
Cell Cycle Reactivation, at the Start of Neurodegeneration, Induced by Forskolin and Aniline in Differentiated Neuroblastoma Cells
 , ,
, , 
 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Results
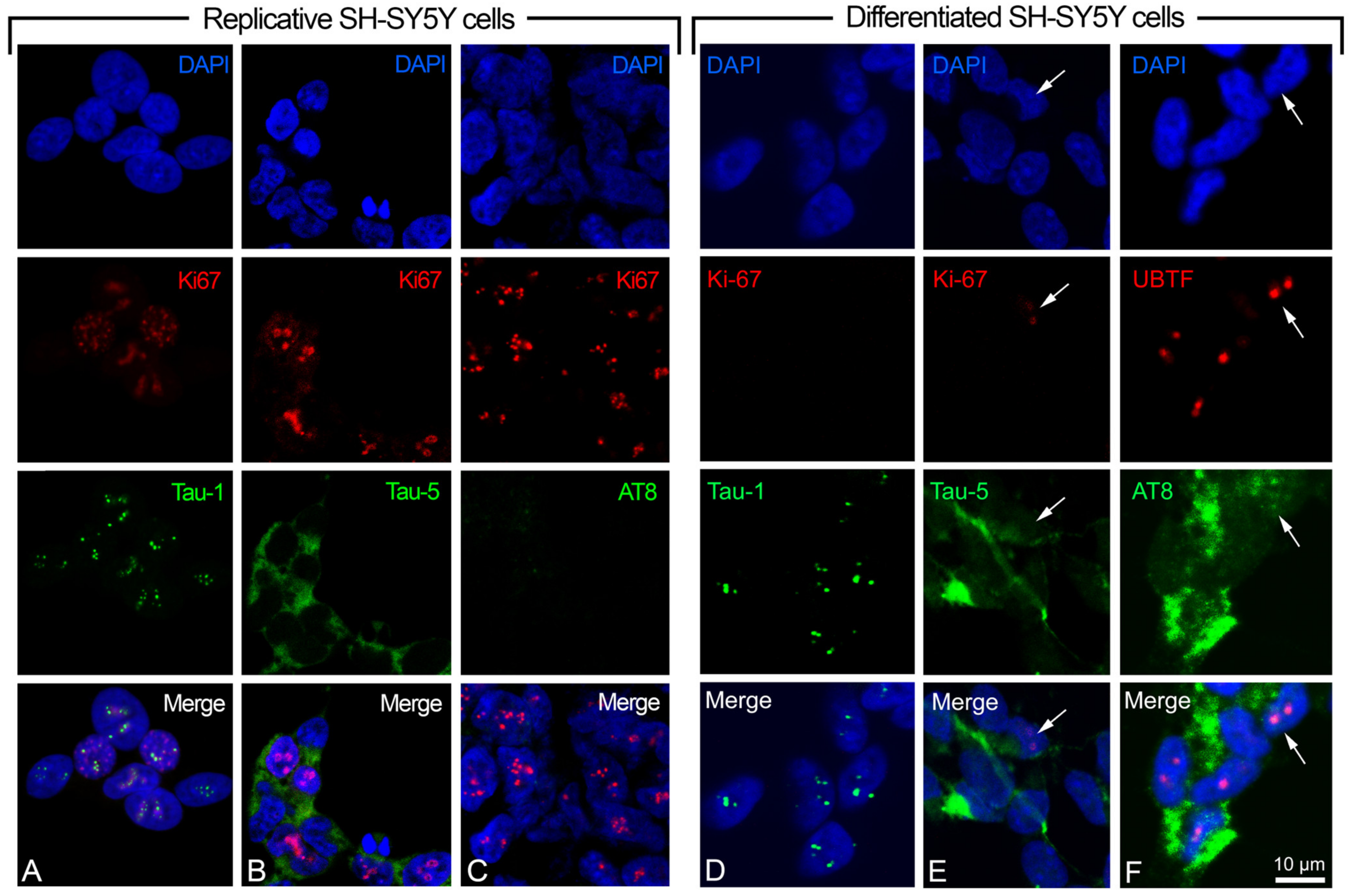
2.1. Neuronal Differentiation of SH-SY5Y Cells, and Detection of Tau1/AT8 Epitopes
2.2. MAPT Gene Expression, and Isoform Detection
2.3. Effect of Forskolin on the SH-SY5Y Cell Line
2.4. Effect of Aniline on the SH-SY5Y Cell Line
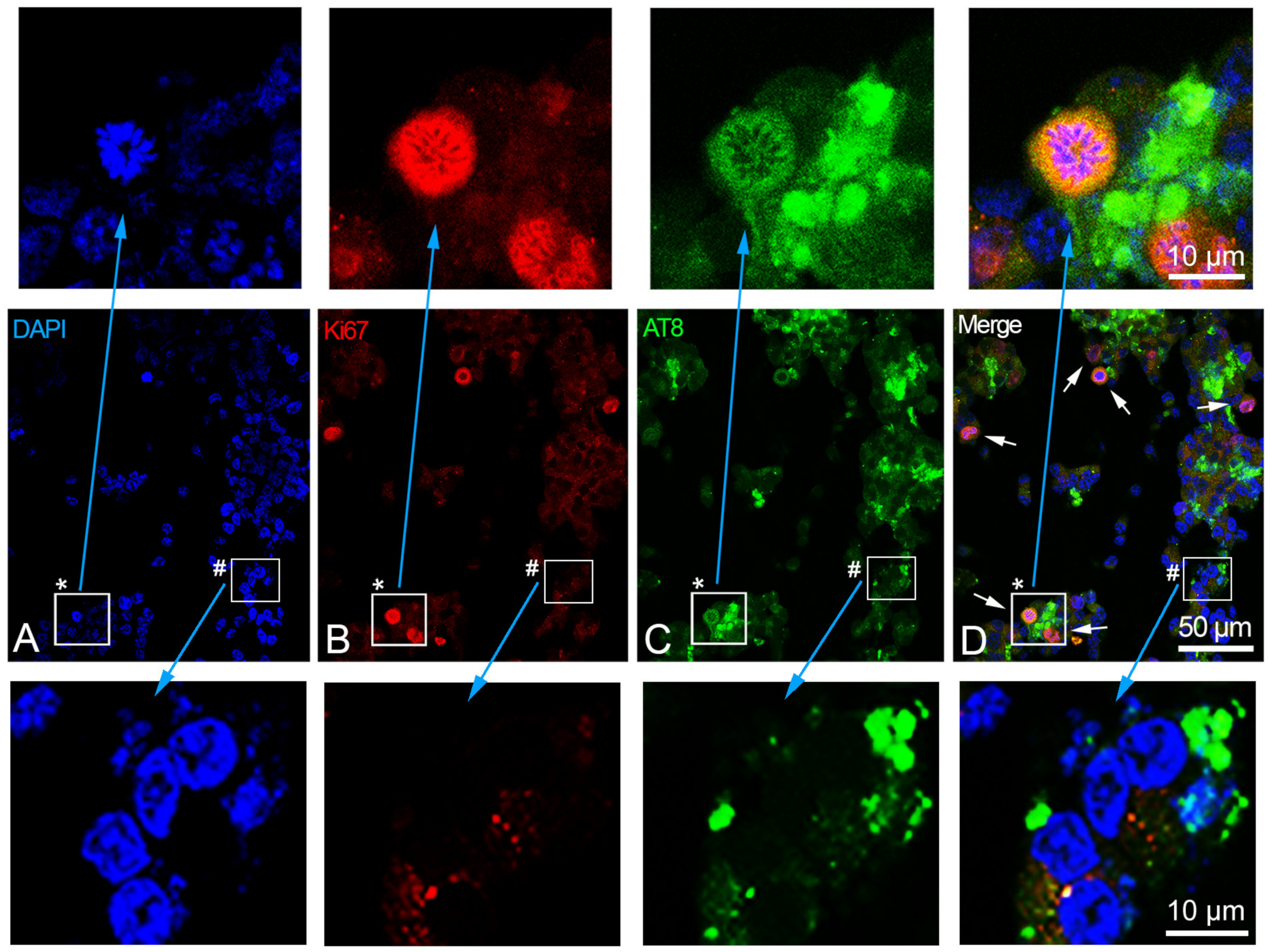
2.5. Blocking of Cell Cycle Reactivation
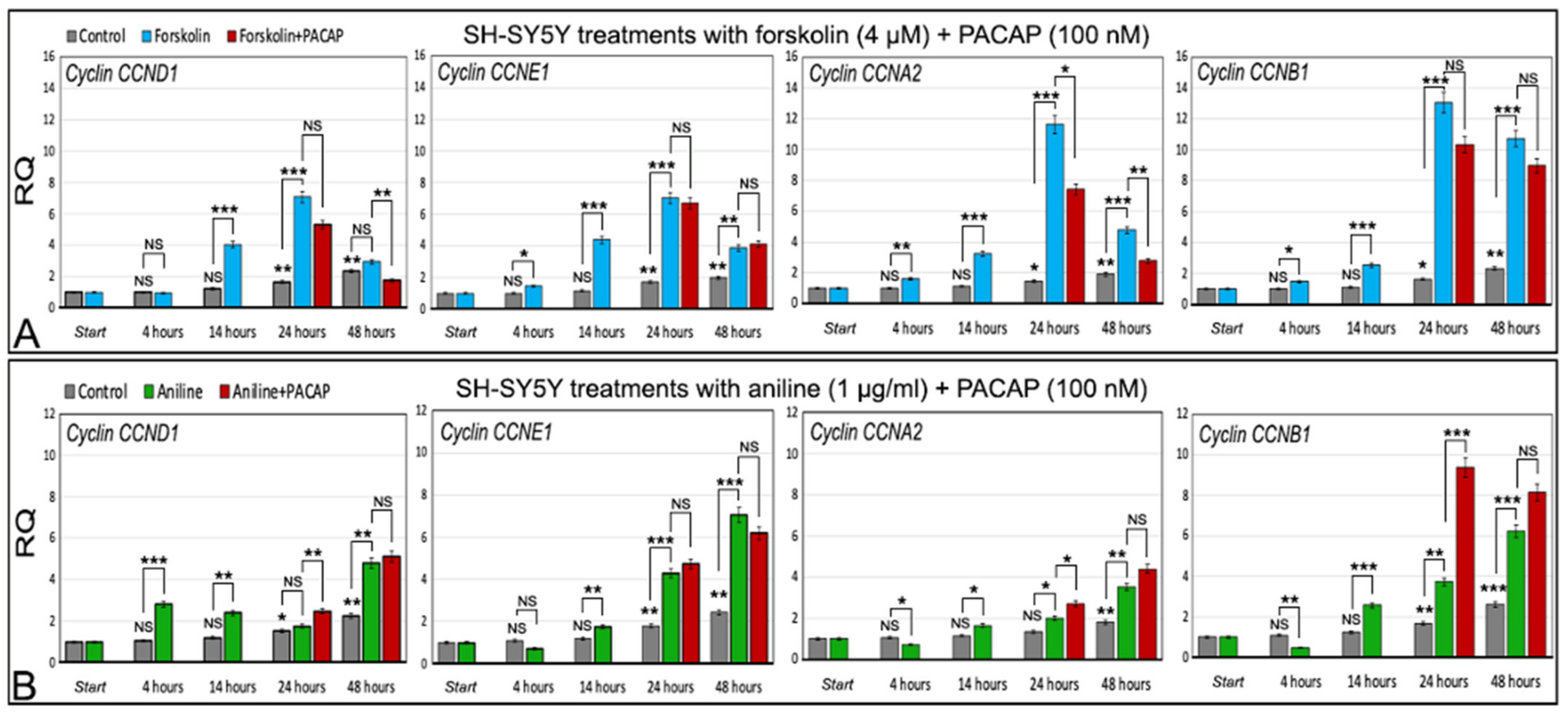
2.5.1. Cyclins in Control Cells Exposed to Forskolin
2.5.2. Cyclins in Control Cells Exposed to Aniline
2.5.3. Forskolin and PACAP
2.5.4. Aniline and PACAP
3. Discussion
4. Materials and Methods
4.1. Cell Cultures, and Neuronal Differentiation
4.2. Detection of the MAPT Gene Isoforms
4.3. Cyclin Expression Analysis
4.4. Indirect Immunofluorescence Staining
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Satyanarayana, A.; Kaldis, P. Mammalian cell-cycle regulation: Several Cdks, numerous cyclins and diverse compensatory mechanisms. Oncogene 2009, 28, 2925–2939. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Michowski, W.; Kolodziejczyk, A.; Sicinski, P. The cell cycle in stem cell proliferation, pluripotency and differentiation. Nat. Cell Biol. 2019, 21, 1060–1067. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.H.; Stoeber, K. The cell cycle and cancer. J. Pathol. 2012, 226, 352–364. [Google Scholar] [CrossRef] [PubMed]
- Vigneron, S.; Sundermann, L.; Labb, J.C.; Pintard, L.; Radulescu, O.; Castro, A.; Lorca, T. Cyclin A-cdk1-Dependent Phosphorylation of Bora Is the Triggering Factor Promoting Mitotic Entry. Dev. Cell 2018, 45, 637–650.e7. [Google Scholar] [CrossRef] [PubMed]
