
Red Blood Cell Fatty Acid Profiles Are Significantly Altered in South Australian Mild Cognitive Impairment and Alzheimer’s Disease Cases Compared to Matched Controls


Abstract
:1. Introduction
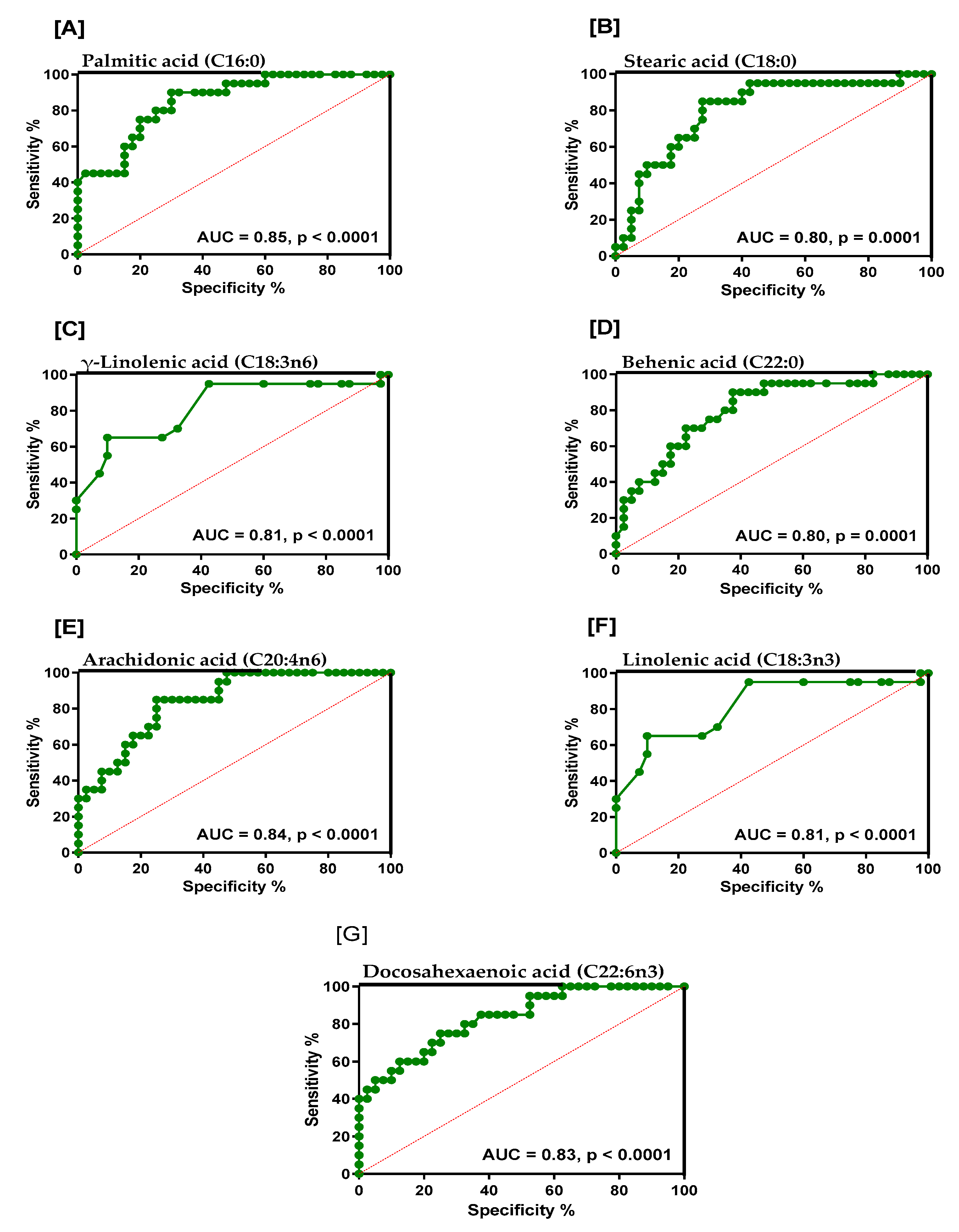
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Design, Recruitment and Characteristics of Participants
4.2. Blood Collection
4.3. Fatty Acid Analysis
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blazer, D.G.; Yaffe, K.; Karlawish, J. Cognitive aging: A report from the Institute of Medicine. JAMA 2015, 313, 2121–2122. [Google Scholar] [CrossRef] [PubMed]
- Juan, S.M.A.; Adlard, P.A. Ageing and Cognition. Subcell. Biochem. 2019, 91, 107–122. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, T.; Arai, H.; Sakurai, T. An update on cognitive frailty: Its definition, impact, associated factors and underlying mechanisms, and interventions. Geriatr. Gerontol. Int. 2022, 22, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Feldman, H.H.; Jacova, C. Mild cognitive impairment. Am. J. Geriatr. Psychiatry 2005, 13, 645–655. [Google Scholar] [CrossRef]
- Gauthier, S.; Reisberg, B.; Zaudig, M.; Petersen, R.C.; Ritchie, K.; Broich, K.; Belleville, S.; Brodaty, H.; Bennett, D.; Chertkow, H.; et al. Mild cognitive impairment. Lancet 2006, 367, 1262–1270. [Google Scholar] [CrossRef]
- Grundman, M.; Petersen, R.C.; Ferris, S.H.; Thomas, R.G.; Aisen, P.S.; Bennett, D.A.; Foster, N.L.; Jack, C.R., Jr.; Galasko, D.R.; Doody, R.; et al. Mild cognitive impairment can be distinguished from Alzheimer disease and normal aging for clinical trials. Arch. Neurol. 2004, 61, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Petersen, R.C. Mild cognitive impairment: Transition between aging and Alzheimer’s disease. Neurologia 2000, 15, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.; O’Connell, T.; Johnson, S.; Cline, S.; Merikle, E.; Martenyi, F.; Simpson, K. Estimating Alzheimer’s Disease Progression Rates from Normal Cognition Through Mild Cognitive Impairment and Stages of Dementia. Curr. Alzheimer Res. 2018, 15, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Burns, A.; Byrne, E.J.; Maurer, K. Alzheimer’s disease. Lancet 2002, 360, 163–165. [Google Scholar] [CrossRef] [PubMed]
- Kawas, C.H. Clinical practice. Early Alzheimer’s disease. N. Engl. J. Med. 2003, 349, 1056–1063. [Google Scholar] [CrossRef]
- Mattson, M.P. Pathways towards and away from Alzheimer’s disease. Nature 2004, 430, 631–639. [Google Scholar] [CrossRef] [PubMed]
- St George-Hyslop, P.H. Piecing together Alzheimer’s. Sci. Am. 2000, 283, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Scheltens, P.; De Strooper, B.; Kivipelto, M.; Holstege, H.; Chételat, G.; Teunissen, C.E.; Cummings, J.; van der Flier, W.M. Alzheimer’s disease. Lancet 2021, 397, 1577–1590. [Google Scholar] [CrossRef] [PubMed]
- Batsch, N.L.; Mittelman, M.S. Overcoming the Stigma of Dementa; Alzheimer’s Disease International (ADI): London, UK, 2012; 80p. [Google Scholar]
- Australian Institute of Health and Welfare. Dementia in Australia: Summary Report 2022; Australian Institute of Health and Welfare: Canberra, Australia, 2023; 38p.
