
In Vitro Assessment of Gold Nanoparticles on Telomerase Activity and Telomere Length in Human Fibroblasts


Abstract
:1. Introduction
2. Results
2.1. Toxicity
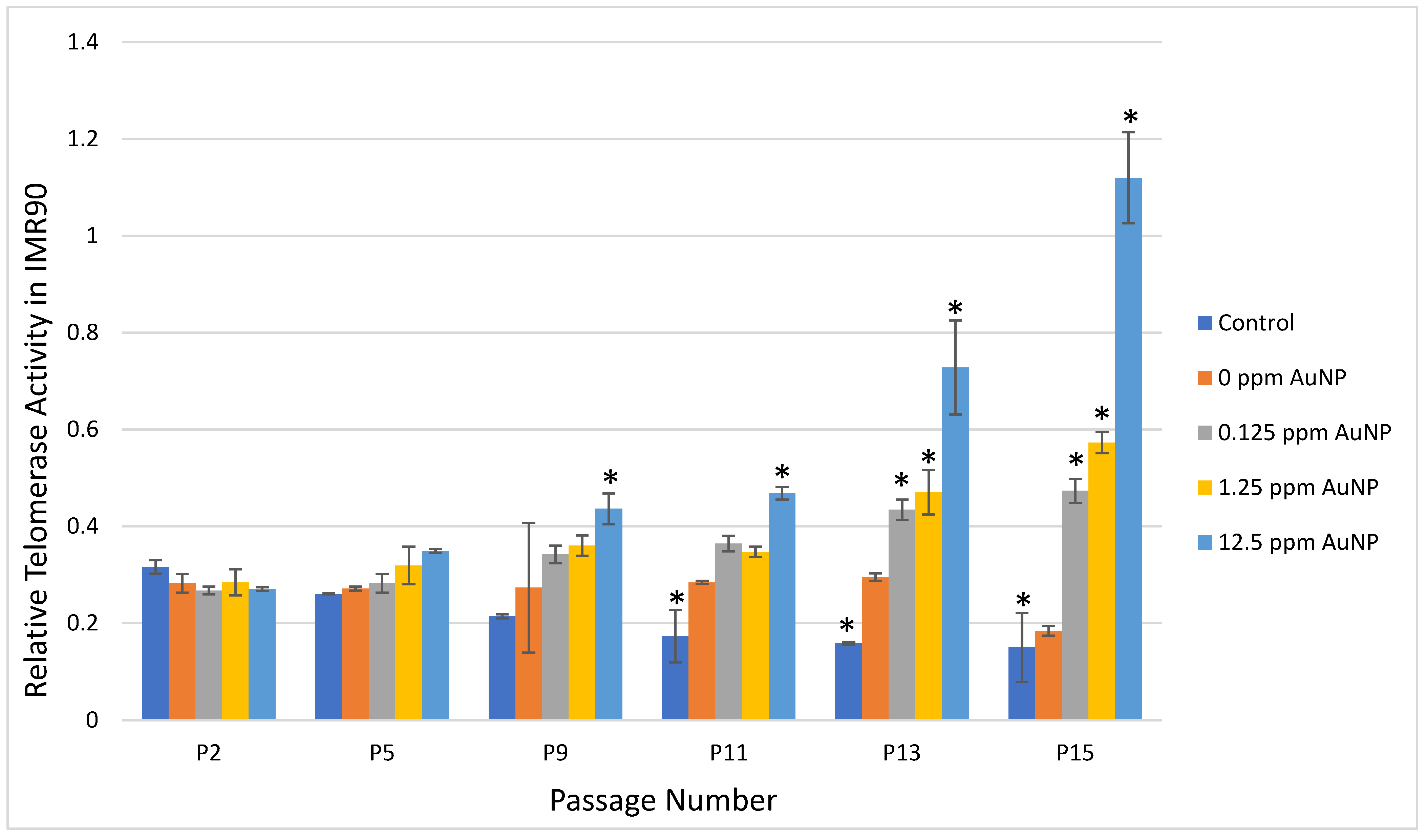
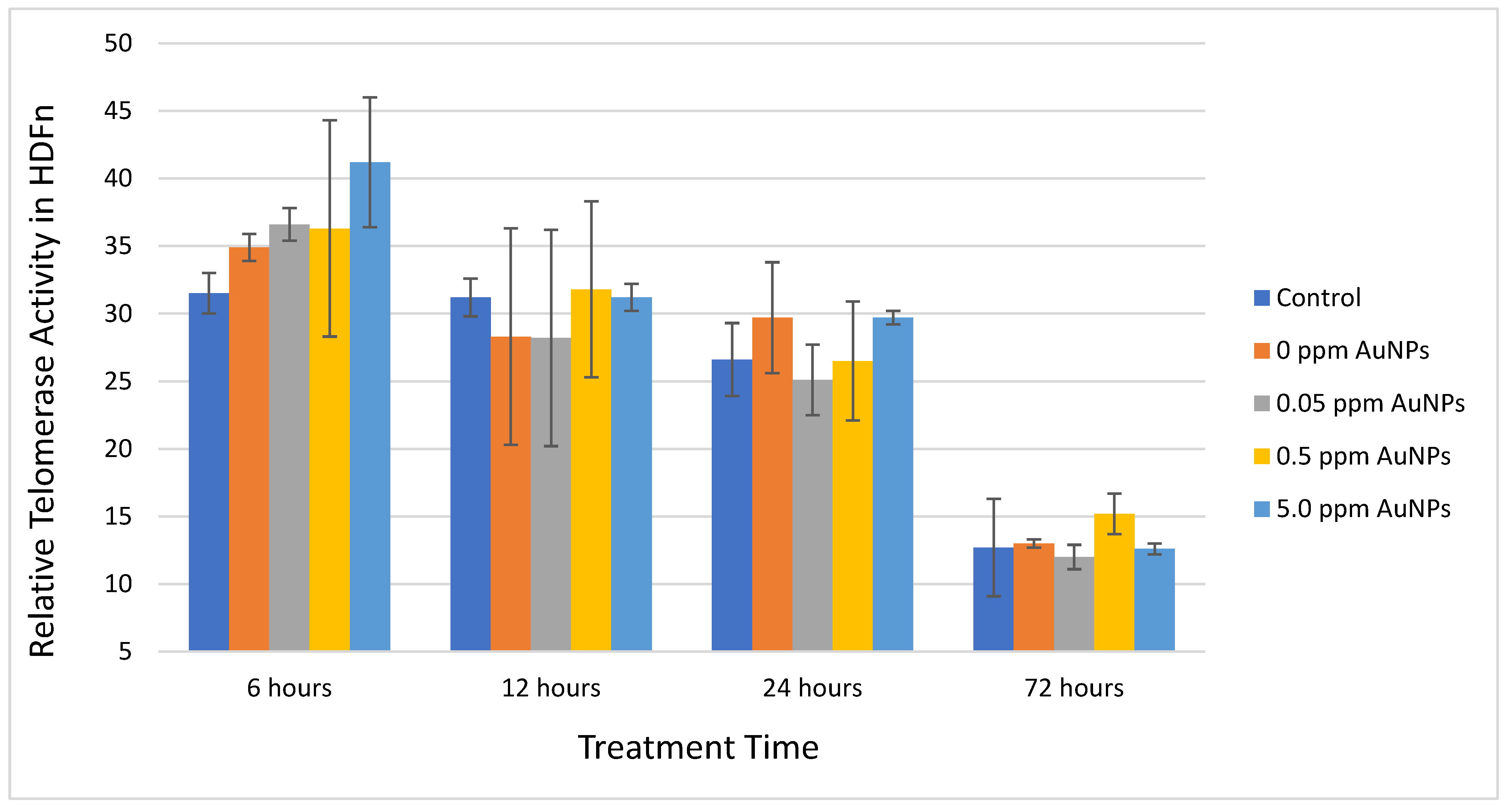
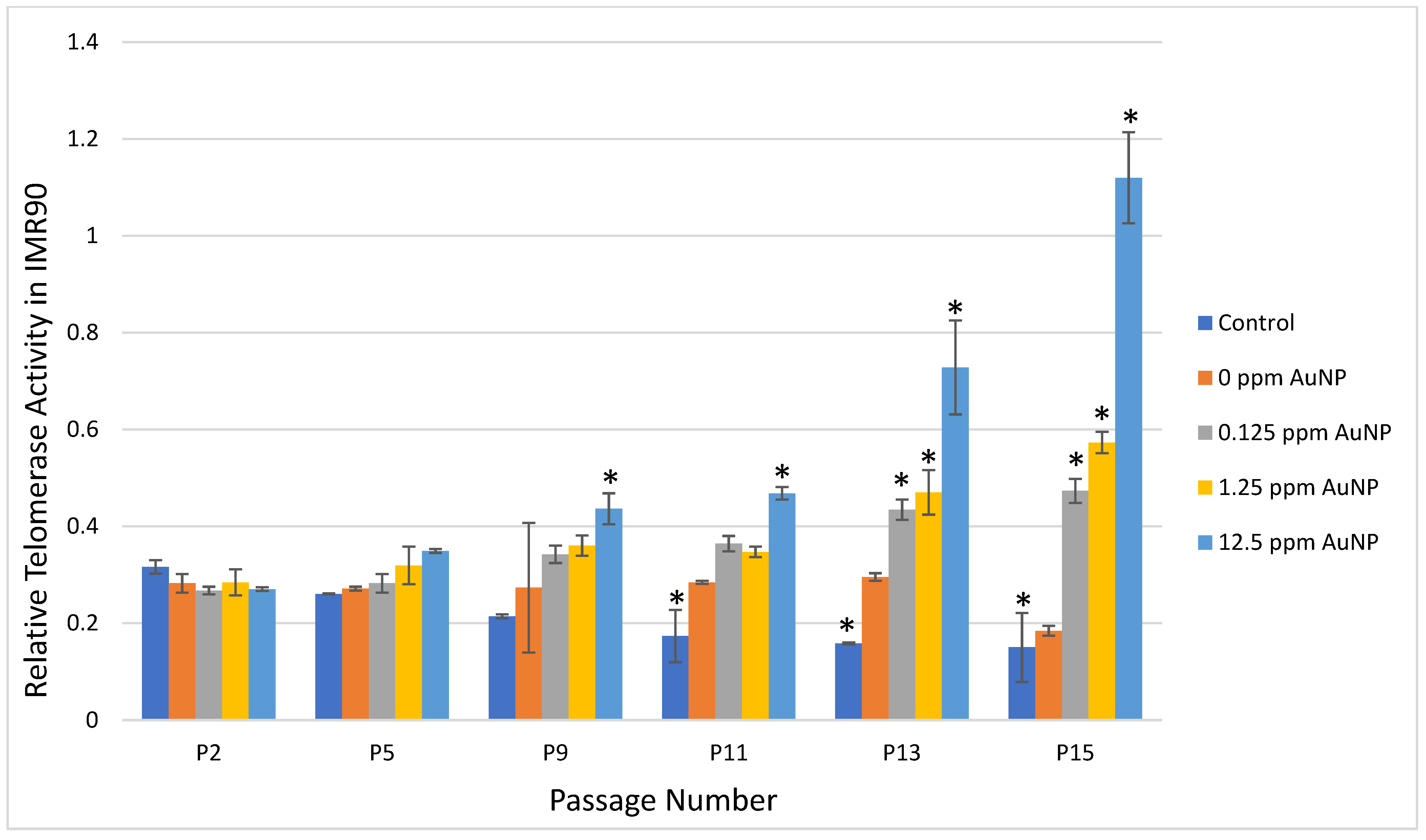
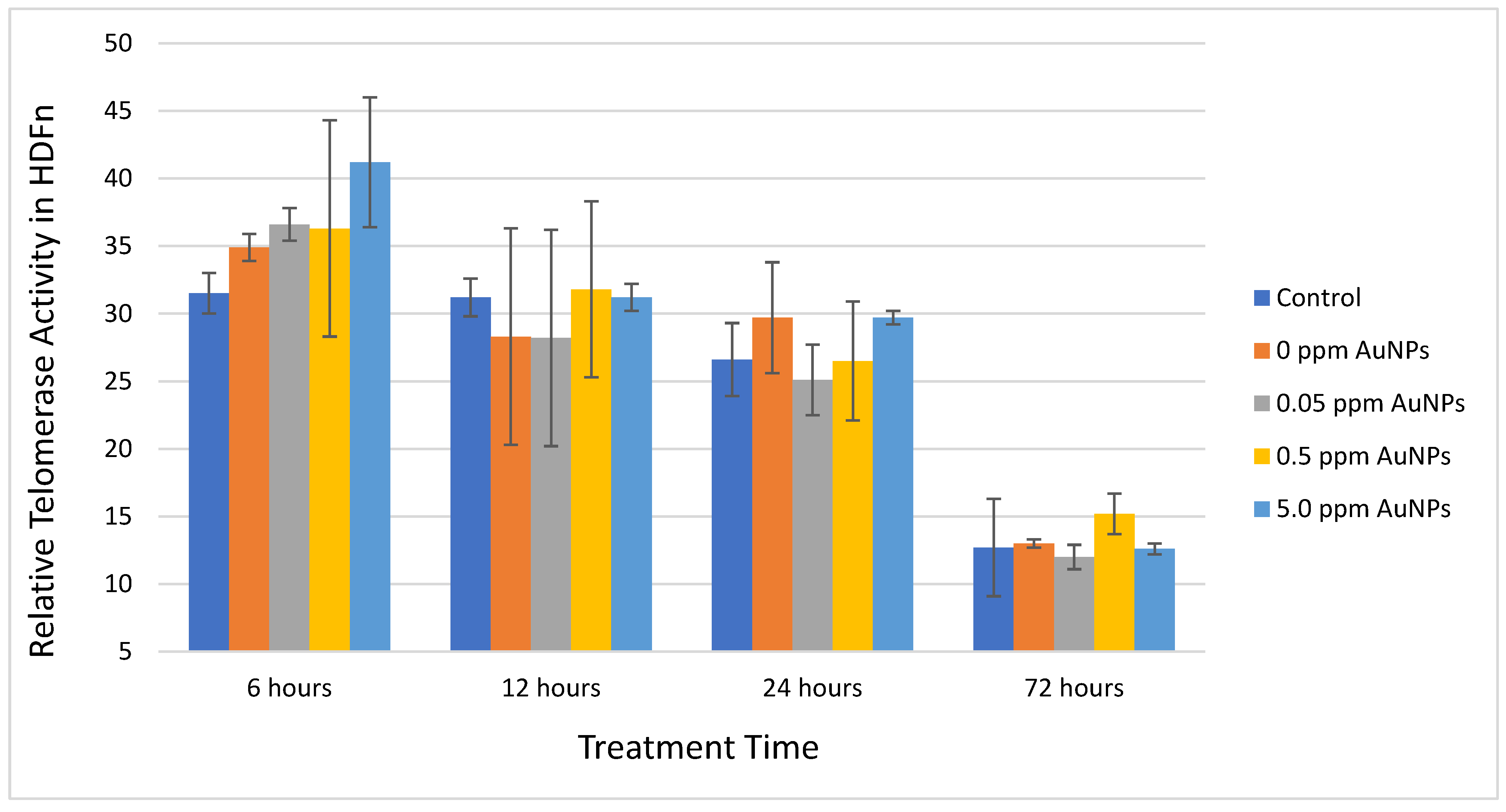
2.2. Telomerase Activity
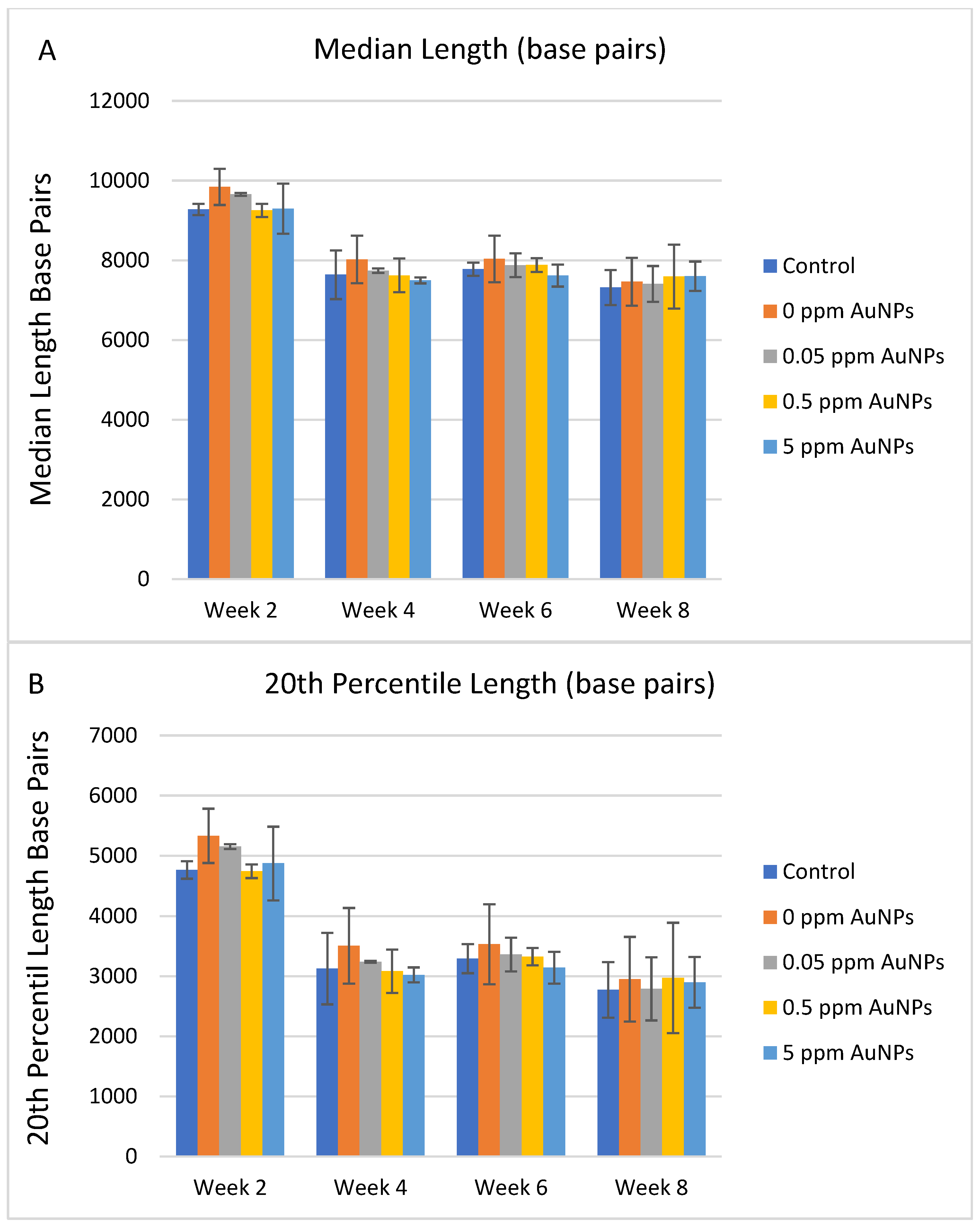
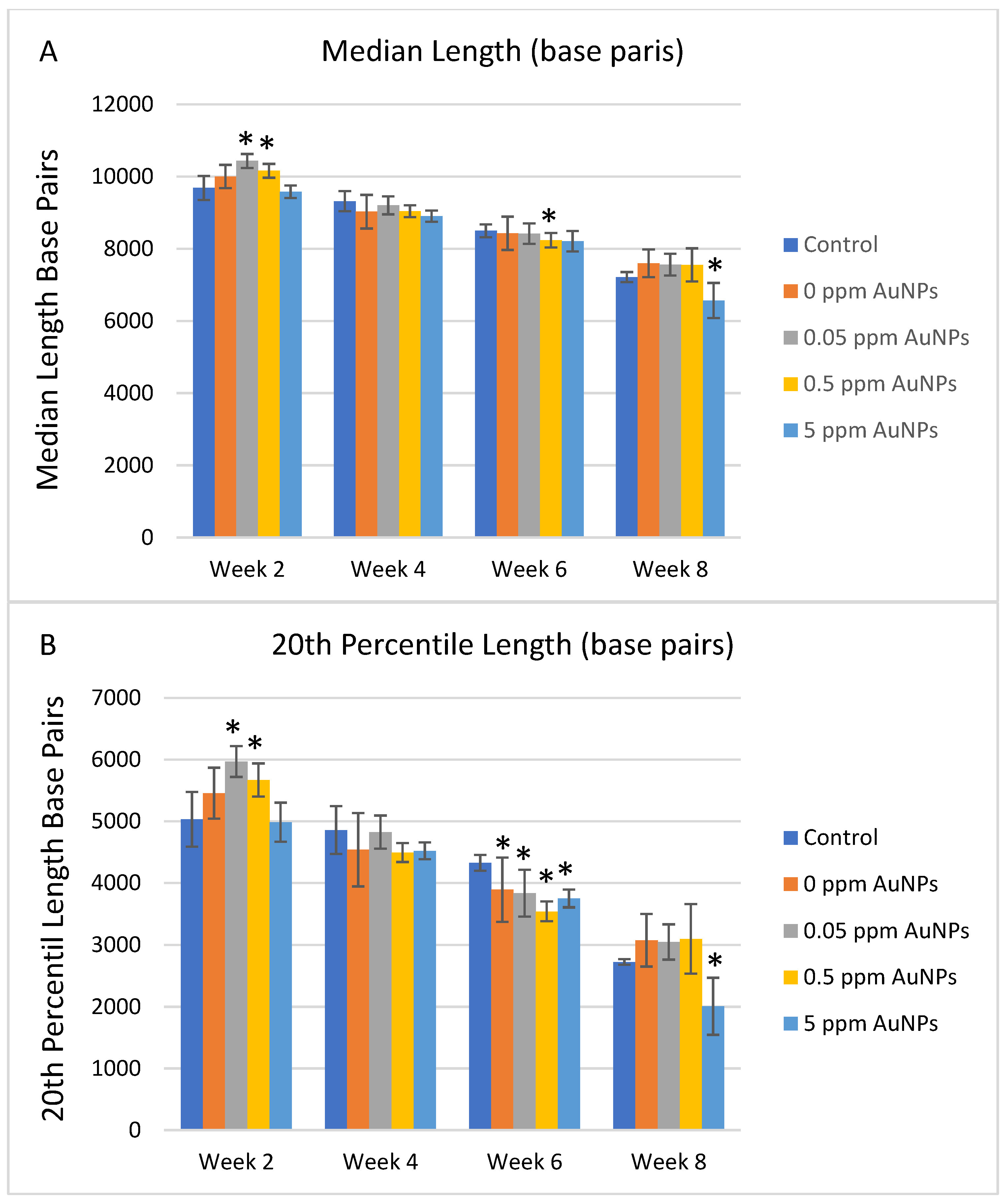
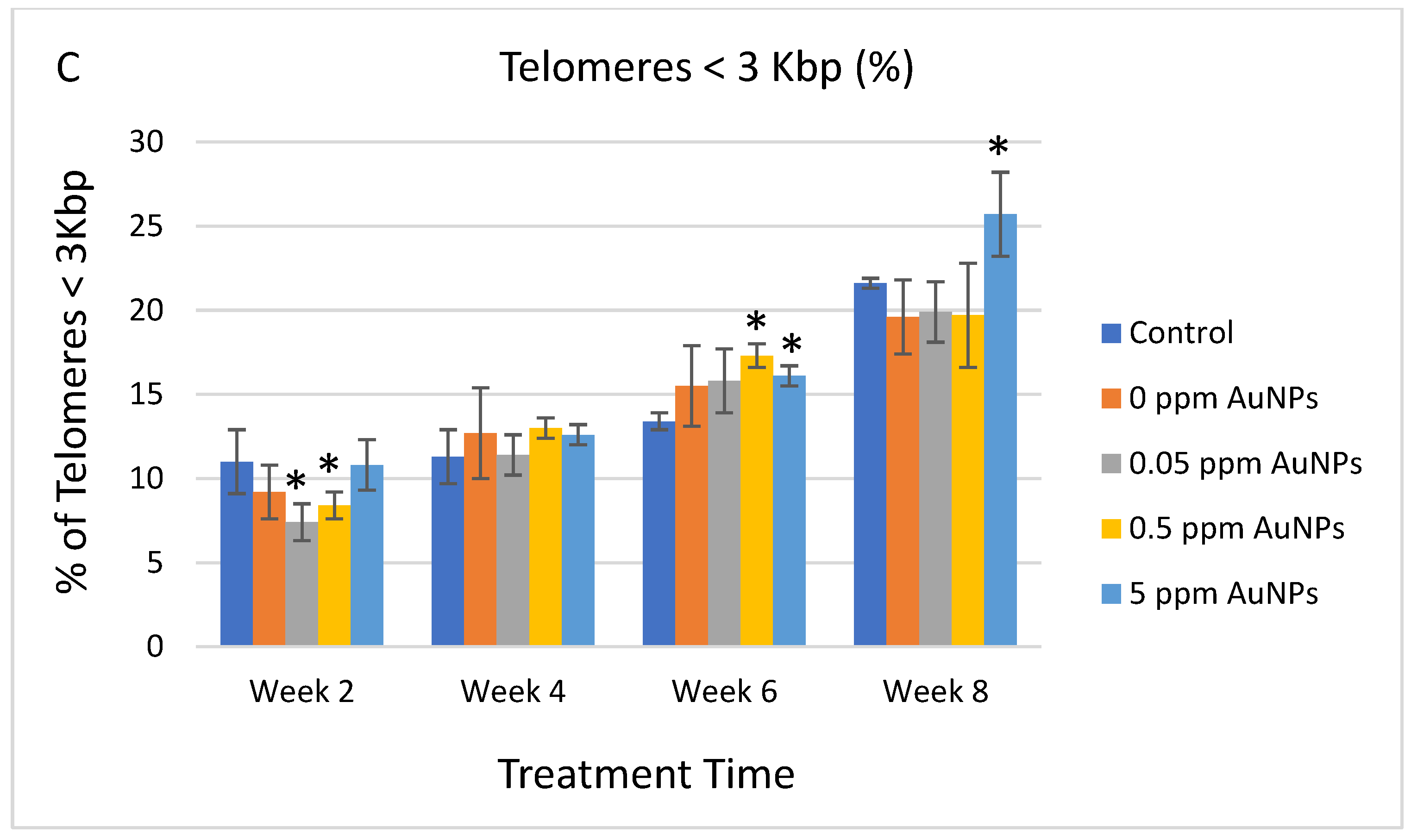
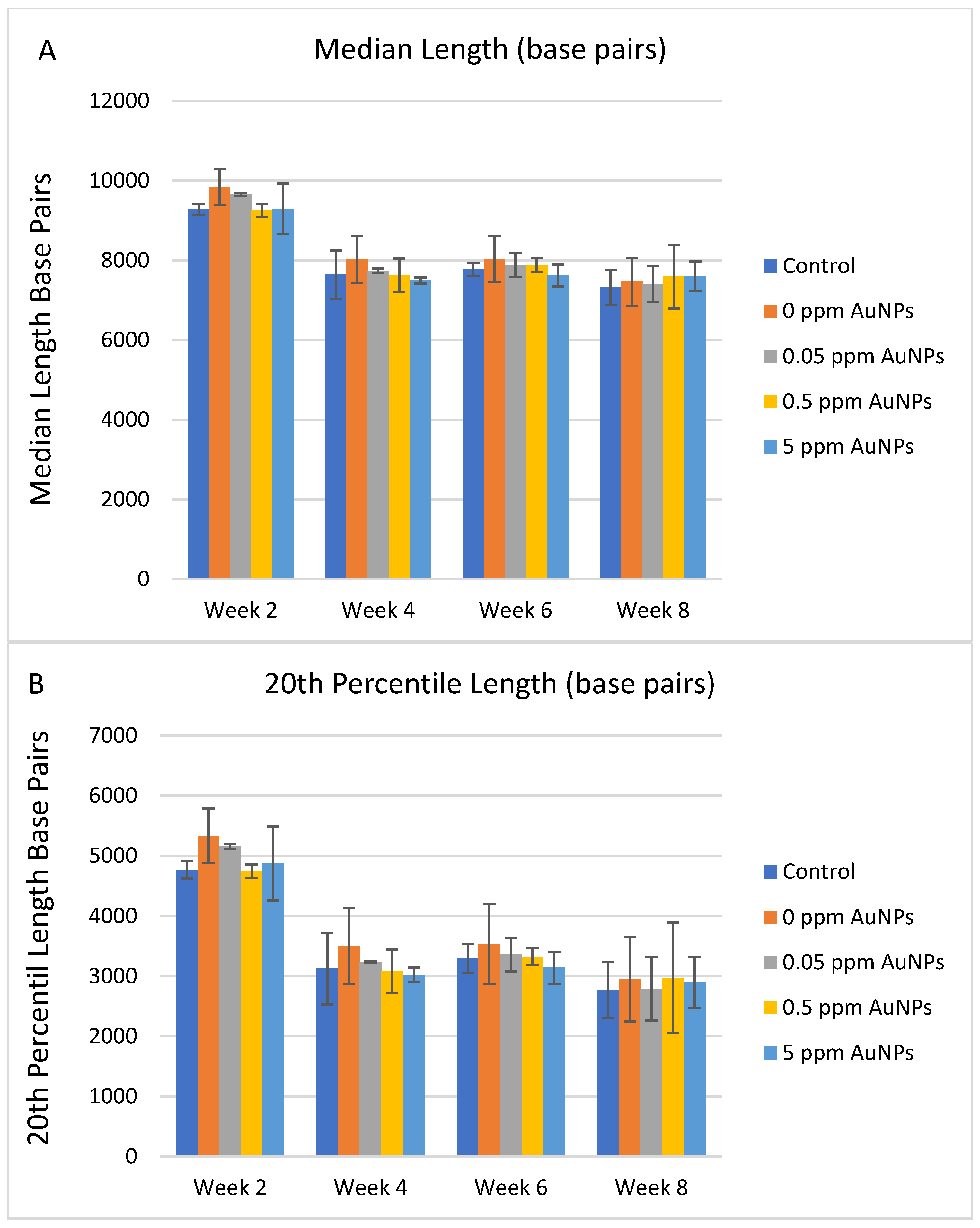
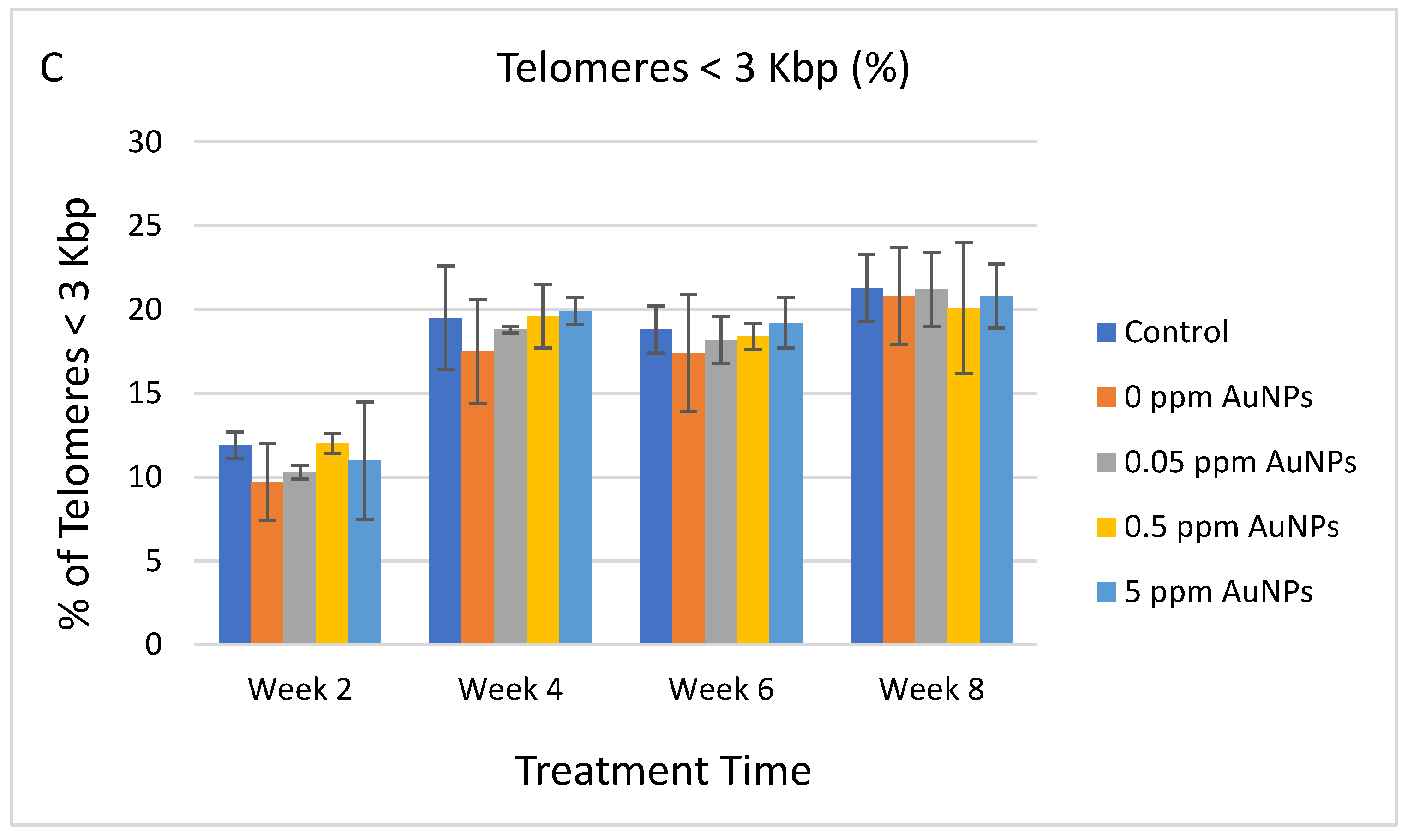
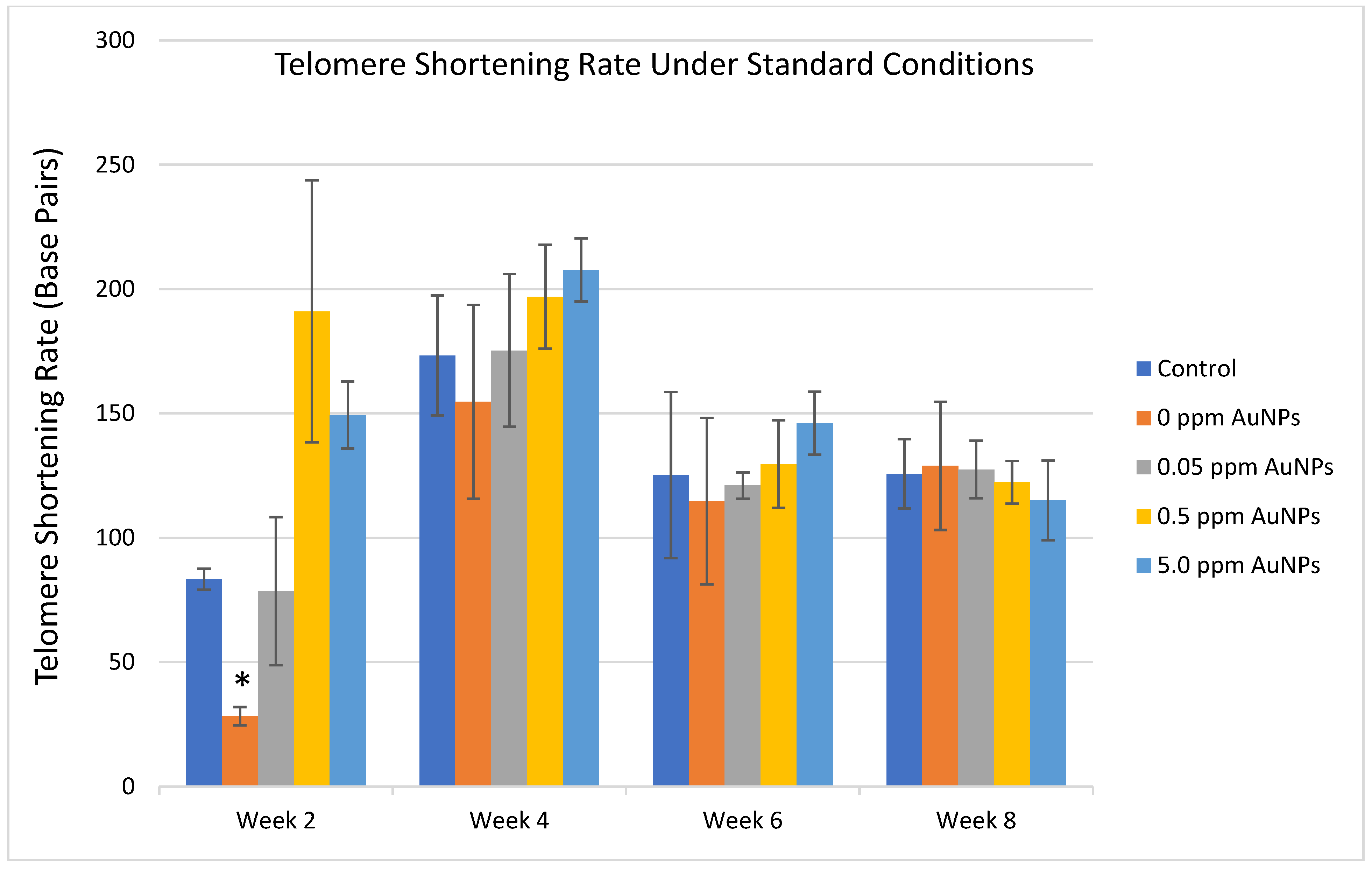
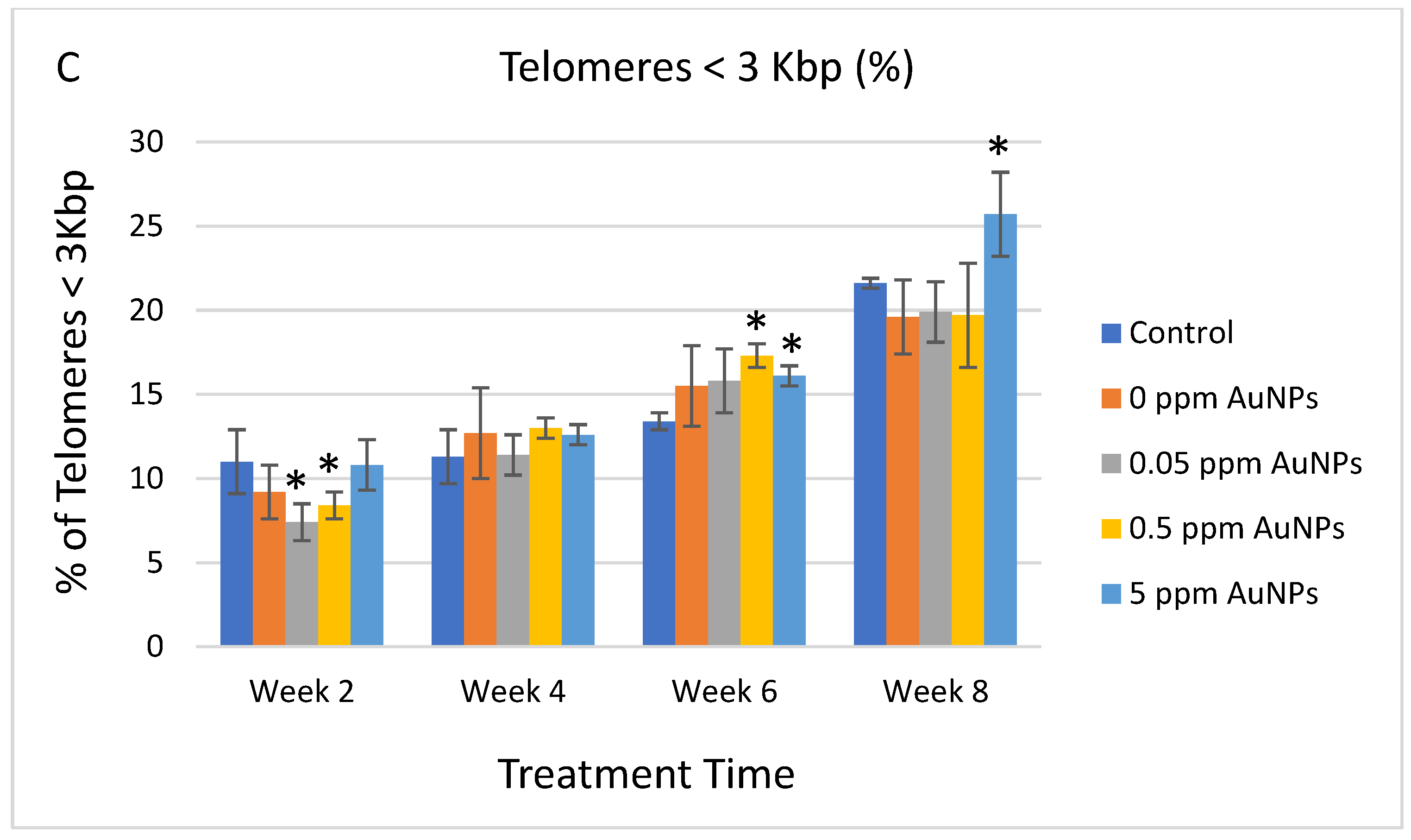
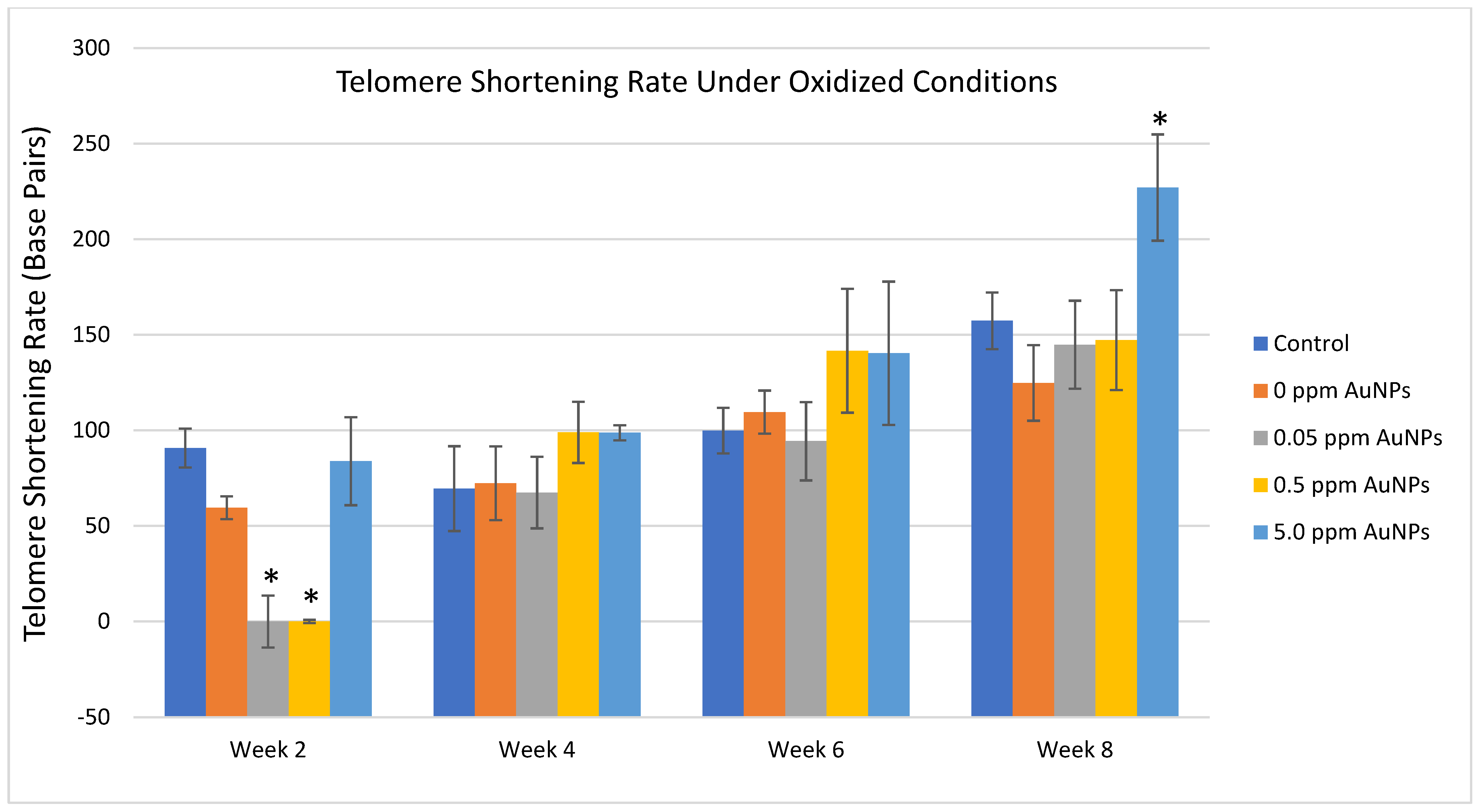
2.3. Telomere Length
3. Discussion
4. Materials and Methods
4.1. Test Articles and Cell Culture
4.2. Toxicity Assays
4.3. Telomerase Activity
4.4. Telomere Length
4.5. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chan, S.R.; Blackburn, E.H. Telomeres and telomerase. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2004, 359, 109–121. [Google Scholar] [CrossRef]
- Kim, N.W.; Piatyszek, M.A.; Prowse, K.R.; Harley, C.B.; West, M.D.; Ho, P.L.; Coviello, G.M.; Wright, W.E.; Weinrich, S.L.; Shay, J.W. Specific association of human telomerase activity with immortal cells and cancer. Science 1994, 266, 2011. [Google Scholar] [CrossRef]
- Cong, Y.S.; Wright, W.E.; Shay, J.W. Human telomerase and its regulation. Microbiol. Mol. Biol. Rev. 2002, 66, 407–425. [Google Scholar] [CrossRef]
- Victorelli, S.; Passos, J.F. Telomeres and cell senescence—Size matters not. eBioMedicine 2017, 21, 14–20. [Google Scholar] [CrossRef]
