, K.; Pedersen, V.; Schwab, M.E.; Brösamle, C. Cervical sprouting of corticospinal fibers after thoracic spinal cord injury accompanies shifts in evoked motor responses. Curr. Biol. 2001, 11, 1766–1770. [Google Scholar] [CrossRef]
- Girgis, J.; Merrett, D.; Kirkland, S.; Metz, G.A.S.; Verge, V.; Fouad, K. Reaching training in rats with spinal cord injury promotes plasticity and task specific recovery. Brain 2007, 130, 2993–3003. [Google Scholar] [CrossRef]
- Raineteau, O.; Schwab, M.E. Plasticity of motor systems after incomplete spinal cord injury. Nat. Rev. Neurosci. 2001, 2, 263–273. [Google Scholar] [CrossRef]
- Florence, S.; Kaas, J. Large-scale reorganization at multiple levels of the somatosensory pathway follows therapeutic amputation of the hand in monkeys. J. Neurosci. 1995, 15, 8083–8095. [Google Scholar] [CrossRef]
- Pons, T.P.; Garraghty, P.E.; Ommaya, A.K.; Kaas, J.H.; Taub, E.; Mishkin, M. Massive cortical reorganization after sensory deafferentation in adult macaques. Science 1991, 252, 1857–1860. [Google Scholar] [CrossRef]
- Raineteau, O.; Fouad, K.; Noth, P.; Thallmair, M.; Schwab, M.E. Functional switch between motor tracts in the presence of the mAb IN-1 in the adult rat. Proc. Natl. Acad. Sci. USA 2001, 98, 6929–6934. [Google Scholar] [CrossRef]
- Aoki, M.; Fujito, Y.; Satomi, H.; Kurosawa, Y.; Kasaba, T. The possible role of collateral sprouting in the functional restitution of corticospinal connections after spinal hemisection. Neurosci. Res. 1986, 3, 617–627. [Google Scholar] [CrossRef]
- Ballermann, M.; Fouad, K. Spontaneous locomotor recovery in spinal cord injured rats is accompanied by anatomical plasticity of reticulospinal fibers. Eur. J. Neurosci. 2006, 23, 1988–1996. [Google Scholar] [CrossRef] [PubMed]
- Bareyre, F.M.; Kerschensteiner, M.; Raineteau, O.; Mettenleiter, T.C.; Weinmann, O.; Schwab, M.E. The injured spinal cord spontaneously forms a new intraspinal circuit in adult rats. Nat. Neurosci. 2004, 7, 269–277. [Google Scholar] [CrossRef]
- Carp, J.S.; Wolpaw, J.R. Motoneuron plasticity underlying operantly conditioned decrease in primate H-reflex. J. Neurophysiol. 1994, 72, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Darian-Smith, C. Primary afferent terminal sprouting after a cervical dorsal rootlet section in the macaque monkey. J. Comp. Neurol. 2004, 470, 134–150. [Google Scholar] [CrossRef] [PubMed]
- Hagg, T.; Oudega, M. Degenerative and spontaneous regenerative processes after spinal cord injury. J. Neurotrauma 2006, 23, 263–280. [Google Scholar] [CrossRef] [PubMed]
- Weidner, N.; Ner, A.; Salimi, N.; Tuszynski, M.H. Spontaneous corticospinal axonal plasticity and functional recovery after adult central nervous system injury. Proc. Natl. Acad. Sci. USA 2001, 98, 3513–3518. [Google Scholar] [CrossRef]
- Jain, N.; Florence, S.L.; Qi, H.-X.; Kaas, J.H. Growth of new brainstem connections in adult monkeys with massive sensory loss. Proc. Natl. Acad. Sci. USA 2000, 97, 5546–5550. [Google Scholar] [CrossRef]
- Vavrek, R.; Girgis, J.; Tetzlaff, W.; Hiebert, G.W.; Fouad, K. BDNF promotes connections of corticospinal neurons onto spared descending interneurons in spinal cord injured rats. Brain 2006, 129, 1534–1545. [Google Scholar] [CrossRef]
- Kim, B.G.; Dai, H.-N.; McAtee, M.; Vicini, S.; Bregman, B.S. Remodeling of synaptic structures in the motor cortex following spinal cord injury. Exp. Neurol. 2006, 198, 401–415. [Google Scholar] [CrossRef] [PubMed]
- Li, G.L.; Farooque, M.; Isaksson, J.; Olsson, Y. Changes in synapses and axons demonstrated by synaptophysin immunohistochemistry following spinal cord compression trauma in the rat and mouse. Biomed. Environ. Sci. 2004, 17, 281–290. [Google Scholar] [PubMed]
- Harvey, P.J.; Lucas-Osma, A.M.; Li, Y.; Lin, S.; Black, S.; Singla, R.; Fouad, K.; Fenrich, K.K.; Bennett, D.J.; Dai, Y.; et al. Persistent sodium currents and repetitive firing in motoneurons of the sacrocaudal spinal cord of adult rats. J. Neurophysiol. 2006, 96, 1141–1157. [Google Scholar] [CrossRef] [PubMed]
- Ceto, S.; Sekiguchi, K.J.; Takashima, Y.; Nimmerjahn, A.; Tuszynski, M.H. Neural Stem Cell Grafts Form Extensive Synaptic Networks that Integrate with Host Circuits after Spinal Cord Injury. Cell Stem Cell 2020, 27, 430–440.e5. [Google Scholar] [CrossRef] [PubMed]
- Oudega, M.; Perez, M.A. Corticospinal reorganization after spinal cord injury. J. Physiol. 2012, 590, 3647–3663. [Google Scholar] [CrossRef]
- Lemon, R.N. Descending pathways in motor control. Annu. Rev. Neurosci. 2008, 31, 195–218. [Google Scholar] [CrossRef]
- Bouvier, J.; Caggiano, V.; Leiras, R.; Caldeira, V.; Bellardita, C.; Balueva, K.; Fuchs, A.; Kiehn, O. Descending Command Neurons in the Brainstem that Halt Locomotion. Cell 2015, 163, 1191–1203. [Google Scholar] [CrossRef]
- Filli, L.; Engmann, A.K.; Zörner, B.; Weinmann, O.; Moraitis, T.; Gullo, M.; Kasper, H.; Schneider, R.; Schwab, M.E. Bridging the gap: A reticulo-propriospinal detour bypassing an incomplete spinal cord injury. J. Neurosci. 2014, 34, 13399–13410. [Google Scholar] [CrossRef]
