
Analysis of Functional Single-Nucleotide Polymorphisms (SNPs) and Leaf Quality in Tea Collection under Nitrogen-Deficient Conditions

 , ,
, , 
 and
and 
Abstract
:1. Introduction
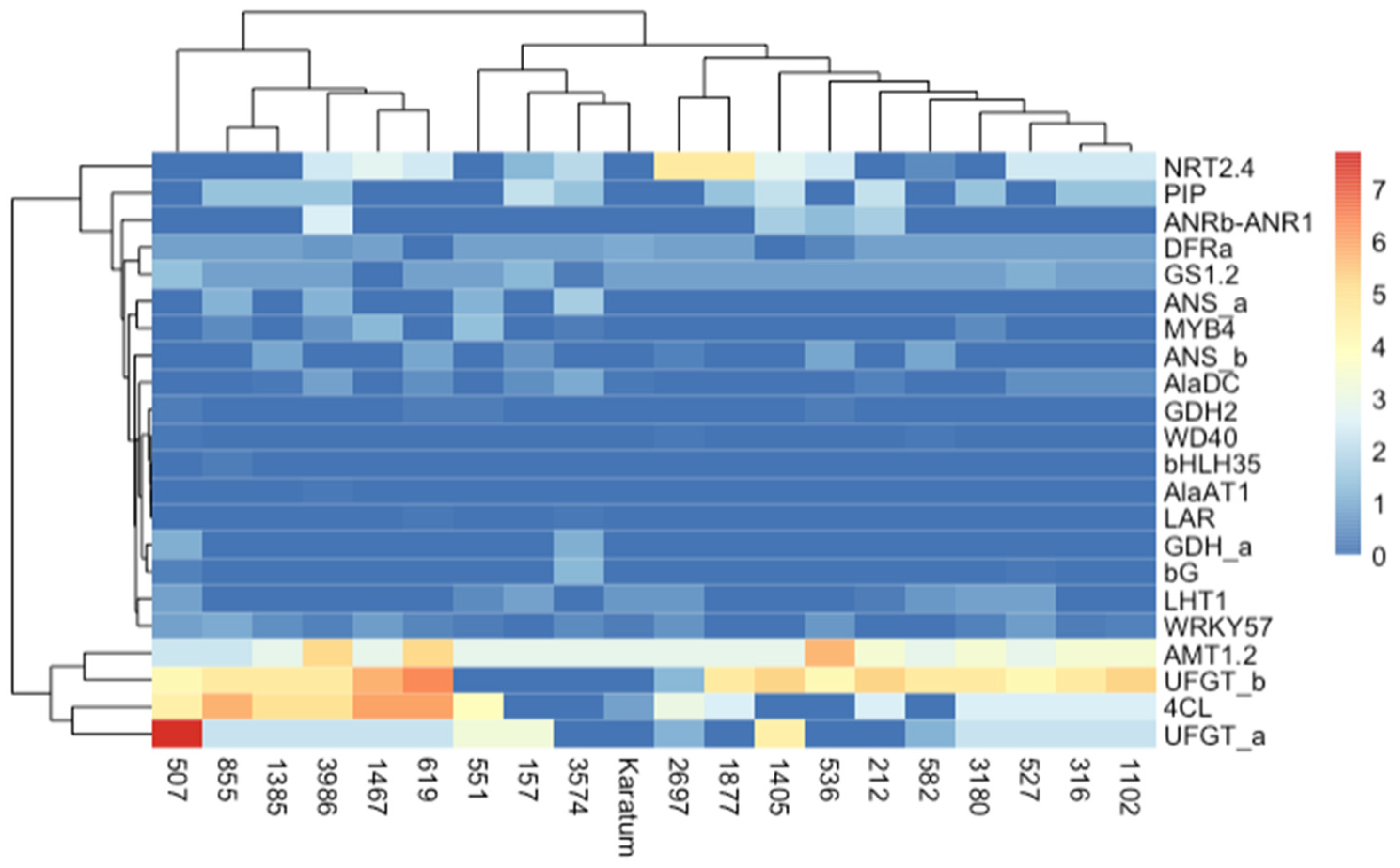
2. Results
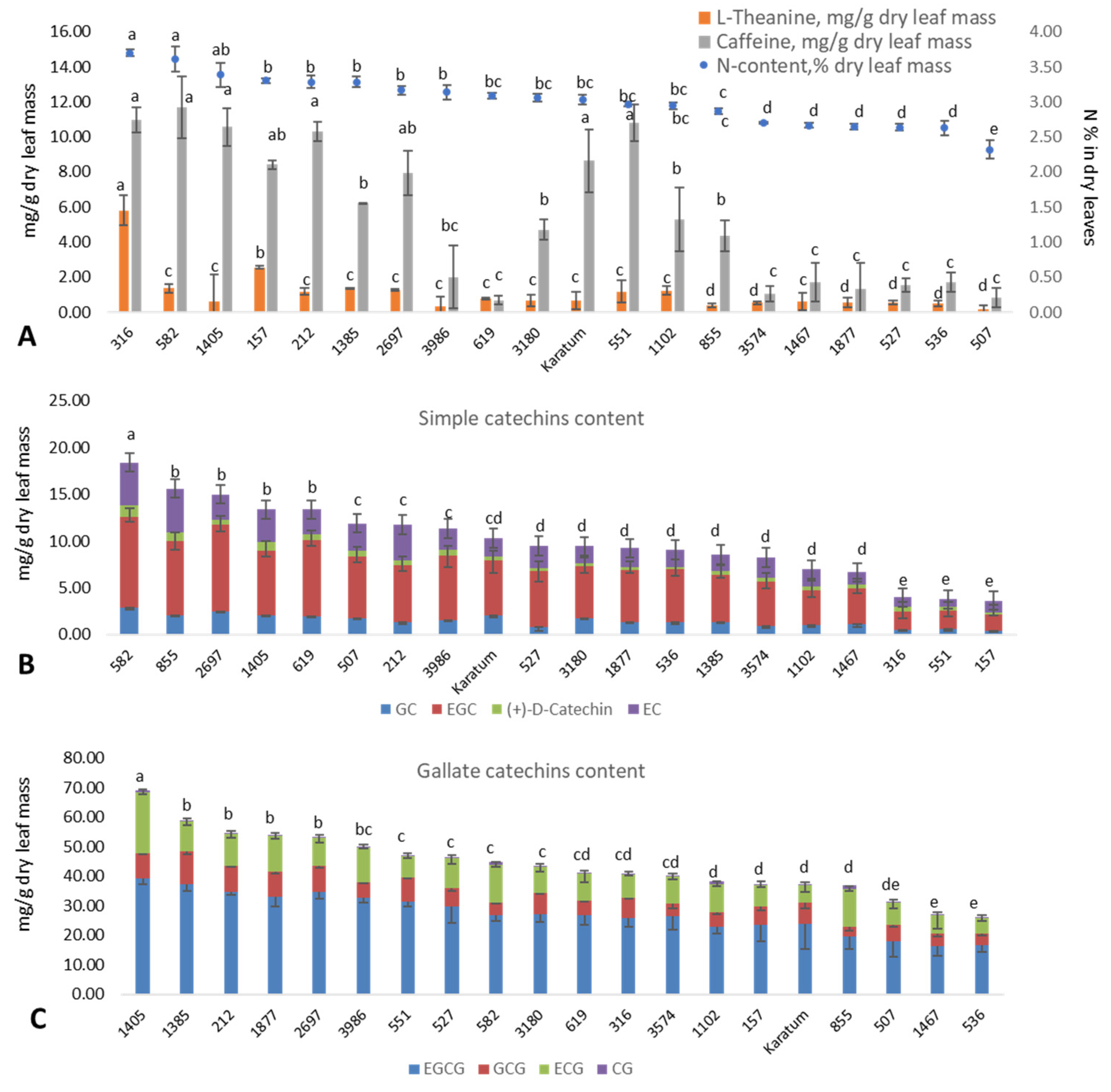
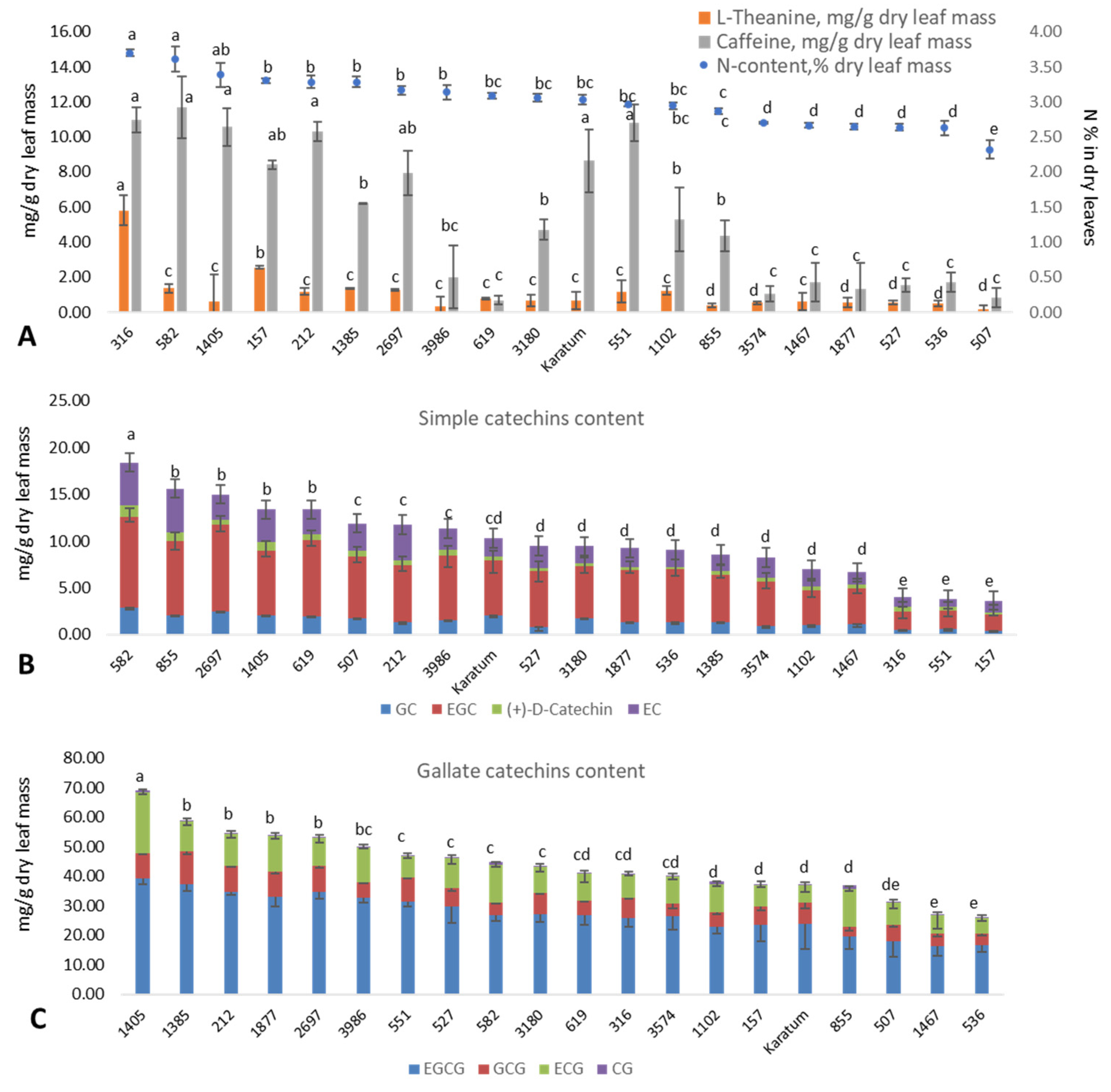
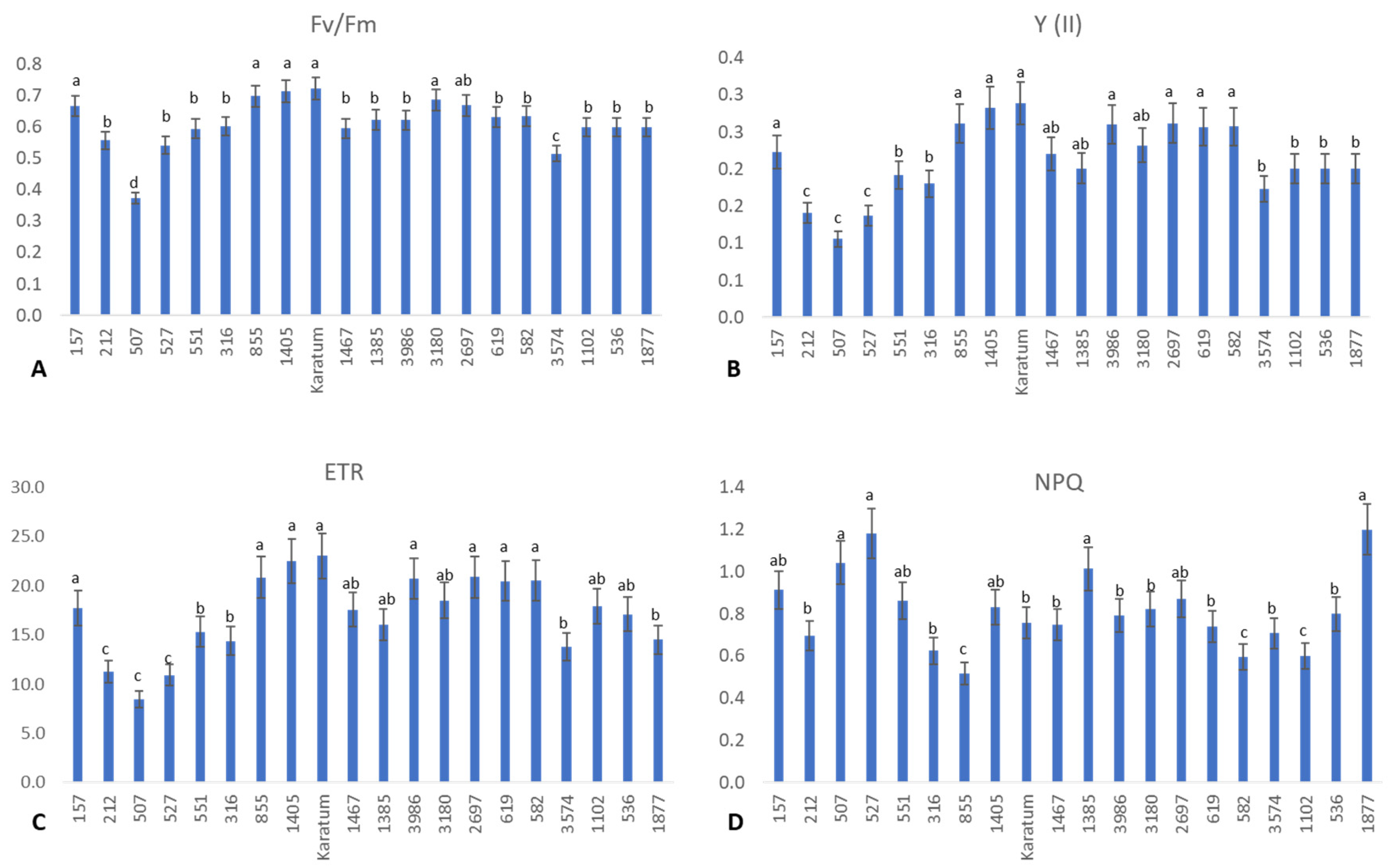
2.1. Phenotypic Evaluation of the Collection
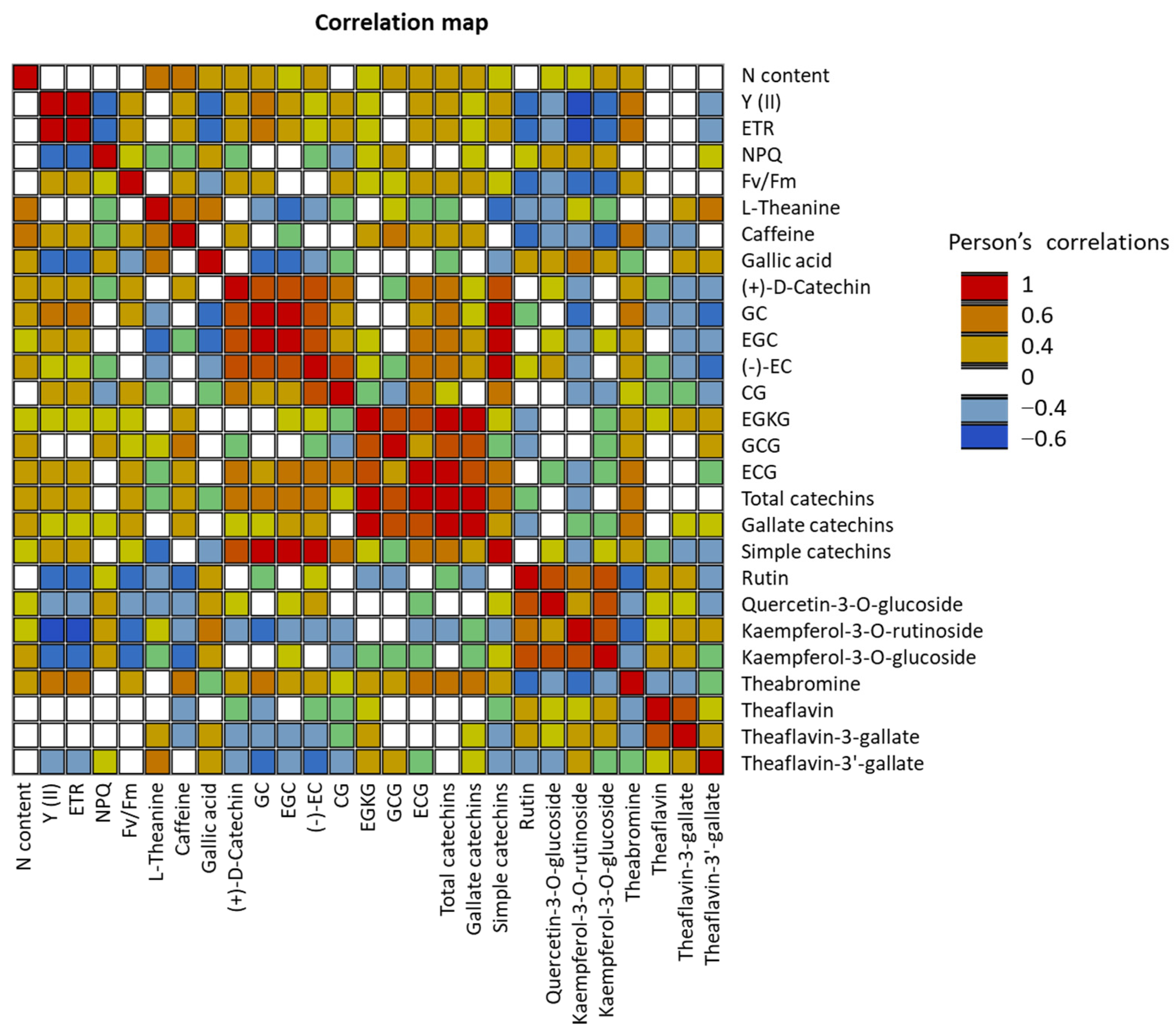
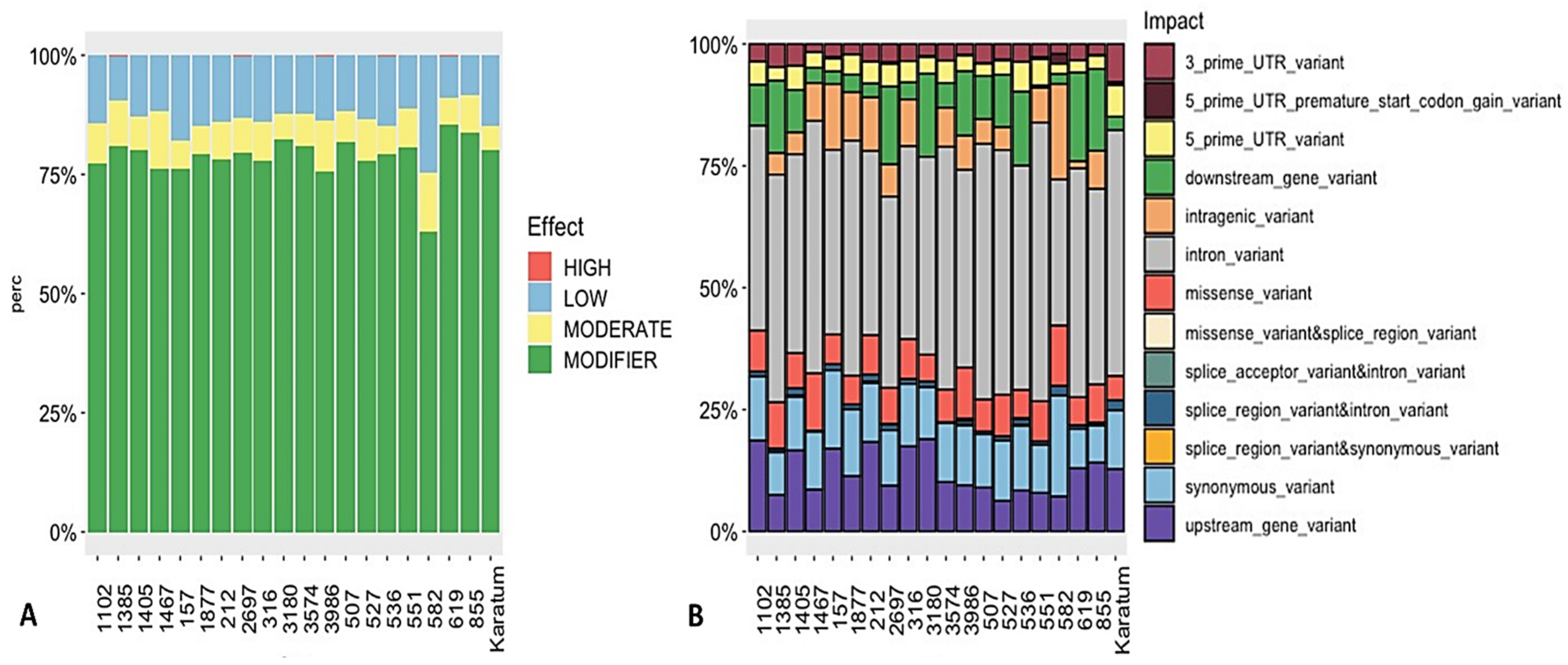
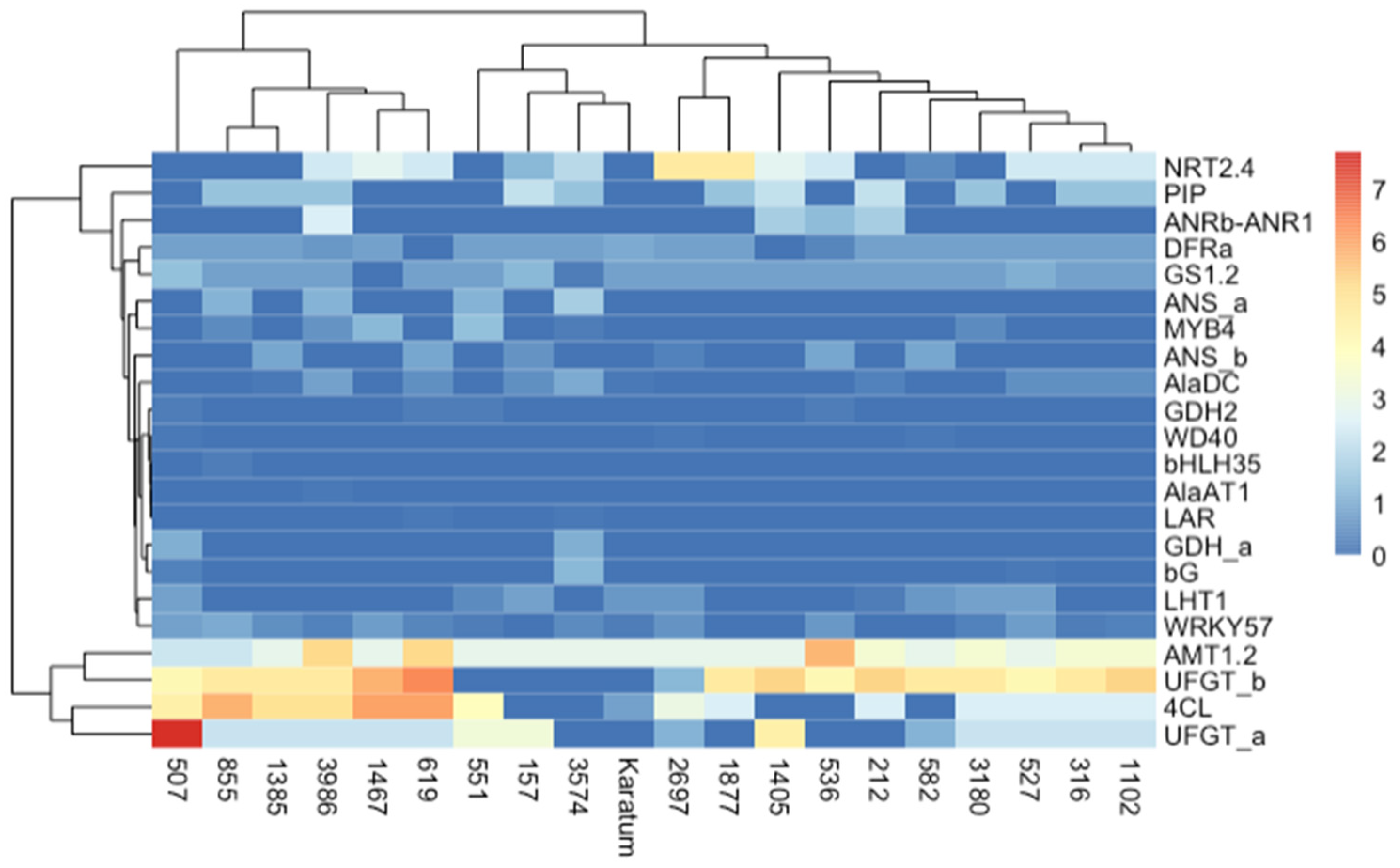
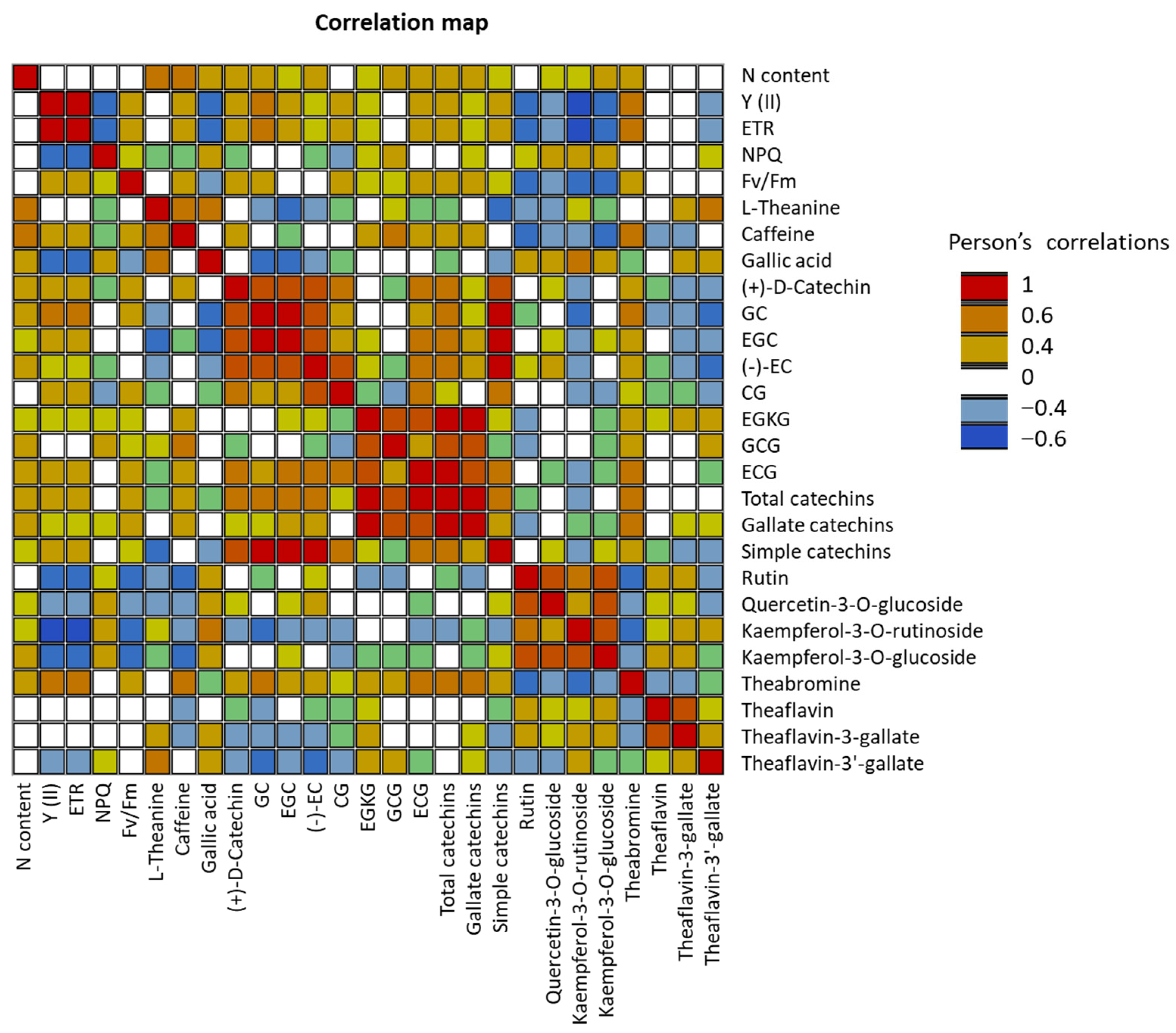
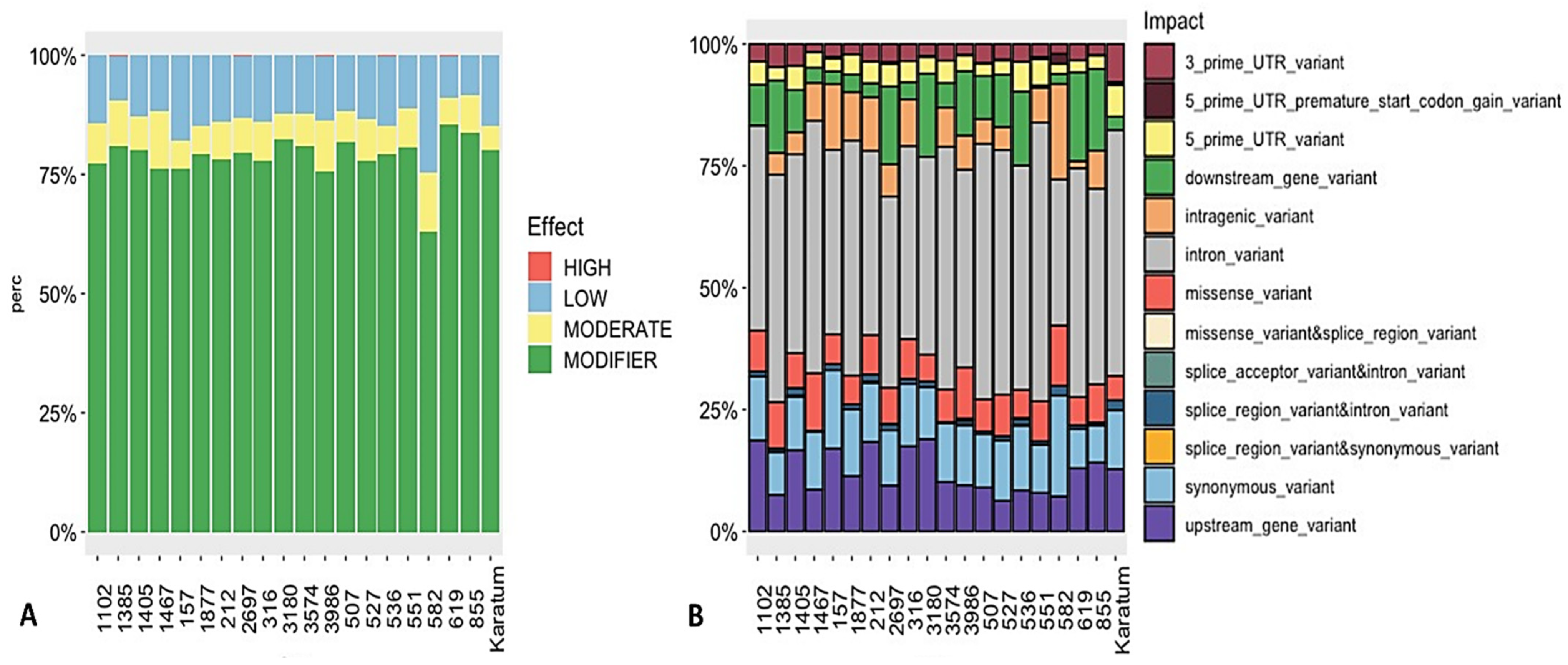
2.2. Detection of SNPs in the Selected Genes and their Relationships with Phenotypes
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Phenotypic Evaluation
4.2. Gene Selection and Primer Design and Long-Range Polymerase Chain Reaction (LR-PCR)
4.3. Pooled Amplicon Sequencing, Filtering and Variation Calling
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sedaghathoor, S.; Torkashvand, A.M.; Hashemabadi, D.; Kaviani, B. Yield and quality response of tea plant to fertilizers. Afr. J. Agric. Res. 2009, 4, 568–570. [Google Scholar]
- Cao, Q.; Cao, Q.; Yang, G.; Duan, D.; Chen, L.; Wang, F.; Xu, B.; Zhao, C.; Niu, F. Combining multispectral and hyperspectral data to estimate nitrogen status of tea plants (Camellia sinensis (L.) O. Kuntze) under field conditions. Comput. Electron. Agric. 2022, 198, 107084. [Google Scholar] [CrossRef]
- Zavalin, A.A.; Sokolov, O.A. Nitrogen Fluxes in the Agroecosystem: From the Ideas of D.N. Pryanishnikov to the Present Day; VNIIA: Moscow, Russia, 2016. [Google Scholar]
- Li, W.; Zhong, M.; Zhou, L.; Liu, H.; Li, S.; Wang, X. Transcriptome and metabolite analysis identifies nitrogen utilization genes in tea plant (Camellia sinensis). Sci. Rep. 2017, 7, 1693. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Yao, Q.; Xia, E.; Gao, L. Metabolomics and Transcriptomics Analyses Reveal Nitrogen Influences on the Accumulation of Flavonoids and Amino Acids in Young Shoots of Tea Plant (Camellia sinensis L.) Associated with Tea Flavor. J. Agric. Food Chem. 2018, 66, 9828–9838. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, H.; Pilon-Smits, E.; Huang, W.; Wang, P.; Wang, M.; Guo, F.; Wang, Y.; Li, R.; Zhao, H.; et al. Transcriptome-wide analysis of nitrogen-regulated genes in tea plant (Camellia sinensis LO Kuntze) and characterization of amino acid transporter CsCAT9. Plants 2020, 9, 1218. [Google Scholar] [CrossRef] [PubMed]
- Cochetel, N.; Escudié, F.; Cookson, S.J.; Dai, Z.; Vivin, P.; Bert, P.-F.; Muñoz, M.S.; Delrot, S.; Klopp, C.; Ollat, N.; et al. Root transcriptomic responses of grafted grapevines to heterogeneous nitrogen availability depend on rootstock genotype. J. Exp. Bot. 2017, 68, 4339–4355. [Google Scholar] [CrossRef] [PubMed]
- Gojon, A.; Krouk, G.; Perrine-Walker, F.; Laugier, E. Nitrate transceptor(s) in plants. J. Exp. Bot. 2011, 62, 2299–2308. [Google Scholar] [CrossRef]
- Medici, A.; Krouk, G. The primary nitrate response: A multifaceted signalling pathway. J. Exp. Bot. 2014, 65, 5567–5576. [Google Scholar] [CrossRef]
- O’Brien, J.A.; Vega, A.; Bouguyon, E.; Krouk, G.; Gojon, A.; Coruzzi, G.; Gutiérrez, R.A. Nitrate transport, sensing, and responses in plants. Mol. Plant 2016, 9, 837–856. [Google Scholar] [CrossRef]
- Ruffela, S.; Krouka, G.; Ristovaa, D.; Shashac, D.; Birnbauma, K.D.; Coruzzi, G.M. Nitrogen economics of root foraging: Transitive closure of the nitrate–cytokinin relay and distinct systemic signaling for N supply vs. demand. Proc. Natl. Acad. Sci. USA 2011, 108, 18524–18529. [Google Scholar] [CrossRef]
