
Therapeutic Effects of Curcumin Derivatives against Obesity and Associated Metabolic Complications: A Review of In Vitro and In Vivo Studies
 ,
,  ,
,
Abstract
:1. Introduction
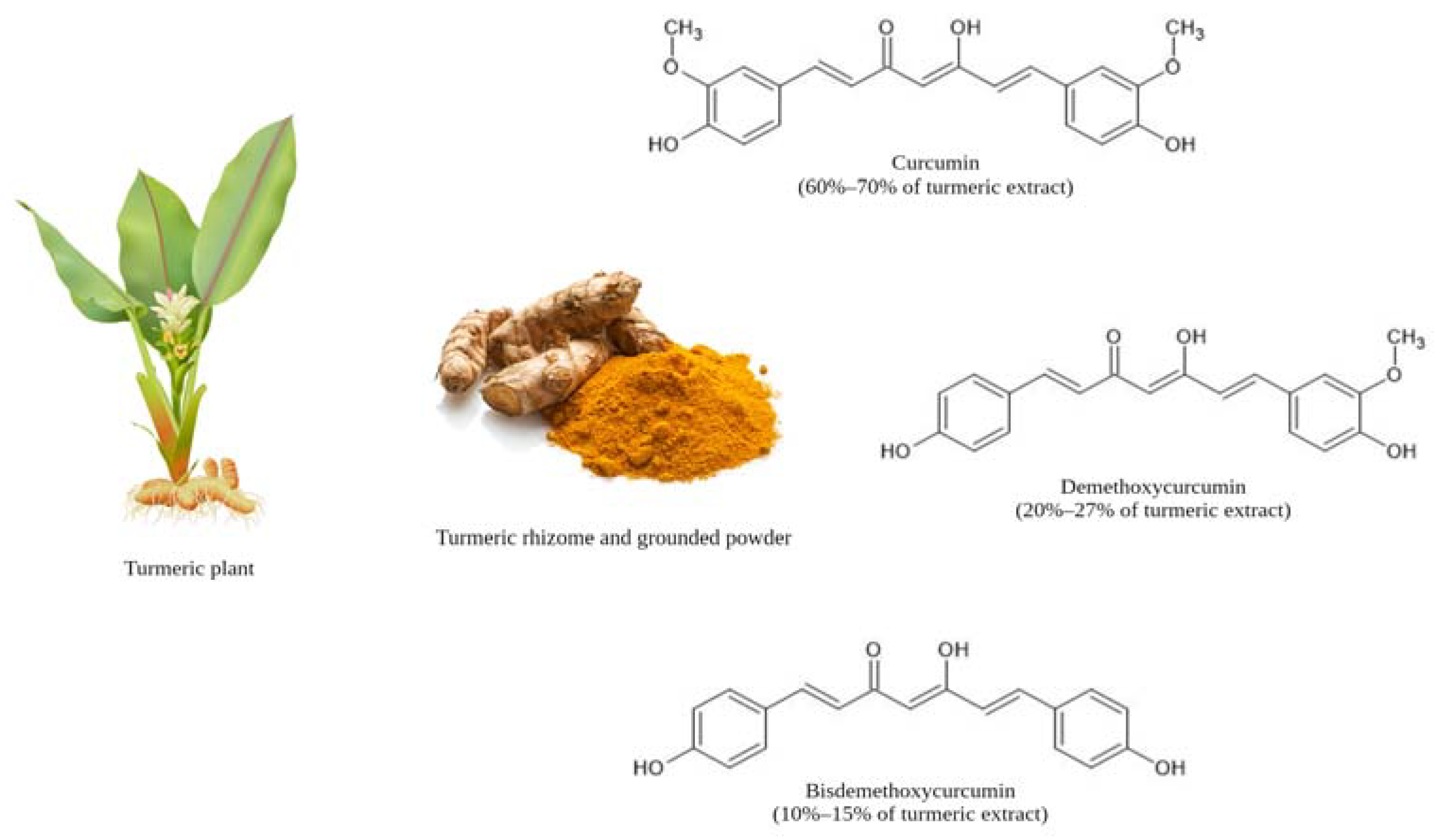
2. Physiochemical, Structural, and Biological Properties of Curcumin
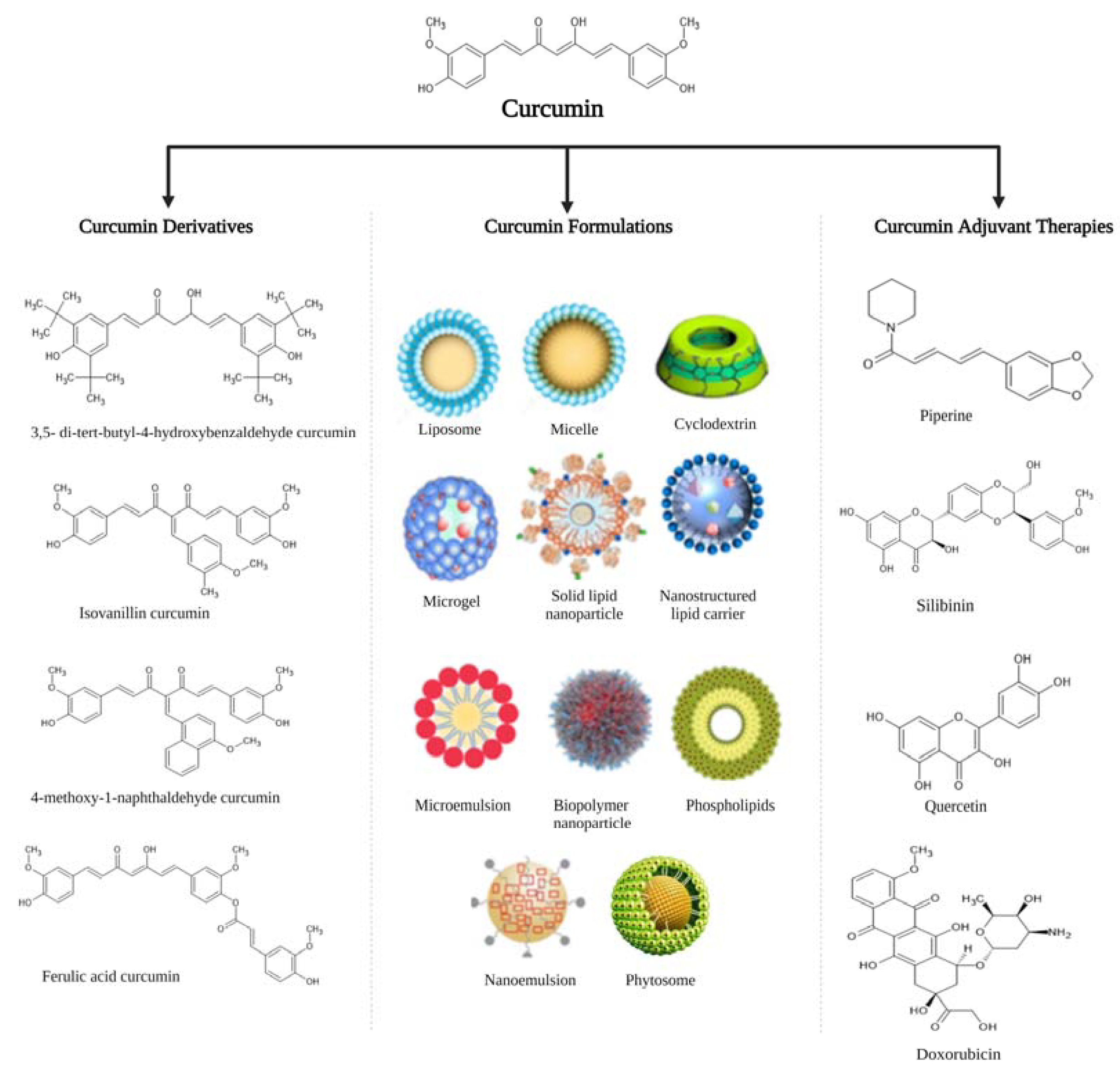
3. The Pharmacokinetic Profile of Curcumin


4. Efficacy of Curcumin Derivatives against Obesity and Associated Metabolic Complications
4.1. In Vitro Studies Reporting on the Therapeutic Effects of Curcumin Derivatives against Obesity and Associated Metabolic Complications

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Curcumin Derivative | Chemical Structure | Aim, Experimental Model, Treatment Dose, and Treatment Period | Outcomes | Reference |
|---|---|---|---|---|
| CNB-001 [4-((1E)-2-(5-(4-hydroxy-3-methoxystyryl-)-1- phenyl-1H-pyrazoyl-3-yl) vinyl)-2-methoxy-phenol] |  | The effect of CNB-001 on insulin resistance was examined. Mouse C2C12 myotubes were co-treated with 400 µM palmitic acid (PA) and 0.1, 1, or 5 μM CNB-001 for 12 h under basal or insulin-stimulated conditions (50 nM insulin, 30 min). | CNB-001 restored PA-induced insulin resistance and impaired insulin signaling by improving insulin-stimulated glucose uptake and increasing insulin-stimulated p-IR, p-IRS-1, and p-AKT expression levels. | [75] |
| Curcumin-3,4-Dichloro Phenyl Pyrazole (CDPP) 4,4′-(1E,1′E)-2,2′-(1-(3,4-dichlorophenyl)-1H-pyrazole-3,5- diyl) bis(ethene-2,1-diyl) bis(2-methoxyphenol) | 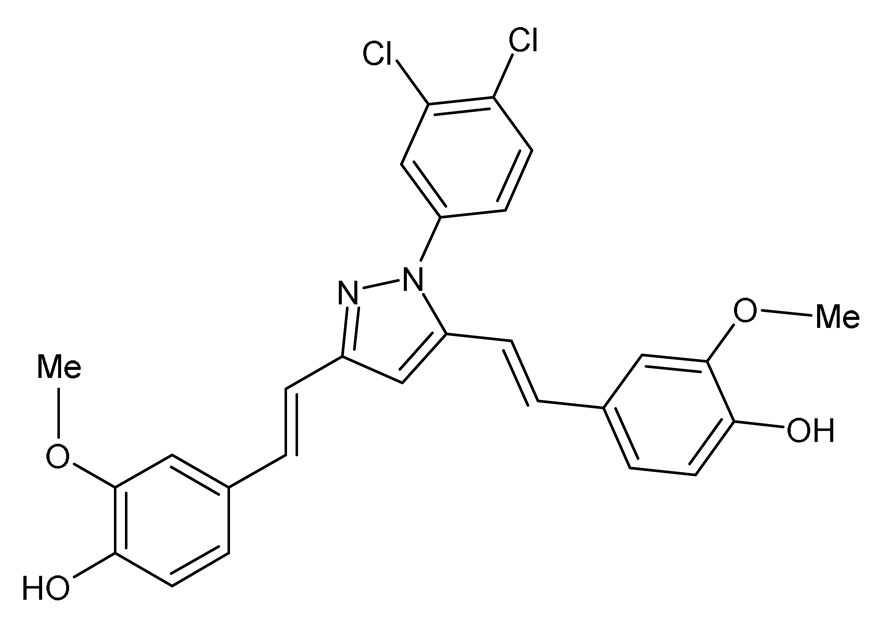 | The effects of CDPP on adipogenesis and lipogenesis were investigated. Mouse 3T3-L1 preadipocytes, mouse C3H10T1/2 mesenchymal stem cells, and human mesenchymal stem cells (hMSC) were treated with 5, 10, and 20 μM CDPP or 20 μM curcumin during differentiation (7 days). Mature 3T3-L1 adipocytes were treated with 20 μM CDPP for 24 h. | CDPP inhibited adipogenesis by dose-dependently decreasing lipid accumulation, reducing C/EBPα, aP2, SREBP1c, PPARγ, and FAS mRNA and protein expression, and downregulating the AKT/mTOR pathway. In 3T3-L1 preadipocytes, CDPP inhibited mitotic clonal expansion by arresting cells at G1-phase and S-phase, decreasing Cyclin D1, Cyclin D3, CDK2, CDK4, CDK6, and p-ERK1/2 expression, and increasing p-p27 expression. CDPP increased oxygen consumption rate and UCP1 and PGC1α expression in 3T3-L1 adipocytes. Compared to curcumin, adipogenesis was significantly inhibited by CDPP. | [78] |
| Curcumin5-8 (CUR5-8) |  | The effect of CUR5-8 on hepatic steatosis was investigated. Mouse AML12 hepatocyte cells were treated with 250 μM PA with or without 10 μM CUR5-8 or curcumin for 24 h. | CUR5-8 decreased PA-induced lipid droplet formation by reducing SREBP1 expression and increasing p-AMPK. CUR5-8 induced autophagy via increased expression of LC3-ATG7, and inhibited apoptosis via increased BCL-2/BAX ratio. The efficacy of CUR5-8 on increased AMPK phosphorylation and autophagy formation and reduced apoptosis was significant compared to curcumin. | [80] |
| Dehydrozingerone (DHZ) Feruloylmethane 1, 7-bis (4-hydroxy-3-methoxyphenyl)-1, 6-heptadiene-3, 5-dione |  | The efficacy of DHZ was evaluated on renal lipotoxicity. Differentiated mouse podocytes were treated with 400 µM PA in the presence or absence of 20 µM DHZ for 24 h. | DHZ inhibited ROS formation, reduced protein expression of NOX-4, NRF2, HO-1, BAX, and SREBP1, and downregulated mRNA expression of Tgf-β and Icam. DHZ increased BCL-2 and podocin protein expression. | [83] |
| The efficacy of DHZ was evaluated on renal lipotoxicity. Mouse glomerular mesangial MES-13 cells were treated with either 30 mM of high glucose (HG) or 250 μM PA in the presence or absence of 20 µM DHZ for 24 h. MES-13 cells were treated with 20 µM DHZ or 20 µM curcumin for 24 h. | DHZ reduced HG-induced ROS production and inflammatory signals by increasing p-AMPK and reducing p-p38 MAPK expression. DHZ decreased PA-induced lipid accumulation, decreased p-p38 MAPK, p-CREB, and COX2 expression, and increased p-AMPK activation. The p-AMPK expression was significantly higher in DHZ-treated cells compared with curcumin. | |||
| The effect of DHZ on the metabolic profile of skeletal muscle was evaluated. Mouse C2C12 myoblasts were treated with 1, 3, 10, 30, or 100 µM DHZ for either 0.1, 0.5, 1, 3, 6, 12, or 24 h. Rat L6 skeletal muscle myotubes were treated with 1, 3, 10, or 30 µM DHZ for 1 h in the presence or absence of 100 nM insulin. | DHZ increased the activation of p-AMPK and p-p38 MAPK signal pathways in C2C12 cells. DHZ upregulated GLUT4, p-IRS-1, and p-AKT expression in C2C12 cells. DHZ induced basal and insulin-stimulated glucose uptake in L6 myotubes. Compared with curcumin, p-AMPK expression was significantly higher in DHZ treated C2C12 cells. | [82] | ||