- Fouad, K.; Schnell, L.; Bunge, M.B.; Schwab, M.E.; Liebscher, T.; Pearse, D.D. Combining schwann cell bridges and olfactory-ensheathing glia grafts with chondroitinase promotes locomotor recovery after complete transection of the spinal cord. J. Neurosci. 2005, 25, 1169–1178. [Google Scholar] [CrossRef]
- Huntley, G.W. Correlation between patterns of horizontal connectivity and the extend of short-term representational plasticity in rat motor cortex. Cereb. Cortex 1997, 7, 143–156. [Google Scholar] [CrossRef]
- Weiss, D.S.; Keller, A. Specific patterns of intrinsic connections between representation zones in the rat motor cortex. Cereb. Cortex 1994, 4, 205–214. [Google Scholar] [CrossRef]
- Murray, K.C.; Nakae, A.; Stephens, M.J.; Rank, M.; D’Amico, J.; Harvey, P.J.; Li, X.; Harris, R.L.W.; Ballou, E.W.; Anelli, R.; et al. Recovery of motoneuron and locomotor function after spinal cord injury depends on constitutive activity in 5-HT2C receptors. Nat. Med. 2010, 16, 694–700. [Google Scholar] [CrossRef]
- Takeoka, A.; Vollenweider, I.; Courtine, G.; Arber, S. Muscle spindle feedback directs locomotor recovery and circuit reorganization after spinal cord injury. Cell 2014, 159, 1626–1639. [Google Scholar] [CrossRef]
- Locke, K.C.; Randelman, M.L.; Hoh, D.J.; Zholudeva, L.V.; Lane, M.A. Respiratory plasticity following spinal cord injury: Perspectives from mouse to man. Neural Regen Res. 2022, 17, 2141–2148. [Google Scholar] [CrossRef]
- Rosenzweig, E.S.; Courtine, G.; Jindrich, D.L.; Brock, J.H.; Ferguson, A.R.; Strand, S.C.; Nout, Y.S.; Roy, R.R.; Miller, D.M.; Beattie, M.S.; et al. Extensive spontaneous plasticity of corticospinal projections after primate spinal cord injury. Nat. Neurosci. 2010, 13, 1505–1510. [Google Scholar] [CrossRef]
- Friedli, L.; Rosenzweig, E.S.; Barraud, Q.; Schubert, M.; Dominici, N.; Awai, L.; Nielson, J.L.; Musienko, P.; Nout-Lomas, Y.; Zhong, H.; et al. Pronounced species divergence in corticospinal tract reorganization and functional recovery after lateralized spinal cord injury favors primates. Sci. Transl. Med. 2015, 7, 302ra134. [Google Scholar] [CrossRef]
- Belhaj-Saïf, A.; Cheney, P.D.; Sugiyama, Y.; Higo, N.; Yoshino-Saito, K.; Murata, Y.; Nishimura, Y.; Oishi, T.; Isa, T.; Umeda, T.; et al. Plasticity in the distribution of the red nucleus output to forearm muscles after unilateral lesions of the pyramidal tract. J. Neurophysiol. 2000, 83, 3147–3153. [Google Scholar] [CrossRef] [PubMed]
- Müllner, A.; Gonzenbach, R.R.; Weinmann, O.; Schnell, L.; Liebscher, T.; Schwab, M.E. Lamina-specific restoration of serotonergic projections after Nogo-A antibody treatment of spinal cord injury in rats. Eur. J. Neurosci. 2008, 27, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Hilton, B.J.; Anenberg, E.; Harrison, T.C.; Boyd, J.D.; Murphy, T.H.; Tetzlaff, W. Re-Establishment of Cortical Motor Output Maps and Spontaneous Functional Recovery via Spared Dorsolaterally Projecting Corticospinal Neurons after Dorsal Column Spinal Cord Injury in Adult Mice. J. Neurosci. 2016, 36, 4080–4092. [Google Scholar] [CrossRef] [PubMed]
- Hollis, E.R.; Ishiko, N.; Yu, T.; Lu, C.-C.; Haimovich, A.; Tolentino, K.; Richman, A.; Tury, A.; Wang, S.-H.; Pessian, M.; et al. Ryk controls remapping of motor cortex during functional recovery after spinal cord injury. Nat. Neurosci. 2016, 19, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Asboth, L.; Friedli, L.; Beauparlant, J.; Martinez-Gonzalez, C.; Anil, S.; Rey, E.; Baud, L.; Pidpruzhnykova, G.; Anderson, M.A.; Shkorbatova, P.; et al. Cortico–reticulo–spinal circuit reorganization enables functional recovery after severe spinal cord contusion. Nat. Neurosci. 2018, 21, 576–588. [Google Scholar] [CrossRef]
- Courtine, G.; Song, B.; Roy, R.R.; Zhong, H.; Herrmann, J.E.; Ao, Y.; Qi, J.; Edgerton, V.R.; Sofroniew, M.V. Recovery of supraspinal control of stepping via indirect propriospinal relay connections after spinal cord injury. Nat. Med. 2008, 14, 69–74. [Google Scholar] [CrossRef]
- Ruder, L.; Takeoka, A.; Arber, S. Long-Distance Descending Spinal Neurons Ensure Quadrupedal Locomotor Stability. Neuron 2016, 92, 1063–1078. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Li, Y.; Yu, B.; Zhang, Z.; Brommer, B.; Williams, P.R.; Liu, Y.; Hegarty, S.V.; Zhou, S.; Zhu, J.; et al. Reactivation of Dormant Relay Pathways in Injured Spinal Cord by KCC2 Manipulations. Cell 2018, 174, 1599, Erratum in Cell 2018, 174, 521–535.e13. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, M.; Matsui, R.; Kato, S.; Hasegawa, T.; Kasahara, H.; Isa, K.; Watakabe, A.; Yamamori, T.; Nishimura, Y.; Alstermark, B.; et al. Genetic dissection of the circuit for hand dexterity in primates. Nature 2012, 487, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Beauparlant, J.; van den Brand, R.; Barraud, Q.; Friedli, L.; Musienko, P.; Dietz, V.; Courtine, G. Undirected compensatory plasticity contributes to neuronal dysfunction after severe spinal cord injury. Brain 2013, 136, 3347–3361. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.-Q.; Zaaimi, B.; Martin, J.H. Competition with Primary Sensory Afferents Drives Remodeling of Corticospinal Axons in Mature Spinal Motor Circuits. J. Neurosci. 2016, 36, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Edgerton, V.R.; Tillakaratne, N.J.; Bigbee, A.J.; de Leon, R.D.; Roy, R.R. Plasticity of the spinal neural circuitry after injury. Annu. Rev. Neurosci. 2004, 27, 145–167. [Google Scholar] [CrossRef] [PubMed]