- Salpea, K.D.; Talmud, P.J.; Cooper, J.A.; Maubaret, C.G.; Stephens, J.W.; Abelak, K.; Humphries, S.E. Association of telomere length with type 2 diabetes, oxidative stress and UCP2 gene variation. Atherosclerosis 2010, 209, 42–50. [Google Scholar] [CrossRef]
- Willeit, P.; Willeit, J.; Mayr, A.; Weger, S.; Oberhollenzer, F.; Brandstatter, A.; Kronenberg, F.; Kiechl, S. Telomere length and risk of incident cancer and cancer mortality. JAMA 2010, 304, 69–75. [Google Scholar] [CrossRef]
- Wentzensen, I.M.; Mirabello, L.; Pfeiffer, R.M.; Savage, S.A. The association of telomere length and cancer: A meta-analysis. Cancer Epidemiol. Biomark. Prev. 2011, 20, 1238–1250. [Google Scholar] [CrossRef]
- Sanders, J.L.; Fitzpatrick, A.L.; Boudreau, R.M.; Arnold, A.M.; Aviv, A.; Kimura, M.; Fried, L.F.; Harris, T.B.; Newman, A.B. Leukocyte telomere length is associated with noninvasively measured agerelated disease: The Cardiovascular Health Study. J. Gerontol. A Biol. Sci. Med. Sci. 2012, 67, 409–416. [Google Scholar] [CrossRef]
- Zhao, J.; Miao, K.; Wang, H.; Ding, H.; Wang, D.W. Association between telomere length and type 2 diabetes mellitus: A meta-analysis. PLoS ONE 2013, 8, e79993. [Google Scholar] [CrossRef] [PubMed]
- von Zglinicki, T.; Saretzki, G.; Docke, W.; Lotze, C. Mild hyperoxia shortens telomeres and inhibits proliferation of fibroblasts: A model for senescence? Exp. Cell Res. 1995, 220, 186–193. [Google Scholar] [CrossRef] [PubMed]
- von Zglinicki, T. Oxidative stress shortens telomeres. Trends Biochem. Sci. 2002, 27, 339–344. [Google Scholar] [CrossRef]