- Hégarat, N.; Crncec, A.; Suarez Peredo Rodriguez, M.F.; Echegaray Iturra, F.; Gu, Y.; Busby, O.; Lang, P.F.; Barr, A.R.; Bakal, C.; Kanemaki, M.T.; et al. Cyclin A triggers Mitosis either via the Greatwall kinase pathway or Cyclin B. EMBO J. 2020, 39, e104419. [Google Scholar] [CrossRef]
- Duronio, R.J.; Xiong, Y. Signaling pathways that control cell proliferation. Cold Spring Harb. Perspect. Biol. 2013, 5, a008904. [Google Scholar] [CrossRef]
- Ohnuma, S.; Harris, W.A. Neurogenesis and the cell cycle. Neuron 2003, 40, 199–208. [Google Scholar] [CrossRef]
- Verdaguer, E.; Garc-Jord, E.; Canudas, A.M.; Domnguez, E.; Jimnez, A.; Pubill, D.; Escubedo, E.; Pallàs, J.C.M.; Camins, A. Kainic acid-induced apoptosis in cerebellar granule neurons: An attempt at cell cycle re-entry. Neuroreport 2002, 13, 413–416. [Google Scholar] [CrossRef]
- Walton, C.C.; Zhang, W.; Pati-Parrado, I.; Barrio-Alonso, E.; Garrido, J.J.; Frade, J.M. Primary neurons can enter M-phase. Sci. Rep. 2019, 9, 4594. [Google Scholar] [CrossRef]
- Goedert, M. Tau protein and neurodegeneration. Semin. Cell Dev. Biol. 2004, 15, 45–49. [Google Scholar] [CrossRef]
- Surguchov, A.; Emamzadeh, F.N.; Titova, M.; Surguchev, A.A. Controversial properties of amyloidogenic proteins and peptides: New data in the COVID era. Biomedicines 2023, 11, 1215. [Google Scholar] [CrossRef]
- Yin, X.; Qiu, Y.; Zhao, C.; Zhou, Z.; Bao, J.; Qian, W. The Role of Amyloid-Beta and Tau in the Early Pathogenesis of Alzheimer’s Disease. Med. Sci. Monit. 2021, 27, e933084. [Google Scholar] [CrossRef] [PubMed]
- Andreadis, A. Tau gene alternative splicing: Expression patterns, regulation and modulation of function in normal brain and neurodegenerative diseases. Biochim. Biophys. Acta 2005, 1739, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Loomis, P.A.; Howard, T.H.; Castleberry, R.P.; Binder, L.I. Identification of nuclear tau isoforms in human neuroblastoma cells. Proc. Natl. Acad. Sci. USA 1990, 87, 8422–8426. [Google Scholar] [CrossRef] [PubMed]
