
Improvement of Rice Blast Resistance in TGMS Line HD9802S through Optimized Anther Culture and Molecular Marker-Assisted Selection

 , , ,
, , , 
Abstract
1. Introduction
2. Results
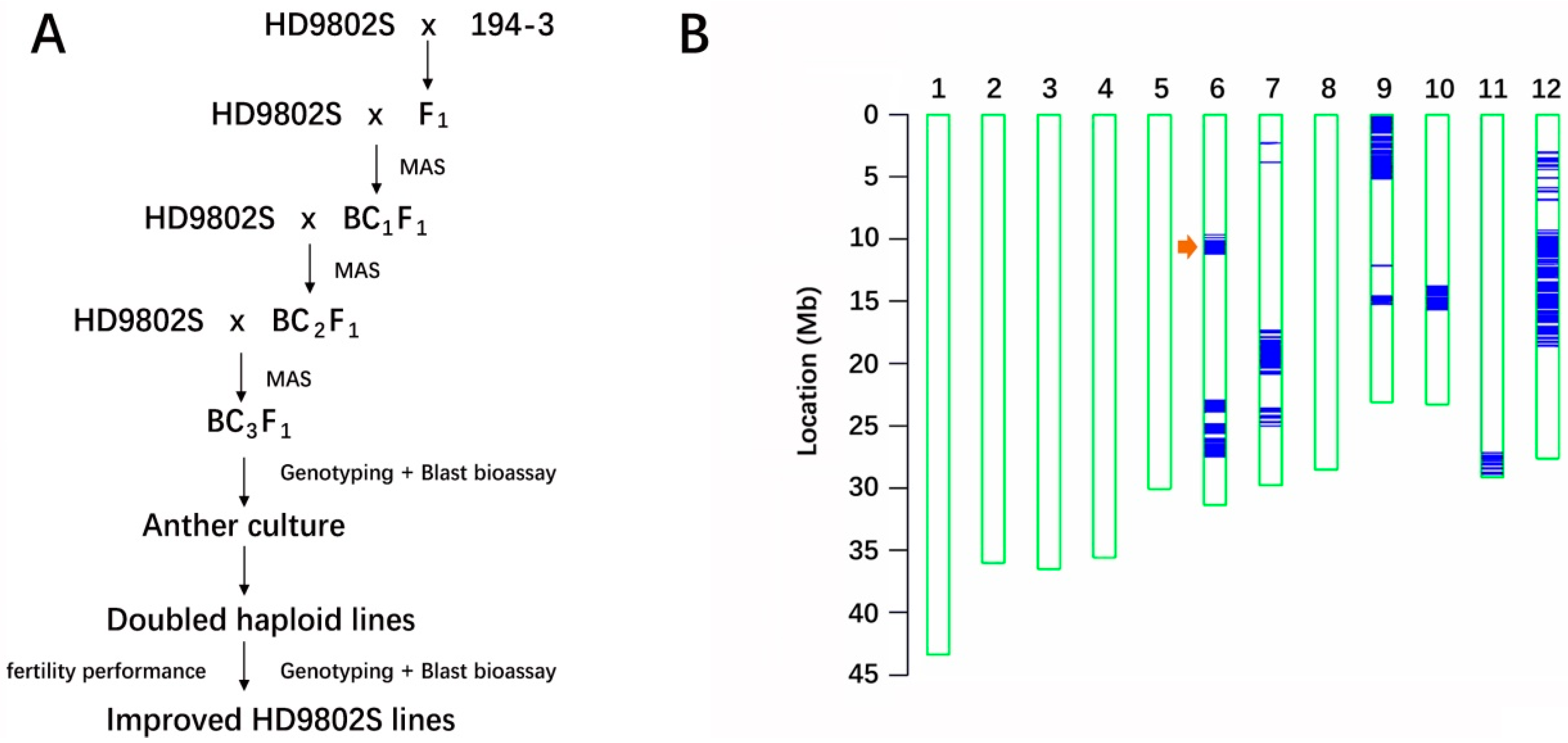
2.1. Introgression of Blast-Resistance Gene R6 into the Background of HD9802S
2.2. Anther Culture
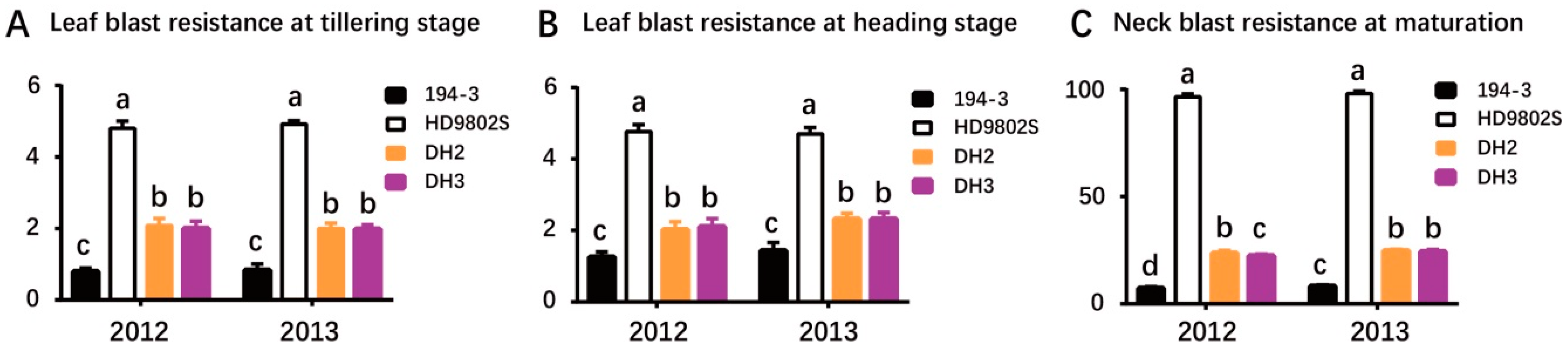
2.3. Evaluation of Rice Blast Resistance
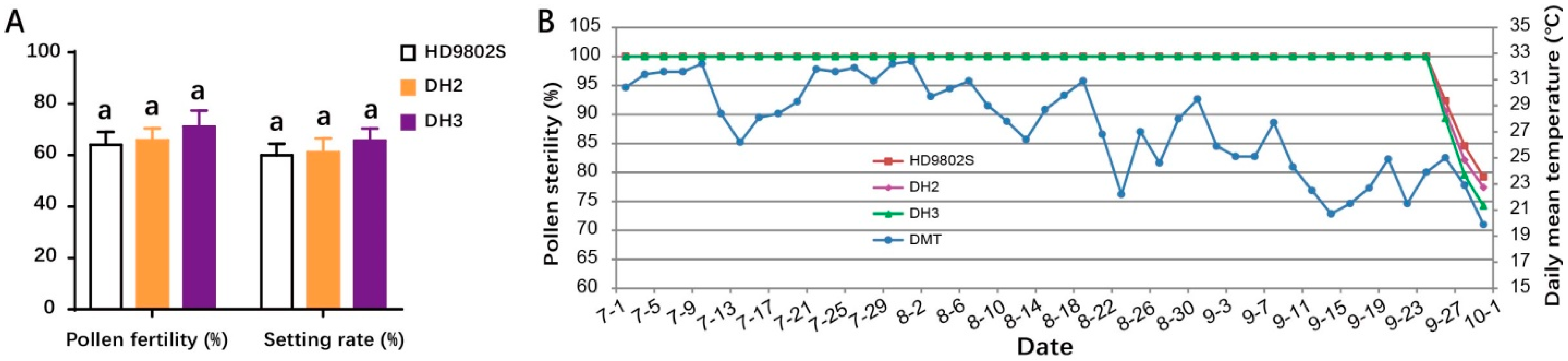
2.4. Characterization of Fertility
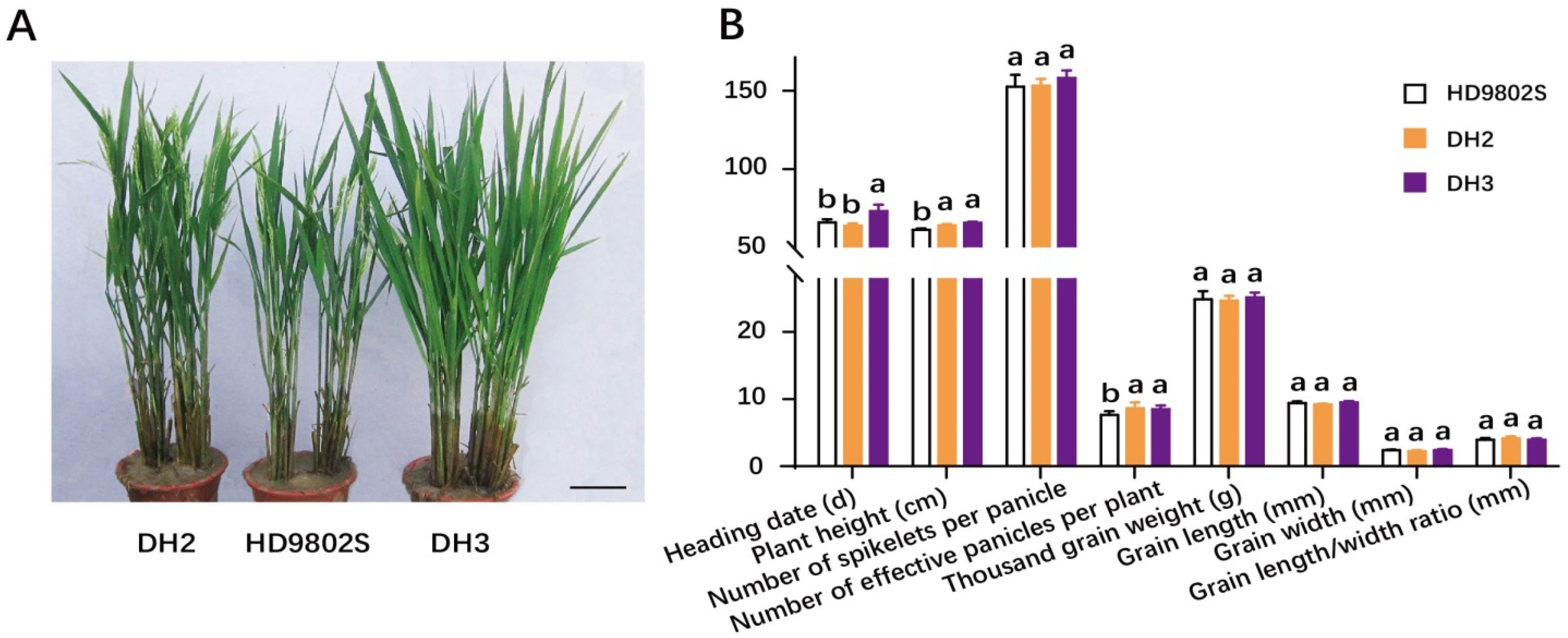
2.5. Investigation of Agronomic and Rice Grain Quality Traits
3. Discussion
3.1. Selection of AC Medium
3.2. Technology Roadmap for the Rapid Improvement of Target Phenotype by Combining MAS with AC
4. Materials and Methods
4.1. Plant Materials and Field Planting
4.2. Molecular Markers and Genotyping Used for MAS
4.3. Anther Culture
4.3.1. Culture Medium
4.3.2. AC Procedure
4.4. Rice Blast Evaluation
4.5. Fertility Assessment
4.6. Ploidy Identification
4.7. Evaluation of Agronomic and Rice Grain Quality Traits
4.8. Data Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kiyosawa, S. Genetics and Epidemiological Modeling of Breakdown of Plant Disease Resistance. Annu. Rev. Phytopathol. 1982, 20, 93–117. [Google Scholar] [CrossRef]
- Mapping of leaf and neck blast resistance genes with resistance gene analog, RAPD and RFLP in rice. Euphytica 2002, 128, 363–370. [CrossRef]
- Dean, R.A.; Talbot, N.J.; Ebbole, D.J.; Farman, M.L.; Mitchell, T.K.; Orbach, M.J.; Thon, M.; Kulkarni, R.; Xu, J.R.; Pan, H. The genome sequence of the rice blast fungus Magnaporthe grisea. Nature 2005, 434, 980. [Google Scholar] [CrossRef] [PubMed]
- Mcclung, A.M.; Marchetti, M.A.; Webb, B.D.; Bollich, C.N. Registration of ‘Jefferson’ Rice. Crop Sci. 1997, 37, 629–630. [Google Scholar] [CrossRef]