| C66 (2E,6E)-2,6-bis (2-(tri- fluoromethyl)benzylidene) cyclohexanone | 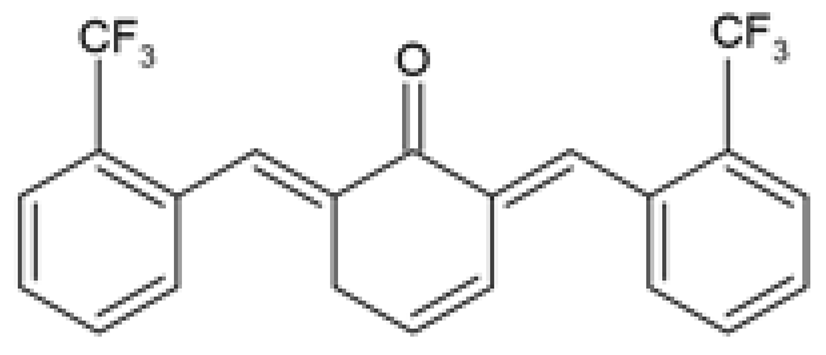 | The protective effect of C66 on renal lipotoxicity and injury was investigated. Mouse glomerular mesangial SV40-MES-13 cells were pre-treated with 2.5, 5, and 10 μM C66 for 2 h followed by treatment with 200 μM PA for 2, 12, or 24 h. | C66 attenuated PA-induced fibrosis by reducing mRNA and protein expression of TGF-β, Col-IV, and MMP-9. C66 ameliorated PA-induced apoptosis by reducing apoptotic positive cells, decreasing BAX and cleaved caspase-3 expression, and increasing BCL-2 expression. C66 protected against PA-induced inflammation by reducing NF-κB/JNK activation via increased IκBα accumulation, reduced p65 phosphorylation and translocation to the nucleus, and reduced JNK phosphorylation. C66 reduced TNF-α, IL-6, and IL-1β secretion levels and mRNA expression. | [88] |
| The protective effect of C66 on cardiomyocyte dysfunction was tested. Rat embryonic heart-derived H9c2 cells were pre-treated with 2.5, 5, and 10 μΜ C66 for 2 h, then treated with 200 μM PA for 24 h. | C66 prevented PA-induced hypertrophy and fibrosis by reducing MyHC, ANP, BNP, TGF-β, Col-I, and MMP-9 mRNA and protein expression. C66 reduced PA-induced apoptosis by decreasing positive apoptotic cells, reducing expression of cleaved caspase-3 and BAX, and increasing BCL-2 expression. C66 ameliorated PA-induced inflammation by reducing NF-κB activation via reduced p65 phosphorylation and translocation to the nucleus and increased IκBα accumulation. C66 also reduced JNK activation and decreased Tnf-α, Il-6, Il-1β, Vcam-1, and Icam-1 mRNA expression. | [89] |
4.2. In Vivo Studies Reporting on the Therapeutic Effects of Curcumin Derivatives on Obesity-Associated Metabolic Complications
| Curcumin Derivative | Chemical Structure | Aim, Experimental Model, Treatment Dose, and Treatment Period | Outcomes | Reference |
|---|---|---|---|---|
| Salicylcurcumin O-hydroxy-substituted analog of curcumin | 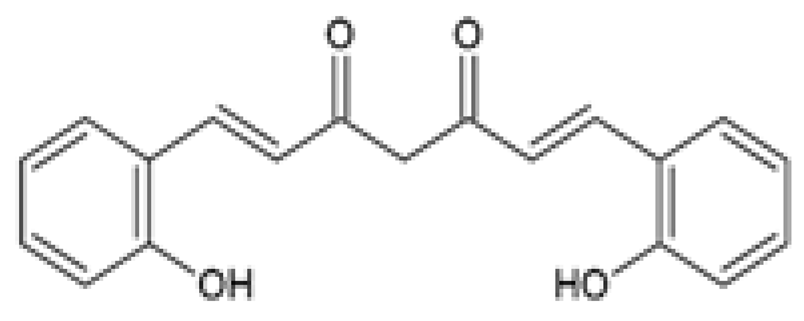 | The antihyperlipidemic effect of salicylcurcumin was investigated. Male albino wistar rats were either orally administered 20% EtOH (alcohol-induced) or fed 15% sunflower oil in a HFD (ΔPUFA-induced) or fed a combination of both in the presence or absence of 80 mg/kg salicylcurcumin or curcumin for 45 days. | Salicylcurcumin reduced TC, TGs, FFAs, and PLs plasma levels. The reduction in TC, TGs, FFAs, and PLs plasma levels was more significant in the salicylcurcumin-treated groups compared to the curcumin-treated groups. | [93] |
| CNB-001 [4-((1E)-2-(5-(4-hydroxy-3-methoxystyryl-)-1- phenyl-1H-pyrazoyl-3-yl) vinyl)-2-methoxy-phenol] | 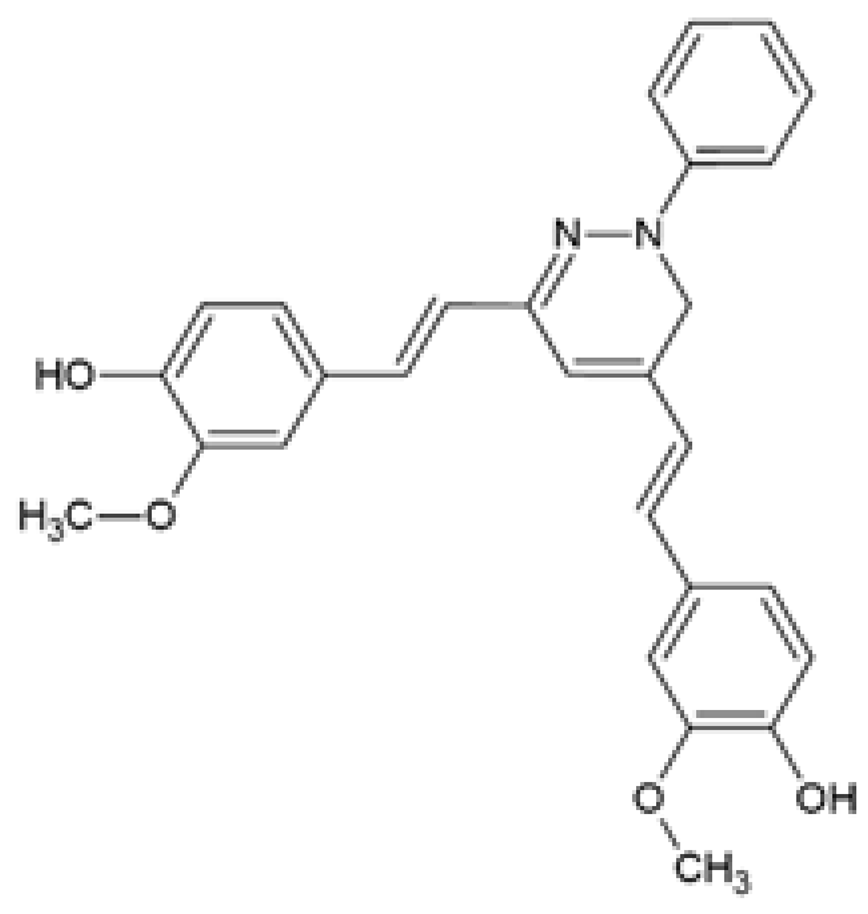 | The effect of CNB-001 on obesity-induced insulin resistance was examined. Male C57BL6 mice were fed a HFD and intraperitoneally injected with CNB-001 (40 mg/kg/day, except weekends) for 22 weeks. | CNB-001 reduced BW gain, adiposity, heart weight, liver weight, serum TGs and IL-6 levels. CNB-001 increased energy expenditure, reduced fasting glucose concentrations, improved glucose tolerance and insulin sensitivity, and increased ex vivo gastrocnemius muscle glucose uptake. CNB-001 attenuated hepatic steatosis by decreasing lipid accumulation and hepatic TG content. CNB-001 restored insulin signaling via upregulation of p-AKT and p-IR expression, reduced ER stress via decreased p-eIF2α and GRP78 expression, and decreased PTP1B expression in gastrocnemius muscles. | [75] |
| Y20 (2E,6E)-2-(2-bromobenzylidene)-6-(2(trifluoromethyl) benzylidene) cyclohexanone | 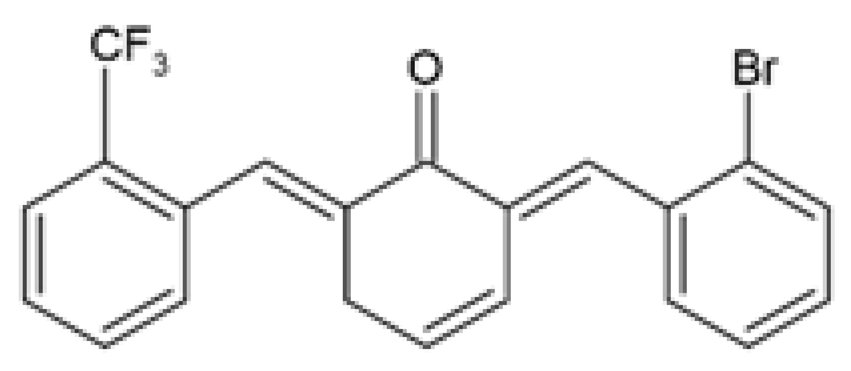 | The protective effects and underlying mechanisms of Y20 on obesity-induced cardiac injury was investigated. HFD-fed male Wistar rats were orally gavaged with 20 mg/kg Y20 or 50 mg/kg curcumin daily for 4 weeks during HFD feeding. | Y20 reduced BW gain and serum TGs levels. Y20 attenuated myocardial inflammation by decreasing the protein and mRNA expression of TNF-α, CD68, IL-6, IL-1β, COX-2, VCAM-1, and ICAM-1, and reversing IκB degradation. Y20 reversed cardiac oxidative stress by reducing superoxide anion production and 3-NT accumulation and increasing the protein and mRNA expression of NRF2 and its downstream antioxidant genes, Ho-1 and Nqo-1. Y20 attenuated cardiac hypertrophy by decreasing cardiomyocyte size and the protein and mRNA expression of cardiac hypertrophic markers, ANP and BNP. Y20 ameliorated cardiac fibrosis by reducing Col-I synthesis and deposition and decreasing the protein and mRNA expression of Col-1, TGF-β, MMP-2, and MMP-9. Y20 reversed myocardial apoptosis by reducing the percentage of apoptotic cardiac cells, increasing BCL-2 expression, and decreasing BAX and cleaved PARP expression levels. Y20 exhibited more anti-inflammatory, antioxidant, anti-fibrosis, and anti-apoptotic efficacy than curcumin. | [94] |
| Curcumin-3,4-Dichloro Phenyl Pyrazole (CDPP) 4,4′-(1E,1′E)-2,2′-(1-(3,4-dichlorophenyl)-1H-pyrazole-3,5- diyl) bis(ethene-2,1-diyl) bis(2-methoxyphenol) |  | The role of CDPP was evaluated on obesity-induced dyslipidemia. Male Syrian golden hamsters were fed a HFD for 4 days then orally administered either 100 mg/kg CDPP or curcumin daily for 7 days during HFD feeding. | CDPP reduced BW gain and serum levels of TGs, TC, LDL-c, ALT, AST, and the TGs/HDL-c ratio. CDPP decreased serum AST and ALT levels. CDPP reduced liver weight, decreased hepatic lipid accumulation, and upregulated hepatic Pparα and Lxrα mRNA expression. CDPP decreased eWAT weight, adipocyte hypertrophy, and reduced the expression of FAS, C/EBPα, aP2 and PPARγ in eWAT. CDPP activated reverse cholesterol transport machinery from adipose tissue to liver by increasing Abca1 and Srb1 and decreasing Abcg1 mRNA expression in eWAT, and increasing hepatic Abcg8, Lcat, Cyp7a1, and Srb1 mRNA expression. The efficacy of CDPP in ameliorating dyslipidemia was more significant compared to curcumin. | [78] |