- Sjoberg, M.K.; Shestakova, E.; Mansuroglu, Z.; Maccioni, R.B.; Bonnefoy, E. Tau protein binds to pericentromeric DNA: A putative role for nuclear tau in nucleolar organization. J. Cell Sci. 2006, 119, 2025–2034. [Google Scholar] [CrossRef] [PubMed]
- Sultan, A.; Nesslany, F.; Violet, M.; Bégard, S.; Loyens, A.; Talahari, S.; Mansuroglu, Z.; Marzin, D.; Sergeant, N.; Humez, S.; et al. Nuclear tau, a key player in neuronal DNA protection. J. Biol. Chem. 2011, 286, 4566–4575. [Google Scholar] [CrossRef]
- Hernández-Ortega, K.; Garcia-Esparcia, P.; Gil, L.; Lucas, J.J.; Ferrer, I. Altered Machinery of Protein Synthesis in Alzheimer’s: From the Nucleolus to the Ribosome. Brain Pathol. 2015, 26, 593–605. [Google Scholar] [CrossRef]
- Mansuroglu, Z.; Benhelli-Mokrani, H.; Marcato, V.; Sultan, A.; Violet, M.; Chauderlier, A.; Delattre, L.; Loyens, A.; Talahari, S.; Bégard, S.; et al. Loss of Tau protein affects the structure, transcription and repair of neuronal pericentromeric heterochromatin. Sci. Rep. 2016, 6, 33047. [Google Scholar] [CrossRef]
- Bukar Maina, M.; Al-Hilaly, Y.K.; Serpell, L.C. Nuclear tau and its potential role in Alzheimer’s disease. Biomolecules 2016, 6, 9. [Google Scholar] [CrossRef]
- Gil, L.; Federico, C.; Pinedo, F.; Bruno, F.; Rebolledo, A.B.; Montoya, J.J.; Olazabal, I.M.; Ferrer, I.; Saccone, S. Aging dependent effect of nuclear tau. Brain Res. 2017, 1677, 129–137. [Google Scholar] [CrossRef]
- Federico, C.; Bruno, F.; Sturiale, V.; D’Amico, A.G.; Maugeri, G.; D’agata, V.; Gil, L.; Saccone, S. Human nuclear tau and aging. In Factors Affecting Nuerological Aging: Genetics, Neurology, Behavior, and Diet; Martin, C.R., Preedy, V.R., Rajendram, R., Eds.; Academic Press: Cambridge, MA, USA; Elsevier Inc.: Oxford, UK, 2021; ISBN 978-0-12-817990-1. [Google Scholar] [CrossRef]