- von Zglinicki, T.; Pilger, R.; Sitte, N. Accumulation of single-strand breaks is the major cause of telomere shortening in human fibroblasts. Free Radic. Biol. Med. 2000, 28, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Furumoto, K.; Inoue, E.; Nagao, N.; Hiyama, E.; Miwa, N. Age-dependent telomere shortening is slowed down by enrichment of intracellular vitamin C via suppression of oxidative stress. Life Sci. 1998, 63, 935–948. [Google Scholar] [CrossRef] [PubMed]
- Jogi, R.; Tager, M.; Perez, D.; Tsapekos, M. Bovine Colostrum, Telomeres, and Skin Aging. J. Drugs Dermatol. 2021, 20, 538–545. [Google Scholar]
- Wu, Y.-Z.; Tsai, Y.-Y.; Chang, L.-S.; Chen, Y.-J. Evaluation of Gallic Acid-Coated Gold Nanoparticles as an Anti-Aging Ingredient. Pharmaceuticals 2021, 14, 1071. [Google Scholar] [CrossRef]
- Omidifar, N.; Moghadami, M.; Mousavi, S.M.; Hashemi, S.A.; Gholami, A.; Shokripour, M.; Sohrabi, Z. Trends in Natural Nutrients for Oxidtative Stress and Cell Senescence. Oxidative Med. Cell. Longev. 2021, 2021, 7501424. [Google Scholar] [CrossRef]
- Crous-Bou, M.; Molinuevo, J.-L.; Sala-Vila, A. Plant-Rich Dietary Patterns, Plant Foods and Nutrients, and Telomere Length. Adv. Nutr. 2019, 10, S296–S303. [Google Scholar] [CrossRef] [PubMed]
- Munteanu, C.; Schwartz, B. The relationship between nutrition and the immune system. Front Nutr. 2022, 9, 1082500. [Google Scholar] [CrossRef]
- Maares, M.; Haase, H. A Guide to Human Zinc Absorption: General Overview and Recent Advances of In Vitro Intestinal Models. Nutrients 2020, 12, 762. [Google Scholar] [CrossRef] [PubMed]
- Sandstrom, B. Dose Dependence of Zinc and Manganese Absorption in Man. Proc. Nutr. Soc. 1992, 51, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Miret, S.; Simpson, R.; McKie, A. Physiology and Molecular Biology of Dietary Iron Absorption. Annu. Rev. Nutr. 2003, 23, 283–301. [Google Scholar] [CrossRef]