- Deng, Y.; Zhai, K.; Xie, Z.; Yang, D.; Zhu, X.; Liu, J.; Wang, X.; Qin, P.; Yang, Y.; Zhang, G.; et al. Epigenetic regulation of antagonistic receptors confers rice blast resistance with yield balance. Science 2017, 355, 962–965. [Google Scholar] [CrossRef]
- Ashkani, S.; Yusop, M.R.; Shabanimofrad, M.; Azadi, A.; Ghasemzadeh, A.; Azizi, P.; Abdul Latif, M. Allele Mining Strategies: Principles and Utilisation for Blast Resistance Genes in Rice (Oryza sativa L.). Curr. Issues Mol. Biol. 2015, 17, 57–74. [Google Scholar]
- Wang, B.H.; Daniel, J.E.; Wang, Z.H. The arms race between Magnaporthe oryzae and rice: Diversity and interaction of Avr and R genes. J. Integr. Agric. 2017, 16, 2746–2760. [Google Scholar] [CrossRef]
- Meng, X.; Xiao, G.; Telebanco-Yanoria, M.J.; Siazon, P.M.; Zhou, B. The broad-spectrum rice blast resistance (R) gene Pita2 encodes a novel R protein unique from Pita. Rice 2020, 13, 19. [Google Scholar] [CrossRef]
- Xiao, G.; Borja, F.N.; Mauleon, R.; Padilla, J.; Telebanco-Yanoria, M.J.; Yang, J.; Lu, G.; Dionisio-Sese, M.; Zhou, B. Identification of resistant germplasm containing novel resistance genes at or tightly linked to the Pi2/9 locus conferring broad-spectrum resistance against rice blast. Rice 2017, 10, 37. [Google Scholar] [CrossRef]
- Yu, Z.H.; Mackill, D.J.; Bonman, J.M.; Tanksley, S.D. Tagging genes for blast resistance in rice via linkage to RFLP markers. Theor. Appl. Genet. 1991, 81, 471–476. [Google Scholar] [CrossRef]
- Jiang, H.; Li, Z.; Liu, J.; Shen, Z.; Gao, G.; Zhang, Q.; He, Y. Development and evaluation of improved lines with broad-spectrum resistance to rice blast using nine resistance genes. Rice 2019, 12, 29. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Mou, T.; Yu, H.; Zhou, F. Molecular breeding of thermo-sensitive genic male sterile (TGMS) lines of rice for blast resistance using Pi2 gene. Rice 2015, 8, 11. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhu, X.; Chen, S.; Yang, J.; Zhou, S.; Zeng, L.; Han, J.; Su, J.; Pan, W.Q. The identification of Pi50(t), a new member of the rice blast resistance Pi2/Pi9 multigene family. Theor. Appl. Genet. 2012, 124, 1295–1304. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Wu, J.; Li, X.; Wang, X.; Liu, X.; Jantasuriyarat, C.; Kudrna, D.; Yu, Y.; Wing, R.A.; Han, B. Genomic structure and evolution of the Pi2/9 locus in wild rice species. Theor. Appl. Genet. 2010, 121, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Lu, G.; Zeng, L.; Wang, G.L. Two broad-spectrum blast resistance genes, Pi9(t) and Pi2(t), are physically linked on rice chromosome 6. Mol. Genet. Genom. 2002, 267, 472–480. [Google Scholar] [CrossRef] [PubMed]
- Sharma, T.R.; Madhav, M.S.; Singh, B.K.; Shanker, P.; Singh, N.K. High-resolution mapping, cloning and molecular characterization of the Pi-kh gene of rice, which confers resistance to Magnaporthe grisea. Mol. Genet. Genom. 2006, 274, 569–578. [Google Scholar] [CrossRef]
- Li, W.; Zhu, Z.; Chern, M.; Yin, J.; Yang, C.; Ran, L.; Cheng, M.; He, M.; Wang, K.; Wang, J. A Natural Allele of a Transcription Factor in Rice Confers Broad-Spectrum Blast Resistance. Cell 2017, 170, 114–126. [Google Scholar] [CrossRef]
- Yang, D.; Tang, J.; Yang, D.; Chen, Y.; Mou, T. Improving rice blast resistance of Feng39S through molecular marker-assisted backcrossing. Rice 2019, 12, 70. [Google Scholar] [CrossRef]
- Wu, Y.P.; Chang, Y.C.; Kuo, H.I.; Lin, B.N.; Wang, S.M.; Tseng, Y.C. The Development of Two High-Yield and High-Quality Functional Rice Cultivars Using Marker-Assisted Selection and Conventional Breeding Methods. Int. J. Mol. Sci. 2022, 23, 4678. [Google Scholar] [CrossRef]
- Wen, S.; Gao, B. Introgressing Blast Resistant Gene Pi-9(t) into Elite Rice Restorer Luhui17 by Marker-Assisted Selection. Mol. Plant Breed. 2012, 2, 31–36. [Google Scholar] [CrossRef][Green Version]
