


A Unique G-Quadruplex Aptamer: A Novel Approach for Cancer Cell Recognition, Cell Membrane Visualization, and RSV Infection Detection


 and
and
Abstract
:1. Introduction
2. Results
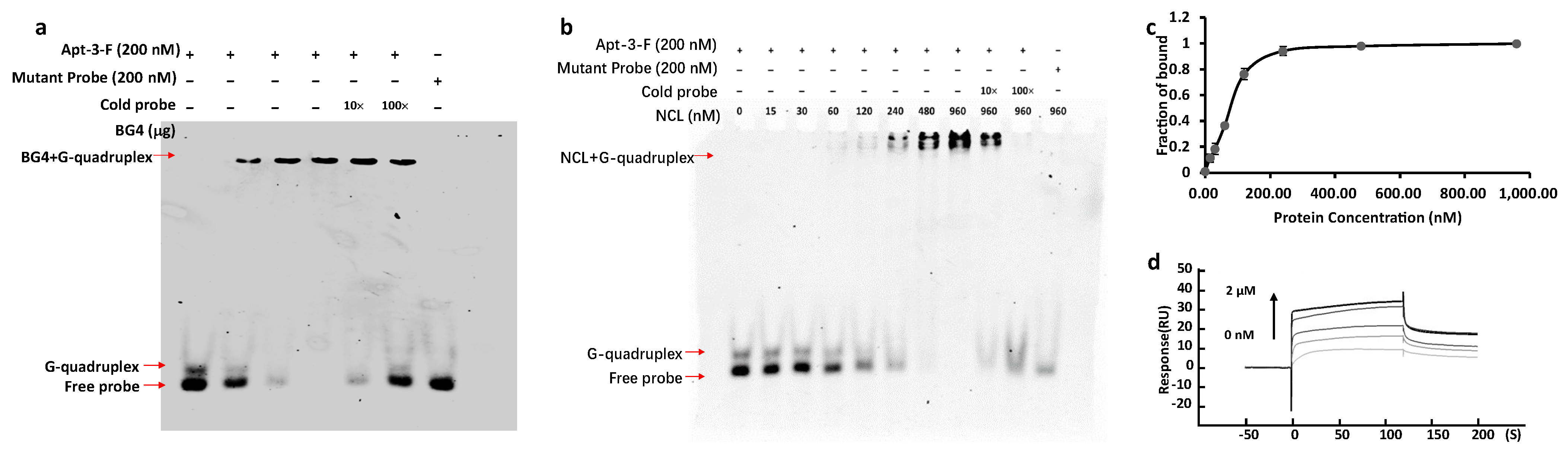
2.1. Finding the Aptamer Binding to NCL on Cell Surface
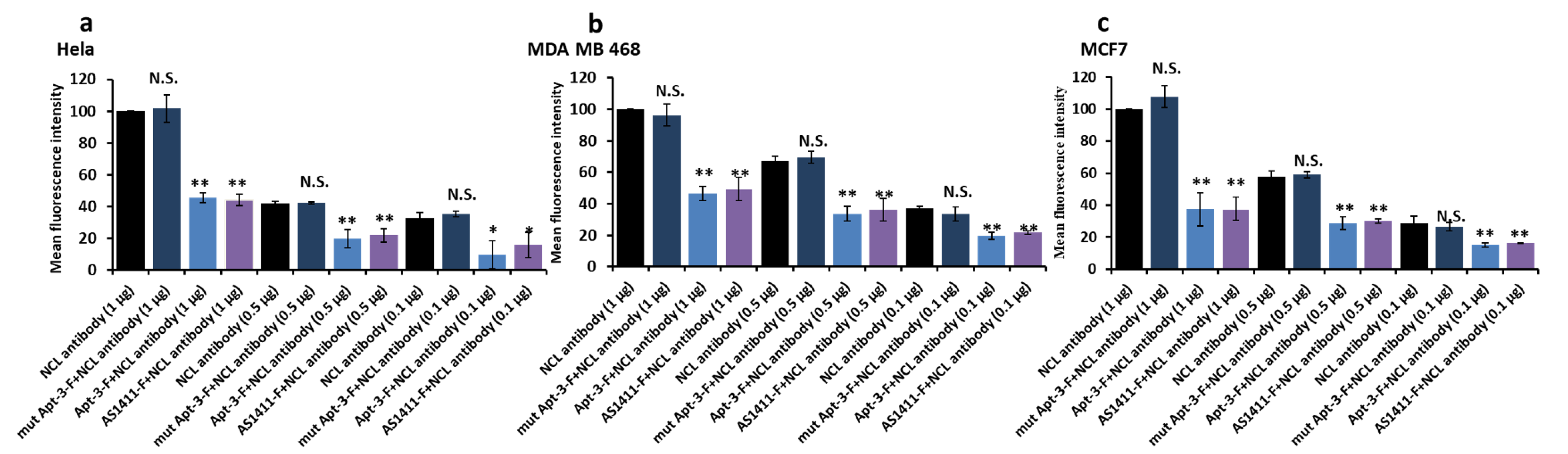
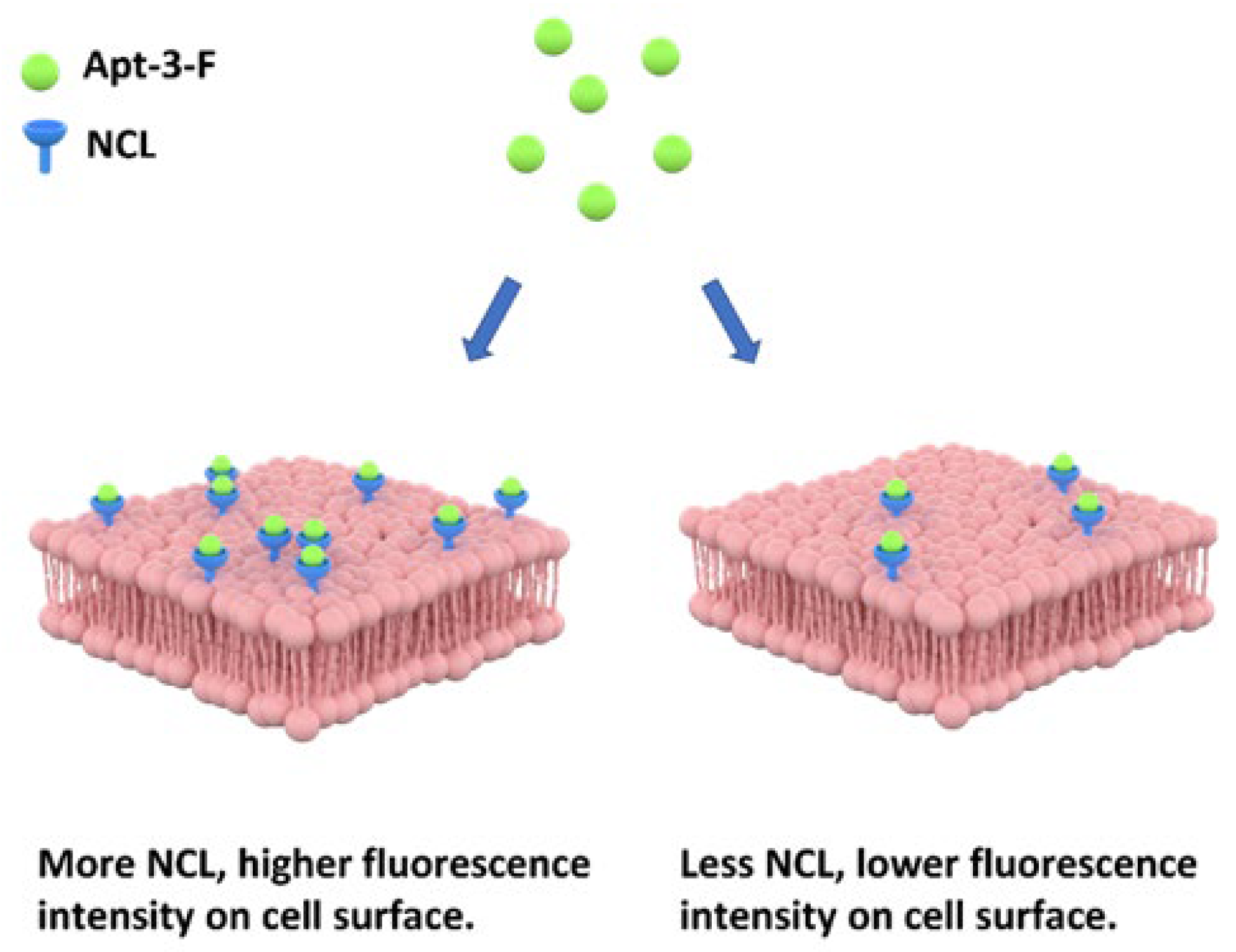
2.2. Detecting the NCL on Cell Surface by Apt-3-F
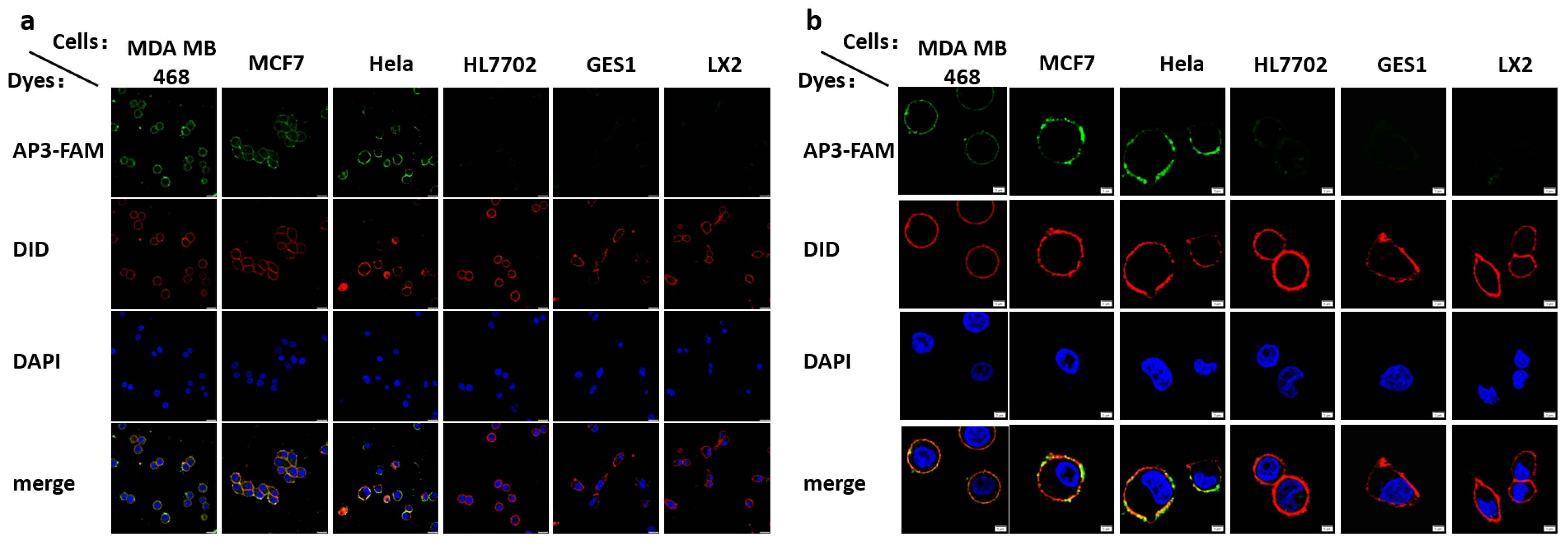
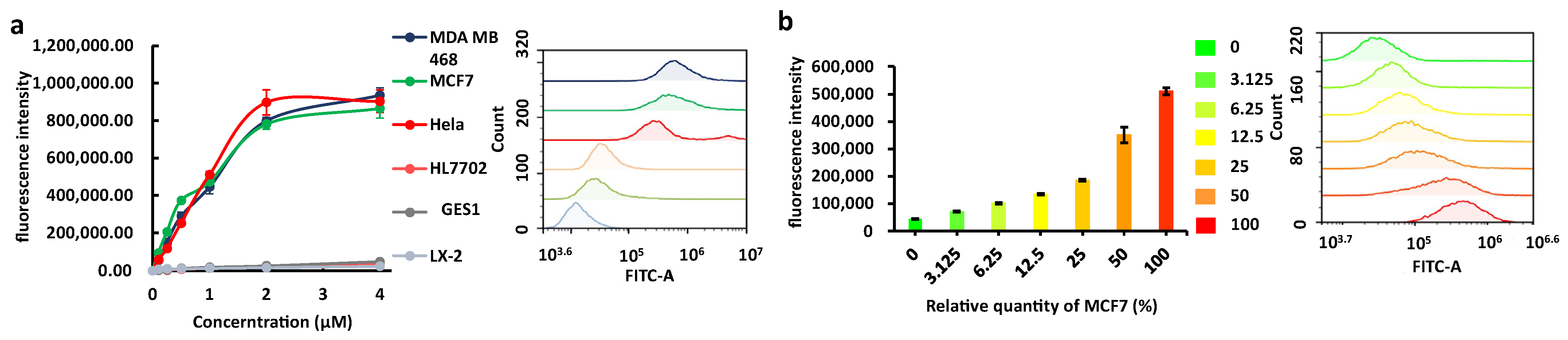
2.3. Cell Membrane Staining and Cancer Cell Indication by Apt-3-F
2.4. RSV Infection Indicated by Apt-3-F
3. Discussion
4. Materials and Methods
4.1. Oligonucleotide Synthesis and Sample Preparation
4.2. Circular Dichroism (CD) Spectroscopy Experiments
4.3. Surface Plasmon Resonance (SPR) Experiments
4.4. Cell Culture and Cell Viability Assays
4.5. Electrophoretic Mobility Shift Assay (EMSA)
4.6. Immunohistochemistry-Based (IHC-Based) Flow Cytometry for Assessing Aptamer Binding to Cell Surface Nucleolin
4.7. Laser Confocal Imaging
4.8. Flow Cytometry Analysis for Aptamer Staining in Cells
4.9. Western Blotting
4.10. Nuclear Magnetic Resonance (NMR) Experiments
4.11. RNA Interference (RNAi)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ginisty, H.; Sicard, H.; Roger, B.; Bouvet, P. Structure and functions of nucleolin. J. Cell Sci. 1999, 112, 761–772. [Google Scholar] [CrossRef]
- Fremerey, J.; Morozov, P.; Meyer, C.; Garzia, A.; Teplova, M.; Tuschl, T.; Borkhardt, A. Nucleolin Controls Ribosome Biogenesis through Its RNA-Binding Properties. Blood 2016, 128, 5056. [Google Scholar] [CrossRef]
- Kim, K.; Dimitrova, D.D.; Carta, K.M.; Saxena, A.; Daras, M.; Borowiec, J.A. Novel Checkpoint Response to Genotoxic Stress Mediated by Nucleolin-Replication Protein A Complex Formation. Mol. Cell. Biol. 2005, 25, 2463–2474. [Google Scholar] [CrossRef] [PubMed]