- Federico, C.; Gil, L.; Bruno, F.; D’Amico, A.G.; D’Agata, V.; Saccone, S. Phosphorylated nucleolar Tau protein is related to the neuronal in vitro differentiation. Gene 2018, 664, 1–11. [Google Scholar] [CrossRef]
- Kovalevich, J.; Langford, D. Considerations for the use of SH-SY5Y neuroblastoma cells in neurobiology. Methods Mol. Biol. 2013, 1078, 9–21. [Google Scholar] [CrossRef]
- Shipley, M.M.; Mangold, C.A.; Szpara, M.L. Differentiation of the SH-SY5Y Human Neuroblastoma Cell Line. J. Vis. Exp. 2016, 108, 53193. [Google Scholar] [CrossRef]
- Krylova, S.M.; Musheev, M.; Nutiu, R.; Li, Y.; Lee, G.; Krylov, S.N. Tau protein binds single-stranded DNA sequence specifically the proof obtained in vitro with non-equilibrium capillary electrophoresis of equilibrium mixtures. FEBS Lett. 2005, 579, 1371–1375. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Qu, M.H.; Wang, X.S.; Chen, L.; Wang, D.L.; Liu, Y.; Hua, Q.; He, R.Q. Binding to the minor groove of the double-strand, tau protein prevents DNA from damage by peroxidation. PLoS ONE 2008, 3, e2600. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.H.; Li, Y.; Li, A.; Yan, F.; Li, Z.L.; Zhang, L.; Zhang, J.; Dong, W.R.; Zhang, L. Forskolin Induces Hyperphosphorylation of Tau Accompanied by Cell Cycle Reactivation in Primary Hippocampal Neurons. Mol. Neurobiol. 2018, 55, 696–706. [Google Scholar] [CrossRef] [PubMed]
- Makhdoumi, P.; Hossini, H.; Ashraf, G.M.; Limoee, M. Molecular Mechanism of Aniline Induced Spleen Toxicity and Neuron Toxicity in Experimental Rat Exposure: A Review. Curr. Neuropharmacol. 2019, 17, 201–213. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, A.G.; Maugeri, G.; Saccone, S.; Federico, C.; Cavallaro, S.; Reglodi, D.; D’agata, V. PACAP Modulates the Autophagy Process in an In Vitro Model of Amyotrophic Lateral Sclerosis. Int. J. Mol. Sci. 2020, 21, 2943. [Google Scholar] [CrossRef]