- Jama, J.W.; Launer, L.J.; Witteman, J.C.; den Breeijen, J.H.; Breteler, M.M.; Grobbee, D.E.; Hofman, A. Dietary antioxidants and cognitive function in a population-based sample of older persons. The Rotterdam Study. Am. J. Epidemiol. 1996, 144, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Dye, L.; Lluch, A.; Blundell, J.E. Macronutrients and mental performance. Nutrition 2000, 16, 1021–1034. [Google Scholar] [CrossRef] [PubMed]
- González-Gross, M.; Marcos, A.; Pietrzik, K. Nutrition and cognitive impairment in the elderly. Br. J. Nutr. 2001, 86, 313–321. [Google Scholar] [CrossRef]
- Chu, C.Q.; Yu, L.L.; Qi, G.Y.; Mi, Y.S.; Wu, W.Q.; Lee, Y.K.; Zhai, Q.X.; Tian, F.W.; Chen, W. Can dietary patterns prevent cognitive impairment and reduce Alzheimer’s disease risk: Exploring the underlying mechanisms of effects. Neurosci. Biobehav. Rev. 2022, 135, 104556. [Google Scholar] [CrossRef]
- Ellouze, I.; Sheffler, J.; Nagpal, R.; Arjmandi, B. Dietary Patterns and Alzheimer’s Disease: An Updated Review Linking Nutrition to Neuroscience. Nutrients 2023, 15, 3204. [Google Scholar] [CrossRef]
- Haag, M. Essential fatty acids and the brain. Can. J. Psychiatry 2003, 48, 195–203. [Google Scholar] [CrossRef]
- Heude, B.; Ducimetière, P.; Berr, C. Cognitive decline and fatty acid composition of erythrocyte membranes—The EVA Study. Am. J. Clin. Nutr. 2003, 77, 803–808. [Google Scholar] [CrossRef] [PubMed]
- Martinat, M.; Rossitto, M.; Di Miceli, M.; Layé, S. Perinatal Dietary Polyunsaturated Fatty Acids in Brain Development, Role in Neurodevelopmental Disorders. Nutrients 2021, 13, 1185. [Google Scholar] [CrossRef]
- Kyle, D.J.; Schaefer, E.; Patton, G.; Beiser, A. Low serum docosahexaenoic acid is a significant risk factor for Alzheimer’s dementia. Lipids 1999, 34 (Suppl. 1), S245. [Google Scholar] [CrossRef] [PubMed]
- Conquer, J.A.; Tierney, M.C.; Zecevic, J.; Bettger, W.J.; Fisher, R.H. Fatty acid analysis of blood plasma of patients with Alzheimer’s disease, other types of dementia, and cognitive impairment. Lipids 2000, 35, 1305–1312. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.Y.; Cheng, C.; Satyanarayanan, S.K.; Chiu, L.T.; Chien, Y.C.; Chuu, C.P.; Lan, T.H.; Su, K.P. Omega-3 fatty acids and blood-based biomarkers in Alzheimer’s disease and mild cognitive impairment: A randomized placebo-controlled trial. Brain Behav. Immun. 2022, 99, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Cooke, S.F.; Bliss, T.V. Plasticity in the human central nervous system. Brain 2006, 129, 1659–1673. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.Y.; Suzuki, H. Intakes of dietary docosahexaenoic acid ethyl ester and egg phosphatidylcholine improve maze-learning ability in young and old mice. J. Nutr. 2000, 130, 1629–1632. [Google Scholar] [CrossRef]
- Berr, C.; Portet, F.; Carriere, I.; Akbaraly, T.N.; Feart, C.; Gourlet, V.; Combe, N.; Barberger-Gateau, P.; Ritchie, K. Olive oil and cognition: Results from the three-city study. Dement. Geriatr. Cogn. Disord. 2009, 28, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Solfrizzi, V.; Colacicco, A.M.; D’Introno, A.; Capurso, C.; Torres, F.; Rizzo, C.; Capurso, A.; Panza, F. Dietary intake of unsaturated fatty acids and age-related cognitive decline: A 8.5-year follow-up of the Italian Longitudinal Study on Aging. Neurobiol. Aging 2006, 27, 1694–1704. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.C.; Evans, D.A.; Bienias, J.L.; Tangney, C.C.; Wilson, R.S. Dietary fat intake and 6-year cognitive change in an older biracial community population. Neurology 2004, 62, 1573–1579. [Google Scholar] [CrossRef] [PubMed]
- Kalmijn, S.; van Boxtel, M.P.; Ocké, M.; Verschuren, W.M.; Kromhout, D.; Launer, L.J. Dietary intake of fatty acids and fish in relation to cognitive performance at middle age. Neurology 2004, 62, 275–280. [Google Scholar] [CrossRef]
- Ballarini, T.; Melo van Lent, D.; Brunner, J.; Schröder, A.; Wolfsgruber, S.; Altenstein, S.; Brosseron, F.; Buerger, K.; Dechent, P.; Dobisch, L.; et al. Mediterranean Diet, Alzheimer Disease Biomarkers and Brain Atrophy in Old Age. Neurology 2021, 96, e2920–e2932. [Google Scholar] [CrossRef] [PubMed]
- Ogbodo, J.O.; Agbo, C.P.; Njoku, U.O.; Ogugofor, M.O.; Egba, S.I.; Ihim, S.A.; Echezona, A.C.; Brendan, K.C.; Upaganlawar, A.B.; Upasani, C.D. Alzheimer’s Disease: Pathogenesis and Therapeutic Interventions. Curr. Aging Sci. 2022, 15, 2–25. [Google Scholar] [CrossRef]