- Alvarez, J.M.; Vidal, E.A.; Gutiérrez, R.A. Integration of local and systemic signaling pathways for plant N responses. Curr. Opin. Plant Biol. 2012, 15, 185–191. [Google Scholar] [CrossRef]
- Bouguyon, E.; Gojon, A.; Nacry, P. Nitrate sensing and signaling in plants. In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2012; Volume 23, pp. 648–654. [Google Scholar] [CrossRef]
- Nacry, P.; Bouguyon, E.; Gojon, A. Nitrogen acquisition by roots: Physiological and developmental mechanisms ensuring plant adaptation to a fluctuating resource. Plant Soil 2013, 370, 1–29. [Google Scholar] [CrossRef]
- Krapp, A.; David, L.C.; Chardin, C.; Girin, T.; Marmagne, A.; Leprince, A.-S.; Chaillou, S.; Ferrario-Méry, S.; Meyer, C.; Daniel-Vedele, F. Nitrate transport and signalling in Arabidopsis. J. Exp. Bot. 2014, 65, 789–798. [Google Scholar] [CrossRef]
- Krapp, A. Plant nitrogen assimilation and its regulation: A complex puzzle with missing pieces. Curr. Opin. Plant Biol. 2015, 25, 115–122. [Google Scholar] [CrossRef]
- Li, Y.; Krouk, G.; Coruzzi, G.M.; Ruffel, S. Finding a nitrogen niche: A systems integration of local and systemic nitrogen signaling in plants. J. Exp. Bot. 2014, 65, 5601–5610. [Google Scholar] [CrossRef]
- Gan, Y.; Bernreiter, A.; Filleur, S.; Abram, B.; Forde, B.G. Overexpressing the ANR1 MADS-box gene in transgenic plants provides new insights into its role in the nitrate regulation of root development. Plant Cell Physiol. 2012, 53, 1003–1016. [Google Scholar] [CrossRef]
- Mounier, E.; Pervent, M.; Ljung, K.; Gojon, A.; Nacry, P. Auxin-mediated nitrate signaling by NRT1.1 participates in the adaptive response of Arabidopsis root architecture to the spatial heterogeneity of nitrate availability. Plant Cell Environ. 2014, 37, 162–174. [Google Scholar] [CrossRef]
- Kiba, T.; Feria-Bourrellier, A.-B.; Lafouge, F.; Lezhneva, L.; Boutet-Mercey, S.; Orsel, M.; Bréhaut, V.; Miller, A.; Daniel-Vedele, F.; Sakakibara, H.; et al. The Arabidopsis nitrate transporter NRT2.4 plays a double role in roots and shoots of nitrogen-starved plants. Plant Cell 2012, 24, 245–258. [Google Scholar] [CrossRef]
- Ruffel, S.; Gojon, A.; Lejay, L. Signal interactions in the regulation of root nitrate uptake. J. Exp. Bot. 2014, 65, 5509–5517. [Google Scholar] [CrossRef]
- Tabata, R.; Sumida, K.; Yoshii, T.; Ohyama, K.; Shinohara, H.; Matsubayashi, Y. Perception of root-derived peptides by shoot LRR-RKs mediates systemic N-demand signaling. Science 2014, 346, 343–346. [Google Scholar] [CrossRef]
- Krouk, G. Hormones and nitrate: A two-way connection. Plant Mol. Biol. 2016, 91, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Ohkubo, Y.; Tanaka, M.; Tabata, R.; Ogawa-Ohnishi, M.; Matsubayashi, Y. Shoot-to-root mobile polypeptides involved in systemic regulation of nitrogen acquisition. Nat. Plants 2017, 3, 17029. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Apostolides, Z.; Chen, Z.M. Global Tea Breeding; Chen, L., Apostolides, Z., Chen, Z.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; Volume 378, p. 588. [Google Scholar]
- Jayasinghe, S.L.; Kumar, L. Potential impact of the current and future climate on the yield, quality, and climate suitability for tea [Camellia sinensis (L.) O. Kuntze]: A systematic review. Agronomy 2021, 11, 619. [Google Scholar] [CrossRef]
- Pan, S.Y.; Nie, Q.; Tai, H.C.; Song, X.L.; Tong, Y.F.; Zhang, L.J.; Wu, X.W.; Lin, Z.H.; Zhang, Y.Y.; Ye, D.Y.; et al. Tea and tea drinking: China’s outstanding contributions to the mankind. Chin. Med. 2022, 17, 27. [Google Scholar] [CrossRef]