- Bharti, A.K.; Olson, M.O.; Kufe, D.W.; Rubin, E.H. Identification of a nucleolin binding site in human topoisomerase I. J. Biol. Chem. 1996, 271, 1993–1997. [Google Scholar] [CrossRef]
- Kobayashi, J.; Fujimoto, H.; Sato, J.; Hayashi, I.; Burma, S.; Matsuura, S.; Chen, D.J.; Komatsu, K. Nucleolin Participates in DNA Double-Strand Break-Induced Damage Response through MDC1-Dependent Pathway. PLoS ONE 2012, 7, e49245. [Google Scholar] [CrossRef] [PubMed]
- Grinstein, E.; Du, Y.; Santourlidis, S.; Christ, J.; Uhrberg, M.; Wernet, P. Nucleolin regulates gene expression in CD34-positive hematopoietic cells. J. Biol. Chem. 2007, 282, 12439–12449. [Google Scholar] [CrossRef] [PubMed]
- Mahotka, C.; Bhatia, S.; Kollet, J.; Grinstein, E. Nucleolin promotes execution of the hematopoietic stem cell gene expression program. Leukemia 2018, 32, 1865–1868. [Google Scholar] [CrossRef] [PubMed]
- Kirman, D.C.; Renganathan, B.; Chui, W.K.; Chen, M.W.; Kaya, N.A.; Ge, R. Cell surface nucleolin is a novel ADAMTS5 receptor mediating endothelial cell apoptosis. Cell Death Dis. 2022, 13, 172. [Google Scholar] [CrossRef]
- Wise, J.F.; Berkova, Z.; Mathur, R.; Zhu, H.; Braun, F.K.; Tao, R.H.; Sabichi, A.L.; Ao, X.; Maeng, H.; Samaniego, F. Nucleolin inhibits Fas ligand binding and suppresses Fas-mediated apoptosis in vivo via a surface nucleolin-Fas complex. Blood 2013, 121, 4729–4739. [Google Scholar] [CrossRef]
- Berger, C.M.; Gaume, X.; Bouvet, P. The roles of nucleolin subcellular localization in cancer. Biochimie 2015, 113, 78–85. [Google Scholar] [CrossRef]