- Arab, L. Biomarkers of fat and fatty acid intake. J. Nutr. 2003, 133 (Suppl. 3), 925s–932s. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.S.; Harris, W.S.; Beiser, A.S.; Au, R.; Himali, J.J.; Debette, S.; Pikula, A.; Decarli, C.; Wolf, P.A.; Vasan, R.S.; et al. Red blood cell ω-3 fatty acid levels and markers of accelerated brain aging. Neurology 2012, 78, 658–664. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.S.; Sands, S.A.; Windsor, S.L.; Ali, H.A.; Stevens, T.L.; Magalski, A.; Porter, C.B.; Borkon, A.M. Omega-3 fatty acids in cardiac biopsies from heart transplantation patients: Correlation with erythrocytes and response to supplementation. Circulation 2004, 110, 1645–1649. [Google Scholar] [CrossRef] [PubMed]
- Hooper, C.; De Souto Barreto, P.; Pahor, M.; Weiner, M.; Vellas, B. The Relationship of Omega 3 Polyunsaturated Fatty Acids in Red Blood Cell Membranes with Cognitive Function and Brain Structure: A Review Focussed on Alzheimer’s Disease. J. Prev. Alzheimers Dis. 2018, 5, 78–84. [Google Scholar] [CrossRef] [PubMed]
- von Schacky, C. Importance of EPA and DHA Blood Levels in Brain Structure and Function. Nutrients 2021, 13, 1074. [Google Scholar] [CrossRef] [PubMed]
- Martinsen, A.; Saleh, R.N.M.; Chouinard-Watkins, R.; Bazinet, R.; Harden, G.; Dick, J.; Tejera, N.; Pontifex, M.G.; Vauzour, D.; Minihane, A.M. The Influence of APOE Genotype, DHA, and Flavanol Intervention on Brain DHA and Lipidomics Profile in Aged Transgenic Mice. Nutrients 2023, 15, 2032. [Google Scholar] [CrossRef]
- Bai, W.; Chen, P.; Cai, H.; Zhang, Q.; Su, Z.; Cheung, T.; Jackson, T.; Sha, S.; Xiang, Y.T. Worldwide prevalence of mild cognitive impairment among community dwellers aged 50 years and older: A meta-analysis and systematic review of epidemiology studies. Age Ageing 2022, 51, afac173. [Google Scholar] [CrossRef]
- Panza, F.; Frisardi, V.; Seripa, D.; Imbimbo, B.P.; Pilotto, A.; Solfrizzi, V. Dietary unsaturated fatty acids and risk of mild cognitive impairment. J. Alzheimers Dis. 2010, 21, 867–870. [Google Scholar] [CrossRef] [PubMed]
- Solfrizzi, V.; Frisardi, V.; Capurso, C.; D’Introno, A.; Colacicco, A.M.; Vendemiale, G.; Capurso, A.; Panza, F. Dietary fatty acids in dementia and predementia syndromes: Epidemiological evidence and possible underlying mechanisms. Ageing Res. Rev. 2010, 9, 184–199. [Google Scholar] [CrossRef] [PubMed]
- Solfrizzi, V.; Capurso, C.; D’Introno, A.; Colacicco, A.M.; Frisardi, V.; Santamato, A.; Ranieri, M.; Fiore, P.; Vendemiale, G.; Seripa, D.; et al. Dietary fatty acids, age-related cognitive decline, and mild cognitive impairment. J. Nutr. Health Aging 2008, 12, 382–386. [Google Scholar] [CrossRef]
- Fan, R.; Zhao, L.; Ding, B.J.; Xiao, R.; Ma, W.W. The association of blood non-esterified fatty acid, saturated fatty acids, and polyunsaturated fatty acids levels with mild cognitive impairment in Chinese population aged 35–64 years: A cross-sectional study. Nutr. Neurosci. 2021, 24, 148–160. [Google Scholar] [CrossRef] [PubMed]
- Currenti, W.; Godos, J.; Alanazi, A.M.; Lanza, G.; Ferri, R.; Caraci, F.; Grosso, G.; Galvano, F.; Castellano, S. Dietary Fats and Cognitive Status in Italian Middle-Old Adults. Nutrients 2023, 15, 1429. [Google Scholar] [CrossRef]
- Luchsinger, J.A.; Tang, M.X.; Shea, S.; Mayeux, R. Caloric intake and the risk of Alzheimer disease. Arch. Neurol. 2002, 59, 1258–1263. [Google Scholar] [CrossRef] [PubMed]
- Kalmijn, S.; Launer, L.J.; Ott, A.; Witteman, J.C.; Hofman, A.; Breteler, M.M. Dietary fat intake and the risk of incident dementia in the Rotterdam Study. Ann. Neurol. 1997, 42, 776–782. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.C.; Evans, D.A.; Bienias, J.L.; Tangney, C.C.; Bennett, D.A.; Wilson, R.S.; Aggarwal, N.; Schneider, J. Consumption of fish and n-3 fatty acids and risk of incident Alzheimer disease. Arch. Neurol. 2003, 60, 940–946. [Google Scholar] [CrossRef] [PubMed]
- Eskelinen, M.H.; Ngandu, T.; Helkala, E.L.; Tuomilehto, J.; Nissinen, A.; Soininen, H.; Kivipelto, M. Fat intake at midlife and cognitive impairment later in life: A population-based CAIDE study. Int. J. Geriatr. Psychiatry 2008, 23, 741–747. [Google Scholar] [CrossRef]
- Albrahim, T. The potential role of nutritional components in improving brain function among patients with Alzheimers disease: A meta-analysis of RCT studies. Neurosciences 2020, 25, 4–17. [Google Scholar] [CrossRef]
- Shen, J.; Li, J.; Hua, Y.; Ding, B.; Zhou, C.; Yu, H.; Xiao, R.; Ma, W. Association between the Erythrocyte Membrane Fatty Acid Profile and Cognitive Function in the Overweight and Obese Population Aged from 45 to 75 Years Old. Nutrients 2022, 14, 914. [Google Scholar] [CrossRef] [PubMed]