| Curc-mPEG454 PEGylated curcumin derivative | 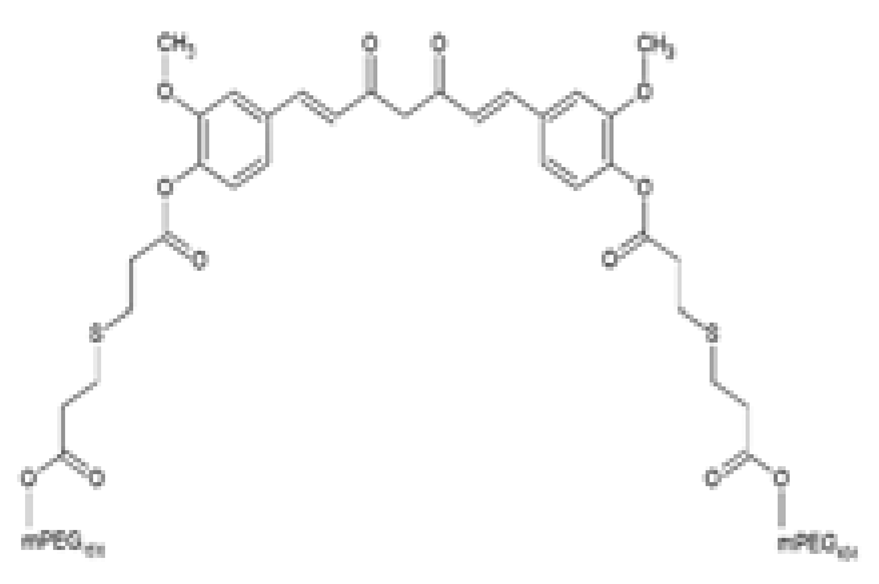 | The effects of Curc-mPEG454 on obesity-associated hepatic steatosis was evaluated. HFD-fed male C57BL/6J mice were intraperitoneally injected every other day with 50 and 100 mg/kg Curc-mPEG454 for 16 weeks during HFD feeding. | Curc-mPEG45 lowered BW gain and serum TGs levels. Curc-mPEG454 reduced hepatic steatosis by decreasing TGs content, decreasing lipid accumulation, and attenuating macrovesicular and microvesicular steatosis. Curc-mPEG454 reduced hepatic PPAR𝛾 and CD36 protein and mRNA expression. Curc-mPEG454 activated hepatic p-CREB. | [98] |
| Curcumin5-8 (CUR5-8) |  | The effect of CUR5-8 on obesity-associated hepatic steatosis and insulin resistance was investigated. Male C57BL/6 mice were fed a HFD chow mixed with either CUR5-8 or curcumin (100 mg/kg/day) for 13 weeks. | CUR5-8 reduced BW gain without suppressing appetite. CUR5 reversed insulin resistance by improving insulin sensitivity, lowering serum insulin levels, and reducing the HOMA-IR index. CUR5-8 protected the liver by decreasing liver weight, reducing fatty liver and TG accumulation, and reducing serum levels of ALT. CUR5-8 regulated hepatic lipid metabolism by decreasing FAS, SREBP1, adipophilin, and PPARγ mRNA and protein expression and increasing p-AMPK expression. CUR5-8 regulated autophagy in the liver by decreasing ATG5 and increasing p62 expression. CUR5-8 reduced apoptosis in the liver by decreasing cleaved caspase-3 staining. Compared to curcumin, the efficacy of CUR5-8 in ameliorating hepatic steatosis and insulin resistance was more significant. | [80] |
| Dehydrozingerone (DHZ) Feruloylmethane 1, 7-bis (4-hydroxy-3-methoxyphenyl)-1, 6-heptadiene-3, 5-dione | 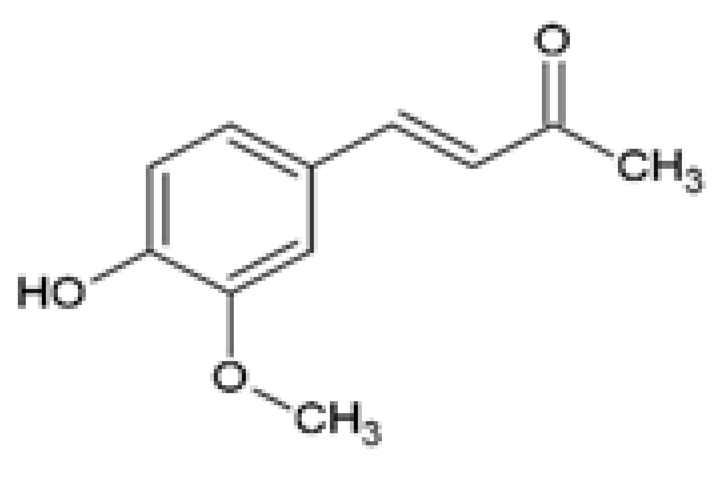 | The efficacy of DHZ on obesity-induced renal lipotoxicity, skeletal muscle metabolism, and the underlying mechanisms was investigated. Male C57BL/6 mice were fed a HFD with or without 100 mg/kg DHZ for 12 weeks. | DHZ reduced BW gain and restored HFD-induced renal damage by decreasing kidney weight, urinary albumin, and urinary ACR. DHZ increased Podocin and decreased Il-1β mRNA expression. DHZ increased nephrin expression and reduced CD68 and ARGINASE 2 expression. DHZ regulated renal lipid metabolism by reducing renal glycerol, FFAs and cholesteryl ester levels, decreasing mRNA and protein expression of SREBP1, SREBP2, FAS, and ACC, and increasing mRNA and protein expression of PPARα, CPT1, and p-AMPK. | [83] |
| DHZ suppressed HFD-induced BW gain, decreased peri-renal fat, epididymal fat, and liver weight, and reduced adipocyte size and lipid accumulation in the liver and epididymal fat. DHZ decreased plasma glucose, insulin, and leptin levels, and improved glucose tolerance. DHZ decreased hepatic mRNA expression of G6Pase, Pepck, and Fetuin. | [82] | |||
| C66 (2E,6E)-2,6-bis (2-(tri- fluoromethyl)benzylidene) cyclohexanone | 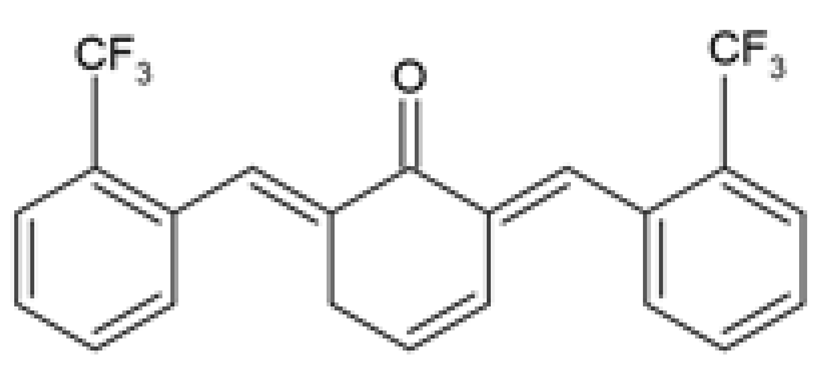 | The protective effect of C66 on obesity-induced renal dysfunction was evaluated. HFD-fed SPF C57BL/6 male mice were orally administered 5, 10, and 20 mg/kg of C66 every 2 days for 11 weeks during HFD feeding. | C66 reduced BW gain, decreased serum TGs and LDL-c levels, and increased serum HDL-c levels. C66 decreased kidney weight, serum BUN and CRE levels, and urine CRE and albumin levels. C66 reduced fibrosis by reducing collagen and ECM deposition and decreasing the mRNA and protein expression of Col-IV, TGF-β, and MMP-9. C66 reversed apoptosis by reducing TUNEL positive stained cells, decreasing cleaved caspase-3 and BAX expression, and increasing BCL-2 expression. C66 attenuated chronic inflammation by reducing macrophage infiltration, decreasing Tnf-α, Il-6, and Il-1β mRNA expression levels, and inhibiting NF-κB/JNK activation via increased IκBα accumulation, and reduced expression of p-p65 and p-JNK. | [88] |
| The protective effect of C66 on obesity-induced cardiomyopathy through inhibition of JNK-mediated inflammation was investigated. Male C57BL/6 mice were fed HFD for 11 weeks followed by oral administration with 5, 10, and 20 mg/kg of C66 every 2 days for a further 7 weeks during HFD feeding. | C66 reduced BW gain and decreased serum TGs, LDL-c, and cholesterol levels, and increased HDL-c serum levels. C66 restored cardiac function by improving EF% and FS%. C66 decreased myocardial hypertrophy by reducing heart weight to tibia length ratio, cardiomyocyte longitudinal and transverse section area, and the circulating and mRNA expression levels of CK-MB, ANP, and BNP. C66 attenuated cardiac fibrosis by decreasing collagen deposition and reducing MyHc, TGF-β, and Col-I protein and mRNA expression levels. C66 suppressed myocardial apoptosis by reducing TUNEL positive stained cells, decreasing cleaved caspase-3 and BAX expression, and increasing BCL-2 expression. C66 ameliorated cardiac inflammation by decreasing macrophage infiltration, reducing Tnf-α, Il-6, and Il-1β mRNA expression levels, and reversing NF-κB/JNK activation via increased IκBα accumulation, and reduced expression of p-p65 and p-JNK. | [89] |