- Cruz, C.; Pedro, A.Q.; Carvalho, J.; Santos, T.; Talhada, D.; Paiva, A.; Queiroz, J.A.; Andrade, M.; Pinto, M.; Montenegro, L.; et al. Nucleolin as a potential biomarker for canine malignant neoplasia. Res. Vet. Sci. 2021, 135, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Romano, S.; Fonseca, N.; Simoes, S.; Goncalves, J.; Moreira, J.N. Nucleolin-based targeting strategies for cancer therapy: From targeted drug delivery to cytotoxic ligands. Drug Discov. Today 2019, 24, 1985–2001. [Google Scholar] [CrossRef] [PubMed]
- Yangngam, S.; Prasopsiri, J.; Hatthakarnkul, P.; Thongchot, S.; Thuwajit, P.; Yenchitsomanus, P.T.; Edwards, J.; Thuwajit, C. Cellular localization of nucleolin determines the prognosis in cancers: A meta-analysis. J. Mol. Med. 2022, 100, 1145–1157. [Google Scholar] [CrossRef] [PubMed]
- Brignole, C.; Bensa, V.; Fonseca, N.A.; Del Zotto, G.; Bruno, S.; Cruz, A.F.; Malaguti, F.; Carlini, B.; Morandi, F.; Calarco, E.; et al. Cell surface Nucleolin represents a novel cellular target for neuroblastoma therapy. J. Exp. Clin. Cancer Res. 2021, 40, 180. [Google Scholar] [CrossRef]
- Huang, F.; Wu, Y.; Tan, H.; Guo, T.; Zhang, K.; Li, D.; Tong, Z. Phosphorylation of nucleolin is indispensable to its involvement in the proliferation and migration of non-small cell lung cancer cells. Oncol. Rep. 2019, 41, 590–598. [Google Scholar] [CrossRef]
- Wu, D.-M.; Zhang, P.; Liu, R.-Y.; Sang, Y.-X.; Zhou, C.; Xu, G.-C.; Yang, J.-L.; Tong, A.-P.; Wang, C.-T. Phosphorylation and changes in the distribution of nucleolin promote tumor metastasis via the PI3K/Akt pathway in colorectal carcinoma. FEBS Lett. 2014, 588, 1921–1929. [Google Scholar] [CrossRef]
- Shi, H.; Huang, Y.; Zhou, H.; Song, X.; Yuan, S.; Fu, Y.; Luo, Y. Nucleolin is a receptor that mediates antiangiogenic and antitumor activity of endostatin. Blood 2007, 110, 2899–2906. [Google Scholar] [CrossRef]
- Tonello, F.; Massimino, M.L.; Peggion, C. Nucleolin: A cell portal for viruses, bacteria, and toxins. Cell. Mol. Life Sci. 2022, 79, 271. [Google Scholar] [CrossRef]