- Olde Rikkert, M.G.; Verhey, F.R.; Sijben, J.W.; Bouwman, F.H.; Dautzenberg, P.L.; Lansink, M.; Sipers, W.M.; van Asselt, D.Z.; van Hees, A.M.; Stevens, M.; et al. Differences in nutritional status between very mild Alzheimer’s disease patients and healthy controls. J. Alzheimers Dis. 2014, 41, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Solfrizzi, V.; Capurso, C.; D’Introno, A.; Colacicco, A.M.; Santamato, A.; Ranieri, M.; Fiore, P.; Capurso, A.; Panza, F. Lifestyle-related factors in predementia and dementia syndromes. Expert Rev. Neurother. 2008, 8, 133–158. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, C.E.; Winocur, G. High-fat diets, insulin resistance and declining cognitive function. Neurobiol. Aging 2005, 26 (Suppl. 1), 42–45. [Google Scholar] [CrossRef]
- Oksman, M.; Iivonen, H.; Hogyes, E.; Amtul, Z.; Penke, B.; Leenders, I.; Broersen, L.; Lütjohann, D.; Hartmann, T.; Tanila, H. Impact of different saturated fatty acid, polyunsaturated fatty acid and cholesterol containing diets on beta-amyloid accumulation in APP/PS1 transgenic mice. Neurobiol. Dis. 2006, 23, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Dong, F.; Ren, J.; Driscoll, M.J.; Culver, B. High dietary fat induces NADPH oxidase-associated oxidative stress and inflammation in rat cerebral cortex. Exp. Neurol. 2005, 191, 318–325. [Google Scholar] [CrossRef]
- Hill, M.; Takechi, R.; Chaliha, D.R.; Albrecht, M.A.; Wright, J.; James, A.P.; Clark, K.; Dhaliwal, S.S.; Lam, V.; Mamo, J.C.L. Dietary saturated fats and apolipoprotein B48 levels are similarly associated with cognitive decline in healthy older aged Australians. Asia Pac. J. Clin. Nutr. 2020, 29, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Molteni, R.; Barnard, R.J.; Ying, Z.; Roberts, C.K.; Gómez-Pinilla, F. A high-fat, refined sugar diet reduces hippocampal brain-derived neurotrophic factor, neuronal plasticity, and learning. Neuroscience 2002, 112, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, A.Z.; Harty, B.; Mukamal, K.J.; Stoddard, A.M.; Vitolins, M.; Dunn, J.E. Monounsaturated, trans, and saturated Fatty acids and cognitive decline in women. J. Am. Geriatr. Soc. 2011, 59, 837–843. [Google Scholar] [CrossRef]
- Shao, F.; Ford, D.A. Elaidic acid increases hepatic lipogenesis by mediating sterol regulatory element binding protein-1c activity in HuH-7 cells. Lipids 2014, 49, 403–413. [Google Scholar] [CrossRef]
- Park, K.H.; Kim, J.M.; Cho, K.H. Elaidic acid (EA) generates dysfunctional high-density lipoproteins and consumption of EA exacerbates hyperlipidemia and fatty liver change in zebrafish. Mol. Nutr. Food Res. 2014, 58, 1537–1545. [Google Scholar] [CrossRef]
- Nazari, B.; Asgary, S.; Azadbakht, L. Fatty acid analysis of Iranian junk food, dairy, and bakery products: Special attention to trans-fats. J. Res. Med. Sci. 2012, 17, 952–957. [Google Scholar] [PubMed]
- Vendel Nielsen, L.; Krogager, T.P.; Young, C.; Ferreri, C.; Chatgilialoglu, C.; Nørregaard Jensen, O.; Enghild, J.J. Effects of elaidic acid on lipid metabolism in HepG2 cells, investigated by an integrated approach of lipidomics, transcriptomics and proteomics. PLoS ONE 2013, 8, e74283. [Google Scholar] [CrossRef] [PubMed]
- Katarey, D.; Francis, I. Reduction of trans-fatty acids from food. JAMA 2012, 308, 1857–1858; author reply 1858–1859. [Google Scholar] [CrossRef] [PubMed]
- Chajès, V.; Biessy, C.; Byrnes, G.; Deharveng, G.; Saadatian-Elahi, M.; Jenab, M.; Peeters, P.H.; Ocké, M.; Bueno-de-Mesquita, H.B.; Johansson, I.; et al. Ecological-level associations between highly processed food intakes and plasma phospholipid elaidic acid concentrations: Results from a cross-sectional study within the European prospective investigation into cancer and nutrition (EPIC). Nutr. Cancer 2011, 63, 1235–1250. [Google Scholar] [CrossRef] [PubMed]
- Honda, T.; Ohara, T.; Shinohara, M.; Hata, J.; Toh, R.; Yoshida, D.; Shibata, M.; Ishida, T.; Hirakawa, Y.; Irino, Y.; et al. Serum elaidic acid concentration and risk of dementia: The Hisayama Study. Neurology 2019, 93, e2053–e2064. [Google Scholar] [CrossRef]
- Briante, R.; Febbraio, F.; Nucci, R. Antioxidant properties of low molecular weight phenols present in the mediterranean diet. J. Agric. Food Chem. 2003, 51, 6975–6981. [Google Scholar] [CrossRef]
- Vitaglione, P.; Savarese, M.; Paduano, A.; Scalfi, L.; Fogliano, V.; Sacchi, R. Healthy virgin olive oil: A matter of bitterness. Crit. Rev. Food Sci. Nutr. 2015, 55, 1808–1818. [Google Scholar] [CrossRef]