- Navarrete-Opazo, A.; Alcayaga, J.; Sepúlveda, O.; Rojas, E.; Astudillo, C. Repetitive Intermittent Hypoxia and Locomotor Training Enhances Walking Function in Incomplete Spinal Cord Injury Subjects: A Randomized, Triple-Blind, Placebo-Controlled Clinical Trial. J. Neurotrauma 2017, 34, 1803–1812. [Google Scholar] [CrossRef] [PubMed]
- Pearse, D.D.; Pereira, F.C.; Marcillo, A.E.; Bates, M.L.; Berrocal, Y.A.; Filbin, M.T.; Bunge, M.B. cAMP and Schwann cells promote axonal growth and functional recovery after spinal cord injury. Nat. Med. 2004, 10, 610–616. [Google Scholar] [CrossRef]
- Williams, R.R.; Henao, M.; Pearse, D.D.; Bunge, M.B. Permissive schwann cell graft/spinal cord interfaces for axon regeneration. Cell Transplant. 2015, 24, 115–131. [Google Scholar] [CrossRef]
- Shih, C.-H.; Lacagnina, M.; Leuer-Bisciotti, K.; Pröschel, C. Astroglial-derived periostin promotes axonal regeneration after spinal cord injury. J. Neurosci. 2014, 34, 2438–2443. [Google Scholar] [CrossRef]
- Zhang, S.; Alvarez, D.J.; Sofroniew, M.V.; Deming, T.J. Design and synthesis of nonionic copolypeptide hydrogels with reversible thermoresponsive and tunable physical properties. Biomacromolecules 2015, 16, 1331–1340. [Google Scholar] [CrossRef]
- Assinck, P.; Duncan, G.J.; Hilton, B.J.; Plemel, J.R.; Tetzlaff, W. Cell transplantation therapy for spinal cord injury. Nat. Neurosci. 2017, 20, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.D.; Guest, J.D.; Dietrich, W.D.; Bunge, M.B.; Curiel, R.; Dididze, M.; Green, B.A.; Khan, A.; Pearse, D.D.; Saraf-Lavi, E.; et al. Safety of Autologous Human Schwann Cell Transplantation in Subacute Thoracic Spinal Cord Injury. J. Neurotrauma 2017, 34, 2950–2963. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Wang, Y.; Graham, L.; McHale, K.; Gao, M.; Wu, D.; Brock, J.; Blesch, A.; Rosenzweig, E.S.; Havton, L.A.; et al. Long-distance growth and connectivity of neural stem cells after severe spinal cord injury. Cell 2012, 150, 1264–1273. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Jones, L.; Snyder, E.; Tuszynski, M. Neural stem cells constitutively secrete neurotrophic factors and promote extensive host axonal growth after spinal cord injury. Exp. Neurol. 2003, 181, 115–129. [Google Scholar] [CrossRef]
- Kadoya, K.; Lu, P.; Nguyen, K.; Lee-Kubli, C.; Kumamaru, H.; Yao, L.; Knackert, J.; Poplawski, G.; Dulin, J.N.; Strobl, H.; et al. Spinal cord reconstitution with homologous neural grafts enables robust corticospinal regeneration. Nat. Med. 2016, 22, 479–487. [Google Scholar] [CrossRef]
- Bonner, J.F.; Connors, T.M.; Silverman, W.F.; Kowalski, D.P.; Lemay, M.A.; Fischer, I. Grafted neural progenitors integrate and restore synaptic connectivity across the injured spinal cord. J. Neurosci. 2011, 31, 4675–4686, Erratum in J. Neurosci. 2013, 33, 6705. [Google Scholar] [CrossRef]
- Abematsu, M.; Tsujimura, K.; Yamano, M.; Saito, M.; Kohno, K.; Kohyama, J.; Namihira, M.; Komiya, S.; Nakashima, K. Neurons derived from transplanted neural stem cells restore disrupted neuronal circuitry in a mouse model of spinal cord injury. J. Clin. Investig. 2010, 120, 3255–3266. [Google Scholar] [CrossRef]
- Fehlings, M.G.; Cadotte, D.W.; Fehlings, L.N. A series of systematic reviews on the treatment of acute spinal cord injury: A foundation for best medical practice. J. Neurotrauma 2011, 28, 1329–1333. [Google Scholar] [CrossRef]
- Fisher, A.R.; Siegelman, E.S. Magnetic resonance imaging: Techniques. Clin. Liver Dis. 2002, 6, 53–72. [Google Scholar] [CrossRef]
- Freund, P.; Seif, M.; Weiskopf, N.; Friston, K.; Fehlings, M.G.; Thompson, A.J.; Curt, A. MRI in traumatic spinal cord injury: From clinical assessment to neuroimaging biomarkers. Lancet Neurol. 2019, 18, 1123–1135. [Google Scholar] [CrossRef]
- Weiskopf, N.; Suckling, J.; Williams, G.; Correia, M.M.; Inkster, B.; Tait, R.; Ooi, C.; Bullmore, E.T.; Lutti, A. Quantitative multi-parameter mapping of R1, PD*, MT, and R2* at 3T: A multi-center validation. Front. Neurosci. 2013, 7, 95. [Google Scholar] [CrossRef]
- Pfyffer, D.; Vallotton, K.; Curt, A.; Freund, P. Tissue bridges predict neuropathic pain emergence after spinal cord injury. J. Neurol. Neurosurg. Psychiatry 2020, 91, 1111–1117. [Google Scholar] [CrossRef]
- Kyathanahally, S.P.; Azzarito, M.; Rosner, J.; Calhoun, V.D.; Blaiotta, C.; Ashburner, J.; Weiskopf, N.; Wiech, K.; Friston, K.; Ziegler, G.; et al. Microstructural plasticity in nociceptive pathways after spinal cord injury. J. Neurol. Neurosurg. Psychiatry 2021, 92, 863–871. [Google Scholar] [CrossRef]
- Villanueva, L.; Le Bars, D. The activation of bulbo-spinal controls by peripheral nociceptive inputs: Diffuse noxious inhibitory controls. Biol. Res. 1995, 28, 113–125. [Google Scholar]