5. Summary and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, Regional, and National Prevalence of Overweight and Obesity in Children and Adults during 1980–2013: A Systematic Analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef] [PubMed]
- Di Cesare, M.; Bentham, J.; Stevens, G.A.; Zhou, B.; Danaei, G.; Lu, Y.; Bixby, H.; Cowan, M.J.; Riley, L.M.; Hajifathalian, K.; et al. Trends in Adult Body-Mass Index in 200 Countries from 1975 to 2014: A Pooled Analysis of 1698 Population-Based Measurement Studies with 19.2 Million Participants. Lancet 2016, 387, 1377–1396. [Google Scholar] [CrossRef]
- Al-Sulaiti, H.; Diboun, I.; Agha, M.V.; Mohamed, F.F.S.; Atkin, S.; Dömling, A.S.; Elrayess, M.A.; Mazloum, N.A. Metabolic Signature of Obesity-Associated Insulin Resistance and Type 2 Diabetes. J. Transl. Med. 2019, 17, 348. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Ballantyne, C.M. Metabolic Inflammation and Insulin Resistance in Obesity. Circ. Res. 2020, 126, 1549–1564. [Google Scholar] [CrossRef]
- Yazıcı, D.; Sezer, H. Insulin Resistance, Obesity and Lipotoxicity. Adv. Exp. Med. Biol. 2017, 960, 277–304. [Google Scholar] [CrossRef]
- Vekic, J.; Zeljkovic, A.; Stefanovic, A.; Jelic-Ivanovic, Z.; Spasojevic-Kalimanovska, V. Obesity and Dyslipidemia. Metabolism 2019, 92, 71–81. [Google Scholar] [CrossRef]
- Polyzos, S.A.; Kountouras, J.; Mantzoros, C.S. Obesity and Nonalcoholic Fatty Liver Disease: From Pathophysiology to Therapeutics. Metabolism 2019, 92, 82–97. [Google Scholar] [CrossRef]
- Leitner, D.R.; Frühbeck, G.; Yumuk, V.; Schindler, K.; Micic, D.; Woodward, E.; Toplak, H. Obesity and Type 2 Diabetes: Two Diseases with a Need for Combined Treatment Strategies—EASO Can Lead the Way. Obes. Facts 2017, 10, 483–492. [Google Scholar] [CrossRef]
- Powell-Wiley, T.M.; Poirier, P.; Burke, L.E.; Després, J.-P.; Gordon-Larsen, P.; Lavie, C.J.; Lear, S.A.; Ndumele, C.E.; Neeland, I.J.; Sanders, P.; et al. Obesity and Cardiovascular Disease: A Scientific Statement from the American Heart Association. Circulation 2021, 143, e984–e1010. [Google Scholar] [CrossRef]
- Davoodi, S.H.; Malek-Shahabi, T.; Malekshahi-Moghadam, A.; Shahbazi, R.; Esmaeili, S. Obesity as an Important Risk Factor for Certain Types of Cancer. Iran. J. Cancer Prev. 2013, 6, 186–194. [Google Scholar]
- Löffler, M.C.; Betz, M.J.; Blondin, D.P.; Augustin, R.; Sharma, A.K.; Tseng, Y.-H.; Scheele, C.; Zimdahl, H.; Mark, M.; Hennige, A.M.; et al. Challenges in Tackling Energy Expenditure as Obesity Therapy: From Preclinical Models to Clinical Application. Mol. Metab. 2021, 51, 101237. [Google Scholar] [CrossRef] [PubMed]
- Lagerros, Y.T.; Rössner, S. Obesity Management: What Brings Success? Therap. Adv. Gastroenterol. 2013, 6, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Tak, Y.J.; Lee, S.Y. Long-Term Efficacy and Safety of Anti-Obesity Treatment: Where Do We Stand? Curr. Obes. Rep. 2021, 10, 14–30. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.D.; Blüher, M.; Tschöp, M.H.; DiMarchi, R.D. Anti-Obesity Drug Discovery: Advances and Challenges. Nat. Rev. Drug Discov. 2022, 21, 201–223. [Google Scholar] [CrossRef]
- Lim, R.; Beekley, A.; Johnson, D.C.; Davis, K.A. Early and Late Complications of Bariatric Operation. Trauma Surg. Acute Care Open 2018, 3, e000219. [Google Scholar] [CrossRef]
- Soleimani, V.; Sahebkar, A.; Hosseinzadeh, H. Turmeric (Curcuma Longa) and Its Major Constituent (Curcumin) as Nontoxic and Safe Substances: Review. Phytother. Res. 2018, 32, 985–995. [Google Scholar] [CrossRef]
- Menon, V.P.; Sudheer, A.R. Antioxidant and Anti-Inflammatory Properties of Curcumin. Adv. Exp. Med. Biol. 2007, 595, 105–125. [Google Scholar] [CrossRef]
- Khan, H.; Ullah, H.; Nabavi, S.M. Mechanistic Insights of Hepatoprotective Effects of Curcumin: Therapeutic Updates and Future Prospects. Food Chem. Toxicol. 2019, 124, 182–191. [Google Scholar] [CrossRef]
- Zhang, Q.; Wu, L. In Vitro and In Vivo Cardioprotective Effects of Curcumin against Doxorubicin-Induced Cardiotoxicity: A Systematic Review. J. Oncol. 2022, 2022, 7277562. [Google Scholar] [CrossRef]
- Xia, Z.-H.; Zhang, S.-Y.; Chen, Y.-S.; Li, K.; Chen, W.-B.; Liu, Y.-Q. Curcumin Anti-Diabetic Effect Mainly Correlates with Its Anti-Apoptotic Actions and PI3K/Akt Signal Pathway Regulation in the Liver. Food Chem. Toxicol. 2020, 146, 111803. [Google Scholar] [CrossRef]
- Chen, Y.; Wu, R.; Chen, W.; Liu, Y.; Liao, X.; Zeng, B.; Guo, G.; Lou, F.; Xiang, Y.; Wang, Y.; et al. Curcumin Prevents Obesity by Targeting TRAF4-induced Ubiquitylation in m 6 A-dependent Manner. EMBO Rep. 2021, 22, e52146. [Google Scholar] [CrossRef]