- Tully, A.M.; Roche, H.M.; Doyle, R.; Fallon, C.; Bruce, I.; Lawlor, B.; Coakley, D.; Gibney, M.J. Low serum cholesteryl ester-docosahexaenoic acid levels in Alzheimer’s disease: A case-control study. Br. J. Nutr. 2003, 89, 483–489. [Google Scholar] [CrossRef]
- Roberts, R.O.; Cerhan, J.R.; Geda, Y.E.; Knopman, D.S.; Cha, R.H.; Christianson, T.J.; Pankratz, V.S.; Ivnik, R.J.; O’Connor, H.M.; Petersen, R.C. Polyunsaturated fatty acids and reduced odds of MCI: The Mayo Clinic Study of Aging. J. Alzheimers Dis. 2010, 21, 853–865. [Google Scholar] [CrossRef]
- Cherubini, A.; Andres-Lacueva, C.; Martin, A.; Lauretani, F.; Iorio, A.D.; Bartali, B.; Corsi, A.; Bandinelli, S.; Mattson, M.P.; Ferrucci, L. Low plasma N-3 fatty acids and dementia in older persons: The InCHIANTI study. J. Gerontol. A Biol. Sci. Med. Sci. 2007, 62, 1120–1126. [Google Scholar] [CrossRef]
- Samieri, C.; Féart, C.; Letenneur, L.; Dartigues, J.F.; Pérès, K.; Auriacombe, S.; Peuchant, E.; Delcourt, C.; Barberger-Gateau, P. Low plasma eicosapentaenoic acid and depressive symptomatology are independent predictors of dementia risk. Am. J. Clin. Nutr. 2008, 88, 714–721. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.C.; Evans, D.A.; Tangney, C.C.; Bienias, J.L.; Wilson, R.S. Fish consumption and cognitive decline with age in a large community study. Arch. Neurol. 2005, 62, 1849–1853. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.L.; Zandi, P.P.; Tucker, K.L.; Fitzpatrick, A.L.; Kuller, L.H.; Fried, L.P.; Burke, G.L.; Carlson, M.C. Benefits of fatty fish on dementia risk are stronger for those without APOE epsilon4. Neurology 2005, 65, 1409–1414. [Google Scholar] [CrossRef]
- Dullemeijer, C.; Durga, J.; Brouwer, I.A.; van de Rest, O.; Kok, F.J.; Brummer, R.J.; van Boxtel, M.P.; Verhoef, P. n 3 fatty acid proportions in plasma and cognitive performance in older adults. Am. J. Clin. Nutr. 2007, 86, 1479–1485. [Google Scholar] [CrossRef]
- Tosi, F.; Sartori, F.; Guarini, P.; Olivieri, O.; Martinelli, N. Delta-5 and delta-6 desaturases: Crucial enzymes in polyunsaturated fatty acid-related pathways with pleiotropic influences in health and disease. Adv. Exp. Med. Biol. 2014, 824, 61–81. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.S.; Hung, C.F.; Ponnusamy, V.K.; Chen, K.C.; Chen, N.C. Higher Serum DHA and Slower Cognitive Decline in Patients with Alzheimer’s Disease: Two-Year Follow-Up. Nutrients 2022, 14, 1159. [Google Scholar] [CrossRef] [PubMed]
- Ammann, E.M.; Pottala, J.V.; Robinson, J.G.; Espeland, M.A.; Harris, W.S. Erythrocyte omega-3 fatty acids are inversely associated with incident dementia: Secondary analyses of longitudinal data from the Women’s Health Initiative Memory Study (WHIMS). Prostaglandins Leukot. Essent. Fatty Acids 2017, 121, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Rouch, L.; Virecoulon Giudici, K.; Cantet, C.; Guyonnet, S.; Delrieu, J.; Legrand, P.; Catheline, D.; Andrieu, S.; Weiner, M.; de Souto Barreto, P.; et al. Associations of erythrocyte omega-3 fatty acids with cognition, brain imaging and biomarkers in the Alzheimer’s disease neuroimaging initiative: Cross-sectional and longitudinal retrospective analyses. Am. J. Clin. Nutr. 2022, 116, 1492–1506. [Google Scholar] [CrossRef] [PubMed]
- Welty, F.K. Omega-3 fatty acids and cognitive function. Curr. Opin. Lipidol. 2023, 34, 12–21. [Google Scholar] [CrossRef]
- He, K.; Rimm, E.B.; Merchant, A.; Rosner, B.A.; Stampfer, M.J.; Willett, W.C.; Ascherio, A. Fish consumption and risk of stroke in men. JAMA 2002, 288, 3130–3136. [Google Scholar] [CrossRef]
- Lim, W.S.; Gammack, J.K.; Van Niekerk, J.; Dangour, A.D. Omega 3 fatty acid for the prevention of dementia. Cochrane Database Syst. Rev. 2006, 2016, Cd005379. [Google Scholar] [CrossRef]
- Hofman, A.; Ott, A.; Breteler, M.M.; Bots, M.L.; Slooter, A.J.; van Harskamp, F.; van Duijn, C.N.; Van Broeckhoven, C.; Grobbee, D.E. Atherosclerosis, apolipoprotein E, and prevalence of dementia and Alzheimer’s disease in the Rotterdam Study. Lancet 1997, 349, 151–154. [Google Scholar] [CrossRef]
- Akiyama, H.; Barger, S.; Barnum, S.; Bradt, B.; Bauer, J.; Cole, G.M.; Cooper, N.R.; Eikelenboom, P.; Emmerling, M.; Fiebich, B.L.; et al. Inflammation and Alzheimer’s disease. Neurobiol. Aging 2000, 21, 383–421. [Google Scholar] [CrossRef] [PubMed]
- Vesga-Jiménez, D.J.; Martin, C.; Barreto, G.E.; Aristizábal-Pachón, A.F.; Pinzón, A.; González, J. Fatty Acids: An Insight into the Pathogenesis of Neurodegenerative Diseases and Therapeutic Potential. Int. J. Mol. Sci. 2022, 23, 2577. [Google Scholar] [CrossRef]