- Djouhri, L.; Koutsikou, S.; Fang, X.; McMullan, S.; Lawson, S.N. Spontaneous Pain, Both Neuropathic and Inflammatory, Is Related to Frequency of Spontaneous Firing in Intact C-Fiber Nociceptors. J. Neurosci. 2006, 26, 1281–1292. [Google Scholar] [CrossRef] [PubMed]
- Hains, B.C.; Waxman, S.G. Sodium channel expression and the molecular pathophysiology of pain after SCI. Prog. Brain Res. 2007, 161, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Detloff, M.R.; Fisher, L.C.; McGaughy, V.; Longbrake, E.E.; Popovich, P.G.; Basso, D.M. Remote activation of microglia and pro-inflammatory cytokines predict the onset and severity of below-level neuropathic pain after spinal cord injury in rats. Exp. Neurol. 2008, 212, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Wiech, K. Deconstructing the sensation of pain: The influence of cognitive processes on pain perception. Science 2016, 354, 584–587. [Google Scholar] [CrossRef]
- Hanna, A.S.; Filipp, M.; Travis, B.J.; Henry, S.S.; Idzikowski, E.C.; Magnuson, S.; Loh, M.Y.; Hellenbrand, D.J. Differences in neuroplasticity after spinal cord injury in varying animal models and humans. Neural Regen. Res. 2019, 14, 7–19. [Google Scholar] [CrossRef]
- Rao, J.-S.; Zhao, C.; Bao, S.-S.; Feng, T.; Xu, M. MRI metrics at the epicenter of spinal cord injury are correlated with the stepping process in rhesus monkeys. Exp. Anim. 2022, 71, 139–149. [Google Scholar] [CrossRef]
- Rao, J.-S.; Zhao, C.; Wei, R.-H.; Feng, T.; Bao, S.-S.; Zhao, W.; Tian, Z.; Liu, Z.; Yang, Z.-Y.; Li, X.-G. Neural regeneration therapy after spinal cord injury induces unique brain functional reorganizations in rhesus monkeys. Ann. Med. 2022, 54, 1867–1883. [Google Scholar] [CrossRef] [PubMed]
- Segal, M.E.; Ditunno, J.F.; Staas, W.E. Interinstitutional agreement of individual functional independence measure (FIM) items measured at two sites on one sample of SCI patients. Spinal Cord 1993, 31, 622–631. [Google Scholar] [CrossRef]
- Bluvshtein, V.; Front, L.; Itzkovich, M.; Aidinoff, E.; Gelernter, I.; Hart, J.; Biering-Soerensen, F.; Weeks, C.; Laramee, M.T.; Craven, C.; et al. SCIM III is reliable and valid in a separate analysis for traumatic spinal cord lesions. Spinal Cord 2010, 49, 292–296. [Google Scholar] [CrossRef]
- Itzkovich, M.; Gelernter, I.; Biering-Sorensen, F.; Weeks, C.; Laramee, M.T.; Craven, B.C.; Tonack, M.; Hitzig, S.L.; Glaser, E.; Zeilig, G.; et al. The Spinal Cord Independence Measure (SCIM) version III: Reliability and validity in a multi-center international study. Disabil. Rehabil. 2007, 29, 1926–1933. [Google Scholar] [CrossRef] [PubMed]
- Bestmann, S.; Krakauer, J.W. The uses and interpretations of the motor-evoked potential for understanding behaviour. Exp. Brain Res. 2015, 233, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Cruccu, G.; Aminoff, M.; Curio, G.; Guerit, J.; Kakigi, R.; Mauguiere, F.; Rossini, P.; Treede, R.-D.; Garcia-Larrea, L. Recommendations for the clinical use of somatosensory-evoked potentials. Clin. Neurophysiol. 2008, 119, 1705–1719. [Google Scholar] [CrossRef]
- Lagerburg, V.; Bakkers, M.; Bouwhuis, A.; Hoeijmakers, J.G.; Smit, A.M.; Berg, S.J.V.D.; Boer, I.H.; Der Lee, M.D.B.; Kranendonk, D.; Reulen, J.P.; et al. Contact heat evoked potentials: Normal values and use in small-fiber neuropathy. Muscle Nerve 2015, 51, 743–749. [Google Scholar] [CrossRef]
- Granovsky, Y.; Anand, P.; Nakae, A.; Nascimento, O.; Smith, B.; Sprecher, E.; Valls-Solé, J. Normative data for Aδ contact heat evoked potentials in adult population. Pain 2016, 157, 1156–1163. [Google Scholar] [CrossRef]
- Horowitz, S.H. Neuropathic pain: Is the emperor wearing clothes. In Current Therapy in Pain; Saunders/Elsevier: Philadelphia, PA, USA, 2009; pp. 9–14. [Google Scholar] [CrossRef]
- Weaver, K.R.; Griffioen, M.A.; Klinedinst, N.J.; Galik, E.; Duarte, A.C.; Colloca, L.; Resnick, B.; Dorsey, S.G.; Renn, C.L. Quantitative Sensory Testing Across Chronic Pain Conditions and Use in Special Populations. Front. Pain Res. 2022, 2, 779068. [Google Scholar] [CrossRef]
- Mücke, M.; Cuhls, H.; Radbruch, L.; Baron, R.; Maier, C.; Tölle, T.; Treede, R.-D.; Rolke, R. Quantitative sensory testing (QST). English version. Der Schmerz 2014, 35, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Ditunno, P.L.; Patrick, M.; Stineman, M.; Ditunno, J.F. Who wants to walk? Preferences for recovery after SCI: A longitudinal and cross-sectional study. Spinal Cord 2008, 46, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Truini, A.; Panuccio, G.; Galeotti, F.; Maluccio, M.; Sartucci, F.; Avoli, M.; Cruccu, G. Laser-evoked potentials as a tool for assessing the efficacy of antinociceptive drugs. Eur. J. Pain 2010, 14, 222–225. [Google Scholar] [CrossRef]
- Barbeau, H.; McCrea, D.A.; O’Donovan, M.J.; Rossignol, S.; Grill, W.M.; Lemay, M.A. Tapping into spinal circuits to restore motor function. Brain Res. Rev. 1999, 30, 27–51. [Google Scholar] [CrossRef] [PubMed]