- Lopresti, A.L. The Problem of Curcumin and Its Bioavailability: Could Its Gastrointestinal Influence Contribute to Its Overall Health-Enhancing Effects? Adv. Nutr. 2018, 9, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Dei Cas, M.; Ghidoni, R. Dietary Curcumin: Correlation between Bioavailability and Health Potential. Nutrients 2019, 11, 2147. [Google Scholar] [CrossRef] [PubMed]
- Racz, L.Z.; Racz, C.P.; Pop, L.-C.; Tomoaia, G.; Mocanu, A.; Barbu, I.; Sárközi, M.; Roman, I.; Avram, A.; Tomoaia-Cotisel, M.; et al. Strategies for Improving Bioavailability, Bioactivity, and Physical-Chemical Behavior of Curcumin. Molecules 2022, 27, 6854. [Google Scholar] [CrossRef] [PubMed]
- Dayton, A.; Selvendiran, K.; Kuppusamy, M.L.; Rivera, B.K.; Meduru, S.; Kálai, T.; Hideg, K.; Kuppusamy, P. Cellular Uptake, Retention and Bioabsorption of HO-3867, a Fluorinated Curcumin Analog with Potential Antitumor Properties. Cancer Biol. Ther. 2010, 10, 1027–1032. [Google Scholar] [CrossRef]
- Zeng, Z.; Shen, Z.L.; Zhai, S.; Xu, J.L.; Liang, H.; Shen, Q.; Li, Q.Y. Transport of Curcumin Derivatives in Caco-2 Cell Monolayers. Eur. J. Pharm. Biopharm. 2017, 117, 123–131. [Google Scholar] [CrossRef]
- Noureddin, S.A.; El-Shishtawy, R.M.; Al-Footy, K.O. Curcumin Analogues and Their Hybrid Molecules as Multifunctional Drugs. Eur. J. Med. Chem. 2019, 182, 111631. [Google Scholar] [CrossRef] [PubMed]
- Ooko, E.; Alsalim, T.; Saeed, B.; Saeed, M.E.M.; Kadioglu, O.; Abbo, H.S.; Titinchi, S.J.J.; Efferth, T. Modulation of P-Glycoprotein Activity by Novel Synthetic Curcumin Derivatives in Sensitive and Multidrug-Resistant T-Cell Acute Lymphoblastic Leukemia Cell Lines. Toxicol. Appl. Pharmacol. 2016, 305, 216–233. [Google Scholar] [CrossRef]
- Abdel Aziz, M.T.; El-Asmar, M.F.; El-Ibrashy, I.N.; Rezq, A.M.; Al-Malki, A.L.; Wassef, M.A.; Fouad, H.H.; Ahmed, H.H.; Taha, F.M.; Hassouna, A.A.; et al. Effect of Novel Water Soluble Curcumin Derivative on Experimental Type-1 Diabetes Mellitus (Short Term Study). Diabetol. Metab. Syndr. 2012, 4, 30. [Google Scholar] [CrossRef]
- Cheng, F.; Chen, Y.; Zhan, Z.; Liu, Y.; Hu, P.; Ren, H.; Tang, H.; Peng, M. Curc-MPEG454, a PEGylated Curcumin Derivative, Improves Anti-Inflammatory and Antioxidant Activities: A Comparative Study. Inflammation 2018, 41, 579–594. [Google Scholar] [CrossRef]
- Wang, Z.S.; Chen, L.Z.; Liu, X.H.; Chen, F.H. Diarylpentadienone Derivatives (Curcumin Analogues): Synthesis and Anti-Inflammatory Activity. Bioorganic. Med. Chem. Lett. 2017, 27, 1803–1807. [Google Scholar] [CrossRef] [PubMed]
- Vogel, A.; Pelletier, J. Examen Chimique de La Racine de Curcuma. J. Pharm. 1815, 1, 289–300. [Google Scholar]
- Vogel, E. Die Bibliothek Der Benediktinerabtei Sponheim. Serapeum 1842, 3, 312–328. [Google Scholar]
- Lampe, V.; Milobedzka, J.; Kostanecki, S.V. Zur Kenntnis Des Curcumins. Berichte Dtsch. Chem. Ges. 1910, 43, 2163–2170. [Google Scholar]
- Lampe, V.; Milobedzka, J. Studien Über Curcumin. Berichte Dtsch. Chem. Ges. 1913, 46, 2235–2240. [Google Scholar] [CrossRef]
- Priyadarsini, K.I. The Chemistry of Curcumin: From Extraction to Therapeutic Agent. Molecules 2014, 19, 20091–20112. [Google Scholar] [CrossRef]
- Kawano, S.I.; Inohana, Y.; Hashi, Y.; Lin, J.M. Analysis of Keto-Enol Tautomers of Curcumin by Liquid Chromatography/Mass Spectrometry. Chin. Chem. Lett. 2013, 24, 685–687. [Google Scholar] [CrossRef]
- Kazakova, O.; Lipkovska, N.; Barvinchenko, V. Keto-Enol Tautomerism of Curcumin in the Preparation of Nanobiocomposites with Fumed Silica. Spectrochim. Acta—Part A Mol. Biomol. Spectrosc. 2022, 277, 121287. [Google Scholar] [CrossRef]
- Nelson, K.M.; Dahlin, J.L.; Bisson, J.; Graham, J.; Pauli, G.F.; Walters, M.A. The Essential Medicinal Chemistry of Curcumin. J. Med. Chem. 2017, 60, 1620–1637. [Google Scholar] [CrossRef]
- Eigner, D.; Scholz, D. Ferula Asa-Foetida and Curcuma Longa in Traditional Medical Treatment and Diet in Nepal. J. Ethnopharmacol. 1999, 67, 1–6. [Google Scholar] [CrossRef]
- EL-Kenawy, A.E.-M.; Hassan, S.M.A.; Mohamed, A.M.M.; Mohammed, H.M.A. Tumeric or Curcuma Longa Linn. In Nonvitamin and Nonmineral Nutritional Supplements; Nabavi, S.M., Silva, A.S., Eds.; Academic Press: Cambridge, MA, USA, 2019; ISBN 978-0-12-812491-8. [Google Scholar]
- Kasprzak-Drozd, K.; Oniszczuk, T.; Gancarz, M.; Kondracka, A.; Rusinek, R.; Oniszczuk, A. Curcumin and Weight Loss: Does It Work? Int. J. Mol. Sci. 2022, 23, 639. [Google Scholar] [CrossRef] [PubMed]
- Maithilikarpagaselvi, N.; Sridhar, M.G.; Swaminathan, R.P.; Sripradha, R. Preventive Effect of Curcumin on Inflammation, Oxidative Stress and Insulin Resistance in High-Fat Fed Obese Rats. J. Complement. Integr. Med. 2016, 13, 137–143. [Google Scholar] [CrossRef]
- Mokgalaboni, K.; Ntamo, Y.; Ziqubu, K.; Nyambuya, T.M.; Nkambule, B.B.; Mazibuko-Mbeje, S.E.; Gabuza, K.B.; Chellan, N.; Tiano, L.; Dludla, P.V. Curcumin Supplementation Improves Biomarkers of Oxidative Stress and Inflammation in Conditions of Obesity, Type 2 Diabetes and NAFLD: Updating the Status of Clinical Evidence. Food Funct. 2021, 12, 12235–12249. [Google Scholar] [CrossRef]