- Available online: www.fao.org/faostat/ (accessed on 30 May 2023).
- Mamati, G.E.; Liang, Y.; Lu, J. Expression of basic genes involved in tea polyphenol synthesis in relation to accumulation of catechins and total tea polyphenols. J. Sci. Food Agric. 2006, 86, 459–464. [Google Scholar] [CrossRef]
- Mahmood, T.; Akhtar, N.; Khan, B.A. The morphology, characteristics and medicinal properties of Camellia sinensis tea. J. Med. Plants Res. 2010, 4, 2028–2033. [Google Scholar] [CrossRef]
- Donlao, N.; Ogawa, Y. The influence of processing conditions on catechin, caffeine and chlorophyll contents of green tea (Camelia sinensis) leaves and infusions. LWT 2019, 116, 108567. [Google Scholar] [CrossRef]
- Zhu, J.Y.; Xu, Q.; Zhao, S.; Xia, X.; Yan, X.; An, Y.; Mi, X.; Guo, L.; Samarina, L.; Wei, C. Comprehensive co-expression analysis provides novel insights into temporal variation of flavonoids in fresh leaves of the tea plant (Camellia sinensis). Plant Sci. 2020, 290, 110306. [Google Scholar] [CrossRef]
- Li, F.; Dong, C.; Yang, T.; Bao, S.; Fang, W.; Lucas, W.J.; Zhang, Z. The tea plant CsLHT1 and CsLHT6 transporters take up amino acids, as a nitrogen source, from the soil of organic tea plantations. Hortic. Res. 2021, 8, 178. [Google Scholar] [CrossRef]
- Samynathan, R.; Thiruvengadam, M.; Nile, S.H.; Shariati, M.A.; Rebezov, M.; Mishra, R.K.; Venkidasamy, B.; Periyasamy, S.; Chung, I.M.; Pateiro, M.; et al. Recent insights on tea metabolites, their biosynthesis and chemo-preventing effects: A review. Crit. Rev. Food Sci. Nutr. 2023, 63, 3130–3149. [Google Scholar] [CrossRef]
- Fang, K.X.; Xia, Z.; Li, H.; Jiang, X.; Qin, D.; Wang, Q.; Wang, Q.; Pan, C.; Li, B.; Wu, H. Genome-wide association analysis identified molecular markers associated with important tea flavor-related metabolites. Hortic. Res. 2021, 8, 42. [Google Scholar] [CrossRef] [PubMed]
- Hollman, P.C.; Feskens, E.J.; Katan, M.B. Tea flavonols in cardiovascular disease and cancer epidemiology. Proc. Soc. Exp. Biol. Med. 1999, 220, 198–202. [Google Scholar] [CrossRef]
- Auvichayapat, P.; Prapochanung, M.; Tunkamnerdthai, O.; Sripanidkulchai, B.O.; Auvichayapat, N.; Thinkhamrop, B.; Kunhasura, S.; Wongpratoom, S.; Sinawat, S.; Hongprapas, P. Effectiveness of green tea on weight reduction in obese Thais: A randomized, controlled trial. Physiol. Behav. 2008, 93, 486–491. [Google Scholar] [CrossRef] [PubMed]
- Kurita, I.; Maeda-Yamamoto, M.; Tachibana, H.; Kamei, M. Antihypertensive effect of Benifuuki tea containing O-methylated EGCG. J. Agric. Food Chem. 2010, 58, 1903–1908. [Google Scholar] [CrossRef] [PubMed]
- Sasazuki, S.; Tamakoshi, A.; Matsuo, K.; Ito, H.; Wakai, K.; Nagata, C.; Mizoue, T.; Tanaka, K.; Tsuji, I.; Inoue, M.; et al. Green tea consumption and gastric cancer risk: An evaluation based on a systematic review of epidemiologic evidence among the Japanese population. JpnJ Clin Oncol. 2012, 42, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, J.M.; Croft, K.D.; Woodman, R.J.; Puddey, I.B.; Fuchs, D.; Draijer, R.; Lukoshkova, E.; Head, G.A. Black tea lowers the rate of blood pressure variation: A randomized controlled trial. Am. J. Clin. Nutr. 2013, 97, 943–950. [Google Scholar] [CrossRef]
- Ng, K.W.; Cao, Z.J.; Chen, H.B.; Zhao, Z.Z.; Zhu, L.; Yi, T. Oolong tea: A critical review of processing methods, chemical composition, health effects, and risk. Crit. Rev. Food Sci. Nutr. 2018, 58, 2957–2980. [Google Scholar] [CrossRef]