- Wernig, A.; Müller, S. Laufband locomotion with body weight support improved walking in persons with severe spinal cord injuries. Spinal Cord 1992, 30, 229–238. [Google Scholar] [CrossRef]
- Grillner, S. Neurobiological Bases of Rhythmic Motor Acts in Vertebrates. Science 1985, 228, 143–149. [Google Scholar] [CrossRef]
- Sherrington, C.S. Flexion-reflex of the limb, crossed extension-reflex, and reflex stepping and standing. J. Physiol. 1910, 40, 28–121. [Google Scholar] [CrossRef]
- Rossignol, S.; Dubuc, R.; Gossard, J.-P. Dynamic Sensorimotor Interactions in Locomotion. Physiol. Rev. 2006, 86, 89–154. [Google Scholar] [CrossRef]
- Dietz, V.; Sinkjaer, T. Spastic movement disorder: Impaired reflex function and altered muscle mechanics. Lancet Neurol. 2007, 6, 725–733. [Google Scholar] [CrossRef]
- Pearson, K.G. Neural Adaptation in the Generation of Rhythmic Behavior. Annu. Rev. Physiol. 2000, 62, 723–753. [Google Scholar] [CrossRef]
- Dietz, V. Proprioception and locomotor disorders. Nat. Rev. Neurosci. 2002, 3, 781–790. [Google Scholar] [CrossRef]
- Versace, V.; Langthaler, P.B.; Höller, Y.; Frey, V.N.; Brigo, F.; Sebastianelli, L.; Saltuari, L.; Nardone, R. Abnormal cortical neuroplasticity induced by paired associative stimulation after traumatic spinal cord injury: A preliminary study. Neurosci. Lett. 2017, 664, 167–171. [Google Scholar] [CrossRef]
- Jo, H.J.; Kizziar, E.; Sangari, S.; Chen, D.; Kessler, A.; Kim, K.; Anschel, A.; Heinemann, A.W.; Mensh, B.D.; Awadalla, S.; et al. Multisite Hebbian Plasticity Restores Function in Humans with Spinal Cord Injury. Ann. Neurol. 2023, 93, 1198–1213. [Google Scholar] [CrossRef]
- Khan, A.S.; Patrick, S.K.; Roy, F.D.; Gorassini, M.A.; Yang, J.F. Training-Specific Neural Plasticity in Spinal Reflexes after Incomplete Spinal Cord Injury. Neural Plast. 2016, 2016, 6718763. [Google Scholar] [CrossRef]
- Seáñez-González, I.; Pierella, C.; Farshchiansadegh, A.; Thorp, E.B.; Wang, X.; Parrish, T.; Mussa-Ivaldi, F.A. Body-Machine Interfaces after Spinal Cord Injury: Rehabilitation and Brain Plasticity. Brain Sci. 2016, 6, 61. [Google Scholar] [CrossRef] [PubMed]
- Jo, H.J.; Perez, M.A. Corticospinal-motor neuronal plasticity promotes exercise-mediated recovery in humans with spinal cord injury. Brain 2020, 143, 1368–1382. [Google Scholar] [CrossRef] [PubMed]
- Faw, T.D.; Lakhani, B.; Schmalbrock, P.; Knopp, M.V.; Lohse, K.R.; Kramer, J.L.; Liu, H.; Nguyen, H.T.; Phillips, E.G.; Bratasz, A.; et al. Eccentric rehabilitation induces white matter plasticity and sensorimotor recovery in chronic spinal cord injury. Exp. Neurol. 2021, 346, 113853. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.; Díaz, F.; Sumich, A. Long-term neuroplasticity in spinal cord injury patients: A study on movement-related brain potentials. Int. J. Psychophysiol. 2013, 87, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Jutzeler, C.R.; Freund, P.; Huber, E.; Curt, A.; Kramer, J.L. Neuropathic Pain and Functional Reorganization in the Primary Sensorimotor Cortex After Spinal Cord Injury. J. Pain 2015, 16, 1256–1267. [Google Scholar] [CrossRef]
- Villiger, M.; Grabher, P.; Hepp-Reymond, M.-C.; Kiper, D.; Curt, A.; Bolliger, M.; Hotz-Boendermaker, S.; Kollias, S.; Eng, K.; Freund, P. Relationship between structural brainstem and brain plasticity and lower-limb training in spinal cord injury: A longitudinal pilot study. Front. Hum. Neurosci. 2015, 9, 254. [Google Scholar] [CrossRef]
- Pascual-Leone, A.; Nguyet, D.; Brasil-Neto, J.P.; Cammarota, A.; Seidel, O.; Carius, D.; Kenville, R.; Ragert, P.; Stöckel, T.; Carroll, T.J.; et al. Modulation of muscle responses evoked by transcranial magnetic stimulation during the acquisition of new fine motor skills. J. Neurophysiol. 1995, 74, 1037–1045. [Google Scholar] [CrossRef] [PubMed]
- Plautz, E.J.; Milliken, G.W.; Nudo, R.J. Effects of Repetitive Motor Training on Movement Representations in Adult Squirrel Monkeys: Role of Use versus Learning. Neurobiol. Learn. Mem. 2000, 74, 27–55. [Google Scholar] [CrossRef] [PubMed]
- Papale, A.E.; Hooks, B.M. Circuit Changes in Motor Cortex During Motor Skill Learning. Neuroscience 2017, 368, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Kleim, J.A.; Bruneau, R.; Calder, K.; Pocock, D.; VandenBerg, P.M.; MacDonald, E.; Monfils, M.H.; Sutherland, R.J.; Nader, K. Functional Organization of Adult Motor Cortex Is Dependent upon Continued Protein Synthesis. Neuron 2003, 40, 167–176. [Google Scholar] [CrossRef]
- Xu, T.; Yu, X.; Perlik, A.J.; Tobin, W.F.; Zweig, J.A.; Tennant, K.; Jones, T.; Zuo, Y. Rapid formation and selective stabilization of synapses for enduring motor memories. Nature 2009, 462, 915–919. [Google Scholar] [CrossRef]
- Kleim, J.A.; Barbay, S.; Cooper, N.R.; Hogg, T.M.; Reidel, C.N.; Remple, M.S.; Nudo, R.J. Motor Learning-Dependent Synaptogenesis Is Localized to Functionally Reorganized Motor Cortex. Neurobiol. Learn. Mem. 2002, 77, 63–77. [Google Scholar] [CrossRef]
- Monfils, M.-H.; Teskey, G. Skilled-learning-induced potentiation in rat sensorimotor cortex: A transient form of behavioural long-term potentiation. Neuroscience 2004, 125, 329–336. [Google Scholar] [CrossRef]