- Zhang, Y.; Shareena Dasari, T.; Deng, H.; Yu, H. Antimicrobial Activity of Gold Nanoparticles and Ionic Gold. J. Environ. Sci. Health 2015, 33, 286–327. [Google Scholar] [CrossRef] [PubMed]
- Jeynes, J.C.G.; Geraki, K.; Jeynes, C.; Zhaohong, M.; Bettiol, A.; Latorre, E.; Harries, L.W.; Soeller, C. Nanoscale Properties of Human Telomeres Measured with a Duel Purpose Z-ray Fluorescence and Super Resolution Microscopy Gold Nanoparticles Probe. Am. Chem. Soc. Nano 2017, 11, 12632–12640. [Google Scholar]
- Zong, S.; Chen, C.; Want, Z.; Zhang, Y.; Cui, Y. Surface Enhanced Raman Scattering Based in Situ Hybridization Strategy for Telomere Length Assessment. Am. Chem. Soc. Nano 2016, 10, 2950–2959. [Google Scholar] [CrossRef]
- Tsoukalas, D.; Fragkiadaki, P.; Docea, A.O.; Alegakis, A.; Sarandi, E.; Thanasoula, M.; Spandidos, D.; Tsaksakis, A.; Razgonova, M.P.; Calina, D. Discovery of potent telomerase activators: Unfolding new therapeutics and anti-aging perspectives. Mol. Med. Rep. 2019, 20, 3701–3708. [Google Scholar] [CrossRef] [PubMed]
- Nemoto, K.; Kondo, Y.; Himeno, S.; Suzuki, Y.; Hara, S.; Akimoto, M.; Imura, N. Modulation of telomerase activity by zinc in human prostatic and renal cancer cells. Mol. Cell. Pharmacol. 2002, 59, 401–405. [Google Scholar] [CrossRef]
- Das, A.; Bhattacharjee, P.; Bhattacharjee, P. Role of arsenic, lead and cadmium on telomere length and the risk of carcinogenesis: A mechanistic insight. Nucleus 2019, 62, 99–107. [Google Scholar] [CrossRef]
- Hornsby, P.J. Telomerase and the aging process. Exp. Gerontol. 2007, 42, 575–581. [Google Scholar] [CrossRef]