- Jiang, H.; Feng, Y.; Bao, L.; Li, X.; Gao, G.; Zhang, Q.; Xiao, J.; Xu, C.; He, Y. Improving blast resistance of Jin 23B and its hybrid rice by marker-assisted gene pyramiding. Mol. Breed. 2012, 30, 1679–1688. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, F.; Luo, X.; Kong, D.; Yu, X. Molecular Breeding of a Novel PTGMS Line of Wdr for Broad-spectrum Resistance to Blast Using Pi9, Pikh, and Pi5 Genes. Rice 2021, 14, 96. [Google Scholar] [CrossRef] [PubMed]
- Lantos, C.; Purgel, S.; Ács, K.; Langó, B.; Bóna, L.; Boda, K.; Békés, F.; Pauk, J. Utilization of in Vitro Anther Culture in Spelt Wheat Breeding. Plants 2019, 8, 436. [Google Scholar] [CrossRef] [PubMed]
- Ohnoutková, L.; Vlčko, T. Homozygous Transgenic Barley (Hordeum vulgare L.) Plants by Anther Culture. Plants 2020, 9, 918. [Google Scholar] [CrossRef]
- Lantos, C.; Jancsó, M. Development of In Vitro Anther Culture for Doubled Haploid Plant Production in Indica Rice (Oryza sativa L.) Genotypes. Plants 2023, 12, 1774. [Google Scholar] [CrossRef]
- Raina, S.K.; Zapata, F.J. Enhanced anther culture efficiency of indica rice (Oryza sativa L.) through modification of the culture media. Plant Breed. 2010, 116, 305–315. [Google Scholar] [CrossRef]
- Bartolome, V.; Grewal, D.; Manito, C. Doubled Haploids Generated through Anther Culture from Crosses of Elite Indica and Japonica Cultivars and/or Lines of Rice: Large-Scale Production, Agronomic Performance, and Molecular Characterization. Crop Sci. 2011, 51, 2544. [Google Scholar]
- Germanà, M.A. Gametic embryogenesis and haploid technology as valuable support to plant breeding. Plant Cell Rep. 2011, 30, 839–857. [Google Scholar] [CrossRef]