- Techaarpornkul, S.; Collins, P.L.; Peeples, M.E. Respiratory syncytial virus with the fusion protein as its only viral glycoprotein is less dependent on cellular glycosaminoglycans for attachment than complete virus. Virology 2002, 294, 296–304. [Google Scholar] [CrossRef]
- Griffiths, C.D.; Bilawchuk, L.M.; McDonough, J.E.; Jamieson, K.C.; Elawar, F.; Cen, Y.; Duan, W.; Lin, C.; Song, H.; Casanova, J.-L.; et al. IGF1R is an entry receptor for respiratory syncytial virus. Nature 2020, 583, 615–619. [Google Scholar] [CrossRef]
- Tayyari, F.; Marchant, D.; Moraes, T.J.; Duan, W.; Mastrangelo, P.; Hegele, R.G. Identification of nucleolin as a cellular receptor for human respiratory syncytial virus. Nat. Med. 2011, 17, 1132–1135. [Google Scholar] [CrossRef] [PubMed]
- Palsson, S.A.; Dondalska, A.; Bergenstrahle, J.; Rolfes, C.; Bjork, A.; Sedano, L.; Power, U.F.; Rameix-Welti, M.A.; Lundeberg, J.; Wahren-Herlenius, M.; et al. Single-Stranded Oligonucleotide-Mediated Inhibition of Respiratory Syncytial Virus Infection. Front. Immunol. 2020, 11, 580547. [Google Scholar] [CrossRef] [PubMed]
- Bao, H.L.; Ishizuka, T.; Yamashita, A.; Furukoji, E.; Asada, Y.; Xu, Y. Improving Thermodynamic Stability and Anticoagulant Activity of a Thrombin Binding Aptamer by Incorporation of 8-trifluoromethyl-2’-deoxyguanosine. J. Med. Chem. 2020, 64, 711–718. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Peng, T.; Peng, Y.; Ai, L.; Deng, Z.; Wang, X.Q.; Tan, W. Molecularly Engineering Triptolide with Aptamers for High Specificity and Cytotoxicity for Triple-Negative Breast Cancer. J. Am. Chem. Soc. 2020, 142, 2699–2703. [Google Scholar] [CrossRef] [PubMed]
- Bates, P.J.; Choi, E.W.; Nayak, L.V. G-rich oligonucleotides for cancer treatment. Methods Mol. Biol. 2009, 542, 379–392. [Google Scholar] [PubMed]