- Torre-Espinosa, Z.Y.; Barredo-Pool, F.; Castano de la Serna, E.; Sanchez-Teyer, L.F. Active telomerase during leaf growth and increase of age in plants from Agave tequilana var. Azul. Physiol. Mol. Biol. Plants 2020, 26, 639–647. [Google Scholar] [CrossRef]
- Samuel, P.; Tsapekos, M.; de Pedro, N.; Liu, A.; Lippmeier, C.; Chen, S. Ergothioneine Mitigates Telomere Shortening under Oxidative Stress Conditions. J. Diet. Suppl. 2022, 19, 212–225. [Google Scholar] [CrossRef]
- Pusceddu, I.; Farrell, C.-J.; Di Pierro, A.M.; Jani, E.; Herrmann, W.; Herrmann, M. The role of telomers and vitamin D in cellular aging and age-related diseases. Clin. Chem. Lab. Med. 2015, 53, 1661–1678. [Google Scholar] [CrossRef] [PubMed]
- Modica, V.; Glavits, R.; Murbach, T.; Endres, J.; Hirka, G.; Vertesi, A.; Beres, E.; Szakonyine, I.P. A toxicological evaluation of 8-28 nm gold nanocrystals. Food Chem. Toxicol. 2022, 161, 112844. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; El-Sayed, I.; Yi, X.; El-Sayed, M. Gold nanoparticles: Catalyst for the oxidation of NADH to NAD+. J. Photochem. Photobiol. B Biol. 2005, 81, 76–83. [Google Scholar] [CrossRef]
- Omran, H.; Almaliki, M. Influence of NAD+ as an ageing-relating immunomodulator on COVID 19 infection: A hypothesis. J. Infect. Public. Health 2020, 13, 1196–1201. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Wang, K.; Stock, A.; Gong, Y.; Demarest, T.; Yang, B.; Giri, N.; Harrington, L.; Alter, B.L.; Savage, S.; et al. Re-equilibration of imbalanced NAD metabolism ameliorates the impact of telomere dysfunction. EMBO J. 2020, 39, e103420. [Google Scholar] [CrossRef]