- Custers; Emma, E.M.; Kiliaan; Amanda, J. Dietary lipids from body to brain. Prog. Lipid Res. 2022, 85, 101144. [Google Scholar] [CrossRef] [PubMed]
- Friedland, R.P. Fish consumption and the risk of Alzheimer disease: Is it time to make dietary recommendations? Arch. Neurol. 2003, 60, 923–924. [Google Scholar] [CrossRef]
- Grimm, M.O.; Kuchenbecker, J.; Grösgen, S.; Burg, V.K.; Hundsdörfer, B.; Rothhaar, T.L.; Friess, P.; de Wilde, M.C.; Broersen, L.M.; Penke, B.; et al. Docosahexaenoic acid reduces amyloid beta production via multiple pleiotropic mechanisms. J. Biol. Chem. 2011, 286, 14028–14039. [Google Scholar] [CrossRef] [PubMed]
- Grimm, M.O.; Haupenthal, V.J.; Mett, J.; Stahlmann, C.P.; Blümel, T.; Mylonas, N.T.; Endres, K.; Grimm, H.S.; Hartmann, T. Oxidized Docosahexaenoic Acid Species and Lipid Peroxidation Products Increase Amyloidogenic Amyloid Precursor Protein Processing. Neurodegener. Dis. 2016, 16, 44–54. [Google Scholar] [CrossRef]
- Puskás, L.G.; Kitajka, K.; Nyakas, C.; Barcelo-Coblijn, G.; Farkas, T. Short-term administration of omega 3 fatty acids from fish oil results in increased transthyretin transcription in old rat hippocampus. Proc. Natl. Acad. Sci. USA 2003, 100, 1580–1585. [Google Scholar] [CrossRef]
- Green, K.N.; Martinez-Coria, H.; Khashwji, H.; Hall, E.B.; Yurko-Mauro, K.A.; Ellis, L.; LaFerla, F.M. Dietary docosahexaenoic acid and docosapentaenoic acid ameliorate amyloid-beta and tau pathology via a mechanism involving presenilin 1 levels. J. Neurosci. 2007, 27, 4385–4395. [Google Scholar] [CrossRef] [PubMed]
- Koch, M.; Furtado, J.D.; DeKosky, S.T.; Fitzpatrick, A.L.; Lopez, O.L.; Kuller, L.H.; Mukamal, K.J.; Jensen, M.K. Case-cohort study of plasma phospholipid fatty acid profiles, cognitive function, and risk of dementia: A secondary analysis in the Ginkgo Evaluation of Memory Study. Am. J. Clin. Nutr. 2021, 114, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Eriksdotter, M.; Vedin, I.; Falahati, F.; Freund-Levi, Y.; Hjorth, E.; Faxen-Irving, G.; Wahlund, L.O.; Schultzberg, M.; Basun, H.; Cederholm, T.; et al. Plasma Fatty Acid Profiles in Relation to Cognition and Gender in Alzheimer’s Disease Patients during Oral Omega-3 Fatty Acid Supplementation: The OmegAD Study. J. Alzheimers Dis. 2015, 48, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.G.; Ijioma, N.; Harris, W. Omega-3 fatty acids and cognitive function in women. Womens Health 2010, 6, 119–134. [Google Scholar] [CrossRef] [PubMed]
- Amminger, G.P.; Schäfer, M.R.; Klier, C.M.; Slavik, J.M.; Holzer, I.; Holub, M.; Goldstone, S.; Whitford, T.J.; McGorry, P.D.; Berk, M. Decreased nervonic acid levels in erythrocyte membranes predict psychosis in help-seeking ultra-high-risk individuals. Mol. Psychiatry 2012, 17, 1150–1152. [Google Scholar] [CrossRef] [PubMed]
- Phung, N.V.; Rong, F.; Xia, W.Y.; Fan, Y.; Li, X.Y.; Wang, S.A.; Li, F.L. Nervonic acid and its sphingolipids: Biological functions and potential food applications. Crit. Rev. Food Sci. Nutr. 2023, 1–20. [Google Scholar] [CrossRef]
- Oda, E.; Hatada, K.; Kimura, J.; Aizawa, Y.; Thanikachalam, P.V.; Watanabe, K. Relationships between serum unsaturated fatty acids and coronary risk factors: Negative relations between nervonic acid and obesity-related risk factors. Int. Heart J. 2005, 46, 975–985. [Google Scholar] [CrossRef]
- Barberger-Gateau, P.; Letenneur, L.; Deschamps, V.; Pérès, K.; Dartigues, J.F.; Renaud, S. Fish, meat, and risk of dementia: Cohort study. BMJ 2002, 325, 932–933. [Google Scholar] [CrossRef]
- Barberger-Gateau, P.; Samieri, C.; Féart, C.; Plourde, M. Dietary omega 3 polyunsaturated fatty acids and Alzheimer’s disease: Interaction with apolipoprotein E genotype. Curr. Alzheimer Res. 2011, 8, 479–491. [Google Scholar] [CrossRef]
- Beydoun, M.A.; Kaufman, J.S.; Satia, J.A.; Rosamond, W.; Folsom, A.R. Plasma n-3 fatty acids and the risk of cognitive decline in older adults: The Atherosclerosis Risk in Communities Study. Am. J. Clin. Nutr. 2007, 85, 1103–1111. [Google Scholar] [CrossRef]
- Laurin, D.; Verreault, R.; Lindsay, J.; Dewailly, E.; Holub, B.J. Omega-3 fatty acids and risk of cognitive impairment and dementia. J. Alzheimers Dis. 2003, 5, 315–322. [Google Scholar] [CrossRef]
- Ishihara, K.; Izawa, K.P.; Kitamura, M.; Shimogai, T.; Kanejima, Y.; Morisawa, T.; Shimizu, I. Serum concentration of dihomo-γ-linolenic acid is associated with cognitive function and mild cognitive impairment in coronary artery disease patients. Prostaglandins Leukot. Essent. Fatty Acids 2020, 158, 102038. [Google Scholar] [CrossRef] [PubMed]