- Bates, P.J.; Laber, D.A.; Miller, D.M.; Thomas, S.D.; Trent, J.O. Discovery and development of the G-rich oligonucleotide AS1411 as a novel treatment for cancer. Exp. Mol. Pathol. 2009, 86, 151–164. [Google Scholar] [CrossRef]
- Bates, P.J.; Reyes-Reyes, E.M.; Malik, M.T.; Murphy, E.M.; O’Toole, M.G.; Trent, J.O. G-quadruplex oligonucleotide AS1411 as a cancer-targeting agent: Uses and mechanisms. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 1414–1428. [Google Scholar]
- Do, N.Q.; Chung, W.J.; Truong, T.H.A.; Heddi, B.; Phan, A.T. G-quadruplex structure of an anti-proliferative DNA sequence. Nucleic Acids Res. 2017, 45, 7487–7493. [Google Scholar] [CrossRef]
- Dailey, M.M.; Miller, M.C.; Bates, P.J.; Lane, A.N.; Trent, J.O. Resolution and characterization of the structural polymorphism of a single quadruplex-forming sequence. Nucleic Acids Res. 2010, 38, 4877–4888. [Google Scholar] [CrossRef]
- Ogloblina, A.M.; Iaccarino, N.; Capasso, D.; Di Gaetano, S.; Garzarella, E.U.; Dolinnaya, N.G.; Yakubovskaya, M.G.; Pagano, B.; Amato, J.; Randazzo, A. Toward G-Quadruplex-Based Anticancer Agents: Biophysical and Biological Studies of Novel AS1411 Derivatives. Int. J. Mol. Sci. 2020, 21, 7781. [Google Scholar] [CrossRef]
- Saha, A.; Duchambon, P.; Masson, V.; Loew, D.; Bombard, S.; Teulade-Fichou, M.P. Nucleolin Discriminates Drastically between Long-Loop and Short-Loop Quadruplexes. Biochemistry 2020, 59, 1261–1272. [Google Scholar] [CrossRef]
- Santos, T.; Salgado, G.F.; Cabrita, E.J.; Cruz, C. Nucleolin: A binding partner of G-quadruplex structures. Trends Cell Biol. 2022, 32, 561–564. [Google Scholar] [CrossRef]
- Xiao, C.D.; Ishizuka, T.; Zhu, X.Q.; Li, Y.; Sugiyama, H.; Xu, Y. Unusual Topological RNA Architecture with an Eight-Stranded Helical Fragment Containing A-, G-, and U-Tetrads. J. Am. Chem. Soc. 2017, 139, 2565–2568. [Google Scholar] [CrossRef]