- Campbell, M.S.; Ouyang, A.; Krishnakumar, I.M.; Charnigo, R.J.; Westgate, P.M.; Fleenor, B.S. Influence of Enhanced Bioavailable Curcumin on Obesity-Associated Cardiovascular Disease Risk Factors and Arterial Function: A Double-Blinded, Randomized, Controlled Trial. Nutrition 2019, 62, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Abdulmalek, S.; Eldala, A.; Awad, D.; Balbaa, M. Ameliorative Effect of Curcumin and Zinc Oxide Nanoparticles on Multiple Mechanisms in Obese Rats with Induced Type 2 Diabetes. Sci. Rep. 2021, 11, 20677. [Google Scholar] [CrossRef] [PubMed]
- Shehzad, A.; Khan, S.; Sup Lee, Y. Curcumin Molecular Targets in Obesity and Obesity-Related Cancers. Futur. Oncol. 2012, 8, 179–190. [Google Scholar] [CrossRef]
- Hewlings, S.J.; Kalman, D.S. Curcumin: A Review of Its Effects on Human Health. Foods 2017, 6, 92. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Rayess, Y.E.; Rizk, A.A.; Sadaka, C.; Zgheib, R.; Zam, W.; Sestito, S.; Rapposelli, S.; Neffe-Skocińska, K.; Zielińska, D.; et al. Turmeric and Its Major Compound Curcumin on Health: Bioactive Effects and Safety Profiles for Food, Pharmaceutical, Biotechnological and Medicinal Applications. Front. Pharmacol. 2020, 11, 01021. [Google Scholar] [CrossRef]
- Tønnesen, H.H.; Másson, M.; Loftsson, T. Studies of Curcumin and Curcuminoids. XXVII. Cyclodextrin Complexation: Solubility, Chemical and Photochemical Stability. Int. J. Pharm. 2002, 244, 127–135. [Google Scholar] [CrossRef]
- Ma, Z.; Wang, N.; He, H.; Tang, X. Pharmaceutical Strategies of Improving Oral Systemic Bioavailability of Curcumin for Clinical Application. J. Control. Release 2019, 316, 359–380. [Google Scholar] [CrossRef]
- Parsamanesh, N.; Moossavi, M.; Bahrami, A.; Butler, A.E.; Sahebkar, A. Therapeutic Potential of Curcumin in Diabetic Complications. Pharmacol. Res. 2018, 136, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Maiti, K.; Mukherjee, K.; Gantait, A.; Saha, B.P.; Mukherjee, P.K. Curcumin-Phospholipid Complex: Preparation, Therapeutic Evaluation and Pharmacokinetic Study in Rats. Int. J. Pharm. 2007, 330, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.Y.; Lin, L.C.; Tseng, T.Y.; Wang, S.C.; Tsai, T.H. Oral Bioavailability of Curcumin in Rat and the Herbal Analysis from Curcuma Longa by LC-MS/MS. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2007, 853, 183–189. [Google Scholar] [CrossRef]
- Lao, C.D.; Ruffin, M.T., IV; Normolle, D.; Heath, D.D.; Murray, S.I.; Bailey, J.M.; Boggs, M.E.; Crowell, J.; Rock, C.L.; Brenner, D.E. Dose Escalation of a Curcuminoid Formulation. BMC Complement. Altern. Med. 2006, 6, 10. [Google Scholar] [CrossRef] [PubMed]
- Vareed, S.K.; Kakarala, M.; Ruffin, M.T.; Crowell, J.A.; Normolle, D.P.; Djuric, Z.; Brenner, D.E. Pharmacokinetics of Curcumin Conjugate Metabolites in Healthy Human Subjects. Cancer Epidemiol. Biomark. Prev. 2008, 17, 1411–1417. [Google Scholar] [CrossRef] [PubMed]
- Ireson, C.R.; Jones, D.J.L.; Boocock, D.J.; Farmer, P.B.; Gescher, A.J.; Orr, S.; Coughtrie, M.W.H.; Williams, M.L.; Steward, W.P. Metabolism of the Cancer Chemopreventive Agent Curcumin in Human and Rat Intestine. Cancer Epidemiol. Biomark. Prev. 2002, 11, 105–111. [Google Scholar]
- Pandey, A.; Chaturvedi, M.; Mishra, S.; Kumar, P.; Somvanshi, P.; Chaturvedi, R. Reductive Metabolites of Curcumin and Their Therapeutic Effects. Heliyon 2020, 6, e05469. [Google Scholar] [CrossRef]
- Pan, M.H.; Huang, T.M.; Lin, J.K. Biotransformation of Curcumin through Reduction and Glucuronidation in Mice. Drug Metab. Dispos. 1999, 27, 486–494. [Google Scholar]
- Hassaninasab, A.; Hashimoto, Y.; Tomita-Yokotani, K.; Kobayashi, M. Discovery of the Curcumin Metabolic Pathway Involving a Unique Enzyme in an Intestinal Microorganism. Proc. Natl. Acad. Sci. USA 2011, 108, 6615–6620. [Google Scholar] [CrossRef]
- Luca, S.V.; Macovei, I.; Bujor, A.; Miron, A.; Skalicka-Woźniak, K.; Aprotosoaie, A.C.; Trifan, A. Bioactivity of Dietary Polyphenols: The Role of Metabolites. Crit. Rev. Food Sci. Nutr. 2020, 60, 626–659. [Google Scholar] [CrossRef]
- Wang, J.; Yu, X.; Zhang, L.; Wang, L.; Peng, Z.; Chen, Y. The Pharmacokinetics and Tissue Distribution of Curcumin and Its Metabolites in Mice. Biomed. Chromatogr. 2018, 32, e4267. [Google Scholar] [CrossRef] [PubMed]
- Stohs, S.J.; Chen, O.; Ray, S.D.; Ji, J.; Bucci, L.R.; Preuss, H.G. Highly Bioavailable Forms of Curcumin and Promising Avenues for Curcumin-Based Research and Application: A Review. Molecules 2020, 25, 1397. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.E.; Ngai, S.C.; Chan, K.G.; Lee, L.H.; Goh, B.H.; Chuah, L.H. Curcumin Nanoformulations for Colorectal Cancer: A Review. Front. Pharmacol. 2019, 10, 152. [Google Scholar] [CrossRef] [PubMed]
- Fança-Berthon, P.; Tenon, M.; Bouter-Banon, S.L.; Manfré, A.; Maudet, C.; Dion, A.; Chevallier, H.; Laval, J.; Van Breemen, R.B. Pharmacokinetics of a Single Dose of Turmeric Curcuminoids Depends on Formulation: Results of a Human Crossover Study. J. Nutr. 2021, 151, 1802–1816. [Google Scholar] [CrossRef] [PubMed]