- Maugeri, G.; D’Amico, A.G.; Amenta, A.; Saccone, S.; Federico, C.; Reibaldi, M.; Russo, A.; Bonfiglio, V.; Avitabile, T.; Longo, A.; et al. Protective effect of PACAP against ultraviolet B radiation-induced human corneal endothelial cell injury. Neuropeptides 2020, 79, 101978. [Google Scholar] [CrossRef]
- Majd, S.; Zarifkar, A.; Rastegar, K.; Takhshid, M.A. Different fibrillar Abeta 1-42 concentrations induce adult hippocampal neurons to reenter various phases of the cell cycle. Brain Res. 2008, 1218, 224–229. [Google Scholar] [CrossRef]
- Majd, S.; Power, J.; Majd, Z. Alzheimer’s Disease and Cancer: When Two Monsters Cannot Be Together. Front. Neurosci. 2019, 13, 155. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, G.; D’Amico, A.G.; Rasà, D.M.; Reitano, R.; Saccone, S.; Federico, C.; Parenti, R.; Magro, G.; D’agata, V. Expression profile of Wilms Tumor 1 (WT1) isoforms in undifferentiated and all-trans retinoic acid differentiated neuroblastoma cells. Genes Cancer 2016, 7, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, H.-L.; Wang, D.-L.; Liu, S.-J.; Wang, J.-Z. A transitory activation of protein kinase-A induces a sustained tau hyperphosphorylation at multiple sites in N2a cells-imply a new mechanism in Alzheimer pathology. J. Neural Transm. 2006, 113, 1487–1497. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, H.; Na, X.L.; Dong, S.Y.; Dong, H.W.; Yu, J.; Jia, L.; Wu, Y.-H. Aniline Induces Oxidative Stress and Apoptosis of Primary Cultured Hepatocytes. Int. J. Environ. Res. Public Health 2016, 13, 1188. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, G.; D’Amico, A.G.; Rasà, D.M.; Saccone, S.; Federico, C.; Cavallaro, S.; D’Agata, V. PACAP and VIP regulate hypoxia-inducible factors in neuroblastoma cells exposed to hypoxia. Neuropeptides 2018, 69, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Souter, S.; Lee, G. Microtubule-associated protein Tau in human prostate cancer cells: Isoforms, phosphorylation and interactions. J. Cell. Bioch. 2009, 108, 555–564. [Google Scholar] [CrossRef]