- Xiao, C.D.; Shibata, T.; Yamamoto, Y.; Xu, Y. An intramolecular antiparallel G-quadruplex formed by human telomere RNA. Chem. Commun. 2018, 54, 3944–3946. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.-D.; Jia, M.-H.; Zhong, M.-Q.; Xu, Y.; Yu, Z.-T.; He, Z.-Y.; Lu, X.; Zhang, Y.; Zhou, X.; Fu, L.-Y.; et al. Unveiling the role of G-quadruplex structure in promoter region: Regulation of ABCA1 expression in macrophages possibly via NONO protein recruitment. Int. J. Biol. Macromol. 2023, 245, 125443. [Google Scholar]
- Gatto, B.; Palumbo, M.; Sissi, C. Nucleic acid aptamers based on the G-quadruplex structure: Therapeutic and diagnostic potential. Curr. Med. Chem. 2009, 16, 1248–1265. [Google Scholar] [CrossRef] [PubMed]
- Roxo, C.; Kotkowiak, W.; Pasternak, A. G-Quadruplex-Forming Aptamers-Characteristics, Applications, and Perspectives. Molecules 2019, 24, 3781. [Google Scholar] [CrossRef]
- Jia, H.R.; Zhu, Y.X.; Duan, Q.Y.; Wu, F.G. Cell surface-localized imaging and sensing. Chem. Soc. Rev. 2021, 50, 6240–6277. [Google Scholar]
- Dash, J.; Shirude, P.S.; Hsu, S.T.; Balasubramanian, S. Diarylethynyl amides that recognize the parallel conformation of genomic promoter DNA G-quadruplexes. J. Am. Chem. Soc. 2008, 130, 15950–15956. [Google Scholar] [CrossRef]
- Palumbo, S.L.; Ebbinghaus, S.W.; Hurley, L.H. Formation of a unique end-to-end stacked pair of G-quadruplexes in the hTERT core promoter with implications for inhibition of telomerase by G-quadruplex-interactive ligands. J. Am. Chem. Soc. 2009, 131, 10878–10891. [Google Scholar]