- Shoba, G.; Joy, D.; Joseph, T.; Majeed, M.; Rajendran, R.; Srinivas, P.S.S.R. Influence of Piperine on the Pharmacokinetics of Curcumin in Animals and Human Volunteers. Planta Med. 1998, 64, 353–356. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, C.; Wang, W.; Liu, J.; Liu, Q.; Huang, F.; Chu, L.; Gao, H.; Li, C.; Kong, D.; et al. Co-Delivery of Doxorubicin and Curcumin by PH-Sensitive Prodrug Nanoparticle for Combination Therapy of Cancer. Sci. Rep. 2016, 6, 21225. [Google Scholar] [CrossRef]
- Moorthi, C.; Kathiresan, K. Curcumin–Piperine/Curcumin–Quercetin/Curcumin–Silibinin Dual Drug-Loaded Nanoparticulate Combination Therapy: A Novel Approach to Target and Treat Multidrug-Resistant Cancers. J. Med. Hypotheses Ideas 2013, 7, 15–20. [Google Scholar] [CrossRef]
- Rivera-Mancía, S.; Lozada-García, M.C.; Pedraza-Chaverri, J. Experimental Evidence for Curcumin and Its Analogs for Management of Diabetes Mellitus and Its Associated Complications. Eur. J. Pharmacol. 2015, 756, 30–37. [Google Scholar] [CrossRef]
- Oliveira, S.; Monteiro-Alfredo, T.; Silva, S.; Matafome, P. Curcumin Derivatives for Type 2 Diabetes Management and Prevention of Complications. Arch. Pharm. Res. 2020, 43, 567–581. [Google Scholar] [CrossRef]
- Idoudi, S.; Bedhiafi, T.; Hijji, Y.M.; Billa, N. Curcumin and Derivatives in Nanoformulations with Therapeutic Potential on Colorectal Cancer. AAPS PharmSciTech 2022, 23, 115. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Deb, L.; Prasad, S. Curcumin Differs from Tetrahydrocurcumin for Molecular Targets, Signaling Pathways and Cellular Responses. Molecules 2014, 20, 185–205. [Google Scholar] [CrossRef] [PubMed]
- Pastorelli, D.; Fabricio, A.S.C.; Giovanis, P.; D’Ippolito, S.; Fiduccia, P.; Soldà, C.; Buda, A.; Sperti, C.; Bardini, R.; Da Dalt, G.; et al. Phytosome Complex of Curcumin as Complementary Therapy of Advanced Pancreatic Cancer Improves Safety and Efficacy of Gemcitabine: Results of a Prospective Phase II Trial. Pharmacol. Res. 2018, 132, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Law, S.K.; Lo, C.; Han, J.; Yang, F.; Leung, A.; Chuanshan, X. Design, Synthesis and Characterization of Novel Curcumin Derivatives. Nat. Prod. Chem. Res. 2020, 8, 367. [Google Scholar] [CrossRef]
- Panzhinskiy, E.; Hua, Y.; Lapchak, P.A.; Topchiy, E.; Lehmann, T.E.; Ren, J.; Nair, S. Novel Curcumin Derivative CNB-001 Mitigates Obesity-Associated Insulin Resistance. J. Pharmacol. Exp. Ther. 2014, 349, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Dargusch, R.; Maher, P.; Schubert, D. A Broadly Neuroprotective Derivative of Curcumin. J. Neurochem. 2008, 105, 1336–1345. [Google Scholar] [CrossRef]
- Selvam, C.; Jachak, S.M.; Thilagavathi, R.; Chakraborti, A.K. Design, Synthesis, Biological Evaluation and Molecular Docking of Curcumin Analogues as Antioxidant, Cyclooxygenase Inhibitory and Anti-Inflammatory Agents. Bioorg. Med. Chem. Lett. 2005, 15, 1793–1797. [Google Scholar] [CrossRef]
- Gupta, A.; Singh, V.K.; Kumar, D.; Yadav, P.; Kumar, S.; Beg, M.; Shankar, K.; Varshney, S.; Rajan, S.; Srivastava, A.; et al. Curcumin-3,4-Dichloro Phenyl Pyrazole (CDPP) Overcomes Curcumin’s Low Bioavailability, Inhibits Adipogenesis and Ameliorates Dyslipidemia by Activating Reverse Cholesterol Transport. Metabolism 2017, 73, 109–124. [Google Scholar] [CrossRef]
- Woo, H.B.; Shin, W.-S.; Lee, S.; Ahn, C.M. Synthesis of Novel Curcumin Mimics with Asymmetrical Units and Their Anti-Angiogenic Activity. Bioorg. Med. Chem. Lett. 2005, 15, 3782–3786. [Google Scholar] [CrossRef]
- Lee, E.S.; Kwon, M.H.; Kim, H.M.; Woo, H.B.; Ahn, C.M.; Chung, C.H. Curcumin Analog CUR5–8 Ameliorates Nonalcoholic Fatty Liver Disease in Mice with High-Fat Diet-Induced Obesity. Metabolism 2020, 103, 154015. [Google Scholar] [CrossRef]
- Kubra, I.R.; Bettadaiah, B.K.; Murthy, P.S.; Rao, L.J.M. Structure-Function Activity of Dehydrozingerone and Its Derivatives as Antioxidant and Antimicrobial Compounds. J. Food Sci. Technol. 2014, 51, 245–255. [Google Scholar] [CrossRef]
- Kim, S.J.; Kim, H.M.; Lee, E.S.; Kim, N.; Lee, J.O.; Lee, H.J.; Park, N.Y.; Jo, J.Y.; Ham, B.Y.; Han, S.H.; et al. Dehydrozingerone Exerts Beneficial Metabolic Effects in High-Fat Diet-Induced Obese Mice via AMPK Activation in Skeletal Muscle. J. Cell. Mol. Med. 2015, 19, 620–629. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.S.; Kang, J.S.; Kim, H.M.; Kim, S.J.; Kim, N.; Lee, J.O.; Kim, H.S.; Lee, E.Y.; Chung, C.H. Dehydrozingerone Inhibits Renal Lipotoxicity in High-Fat Diet–Induced Obese Mice. J. Cell. Mol. Med. 2021, 25, 8725–8733. [Google Scholar] [CrossRef]
- Pan, Y.; Wang, Y.; Cai, L.; Cai, Y.; Hu, J.; Yu, C.; Li, J.; Feng, Z.; Yang, S.; Li, X.; et al. Inhibition of High Glucose-Induced Inflammatory Response and Macrophage Infiltration by a Novel Curcumin Derivative Prevents Renal Injury in Diabetic Rats. Br. J. Pharmacol. 2012, 166, 1169–1182. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Zhang, X.; Wang, Y.; Cai, L.; Ren, L.; Tang, L.; Wang, J.; Zhao, Y.; Wang, Y.; Liu, Q.; et al. Targeting JNK by a New Curcumin Analog to Inhibit NF-KB-Mediated Expression of Cell Adhesion Molecules Attenuates Renal Macrophage Infiltration and Injury in Diabetic Mice. PLoS ONE 2013, 8, e79084. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Huang, Y.; Wang, Z.; Fang, Q.; Sun, Y.; Tong, C.; Peng, K.; Wang, Y.; Miao, L.; Cai, L.; et al. Inhibition of MAPK-Mediated ACE Expression by Compound C66 Prevents STZ-Induced Diabetic Nephropathy. J. Cell. Mol. Med. 2014, 18, 231–241. [Google Scholar] [CrossRef]