- Kanoski, S.E.; Davidson, T.L. Western diet consumption and cognitive impairment: Links to hippocampal dysfunction and obesity. Physiol. Behav. 2011, 103, 59–68. [Google Scholar] [CrossRef]
- Stoll, B.A. Western diet, early puberty, and breast cancer risk. Breast Cancer Res. Treat. 1998, 49, 187–193. [Google Scholar] [CrossRef]
- Morris, M.C.; Evans, D.A.; Bienias, J.L.; Tangney, C.C.; Bennett, D.A.; Aggarwal, N.; Schneider, J.; Wilson, R.S. Dietary fats and the risk of incident Alzheimer disease. Arch. Neurol. 2003, 60, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.C.; Evans, D.A.; Tangney, C.C.; Bienias, J.L.; Schneider, J.A.; Wilson, R.S.; Scherr, P.A. Dietary copper and high saturated and trans fat intakes associated with cognitive decline. Arch. Neurol. 2006, 63, 1085–1088. [Google Scholar] [CrossRef]
- Stefaniak, O.; Dobrzyńska, M.; Drzymała-Czyż, S.; Przysławski, J. Diet in the Prevention of Alzheimer’s Disease: Current Knowledge and Future Research Requirements. Nutrients 2022, 14, 4564. [Google Scholar] [CrossRef] [PubMed]
- Więckowska-Gacek, A.; Mietelska-Porowska, A.; Wydrych, M.; Wojda, U. Western diet as a trigger of Alzheimer’s disease: From metabolic syndrome and systemic inflammation to neuroinflammation and neurodegeneration. Ageing Res. Rev. 2021, 70, 101397. [Google Scholar] [CrossRef] [PubMed]
- Solfrizzi, V.; Panza, F.; Torres, F.; Mastroianni, F.; Del Parigi, A.; Venezia, A.; Capurso, A. High monounsaturated fatty acids intake protects against age-related cognitive decline. Neurology 1999, 52, 1563–1569. [Google Scholar] [CrossRef]
- Pottala, J.V.; Talley, J.A.; Churchill, S.W.; Lynch, D.A.; von Schacky, C.; Harris, W.S. Red blood cell fatty acids are associated with depression in a case-control study of adolescents. Prostaglandins Leukot. Essent. Fatty Acids 2012, 86, 161–165. [Google Scholar] [CrossRef]
- Pastor, S.; Creus, A.; Parrón, T.; Cebulska-Wasilewska, A.; Siffel, C.; Piperakis, S.; Marcos, R. Biomonitoring of four European populations occupationally exposed to pesticides: Use of micronuclei as biomarkers. Mutagenesis 2003, 18, 249–258. [Google Scholar] [CrossRef]
- Roe, C.M.; Mintun, M.A.; D’Angelo, G.; Xiong, C.; Grant, E.A.; Morris, J.C. Alzheimer disease and cognitive reserve: Variation of education effect with carbon 11-labeled Pittsburgh Compound B uptake. Arch. Neurol. 2008, 65, 1467–1471. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.B.; Derby, C.; LeValley, A.; Katz, M.J.; Verghese, J.; Lipton, R.B. Education delays accelerated decline on a memory test in persons who develop dementia. Neurology 2007, 69, 1657–1664. [Google Scholar] [CrossRef] [PubMed]
- Cunnane, S.C.; Schneider, J.A.; Tangney, C.; Tremblay-Mercier, J.; Fortier, M.; Bennett, D.A.; Morris, M.C. Plasma and brain fatty acid profiles in mild cognitive impairment and Alzheimer’s disease. J. Alzheimers Dis. 2012, 29, 691–697. [Google Scholar] [CrossRef]
- Nana, G.; Mitra, S.; Watson, H.; Young, C.; Wood, H.M.; Perry, S.L.; Race, A.D.; Quirke, P.; Toogood, G.J.; Loadman, P.M.; et al. Luminal Bioavailability of Orally Administered ω-3 PUFAs in the Distal Small Intestine, and Associated Changes to the Ileal Microbiome, in Humans with a Temporary Ileostomy. J. Nutr. 2021, 151, 2142–2152. [Google Scholar] [CrossRef]
- Leblhuber, F.; Ehrlich, D.; Steiner, K.; Geisler, S.; Fuchs, D.; Lanser, L.; Kurz, K. The Immunopathogenesis of Alzheimer’s Disease Is Related to the Composition of Gut Microbiota. Nutrients 2021, 13, 361. [Google Scholar] [CrossRef] [PubMed]
- Crisi, P.E.; Luciani, A.; Di Tommaso, M.; Prasinou, P.; De Santis, F.; Chatgilialoglu, C.; Pietra, M.; Procoli, F.; Sansone, A.; Giordano, M.V.; et al. The Fatty Acid-Based Erythrocyte Membrane Lipidome in Dogs with Chronic Enteropathy. Animals 2021, 11, 2604. [Google Scholar] [CrossRef]
- Saji, N.; Tsuduki, T.; Murotani, K.; Hisada, T.; Sugimoto, T.; Kimura, A.; Niida, S.; Toba, K.; Sakurai, T. Relationship between the Japanese-style diet, gut microbiota, and dementia: A cross-sectional study. Nutrition 2022, 94, 111524. [Google Scholar] [CrossRef]
- Satizabal, C.L.; Himali, J.J.; Beiser, A.S.; Ramachandran, V.; Melo van Lent, D.; Himali, D.; Aparicio, H.J.; Maillard, P.; DeCarli, C.S.; Harris, W.; et al. Association of Red Blood Cell Omega-3 Fatty Acids with MRI Markers and Cognitive Function in Midlife: The Framingham Heart Study. Neurology 2022, 99, e2572–e2582. [Google Scholar] [CrossRef]