- Niu, Z.; Jiang, A.; Abu Hammad, W.; Oladzadabbasabadi, A.; Xu, S.S.; Mergoum, M.; Elias, E.M.; Hartl, L. Review of doubled haploid production in durum and common wheat through wheat × maize hybridization. Plant Breed. 2014, 133, 313–320. [Google Scholar] [CrossRef]
- Wijerathna-Yapa, A.; Ramtekey, V.; Ranawaka, B.; Basnet, B.R. Applications of In Vitro Tissue Culture Technologies in Breeding and Genetic Improvement of Wheat. Plants 2022, 11, 2273. [Google Scholar] [CrossRef]
- He, Y.; Chen, L.; Xu, C. Variability of Anther Culture and the Technology Strategies for Developing Pract ical Photoperiod Sensitive Male Sterile Rice. Acta Agron. Sin. 2000, 26, 879–884. [Google Scholar]
- Yuan, L.; Song, D.; Gao, G.; He, Y. Improvement of Resistance to Rice Blast in PGMS Line Y58S by Molecular Marker-assisted Selection and Anther Culture. Genom. Appl. Biol. 2011, 30, 620–625. [Google Scholar]
- Zhou, Y.; Ju, C.; Xu, G.; Huang, Z.; Zhao, H.; Xie, P.; Gao, M.X. Breeding and Utilization of Fine Quality Indica TGMS Line HD9802S in Rice. Hybrid Rice 2008, 23, 7–10. [Google Scholar]
- Zhang, X.; Zuo, B.; Song, Z.; Wang, W.; Cai, D. Breeding and study of two new photoperiod- and thermo-sensitive genic male sterile lines of polyploid rice (Oryza sativa L.). Sci. Rep. 2017, 7, 14744. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Li, G.; Bao, L.; He, Y. Genetic Analysis of Rice Blast Resistance and Identification of Resistance Genes Throughout all Stages in Rice. Mol. Plant Breed. 2008, 6, 825–829. [Google Scholar]
- Jiang, H.; Feng, Y.; Qiu, L.; Gao, G.; He, Y. Identification of Blast Resistance QTLs Based on Two Advanced Backcross Populations in Rice. Rice 2020, 13, 31. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Xie, W.; Li, J.; Zhou, F.; Zhang, Q. A whole-genome SNP array (RICE6K) for genomic breeding in rice. Plant Biotechnol. J. 2013, 12, 28–37. [Google Scholar] [CrossRef]
- Leslie, K. Improved Anther Culture Media for Enhanced Callus Formation and Plant Regeneration in Rice (Oryza sativa L.). Plants 2021, 10, 839. [Google Scholar]