- Spicakova, T.; O’Brien, M.M.; Duran, G.E.; Sweet-Cordero, A.; Sikic, B.I. Expression and silencing of the microtubule-associated protein Tau in breast cancer cells. Mol. Cancer Ther. 2010, 9, 2970–2981. [Google Scholar] [CrossRef]
- Yang, C.; Ionescu-Tiba, V.; Burns, K.; Gadd, M.; Zukerberg, L.; Spicakova, T.; O’Brien, M.M.; Duran, G.E.; Sweet-Cordero, A.; Sikic, B.I. The role of the cyclin D1-dependent kinases in ErbB2-mediated breast cancer. Am. J. Pathol. 2004, 164, 1031–1038. [Google Scholar] [CrossRef]
- Sherr, C.J. Mammalian G1 cyclins. Cell 1993, 73, 1059–1065. [Google Scholar] [CrossRef]
- Tessari, M.A.; Gostissa, M.; Altamura, S.; Sgarra, R.; Rustighi, A.; Salvagno, C.; Caretti, G.; Imbriano, C.; Mantovani, R.; Del Sal, G.; et al. Transcriptional activation of the cyclin A gene by the architectural transcription factor HMGA2. Mol. Cell Biol. 2003, 23, 9104–9116. [Google Scholar] [CrossRef]
- He, X.; Jiang, W.; Luo, Z.; Qu, T.; Wang, Z.; Liu, N.; Zhang, Y.; Cooper, P.R.; He, W. IFN-Πregulates human dental pulp stem cells behavior via NF-κB and MAPK signaling. Sci. Rep. 2017, 7, 40681. [Google Scholar] [CrossRef] [PubMed]
- Leotta, C.G.; Federico, C.; Brundo, M.V.; Tosi, S.; Saccone, S. HLXB9 gene expression, and nuclear location during in vitro neuronal differentiation in the SK-N-BE neuroblastoma cell line. PLoS ONE 2014, 9, e105481. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Type | Nucleotide Sequence 5′-3′ | Reference |
|---|---|---|---|
| MAPT-F1 | Forward | AACCAGGATGGCTGAGCCCC | [37] |
| MAPT-R4 | Reverse | GTGACCAGCAGCTTCGTCTT | [37] |
| MAPT-F9/10 | Forward | CGGGAAGGTGCAGATAATTAA | [38] |
| MAPT-F9/11 | Forward | AGGCGGGAAGGTGCAAATA | [38] |
| MAPT-R12 | Reverse | CCCAATCTTCGACTGGACTC | [37] |
| CCND1 | Forward | GGATGCTGGAGGTCTGCGA | [39] |
| Reverse | AGAGGCCACGAACATGCAAG | ||