- Hess, G.; Donoghue, J.P.; Sato, D.; Yamashiro, K.; Onishi, H.; Yasuhiro, B.; Shimoyama, Y.; Maruyama, A.; Mang, C.S.; Snow, N.J.; et al. Long-term potentiation of horizontal connections provides a mechanism to reorganize cortical motor maps. J. Neurophysiol. 1994, 71, 2543–2547. [Google Scholar] [CrossRef] [PubMed]
- Rioult-Pedotti, M.-S.; Friedman, D.; Donoghue, J.P. Learning-Induced LTP in Neocortex. Science 2000, 290, 533–536. [Google Scholar] [CrossRef]
- Strens, L.H.; Fogelson, N.; Shanahan, P.; Rothwell, J.C.; Brown, P. The Ipsilateral Human Motor Cortex Can Functionally Compensate for Acute Contralateral Motor Cortex Dysfunction. Curr. Biol. 2003, 13, 1201–1205. [Google Scholar] [CrossRef]
- Nishimura, Y.; Onoe, H.; Morichika, Y.; Perfiliev, S.; Tsukada, H.; Isa, T. Time-Dependent Central Compensatory Mechanisms of Finger Dexterity After Spinal Cord Injury. Science 2007, 318, 1150–1155. [Google Scholar] [CrossRef]
- Zörner, B.; Bachmann, L.C.; Filli, L.; Kapitza, S.; Gullo, M.; Bolliger, M.; Starkey, M.L.; Röthlisberger, M.; Gonzenbach, R.R.; Schwab, M.E. Chasing central nervous system plasticity: The brainstem’s contribution to locomotor recovery in rats with spinal cord injury. Brain 2014, 137, 1716–1732. [Google Scholar] [CrossRef]
- Matsuyama, K.; Mori, F.; Nakajima, K.; Drew, T.; Aoki, M.; Mori, S. Locomotor role of the corticoreticular-reticulospinal-spinal in-terneuronal system. Prog. Brain Res. 2004, 143, 239–249. [Google Scholar] [CrossRef]
- Dimitrijevic, M.R.; Gerasimenko, Y.; Pinter, M.M. Evidence for a Spinal Central Pattern Generator in Humans. Ann. N. Y. Acad. Sci. 1998, 860, 360–376. [Google Scholar] [CrossRef]
- Nadeau, S.; Jacquemin, G.; Fournier, C.; Lamarre, Y.; Rossignol, S. Spontaneous Motor Rhythms of the Back and Legs in a Patient With a Complete Spinal Cord Transection. Neurorehabilit. Neural Repair 2009, 24, 377–383. [Google Scholar] [CrossRef]
- Martinez, M.; Brown, A. From cortex to cord: Motor circuit plasticity after spinal cord injury. Neural Regen. Res. 2019, 14, 2054–2062. [Google Scholar] [CrossRef] [PubMed]
- Fluri, F.; Malzahn, U.; Homola, G.A.; Schuhmann, M.K.; Kleinschnitz, C.; Volkmann, J. Stimulation of the mesencephalic locomotor region for gait recovery after stroke. Ann. Neurol. 2017, 82, 828–840. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, L.C.; Matis, A.; Lindau, N.T.; Felder, P.; Gullo, M.; Schwab, M.E. Deep Brain Stimulation of the Midbrain Locomotor Region Improves Paretic Hindlimb Function After Spinal Cord Injury in Rats. Sci. Transl. Med. 2013, 5, 208ra146. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, C.S.; Fehlings, M. Concise Review: Bridging the Gap: Novel Neuroregenerative and Neuroprotective Strategies in Spinal Cord Injury. Stem Cells Transl. Med. 2016, 5, 914–924. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Okano, H. Cell transplantation therapies for spinal cord injury focusing on induced pluripotent stem cells. Cell Res. 2013, 23, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Massoto, T.B.; Santos, A.C.R.; Ramalho, B.S.; Almeida, F.M.; Martinez, A.M.B.; Marques, S.A. Mesenchymal stem cells and treadmill training enhance function and promote tissue preservation after spinal cord injury. Brain Res. 2019, 1726, 146494. [Google Scholar] [CrossRef]
- Younsi, A.; Zheng, G.; Scherer, M.; Riemann, L.; Zhang, H.; Tail, M.; Hatami, M.; Skutella, T.; Unterberg, A.; Zweckberger, K. Treadmill training improves survival and differentiation of transplanted neural precursor cells after cervical spinal cord injury. Stem Cell Res. 2020, 45, 101812. [Google Scholar] [CrossRef]
- Hwang, D.H.; Shin, H.Y.; Kwon, M.J.; Choi, J.Y.; Ryu, B.-Y.; Kim, B.G. Survival of Neural Stem Cell Grafts in the Lesioned Spinal Cord Is Enhanced by a Combination of Treadmill Locomotor Training via Insulin-Like Growth Factor-1 Signaling. J. Neurosci. 2014, 34, 12788–12800. [Google Scholar] [CrossRef]
- Tashiro, S.; Nishimura, S.; Iwai, H.; Sugai, K.; Zhang, L.; Shinozaki, M.; Iwanami, A.; Toyama, Y.; Liu, M.; Okano, H.; et al. Functional Recovery from Neural Stem/Progenitor Cell Transplantation Combined with Treadmill Training in Mice with Chronic Spinal Cord Injury. Sci. Rep. 2016, 6, 30898. [Google Scholar] [CrossRef]
- Sachdeva, R.; Theisen, C.C.; Ninan, V.; Twiss, J.L.; Houlé, J.D. Exercise dependent increase in axon regeneration into peripheral nerve grafts by propriospinal but not sensory neurons after spinal cord injury is associated with modulation of regeneration-associated genes. Exp. Neurol. 2015, 276, 72–82. [Google Scholar] [CrossRef]
- Theisen, C.C.; Sachdeva, R.; Austin, S.; Kulich, D.; Kranz, V.; Houle, J.D. Exercise and Peripheral Nerve Grafts as a Strategy To Promote Regeneration after Acute or Chronic Spinal Cord Injury. J. Neurotrauma 2017, 34, 1909–1914. [Google Scholar] [CrossRef]