- Zhong, L.; Yeh, T.-Y.; Hao, J.; Pourtabatabaei, N.; Mahata, S.; Shao, J.; Chessler, S.; Chi, N.-W. Nutritional Energy Stimulates NAD+ Production to Promote Tankyrase-Mediated PARsylation in Insulinoma Cells. PLoS ONE 2015, 10, e0122948. [Google Scholar] [CrossRef]
- Miglani, M.; Pasha, Q.; Gupta, A.; Priyadarshini, A.; Pandey, R.P.; Vibhuti, A. Seeding drug discovery: Telomeric tankyrase as a pharmacological target for the pathophysiology of high-altitude hypoxia. Drug Discov. Today 2021, 26, 2774–2781. [Google Scholar] [CrossRef]
- Robinson, A.P.; Zhang, J.Z.; Titus, H.E.; Karl, M.; Merzliakov, M.; Dorfman, A.R.; Karlik, S.; Stewart, M.G.; Watt, R.K.; Facer, B.D.; et al. Nanocatalytic activity of clean-surfaced, faceted nanocrystalline gold enhances remyelination in animal models of multiple sclerosis. Sci. Rep. 2020, 10, 1936. [Google Scholar] [CrossRef]
- Huffman, K.; Levene, S.; Tesmer, V.; Shaw, J.; Wright, W. Telomere shortening is proportional to the size of the G-rich telomeric 3′-overhang. J. Biol. Chem. 2000, 275, 19719–19722. [Google Scholar] [CrossRef]
- Ait-Ghezala, G.; Hassan, S.; Tweed, M.; Paris, D.; Crynen, G.; Zakirova, Z.; Crynen, S.; Crawford, F. Identification of telomerase-activating blends from naturally occurring compounds. Altern. Ther. 2016, 22, 6–14. [Google Scholar]
- Mehkri, S.; Perez, D.; Najarro, P.; Tsapekos, M.; Bopanna, K. Effect of Lutein (Lute-gen®) on proliferation rate and telomere length in vitro and possible mechanism of action. Int. J. Biol. Res. 2019, 4, 84–91. [Google Scholar]