- Ambrus, A.; Chen, D.; Dai, J.; Bialis, T.; Jones, R.A.; Yang, D. Human telomeric sequence forms a hybrid-type intramolecular G-quadruplex structure with mixed parallel/antiparallel strands in potassium solution. Nucleic Acids Res. 2006, 34, 2723–2735. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, C.; Cui, Y.; Mao, H.; Hurley, L.H. A Mechanosensor Mechanism Controls the G-Quadruplex/i-Motif Molecular Switch in the MYC Promoter NHE III. J. Am. Chem. Soc. 2016, 138, 14138–14151. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Zhao, G.; Zhang, S.; Nigim, F.; Zhou, G.; Yu, Z.; Song, Y.; Chen, Y.; Li, Y. AS1411-Induced Growth Inhibition of Glioma Cells by Up-Regulation of p53 and Down-Regulation of Bcl-2 and Akt1 via Nucleolin. PLoS ONE 2016, 11, e0167094. [Google Scholar]
- Soundararajan, S.; Chen, W.; Spicer, E.K.; Courtenay-Luck, N.; Fernandes, D.J. The nucleolin targeting aptamer AS1411 destabilizes Bcl-2 messenger RNA in human breast cancer cells. Cancer Res. 2008, 68, 2358–2365. [Google Scholar] [CrossRef]
- Ishimaru, D.; Zuraw, L.; Ramalingam, S.; Sengupta, T.K.; Bandyopadhyay, S.; Reuben, A.; Fernandes, D.J.; Spicer, E.K. Mechanism of regulation of bcl-2 mRNA by nucleolin and A+U-rich element-binding factor 1 (AUF1). J. Biol. Chem. 2010, 285, 27182–27191. [Google Scholar] [PubMed]
- Voskuil, J.L.A.; Bandrowski, A.; Begley, C.G.; Bradbury, A.R.M.; Chalmers, A.D.; Gomes, A.V.; Hardcastle, T.; Lund-Johansen, F.; Plückthun, A.; Roncador, G.; et al. The Antibody Society’s antibody validation webinar series. MAbs 2020, 12, 1794421. [Google Scholar] [CrossRef]
- Alamudi, S.H.; Kimoto, M.; Hirao, I. Uptake mechanisms of cell-internalizing nucleic acid aptamers for applications as pharmacological agents. RSC Med. Chem. 2021, 12, 1640–1649. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, W.; Chen, S.; Zhuang, Z.; Zhang, Y.; Jiang, L.; Lin, J.S. SELEX tool: A novel and convenient gel-based diffusion method for monitoring of aptamer-target binding. J. Biol. Eng. 2020, 14, 1–13. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aptamer Names | Sequence |