- Kuban-Jankowska, A.; Kostrzewa, T.; Musial, C.; Barone, G.; Lo Bosco, G.; Lo Celso, F.; Gorska-Ponikowska, M. Green tea catechins induce inhibition of PTP1B phosphatase in breast cancer cells with potent anti-cancer properties: In vitro assay, molecular docking, and dynamics studies. Antioxidants 2020, 9, 1208. [Google Scholar] [CrossRef]
- Fujimura, Y.; Fujino, K.; Yoshimoto, T.; Nezu, A.; Marugame, Y.; Bae, J.; Kumazoe, M.; Tachibana, H. Eriodictyol-Amplified 67-kDa laminin receptor signaling potentiates the antiallergic effect of O-Methylated catechin. J. Nat. Prod. 2021, 84, 1823–1830. [Google Scholar] [CrossRef]
- Zhang, L.Q.; Wei, K.; Cheng, H.; Wang, L.-Y.; Zhang, C.-C. Accumulation of catechins and expression of catechin synthetic genes in Camellia sinensis at different developmental stages. Bot. Stud. 2016, 57, 31. [Google Scholar] [CrossRef]
- Fang, R.; Redfern, S.P.; Kirkup, D.; Porter, E.A.; Kite, G.C.; Terry, L.A.; Berry, M.J.; Simmonds, M.S. Variation of theanine, phenolic, and methylxanthine compounds in 21 cultivars of Camellia sinensis harvested in different seasons. Food Chem. 2017, 220, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.S.; Wei, K.; Wang, L.; Ruan, L.; Li, H.; Zhou, X.; Lin, Z.; Shan, R.; Cheng, H. Expression of key structural genes of the phenylpropanoid pathway associated with catechin epimerization in tea cultivars. Front. Plant Sci. 2017, 8, 702. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Yu, S.; Li, J.; Li, Q.; Wang, K.; Huang, J.; Liu, Z. Molecular characterization of WRKY transcription factors that act as negative regulators of O-methylated catechin biosynthesis in tea plants (Camellia sinensis L.). J. Agric. Food Chem. 2018, 66, 11234–11243. [Google Scholar] [CrossRef]
- Wang, W.Z.; Zhou, Y.; Wu, Y.; Dai, X.; Liu, Y.; Qian, Y.; Li, M.; Jiang, X.; Wang, Y.; Gao, L.; et al. Insight into catechins metabolic pathways of Camellia sinensis based on genome and transcriptome analysis. J. Agric. Food Chem. 2018, 66, 4281–4293. [Google Scholar] [CrossRef]
- Cui, X.; Zhao, P.; Liang, W.; Cheng, Q.; Mu, B.; Niu, F.; Yan, J.; Liu, C.; Xie, H.; Kav, N.N.V.; et al. A Rapeseed WRKY transcription factor phosphorylated by CPK modulates cell death and leaf senescence by regulating the expression of ROS and SA-Synthesis-Related genes. J. Agric. Food Chem. 2020, 68, 7348–7359. [Google Scholar] [CrossRef]
- Su, H.; Zhang, X.; He, Y.; Li, L.; Wang, Y.; Hong, G.; Xu, P. Transcriptomic analysis reveals the molecular adaptation of three major secondary metabolic pathways to multiple macronutrient starvation in tea (Camellia sinensis). Genes 2020, 11, 241. [Google Scholar] [CrossRef]
- Malyukova, L.S.; Samarina, L.S.; Zagoskina, N.V. Genetic mechanisms of the biosynthesis of catechins, caffeine and L-theanine in the tea plant Camellia sinensis (L.) Kuntze (review). Sel’skokhozyaistvennaya Biol. 2022, 57, 882–896. [Google Scholar] [CrossRef]
- Kong, W.; Jiang, M.; Wang, Y.; Chen, S.; Zhang, S.; Lei, W.; Chai, K.; Wang, P.; Liu, R.; Zhang, X. Pan-transcriptome assembly combined with multiple association analysis provides new insights into the regulatory network of specialized metabolites in the tea plant Camellia sinensis. Hortic. Res. 2022, 9, uhac100. [Google Scholar] [CrossRef]
- Chen, C.F.; Lin, J.Y. Estimating the gross budget of applied nitrogen and phosphorus in tea plantations. Sustain. Environ. Res. 2016, 26, 124–130. [Google Scholar] [CrossRef]