- Sun, X.; Liu, Y.; Li, C.; Wang, X.; Zhu, R.; Liu, C.; Liu, H.; Wang, L.; Ma, R.; Fu, M.; et al. Recent Advances of Curcumin in the Prevention and Treatment of Renal Fibrosis. Biomed Res. Int. 2017, 2017, 2418671. [Google Scholar] [CrossRef]
- Ye, L.; Hu, X.; Hu, X.; Yin, S.; Chen, J.; He, H.; Hong, S.; Yang, B.; Singh, K.K.K.; Feng, J.; et al. Curcumin Analogue C66 Attenuates Obesity-Induced Renal Injury by Inhibiting Chronic Inflammation. Biomed. Pharmacother. 2021, 137, 111418. [Google Scholar] [CrossRef]
- Ye, L.; Chen, X.; Wang, M.; Jin, L.; Zhuang, Z.; Yang, D.; Guan, X.; Samorodov, A.V.; Pavlov, V.N.; Chattipakorn, N.; et al. Curcumin Analogue C66 Attenuates Obesity-Induced Myocardial Injury by Inhibiting JNK-Mediated Inflammation. Biomed. Pharmacother. 2021, 143, 112121. [Google Scholar] [CrossRef]
- Babu, K.V.; Rajasekharan, K.N. Simplified Condition for Synthesis of Curcumin I and Other Curcuminoids. Org. Prep. Proced. Int. 1994, 26, 674–677. [Google Scholar] [CrossRef]
- Anto, R.J.; George, J.; Dinesh Babu, K.V.; Rajasekharan, K.N.; Kuttan, R. Antimutagenic and Anticarcinogenic Activity of Natural and Synthetic Curcuminoids. Mutat. Res.—Genet. Toxicol. 1996, 370, 127–131. [Google Scholar] [CrossRef]
- Rukkumani, R.; Aruna, K.; Suresh Varma, P.; Viswanathan, P.; Rajasekaran, K.N.N.; Menon, V.P. Protective Role of a Novel Curcuminoid on Alcohol and PUFA-Induced Hyperlipidemia. Toxicol. Mech. Methods 2005, 15, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Rukkumani, R.; Aruna, K.; Varma, P.S.; Rajasekaran, K.N.; Menon, V.P. Comparative Effects of Curcumin and Its Analog on Alcohol- and Polyunsaturated Fatty Acid-Induced Alterations in Circulatory Lipid Profiles. J. Med. Food 2005, 8, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Zhong, P.; Liang, D.; Xu, Z.; Skibba, M.; Zeng, C.; Li, X.; Wei, T.; Wu, L.; Liang, G. A Newly Designed Curcumin Analog Y20 Mitigates Cardiac Injury via Anti-Inflammatory and Anti-Oxidant Actions in Obese Rats. PLoS ONE 2015, 10, e0120215. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, H. Current Status and Prospects of Polyethyleneglycol-Modified Medicines. Biol. Pharm. Bull. 2013, 36, 883–888. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Murphy, C.J.; Zhang, B.; Shen, Y.; Sui, M.; Van Kirk, E.A.; Feng, X.; Murdoch, W.J. Amphiphilic Curcumin Conjugate-Forming Nanoparticles as Anticancer Prodrug and Drug Carriers: In Vitro and In Vivo Effects. Nanomedicine 2010, 5, 855–865. [Google Scholar] [CrossRef]
- Xiao, S.; Deng, Y.; Shen, N.; Sun, Y.; Tang, H.; Hu, P.; Ren, H.; Peng, M. Curc-MPEG454, a PEGylated Curcumin Derivative, as a Multi-Target Anti-Fibrotic Prodrug. Int. Immunopharmacol. 2021, 101, 108166. [Google Scholar] [CrossRef]
- Liu, Y.; Cheng, F.; Luo, Y.; Zhan, Z.; Hu, P.; Ren, H.; Tang, H.; Peng, M. PEGylated Curcumin Derivative Attenuates Hepatic Steatosis via CREB/PPAR-γ/CD36 Pathway. Biomed Res. Int. 2017, 2017, 8234507. [Google Scholar] [CrossRef]
- Mapoung, S.; Suzuki, S.; Fuji, S.; Naiki-Ito, A.; Kato, H.; Yodkeeree, S.; Sakorn, N.; Ovatlarnporn, C.; Takahashi, S.; Limtrakul Dejkriengkraikul, P. Dehydrozingerone, a Curcumin Analog, as a Potential Anti-Prostate Cancer Inhibitor In Vitro and In Vivo. Molecules 2020, 25, 2737. [Google Scholar] [CrossRef]
- Lapchak, P.A.; McKim, J.M. CeeToxTM Analysis of CNB-001 a Novel Curcumin-Based Neurotrophic/Neuroprotective Lead Compound to Treat Stroke: Comparison with NXY-059 and Radicut. Transl. Stroke Res. 2011, 2, 51–59. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moetlediwa, M.T.; Ramashia, R.; Pheiffer, C.; Titinchi, S.J.J.; Mazibuko-Mbeje, S.E.; Jack, B.U. Therapeutic Effects of Curcumin Derivatives against Obesity and Associated Metabolic Complications: A Review of In Vitro and In Vivo Studies. Int. J. Mol. Sci. 2023, 24, 14366. https://doi.org/10.3390/ijms241814366
Moetlediwa MT, Ramashia R, Pheiffer C, Titinchi SJJ, Mazibuko-Mbeje SE, Jack BU. Therapeutic Effects of Curcumin Derivatives against Obesity and Associated Metabolic Complications: A Review of In Vitro and In Vivo Studies. International Journal of Molecular Sciences. 2023; 24(18):14366. https://doi.org/10.3390/ijms241814366
Chicago/Turabian StyleMoetlediwa, Marakiya T., Rudzani Ramashia, Carmen Pheiffer, Salam J. J. Titinchi, Sithandiwe E. Mazibuko-Mbeje, and Babalwa U. Jack. 2023. "Therapeutic Effects of Curcumin Derivatives against Obesity and Associated Metabolic Complications: A Review of In Vitro and In Vivo Studies" International Journal of Molecular Sciences 24, no. 18: 14366. https://doi.org/10.3390/ijms241814366
APA StyleMoetlediwa, M. T., Ramashia, R., Pheiffer, C., Titinchi, S. J. J., Mazibuko-Mbeje, S. E., & Jack, B. U. (2023). Therapeutic Effects of Curcumin Derivatives against Obesity and Associated Metabolic Complications: A Review of In Vitro and In Vivo Studies. International Journal of Molecular Sciences, 24(18), 14366. https://doi.org/10.3390/ijms241814366