- Verhulst, C.; Coiffard, C.; Coiffard, L.J.; Rivalland, P.; De Roeck-Holtzhauer, Y. In vitro correlation between two colorimetric assays and the pyruvic acid consumption by fibroblasts cultured to determine the sodium laurylsulfate cytotoxicity. J. Pharmacol. Toxicol. Methods 1998, 39, 143–146. [Google Scholar] [CrossRef]
- Liu, Y.; Nair, M.G. An efficient and economical MTT assay for determining the antioxidant activity of plant natural product extracts and pure compounds. J. Nat. Prod. 2010, 73, 1193–1195. [Google Scholar] [CrossRef] [PubMed]
- Van Kaspers, G.; Cloos, J. Cell sensitivity assays: The MTT assay. Methods Mol. Biol. 2011, 731, 237–245. [Google Scholar]
- Riss, T.; Moravec, R.; Niles, A.; Duellman, S.; Benink, H.; Worzella, T.; Minor, L. Cell Viability Assay. In Assay Guidance Manual; Eli Lilly & Company; the National Center for Advancing Translational Sciences: Bethesda, MD, USA, 2013. [Google Scholar]
- Hou, M.; Xu, D.; Bjorkholm, M.; Gruber, A. Real-time quantitative telomeric repeat amplification protocol assay for the detection of telomerase activity. Clin. Chem. 2001, 47, 519–524. [Google Scholar] [CrossRef]
- Wege, H.; Chui, M.S.; Le, H.T.; Tran, J.M.; Zern, M.A. SYBR Green real-time telomeric repeat amplification protocol for the rapid quantification of telomerase activity. Nucleic Acids Res. 2003, 31, e3. [Google Scholar] [CrossRef]
- Herbert, B.S.; Hochreiter, A.E.; Wright, W.E.; Shay, J.W. Nonradioactive detection of telomerase activity using the telomeric repeat amplification protocol. Nat. Protoc. 2006, 1, 1583–1590. [Google Scholar] [CrossRef] [PubMed]