- Yan, P.; Shen, C.; Fan, L.; Li, X.; Zhang, L.; Zhang, L.; Han, W. Tea planting affects soil acidification and nitrogen and phosphorus distribution in soil. Agric Ecosyst. Environ. 2018, 254, 20–25. [Google Scholar] [CrossRef]
- Ruan, J.Y.; Ma, L.; Shi, Y.; Han, W. Uptake of fluoride by tea plant (Camellia sinensis L) and the impact of aluminium. J. Sci. Food Agric. 2003, 83, 1342–1348. [Google Scholar] [CrossRef]
- Yang, X.D.; Ma, L.; Ji, L.; Shi, Y.; Yi, X.; Yang, Q.; Ni, K.; Ruan, J. Long-term nitrogen fertilization indirectly affects soil fungi community structure by changing soil and pruned litter in a subtropical tea (Camellia sinensis L.) plantation in China. Plant Soil. 2019, 444, 409–426. [Google Scholar] [CrossRef]
- Fan, K.; Zhang, Q.; Tang, D.; Shi, Y.; Ma, L.; Liu, M.; Ruan, J. Dynamics of nitrogen translocation from mature leaves to new shoots and related gene expression during spring shoots development in tea plants (Camellia sinensis L.). J. Plant Nutr. Soil Sci. 2020, 183, 180–191. [Google Scholar] [CrossRef]
- Zhang, F.; He, W.; Yuan, Q.; Wei, K.; Ruan, L.; Wang, L.; Cheng, H. Transcriptome analysis identifies CsNRT genes involved in nitrogen uptake in tea plants, with a major role of CsNRT2. 4. Plant Physiol. Biochem. 2021, 167, 970–979. [Google Scholar] [CrossRef]
- Wang, Y.; Cheng, Y.; Yang, T.; Su, Y.; Lin, S.; Zhang, S.; Zhang, Z. Nitrogen-regulated theanine and flavonoid biosynthesis in tea plant roots: Protein-level regulation revealed by multiomics analyses. J. Agric. Food Chem. 2021, 69, 10002–10016. [Google Scholar] [CrossRef] [PubMed]
- Dinkeloo, K.; Boyd, S.; Pilot, G. Update on amino acid transporter functions and on possible amino acid sensing mechanisms in plants. Semin Cell Dev. Biol. 2018, 74, 105–113. [Google Scholar] [CrossRef]
- Fernández-Escobar, R.; Antonaya-Baena, F.; Almeida-Lavado, S. Nitrogen Uptake Efficiency of Olive Cultivars. Horticulturae 2021, 7, 136. [Google Scholar] [CrossRef]
- Kitajima, M.; Butler, W.L. Quenching of chlorophyll fluorescence and primary photochemistry in chloroplasts by dibromothymoquinone. Biochim. Biophys. Acta 1975, 376, 105–115. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim Biophys Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Brugnoli, E.; Björkman, O. Chloroplast movements in leaves: Influence on chlorophyll fluorescence and measurements of light-induced absorbance changes related to ΔpH and zeaxanthin formation. Photosynth. Res. 1992, 32, 23–35. [Google Scholar] [CrossRef]
- Ye, J.; Wang, Y.; Kang, J.; Chen, Y.; Hong, L.; Li, M.; Jia, Y.; Wang, Y.; Jia, X.; Wu, Z.; et al. Effects of Long-Term Use of Organic Fertilizer with Different Dosages on Soil Improvement, Nitrogen Transformation, Tea Yield and Quality in Acidified Tea Plantations. Plants 2022, 12, 122. [Google Scholar] [CrossRef]
- Wang, W.; Xin, H.; Wang, M.; Ma, Q.; Wang, L.; Kaleri, N.A.; Wang, Y.; Li, X. Transcriptomic Analysis Reveals the Molecular Mechanisms of Drought-Stress-Induced Decreases in Camellia sinensis Leaf Quality. Front Plant Sci. 2016, 7, 385. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Sun, Z.; Zhang, X.; Han, X. Novel Aspects of Regulation of Nitrogen Responses in the Tea Plant (Camellia sinensis (L.)). Agronomy 2023, 13, 144. [Google Scholar] [CrossRef]
- Li, M.; Guo, L.; Wang, Y.; Li, Y.; Jiang, X.; Liu, Y.; Xie, D.; Gao, L.; Xia, T. Molecular and biochemical characterization of two 4-coumarate: CoA ligase genes in tea plant (Camellia sinensis). Plant Mol. Biol. 2022, 109, 579–593. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.Y.; Gerendas, J.; Hardter, R.; Sattelmacher, B. Effect of nitrogen form and root-zone pH on growth and nitrogen uptake of tea (Camellia sinensis L.) plants. Ann. Bot. 2008, 99, 301–310. [Google Scholar] [CrossRef]