|---|---|
| Apt-1 | GUUAGGGUUUGGGAUUG |
| Apt-2 | GUUAGGGUUUUGGGAUUG |
| Apt-3 | GUUAGGGUUUAUGGGAUUG |
| Apt-4 | GUUAGGGUUUAUUGGGAUUG |
| Apt-5 | GUUAGGGUUUAUUUGGGAUUG |
| Apt-6 | GUUAGGGUUUAUUAUGGGAUUG |
| AS411 | GGTGGTGGTGGTTGTGGTGGTGGTGG |
| Cells | Apt-3 CC50 (μM) | AS1411 CC50 (μM) |
|---|---|---|
| MDA MB 468 | >128 | 11.10 |
| MCF7 | >128 | 15.60 |
| Hela | 119.20 | 4.49 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, C.-D.; Zhong, M.-Q.; Gao, Y.; Yang, Z.-L.; Jia, M.-H.; Hu, X.-H.; Xu, Y.; Shen, X.-C. A Unique G-Quadruplex Aptamer: A Novel Approach for Cancer Cell Recognition, Cell Membrane Visualization, and RSV Infection Detection. Int. J. Mol. Sci. 2023, 24, 14344. https://doi.org/10.3390/ijms241814344
Xiao C-D, Zhong M-Q, Gao Y, Yang Z-L, Jia M-H, Hu X-H, Xu Y, Shen X-C. A Unique G-Quadruplex Aptamer: A Novel Approach for Cancer Cell Recognition, Cell Membrane Visualization, and RSV Infection Detection. International Journal of Molecular Sciences. 2023; 24(18):14344. https://doi.org/10.3390/ijms241814344
Chicago/Turabian StyleXiao, Chao-Da, Ming-Qing Zhong, Yue Gao, Zheng-Lin Yang, Meng-Hao Jia, Xiao-Hui Hu, Yan Xu, and Xiang-Chun Shen. 2023. "A Unique G-Quadruplex Aptamer: A Novel Approach for Cancer Cell Recognition, Cell Membrane Visualization, and RSV Infection Detection" International Journal of Molecular Sciences 24, no. 18: 14344. https://doi.org/10.3390/ijms241814344
APA StyleXiao, C.-D., Zhong, M.-Q., Gao, Y., Yang, Z.-L., Jia, M.-H., Hu, X.-H., Xu, Y., & Shen, X.-C. (2023). A Unique G-Quadruplex Aptamer: A Novel Approach for Cancer Cell Recognition, Cell Membrane Visualization, and RSV Infection Detection. International Journal of Molecular Sciences, 24(18), 14344. https://doi.org/10.3390/ijms241814344