- De Pedro, N.; Diez, M.; Garcia, I.; Garcia, J.; Otero, L.; Fernandez, L.; Garcia, B.; Gonzalez, R.; Rincon, S.; Perez, D.; et al. Analytical validation of telomere analysis technology® for the high-throughput analysis of m ultiple telomere-associated variables. Biol. Proced. Online 2020, 22, 2. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Dose | Compound |
|---|---|
| 0 ppm | Control |
| 0 ppm | Control, with AuNP buffer |
| 0.125 ppm | AuNPs |
| 1.25 ppm | AuNPs |
| 12.5 ppm | AuNPs |
| Treatment Dose | Compound |
|---|---|
| 0 ppm | Control, no treatment |
| 0 ppm | Control, with AuNP buffer |
| 0.05 ppm | AuNP |
| 0.5 ppm | AuNP |
| 5.0 ppm | AuNP |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, X.; Hirschel, A.; Tsapekos, M.; Perez, D.; Vollmer, D. In Vitro Assessment of Gold Nanoparticles on Telomerase Activity and Telomere Length in Human Fibroblasts. Int. J. Mol. Sci. 2023, 24, 14273. https://doi.org/10.3390/ijms241814273
Han X, Hirschel A, Tsapekos M, Perez D, Vollmer D. In Vitro Assessment of Gold Nanoparticles on Telomerase Activity and Telomere Length in Human Fibroblasts. International Journal of Molecular Sciences. 2023; 24(18):14273. https://doi.org/10.3390/ijms241814273
Chicago/Turabian StyleHan, Xuesheng, Alice Hirschel, Menelaos Tsapekos, Diego Perez, and David Vollmer. 2023. "In Vitro Assessment of Gold Nanoparticles on Telomerase Activity and Telomere Length in Human Fibroblasts" International Journal of Molecular Sciences 24, no. 18: 14273. https://doi.org/10.3390/ijms241814273
APA StyleHan, X., Hirschel, A., Tsapekos, M., Perez, D., & Vollmer, D. (2023). In Vitro Assessment of Gold Nanoparticles on Telomerase Activity and Telomere Length in Human Fibroblasts. International Journal of Molecular Sciences, 24(18), 14273. https://doi.org/10.3390/ijms241814273