- Takeoka, A.; Jindrich, D.L.; Muñoz-Quiles, C.; Zhong, H.; Brand, R.v.D.; Pham, D.L.; Ziegler, M.D.; Ramón-Cueto, A.; Roy, R.R.; Edgerton, V.R.; et al. Axon Regeneration Can Facilitate or Suppress Hindlimb Function after Olfactory Ensheathing Glia Transplantation. J. Neurosci. 2011, 31, 4298–4310. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Ye, C.; Zhang, Z.; Wu, J.; Huang, H. Cotransplantation of Olfactory Ensheathing Cells and Schwann Cells Combined with Treadmill Training Promotes Functional Recovery in Rats with Contused Spinal Cords. Cell Transplant. 2013, 22, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Fan, X.; Wang, J.-Z.; Lin, X.-M. Stem cell transplantation for spinal cord injury: A meta-analysis of treatment effectiveness and safety. Neural Regen. Res. 2017, 12, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, O.; Sugai, K.; Yamaguchi, R.; Tashiro, S.; Nagoshi, N.; Kohyama, J.; Iida, T.; Ohkubo, T.; Itakura, G.; Isoda, M.; et al. Concise Review: Laying the Groundwork for a First-In-Human Study of an Induced Pluripotent Stem Cell-Based Intervention for Spinal Cord Injury. Stem Cells 2018, 37, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.S.; Tang, C.; Rao, M.S.; Weissman, I.L.; Wu, J.C. Tumorigenicity as a clinical hurdle for pluripotent stem cell therapies. Nat. Med. 2013, 19, 998–1004. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Evoked Potentials and Tests | Description |
|---|---|
| Motor Evoked Potential (MEP) | MEPs are signals originating from descending motor pathways or muscles, which are recorded after the stimulation of motor paths in the brain. MEPs triggered by transcranial magnetic stimulation of the human motor cortex provide information about corticospinal excitability during stimulation [88]. |
| Somatosensory Evoked Potential (SEP) | SEPs are electrical signals that provide somatosensory information and are transmitted through two major pathways within the spinal cord: the spinal lemniscus system and the spinothalamic system. SEP monitors only the dorsal lemniscal system, which transmits mechanoreception and proprioception. The recording of SEPs typically begins at the upper or lower extremity nerves, which are then picked up by the surgeon upon completion of surgery. Different parts of the sensory pathway receive electrical energy from electrodes. Sensory pathways are characterized by the transmission of evoked potentials from electrodes to the cortex, where waveforms are recorded [89]. |
| Contact Heat Evoked Potential (CHEP) | To induce a brain EEG response, this neuroelectrophysiological technique involves applying neo-conscious tissue onto skin surfaces through rapid temperature changes (70 °C/s) via a fiber transmission. By examining the cortical responses, which include N2 latency, P2 latency, and N2-P2 amplitude, it is possible to determine the function of peripheral small nerve fibers [90,91]. |
| Quantitative Sensory Testing (QST) | The use of this testing method involves a systematic approach to psychophysical testing that measures sensory thresholds such as pain, touch, vibration, and temperature. It measures personal sensation based on direct feedback from patients. It tests sensory loss (hypoesthesia, hypoesthesia) sensory gain (hyperalgesia, hyperalgesia, allodynia) nociception, and nociception of different afferent nerve fibers and central pathways [92,93,94]. |
| Laser Evoked Potential (LEP) | LEP is a result of the brain’s response to laser-radiated heat pulses, which also trigger Aδ nociceptors. It is the most commonly used approach to investigate the function of nociceptive pathways in NP patients. Aδ and C nociceptors are activated by laser-generated radiant heat pulses, which generate “late” brain potential that is dependent on the activation of adjacent Aδ fibers [95]. |
| Author | Aim | Treatment Period | Sample Size | Outcomes Measures | Main Findings | Study Limitations |
|---|---|---|---|---|---|---|
| Versace et al., 2017 [105] | To assess whether SCI patients show altered sensory-motor plasticity within the M1. | Not Specificated. | 10 Subjects with chronic SCIs and 10 Healthy Volunteers. | PAS, TMS. SCIM. | The PAS protocol significantly increased corticospinal excitability within 30 min in healthy subjects and SCI patients with good motor recovery but not in SCI patients with poor functional recovery. | PAS may not be beneficial in SCI patients with poor recovery due to difficulty in finding the right hotspot and potential differences in conductivity between patients and healthy controls. |
| Jo et al., 2023 [106] | The study aimed to enhance corticospinal-motor neuronal synapses at multiple spinal levels through Hebbian plasticity, thereby promoting functional recovery in the legs and arms. | 8 to14 weeks. | 20 Participants with chronic SCIs. | ISNCSCI. | Participants with Hebbian stimulation showed improved walking speed, corticospinal function, grip, gait, and quality of life compared to sham stimulation, with further improvements observed after nine months. | Small sample to generalize the data. |