- Yang, Y.Y.; Li, X.H.; Ratcliffe, R.G.; Ruan, J.Y. Characterization of ammonium and nitrate uptake and assimilation in roots of tea plants. Russ. J. Plant Physiol. 2013, 60, 91–99. [Google Scholar]
- Fan, K.; Fan, D.; Ding, Z.; Su, Y.; Wang, X. Cs-miR156 is involved in thenitrogen form regulation of catechins accumulation in tea plant (Camellia sinensis L.). Plant Physiol. Bioch. 2015, 97, 350–360. [Google Scholar] [CrossRef]
- Cingolani, P.; Platts, A.; Wang, L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef]
- Tang, D.; Jiao, Z.; Zhang, Q.; Liu, M.Y.; Ruan, J. Glutamate dehydrogenase isogenes CsGDHs cooperate with glutamine synthetase isogenes CsGSs to assimilate ammonium in tea plant (Camellia sinensis L.). Plant Sci. 2021, 312, 111031. [Google Scholar] [CrossRef]
- Wang, C.M.; Tang, Q. Investigation of wild germplasm resources of loquat tea in Chongzhou. Southwest China J. Agric. Sci. 2012, 25, 642–648. [Google Scholar]
- Ates, F.; Kaya, O. The Relationship Between Iron and Nitrogen Concentrations Based On Kjeldahl Method and SPAD-502 Readings in Grapevine (Vitis vinifera L. cv. ‘Sultana Seedless’). Erwerbs-Obstbau 2021, 63, S53–S59. [Google Scholar] [CrossRef]
- Wei, C.; Yang, H.; Wang, S.; Zhao, J.; Liu, C.; Gao, L.; Xia, E.; Lu, Y.; Tai, Y.; She, G.; et al. Draft genome sequence of Camellia sinensis var. sinensis provides insights into the evolution of the tea genome and tea quality. Proc. Natl. Acad. Sci. USA 2018, 115, E4151–E4158. [Google Scholar] [CrossRef] [PubMed]
- Xia, E.H.; Li, F.D.; Tong, W.; Li, P.H.; Wu, Q.; Zhao, H.J.; Ge, R.H.; Li, R.P.; Li, Y.Y.; Zhang, Z.Z.; et al. Tea Plant Information Archive: A comprehensive genomics and bioinformatics platform for tea plant. Plant Biotechnol. J. 2019, 17, 1938–1953. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Sriramajayam, K.; Luo, D.; Liao, D.J. A Quick, Cost-Free Method of Purification of DNA Fragments from Agarose Gel. J. Cancer 2012, 3, 93–95. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 15 September 2023).
- Schubert, M.; Lindgreen, S.; Orlando, L. AdapterRemoval v2: Rapid adapter trimming, identification, and read merging. BMC Res. Notes 2016, 9, 88. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Picard Toolkit [Online Tool]. Available online: http://broadinstitute.github.io/picard/ (accessed on 30 May 2023).
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Breeding Status | Origin | Leaf Area, cm2 ± SD | Group |
|---|---|---|---|---|
| #507 | mutant form | ɣ-mutant | 40.8 ± 4.1 c | Middle |
| #3574 | mutant form | ɣ-mutant | 46.1 ± 4.6 c | Large |
| #855 | mutant form | ɣ-mutant | 55.6 ± 3.9 bc | Large |
| #527 | mutant form | ɣ-mutant | 57.4 ± 5.0 bc | Large |
| #3986 | mutant form | ɣ-mutant | 58.2 ± 5.1 bc | Large |
| #551 | mutant form | ɣ-mutant | 65.6 ± 5.9 b | Extra-large |
| #157 | mutant form | ɣ-mutant | 66.5 ± 5.8 b | Extra-large |
| #3180 | mutant form | ɣ-mutant | 68.0 ± 6.0 b | Extra-large |
| #536 | mutant form | ɣ-mutant | 68.7 ± 6.2 b | Extra-large |
| #1405 | mutant form | ɣ-mutant | 69.8 ± 6.0 b | Extra-large |
| #1877 | mutant form | ɣ-mutant | 70.6 ± 6.3 b | Extra-large |
| #582 | mutant form | ɣ-mutant | 71.0 ± 6.3 b | Extra-large |
| #1467 | mutant form | ɣ-mutant | 74.9 ± 6.7 b | Extra-large |
| Karatum | Cultivar | Sport mutation | 83.3 ± 6.5 ab | Extra-large |
| #1385 | mutant form | ɣ-mutant | 83.8 ± 7.1 ab | Extra-large |