- Haiming, X.U.; Jun, Z. Statistical approaches in QTL mapping and molecular breeding for complex traits. Chin. Sci. Bull. 2012, 57, 8. [Google Scholar]
- Bonman, J.M.; Vergel, D.; Dios, T.I.; Khin, M.M. Physiologic specialization of Pyricularia oryzae in the Philippines. Plant Dis. 1986, 70, 767–769. [Google Scholar] [CrossRef]
- Dolezel, J.; Greilhuber, J.; Suda, J. Estimation of nuclear DNA content in plants using flow cytometry. Nat. Protoc. 2007, 2, 2233–2244. [Google Scholar] [CrossRef] [PubMed]
- Pang, H.H.; Liu, X. Induction of pollen plant of hybrid offspring from cultivated and wild rice and their genetic expression. Acta Agron. Sin. 2000, 26, 47–52. [Google Scholar]
- Song, W.C.; Zhang, Y.H. Tetraploidy of rice and its impact on agronomic traits and nutritional composition. Acta Agron. Sin. 1992, 18, 138–144. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium | MS | N6 | 2,4-D mg/L | NAA mg/L | KT mg/L | CH mg/L | Pro mg/L | Glu mg/L | Surose g/L | Maltose g/L | No. of Anther | CIR (%) | No. of Plant | GSR (%) | WSR (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IM1 | + | − | 3.00 | - | - | 800 | 600 | 500 | 20 | 30 | 3000 | 4.00 | 2 | 1.67 | 29.17 |
| IM2 | + | − | 3.00 | 2.00 | - | 800 | 600 | 500 | - | 30 | 4300 | 3.09 | 2 | 1.50 | 24.06 |
| IM3 | + | − | 3.00 | 2.00 | - | 800 | 600 | 500 | 20 | 30 | 4600 | 4.00 | 5 | 2.72 | 21.74 |
| IM4 | + | − | 3.00 | 2.00 | 1.00 | 800 | 600 | 500 | 20 | 30 | 5150 | 7.03 | 11 | 3.04 | 20.72 |
| IM5 | − | + | 2.00 | 3.00 | 1.00 | 800 | - | - | 60 | - | 5600 | 3.21 | 4 | 2.22 | 30.00 |
| IM6 | − | + | 2.00 | 3.00 | 1.00 | 800 | 600 | 500 | 60 | - | 3950 | 7.06 | 9 | 3.23 | 26.88 |
| Trait | Year | 194-3 | HD9802S | DH2 | DH3 |
|---|---|---|---|---|---|
| LT | 2012 | 0.79 ± 0.10 c | 4.80 ± 0.21 a | 2.08 ± 0.20 b | 2.00 ± 0.20 b |