| CCNE1 | Forward | GGTTCCATTTGCCATGGTTA | [40] |
| Reverse | CCCTATTTTGTTCAGACAACATGGC | ||
| CCNA2 | Forward | AGGGAAATGGAGGTTAAATG | [41] |
| Reverse | CACTGACATGGAAGACAGGAACCT | ||
| CCNB1 | Forward | AATGAAATTCAGGTTGTTGCAGGAG | [42] |
| Reverse | CATGGCAGTGACACCAACCAG | ||
| ACTB | Forward | GACGACATGGAGAAAATCTG | Sigma-Aldrich |
| Reverse | ATGATCTGGGTCATCTTCTC | ||
| GAP-43 | Forward | GAGGAAAAATCTTCAGAGACC | [43] |
| Reverse | AACCCTTGAAATCCAGAAAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sturiale, V.; Bruno, F.; Brancato, D.; D’Amico, A.G.; Maugeri, G.; D’Agata, V.; Saccone, S.; Federico, C. Cell Cycle Reactivation, at the Start of Neurodegeneration, Induced by Forskolin and Aniline in Differentiated Neuroblastoma Cells. Int. J. Mol. Sci. 2023, 24, 14373. https://doi.org/10.3390/ijms241814373
Sturiale V, Bruno F, Brancato D, D’Amico AG, Maugeri G, D’Agata V, Saccone S, Federico C. Cell Cycle Reactivation, at the Start of Neurodegeneration, Induced by Forskolin and Aniline in Differentiated Neuroblastoma Cells. International Journal of Molecular Sciences. 2023; 24(18):14373. https://doi.org/10.3390/ijms241814373
Chicago/Turabian StyleSturiale, Valentina, Francesca Bruno, Desiree Brancato, Agata Grazia D’Amico, Grazia Maugeri, Velia D’Agata, Salvatore Saccone, and Concetta Federico. 2023. "Cell Cycle Reactivation, at the Start of Neurodegeneration, Induced by Forskolin and Aniline in Differentiated Neuroblastoma Cells" International Journal of Molecular Sciences 24, no. 18: 14373. https://doi.org/10.3390/ijms241814373
APA StyleSturiale, V., Bruno, F., Brancato, D., D’Amico, A. G., Maugeri, G., D’Agata, V., Saccone, S., & Federico, C. (2023). Cell Cycle Reactivation, at the Start of Neurodegeneration, Induced by Forskolin and Aniline in Differentiated Neuroblastoma Cells. International Journal of Molecular Sciences, 24(18), 14373. https://doi.org/10.3390/ijms241814373