| #212 | breeding line | Clonal selection | 84.6 ± 7.1 ab | Extra-large |
| #1102 | mutant form | ɣ-mutant | 92.4 ± 8.3 a | Extra-large |
| #316 | mutant form | ɣ-mutant | 97.3 ± 7.2 a | Extra-large |
| #2697 | mutant form | ɣ-mutant | 112.3 ± 7.0 a | Extra-large |
| #619 | mutant form | ɣ-mutant | 149.1 ± 7.3 a | Extra-large |
| Gene | Fragment Length, bp | Mean SNPs Number in Introns | Mean SNPs Number in Exons | SNP Density in Introns | SNP Density in Exons | SNP % in Exons |
|---|---|---|---|---|---|---|
| F3′5′Hb | 4435 | 6.25 | 7.60 | 1.41 | 1.71 | 54.87 |
| MYB4 | 5342 | 8.85 | 01.05 | 1.66 | 0.20 | 10.61 |
| LHT1 | 5107 | 3.50 | 1.40 | 0.69 | 0.27 | 28.57 |
| 4CL | 5264 | 31.85 | 9.95 | 6.05 | 1.89 | 23.80 |
| AMT1.2 | 2643 | 9.45 | 5.75 | 3.58 | 2.18 | 37.83 |
| GS1.2 | 6202 | 58.05 | 4.85 | 9.36 | 0.78 | 7.71 |
| DFRa | 6600 | 4.45 | 4.35 | 0.67 | 0.66 | 49.43 |
| WRKY57 | 11,214 | 41.85 | 4.25 | 3.73 | 0.38 | 9.22 |
| NRT2.4 | 3060 | 10.50 | 4.15 | 3.43 | 1.36 | 28.33 |
| F3′5′H_a | 5118 | 11.35 | 3.25 | 2.22 | 0.64 | 22.26 |
| AlaDC | 7227 | 5.00 | 1.85 | 0.69 | 0.26 | 27.01 |
| PIP | 2006 | 2.25 | 1.25 | 1.12 | 0.62 | 35.71 |
| bG | 8605 | 0.20 | 0.65 | 0.02 | 0.08 | 76.47 |
| WD40 | 3844 | 02.05 | 0.55 | 0.53 | 0.14 | 21.15 |
| GDH2 | 4915 | 4.50 | 0.20 | 0.92 | 0.04 | 4.26 |
| LAR | 8600 | 0.45 | 0.1 | 0.05 | 0.01 | 18.18 |
| AlaAT1 | 8058 | 1.85 | 0.05 | 0.23 | 0.01 | 2.63 |
| bHLH35 | 5743 | 3.35 | 0.05 | 0.58 | 0.01 | 1.47 |
| MYB7 | 3376 | 0.05 | 0.00 | 0.01 | 0.00 | 0.00 |
| bHLH36 | 2953 | 0.25 | 0.00 | 0.08 | 0.00 | 0.00 |
| SNPs | Gene | Position | Genotype (REF/ALT) | Phenotype | p Value | Amino (REF/ALT) | Property (REF/ALT) |
|---|---|---|---|---|---|---|---|
| SNP2 | ANSa | 1026570 | G/T | L-Theanine | 6.376794 × 10−8 | ||
| SNP3 | ANSa | 1026588 | A/C | Theaflavin | 3.458851 × 10−6 | ||
| SNP6 | ANSa | 1027478 | A/G | Theaflavin | 3.458851 × 10−6 | p.Leu165Pro | Hydrophobic/ Unique |
| SNP11 | DFRa | 1286200 | C/T | ECG/ Theabromine | 0.0001012753/ 1.208252 × 10−6 | ||
| SNP16 | GDH2 | 1343623 | T/C | ECG/ Theabromine | 0.0001012753/ 1.208252 × 10−6 | p.Lys368Arg | Charged_basic/ Charged_basic |
| SNP67 | 4CL | 2130618 | C/A | L-Theanine | 1.841082 × 10−6 | ||
| SNP184 | AlaAT1 | 531832 | A/G | L-Theanine | 6.376794 × 10−8 | ||
| SNP59 | 4CL | 2130419 | A/G | Theaflavin | 3.458851 × 10−6 | p.Asn15Ser | Neutral/Neutral |
| SNP60 | 4CL | 2130421 | A/T | Theaflavin | 3.458851 × 10−6 | p.Thr16Ser | Neutral/Neutral |
| SNP68 | 4CL | 2130888 | T/C | Theaflavin | 3.458851 × 10−6 | ||
| SNP120 | MYB4 | 254145 | A/G | Theaflavin | 3.458851 × 10−6 | ||
| SNP187 | LHT1 | 565216 | G/A | Theaflavin | 3.458851 × 10−6 | ||
| SNP193 | F3′5′Hb | 649286 | T/A | Theaflavin | 3.458851 × 10−6 | p.Lys505Met | Charged_basic/ Hydrophobic |
| SNP194 | F3′5′Hb | 649465 | G/A | Theaflavin | 0.0001046889 | ||
| SNP195 | F3′5′Hb | 649575 | G/A | Theaflavin | 3.458851 × 10−6 | p.Pro409Ser | Hydrophobic+ Unique/ Neutral |
| SNP260 | UFGTa | 883919 | A/G | Theaflavin | 3.458851 × 10−6 | p.Thr274Ala | Neutral/ Hydrophobic |
| Gene | Primer F 5′-3′ | Primer R 5′-3′ | Fragment Length, bp | Annealing Temp | Elongation, Min |
|---|---|---|---|---|---|
| UFGT_a | ACTCACAGGGATAAGAACACTC | CCATCTGGCAACATCTCCTC | 1455 | 58 | 3 |
| UFGT_a | ACTCACAGGGATAAGAACACTC | CACACTCCTCACCGCTCTT | 1747 | 58 | 3 |
| PIP | CCAAATGCCAAATATAGGGAGGC | GACCATAGAGAGAGTGGGTGG | 2006 | 62 | 2.5 |
| CHS | TTCGACCGCTTTGACCCGA | TGTTGCAGGCTCACATCACTC | 2201 | 59 | 4 |
| UFGT_b | TGATTGCTCATGTTATTTGGTCTC | TTGCCCATAACCTCTCCCTC | 2600 | 62 | 3 |