| 2013 | 0.83 ± 0.18 c | 4.92 ± 0.10 a | 2.00 ± 0.15 b | 1.98 ± 0.12 b | |
| LH | 2012 | 1.25 ± 0.15 c | 4.77 ± 0.19 a | 2.02 ± 0.22 b | 2.10 ± 0.23 b |
| 2013 | 1.43 ± 0.23 c | 4.70 ± 0.18 a | 2.32 ± 0.16 b | 2.32 ± 0.18 b | |
| NB | 2012 | 7.18 ± 0.80 d | 96.50 ± 1.38 a | 23.50 ± 1.38 b | 22.33 ± 0.82 c |
| 2013 | 8.12 ± 0.66 c | 98.00 ± 1.55 a | 24.67 ± 0.82 b | 24.33 ± 1.03 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, P.; Gao, G.; Lou, G.; Hu, J.; Wang, Y.; Liu, R.; Zhao, D.; Liu, Q.; Sun, B.; Mao, X.; et al. Improvement of Rice Blast Resistance in TGMS Line HD9802S through Optimized Anther Culture and Molecular Marker-Assisted Selection. Int. J. Mol. Sci. 2023, 24, 14446. https://doi.org/10.3390/ijms241914446
Chen P, Gao G, Lou G, Hu J, Wang Y, Liu R, Zhao D, Liu Q, Sun B, Mao X, et al. Improvement of Rice Blast Resistance in TGMS Line HD9802S through Optimized Anther Culture and Molecular Marker-Assisted Selection. International Journal of Molecular Sciences. 2023; 24(19):14446. https://doi.org/10.3390/ijms241914446
Chicago/Turabian StyleChen, Pingli, Guanjun Gao, Guangming Lou, Jie Hu, Yufu Wang, Rongjia Liu, Da Zhao, Qing Liu, Bingrui Sun, Xingxue Mao, and et al. 2023. "Improvement of Rice Blast Resistance in TGMS Line HD9802S through Optimized Anther Culture and Molecular Marker-Assisted Selection" International Journal of Molecular Sciences 24, no. 19: 14446. https://doi.org/10.3390/ijms241914446
APA StyleChen, P., Gao, G., Lou, G., Hu, J., Wang, Y., Liu, R., Zhao, D., Liu, Q., Sun, B., Mao, X., Jiang, L., Zhang, J., Lv, S., Yu, H., Chen, W., Fan, Z., Li, C., & He, Y. (2023). Improvement of Rice Blast Resistance in TGMS Line HD9802S through Optimized Anther Culture and Molecular Marker-Assisted Selection. International Journal of Molecular Sciences, 24(19), 14446. https://doi.org/10.3390/ijms241914446