| Khan et al., 2016 [107] | To determine the neural plasticity of spinal reflexes after two contrasting forms of walking training in individuals with chronic, motor-incomplete SCI. | 6 months | 20 Participants. | EMG, Electodes. | Reflex excitability, a specific response to training, was found to improve walking function, with participants with lower reflex excitability showing higher walking speed and distance. | Participants’ biases may have influenced the results, but they likely did not prefer one type of training over another, as they were unaware of the best exercise intervention. |
| Gonzalez et al., 2016 [108] | This study aimed to assess the rehabilitation effects and changes in white matter microstructure in patients with high SCI after bilateral upper extremity motor skill training. | Participants performed the visuospatial-motor training task in 12 sessions of 1.5 h: 2–3 times a week for a total of 4–6 weeks. | 5 Subjects and 14 Control Subjects. | MMT | Exercise training enhances shoulder and upper arm MMT scores, isometric muscle strength, and FA values of the left hemisphere cingulate, indicating local white matter microstructural changes. | No additional clinical assessment was performed in the study. Motor dysfunction, muscle weakness, muscle atrophy, and cortical atrophy were shown to progress further with immobilization following SCIs. |
| Jo et al., 2020 [109] | The aim was to enhance functional recovery by working on the remaining neural networks. | 10 sessions in 2–3 weeks. | 25 Individuals with SCIs. | GRASSP | The GRASSP and 10 m walking tests were reduced by 20% in all protocols, but corticospinal responses and muscle contraction amplitude increased by 40–50% after PCMS with or without exercise. | The effectiveness of neuromodulatory approaches in enhancing exercise effects is not fully understood due to limited studies combining exercise with sham neurostimulation and stimulus intensity. |
| Faw et al., 2021 [110] | To examine whether the system promotes white matter plasticity and recovery in chronic incomplete SCIs. | 12 weeks (3 times/week). | 20 Individuals with SCIs. | MRI | This study indicates that eccentricity-focused downhill rehabilitation enhances white matter plasticity and functional recovery in chronic SCIs through oligodendrogenesis in neuronal regions activated by the training approach. | Many people with SCIs were excluded from human trials for the following reasons: spinal hardware, claustrophobia, and motion artifacts. |
| Castro et al., 2013 [111] | Look for detectable changes in neuroplasticity immediately after trauma. | Non Specificated. | 20 Patients. | Stimuli Tasks, EEG. | The study found that the SCI group had smaller preparation and movement potential amplitudes and a more similar topographic distribution of movement potentials compared to the exercise control group. | Not detected. |
| Jutzeler et al., 2015 [112] | The study aimed to investigate the relationship between cortical reorganization and NP after SCI. | Not Specificated. | 57 Subjects (26 with NP and 31 Healthy Individuals). | FMR, Sensory Tasks, EI. | The results suggest that NP is not associated with increased plasticity in motor and sensory tasks above the lesion level. | The study focused on the base of the thumb using various sensory modalities, lacking reorganization responses and addressing heterogeneity in the SCI sample compared to Wrigley. |
| Villiger et al., 2015 [113] | This study used longitudinal MRI to assess structural brain plasticity induced by improved training in patients with chronic inflammatory SCI. | Between August 2010 and March 2012. | 9 Patients with iSCI. | MRI, TBM. | TBM volume increases in various brain regions, particularly in patients with iSCIs, with significant improvements observed in the left middle temporal and occipital gyrus, hippocampus, cerebellum, corpus callosum, and brainstem. | Small sample size. The lack of an SCI control group for training with virtual reality means that the changes induced by the training in patients could have occurred not only as part of the training but also as part of the placebo effect. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calderone, A.; Cardile, D.; De Luca, R.; Quartarone, A.; Corallo, F.; Calabrò, R.S. Brain Plasticity in Patients with Spinal Cord Injuries: A Systematic Review. Int. J. Mol. Sci. 2024, 25, 2224. https://doi.org/10.3390/ijms25042224
Calderone A, Cardile D, De Luca R, Quartarone A, Corallo F, Calabrò RS. Brain Plasticity in Patients with Spinal Cord Injuries: A Systematic Review. International Journal of Molecular Sciences. 2024; 25(4):2224. https://doi.org/10.3390/ijms25042224
Chicago/Turabian StyleCalderone, Andrea, Davide Cardile, Rosaria De Luca, Angelo Quartarone, Francesco Corallo, and Rocco Salvatore Calabrò. 2024. "Brain Plasticity in Patients with Spinal Cord Injuries: A Systematic Review" International Journal of Molecular Sciences 25, no. 4: 2224. https://doi.org/10.3390/ijms25042224
APA StyleCalderone, A., Cardile, D., De Luca, R., Quartarone, A., Corallo, F., & Calabrò, R. S. (2024). Brain Plasticity in Patients with Spinal Cord Injuries: A Systematic Review. International Journal of Molecular Sciences, 25(4), 2224. https://doi.org/10.3390/ijms25042224