| AMT1.2 | CCATTGCTGCTGAGACTGATAA | TGAGGTGATGGTGATTTGACGG | 2643 | 61 | 2.5 |
| ANSa | GGATGAAGGTGAGGGCAGAG | TGTCAGGGCTTTTTCTAAATCGT | 2707 | 62 | 2.5 |
| bHLH36 | TCCATCAAAGAAGACAGACCACC | CCATTTGTCACCTTCTTGGTTTG | 2953 | 60 | 3 |
| NRT2.4 | TTGTCCTTCCTTGGCTCCTTG | ACCAACTGTGCTCATTAACAAATC | 3060 | 61 | 2.5 |
| MYB7 | GTTTGTGCTCTCACCGATTTCA | GTAGTAAACCCATACGTGGCCT | 3376 | 59 | 3.5 |
| WD40 | AGGTCGGTGTAATTTCCGCT | ACAAGTGGCTAACTCTCATTGGT | 3844 | 60 | 3 |
| ANS_b | GTTGAAGGTTGCATTATTGAAGTTG | CCATGAGGTTTTGATTAGTGGAGT | 3900 | 60 | 3 |
| F’3’5’Hb | AGCCTCCTCTGTGGTAAGTG | CTCGACTCCACGAATACTAACA | 4435 | 60 | 3.5 |
| GDH2 | GTCCTCCTATAACCTCCATTG | ATATTTGTTTTCTTGGTCACGTTTG | 4915 | 58 | 3.5 |
| ANRb | AGTCGTTGGTGCTGTGTGTT | AGGTTTACTTGGGTCTTCTTCAGG | 5000 | 61 | 3 |
| ANRa | GGTCCGTATGTGTGAATGAATCT | TGACTTGATCGTCGGAGTGC | 5023 | 59 | 4 |
| LHT1 | GCTGTCTTCAGTAGCGGACTT | TTGCACGTTAGAATCAGGGTCAT | 5107 | 58 | 5.5 |
| F3’5’Ha | TGTGGTTGAGAAGAAAGGATACCA | GGAGACACCATAGCCGAAAGA | 5118 | 58 | 3.5 |
| 4CL | GAGCTGCAATAGGCCTCACC | ACGACAGTAAAACCATAACAGAGT | 5264 | 61 | 4 |
| MYB4 | TGACCCCTCATCACATTTAGT | CGTAAGTTCGTGGTGGTAGTG | 5342 | 59 | 3.5 |
| FLS | AGCAAGGCTATCGAAGGTGT | AGGGGAAGGTTTTAGGCAAG | 5568 | 58 | 3.5 |
| bHLH35 | CGGAGAAATGATGGCGTGGT | GGCAAGTGGTTATGGGCGAA | 5743 | 59 | 3.5 |
| GS1.2 | TGTTGGTTGATGTTGATGATGTT | CCATTTAAGTAGGAAGATTTCGCC | 6202 | 61 | 4 |
| DFRa | AATGTGGTGAAAGTGGACGTAG | TCTTGTTGCTGGATGACTGAC | 6600 | 59 | 4 |
| AlaDC | TGACCCACCCAAACAACGAG | TGGCACACACAAACGATAGTAGA | 7227 | 59 | 6 |
| GDHa | ACCATCCTAGTCACGCCCAT | CATGTATTTCCACAACAGAACAAG | 7617 | 58 | 5.5 |
| AlaAT1 | GCTTCGTAAATGGAATCGCC | CCTTGGGCATTCGCAGCTT | 8058 | 59 | 7 |
| LAR | ACCTACACCAAAGACAACGAA | TGTTGTTATGGTGTTTGGTTGGTT | 8600 | 58 | 6 |
| bG | TCAGTAAGCCTTGAGAAAAGCA | GATGAAGTGTCACCTATTATGAGC | 8605 | 59 | 6 |
| WRKY57 | TGATTCTTCTTGGGTCTAAACAG | GCCCACACTCACTCAAATCC | 11,214 | 58 | 9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samarina, L.; Fedorina, J.; Kuzmina, D.; Malyukova, L.; Manakhova, K.; Kovalenko, T.; Matskiv, A.; Xia, E.; Tong, W.; Zhang, Z.; et al. Analysis of Functional Single-Nucleotide Polymorphisms (SNPs) and Leaf Quality in Tea Collection under Nitrogen-Deficient Conditions. Int. J. Mol. Sci. 2023, 24, 14538. https://doi.org/10.3390/ijms241914538
Samarina L, Fedorina J, Kuzmina D, Malyukova L, Manakhova K, Kovalenko T, Matskiv A, Xia E, Tong W, Zhang Z, et al. Analysis of Functional Single-Nucleotide Polymorphisms (SNPs) and Leaf Quality in Tea Collection under Nitrogen-Deficient Conditions. International Journal of Molecular Sciences. 2023; 24(19):14538. https://doi.org/10.3390/ijms241914538
Chicago/Turabian StyleSamarina, Lidiia, Jaroslava Fedorina, Daria Kuzmina, Lyudmila Malyukova, Karina Manakhova, Tatyana Kovalenko, Alexandra Matskiv, Enhua Xia, Wei Tong, Zhaoliang Zhang, and et al. 2023. "Analysis of Functional Single-Nucleotide Polymorphisms (SNPs) and Leaf Quality in Tea Collection under Nitrogen-Deficient Conditions" International Journal of Molecular Sciences 24, no. 19: 14538. https://doi.org/10.3390/ijms241914538
APA StyleSamarina, L., Fedorina, J., Kuzmina, D., Malyukova, L., Manakhova, K., Kovalenko, T., Matskiv, A., Xia, E., Tong, W., Zhang, Z., Ryndin, A., Orlov, Y. L., & Khlestkina, E. K. (2023). Analysis of Functional Single-Nucleotide Polymorphisms (SNPs) and Leaf Quality in Tea Collection under Nitrogen-Deficient Conditions. International Journal of Molecular Sciences, 24(19), 14538. https://doi.org/10.3390/ijms241914538