- Sullivan, B.L.; Williams, P.G.; Meyer, B.J. Biomarker validation of a long-chain omega-3 polyunsaturated fatty acid food frequency questionnaire. Lipids 2006, 41, 845–850. [Google Scholar] [CrossRef]
- McKhann, G.; Drachman, D.; Folstein, M.; Katzman, R.; Price, D.; Stadlan, E.M. Clinical diagnosis of Alzheimer’s disease: Report of the NINCDS-ADRDA Work Group under the auspices of Department of Health and Human Services Task Force on Alzheimer’s Disease. Neurology 1984, 34, 939–944. [Google Scholar] [CrossRef]
- Ridges, L.; Sunderland, R.; Moerman, K.; Meyer, B.; Astheimer, L.; Howe, P. Cholesterol lowering benefits of soy and linseed enriched foods. Asia Pac. J. Clin. Nutr. 2001, 10, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Lepage, G.; Roy, C.C. Direct transesterification of all classes of lipids in a one-step reaction. J. Lipid Res. 1986, 27, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Deo, P.; Dhillon, V.S.; Thomas, P.; Fenech, M. The Association of Nε-Carboxymethyllysine With Polyunsaturated and Saturated Fatty Acids in Healthy Individuals. J. Gerontol. A Biol. Sci. Med. Sci. 2022, 77, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, V.S.; Deo, P.; Chua, A.; Thomas, P.; Fenech, M. Telomere Length in Healthy Adults Is Positively Associated With Polyunsaturated Fatty Acids, Including Arachidonic Acid, and Negatively With Saturated Fatty Acids. J. Gerontol. A Biol. Sci. Med. Sci. 2021, 76, 3–6. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Fatty Acids | Controls | MCI | AD | p Value (ANOVA) |
|---|---|---|---|---|
| SATFAs | ||||
| C14:0 (Myristic acid) | 0.32 ± 0.15 | 0.41 ± 0.10 | 0.36 ± 1.4 | 0.061 |
| C15:0 (Pentadecanoic acid) | 0.34 ± 0.13 | 0.36 ± 0.10 | 0.31 ± 0.1 | 0.67 |
| C16:0 (Palmitic acid) | 21.07 ± 2.88 a | 25.48 ± 3.40 b | 23.36 ± 3.86 b | <0.00001 |
| C18:0 (Stearic acid) | 18.00 ± 2.17 a | 20.61 ± 2.32 b | 18.73 ± 2.11 a | 0.0001 |
| C20:0 (Arachidic acid) | 0.71 ± 0.1 a | 0.81 ± 0.11 b | 0.74 ± 0.09 a | 0.003 |
| C22:0 (Behenic acid) | 2.60 ± 0.41 a | 3.13 ± 0.51 b | 2.66 ± 0.53 a | 0.0002 |
| C23:0 (Tricosanoic acid) | 0.39 ± 0.08 a | 0.47 ± 0.07 b | 0.38 ± 0.10 a | 0.007 |
| C24:0 (Lignoceric acid) | 8.70 ± 0.99 a | 9.76 ± 1.16 b | 8.69 ± 1.04 a | 0.001 |
| Total SATFAs | 52.14 ± 6.17 a | 61.05 ± 7.14 b | 55.26 ± 7.34 a | 0.00002 |
| MUFAs | ||||
| C16:1n-9 (Palmitoleic acid) | 0.30 ± 0.10 | 0.31 ± 0.09 | 0.34 ± 0.30 | 0.64 |
| C18:1n-9 (Oleic acid) | 14.88 ± 1.10 a | 16.08 ± 1.15 b | 16.06 ± 1.63 b | 0.002 |
| C22:1n-9 (Erucic acid) | 0.15 ± 0.03 | 0.13 ± 0.02 | 0.14 ± 0.03 | 0.08 |
| C24:1n-9 (Nervonic acid) | 6.86 ± 0.72 | 7.18 ± 0.84 | 6.8 ± 0.93 | 0.29 |
| Total MUFAs | 22.20 ± 1.54 a | 23.72 ± 1.58 b | 23.39 ± 2.57 ab | 0.012 |
| n-6 PUFAs | ||||
| C18:2n-6 (Linoleic acid) | 6.11 ± 1.22 a | 4.76 ± 1.38 b | 5.40 ± 1.36 ab | 0.001 |
| C18:3n-6 (γ-Linolenic acid) | 0.09 ± 0.02 a | 0.06 ± 0.02 b | 0.08 ± 0.05 ab | 0.03 |
| C20:3n-6 (Eicosatrienoic acid) | 1.05 ± 0.34 a | 0.74 ± 0.42 b | 0.92 ± 0.34 ab | 0.009 |
| C20:4n-6 (Arachidonic acid) | 10.18 ± 3.39 a | 5.54 ± 3.34 b | 8.41 ± 4.19 a | 0.00004 |
| Total n-6 PUFA | 17.44 ± 4.40 a | 11.12 ± 5.1 b | 14.83 ± 5.40 a | 0.00003 |
| n-3 PUFAs | ||||
| C18:3n-3 (α-Linolenic acid) | 0.08 ± 0.03 a | 0.03 ± 0.03 b | 0.05 ± 0.04 b | 0.0005 |
| C20:5n-3 (Eicosapentaenoic acid) | 1.01 ± 0.69 | 0.49 ± 0.50 | 0.96 ± 1.19 | 0.06 |
| C22:6n-3 (Docosahexaenoic acid) | 5.07 ± 2.06 a | 2.47 ± 1.77 b | 3.75 ± 2.13 b | 0.00003 |
| Total n-3 PUFAs | 6.17 ± 2.60 a | 3.00 ± 2.25 b | 4.77 ± 3.17 ab | 0.0002 |
| Total PUFAs | 23.61 ± 6.55 a | 14.13 ± 7.16 b | 19.60 ± 8.01 a | <0.00002 |
| n-3/n-6 ratio | 0.35 | 0.27 | 0.32 | 0.004 |
| Total PUFA/Total SATFA ratio | 0.45 | 0.23 | 0.35 | 0.00001 |
| Fatty Acids | MCI Mean ± SEM | Con MCI Mean ± SEM | p Value | AD Mean ± SEM | Con AD Mean ± SEM | p Value |
|---|---|---|---|---|---|---|
| 14:0 (Myristic acid) | 0.42 ± 0.02 | 0.37 ± 0.04 | 0.41 | 0.36 ± 0.03 | 0.28 ± 0.01 | 0.03 |
| 15:0 (Pentadecanoic acid) | 0.36 ± 0.03 | 0.35 ± 0.03 | 0.92 | 0.32 ± 0.03 | 0.32 ± 0.03 | 0.99 |
| 16:0 (Palmitic acid) | 25.48 ± 0.76 | 22.20 ± 0.77 | 0.003 | 23.36 ± 0.87 | 19.94 ± 0.35 | 0.0004 |
| 18:0 (Stearic acid) | 20.61 ± 0.52 | 19.09 ± 0.56 | 0.06 | 18.73 ± 0.47 | 16.91 ± 0.20 | 0.001 |
| 20:0 (Arachidic acid) | 0.81 ± 0.03 | 0.75 ± 0.03 | 0.11 | 0.74 ± 0.02 | 0.68 ± 0.02 | 0.01 |
| 22:0 (Behenic acid) | 3.14 ± 0.12 | 2.78 ± 0.11 | 0.06 | 2.66 ± 0.12 | 2.43 ± 0.06 | 0.08 |
| 23:0 (Tricosanoic acid) | 0.47 ± 0.02 | 0.41 ± 0.02 | 0.11 | 0.38 ± 0.01 | 0.38 ± 0.01 | 0.69 |
| 24:0 (Lignoceric acid) | 9.77 ± 0.26 | 8.99 ± 0.27 | 0.08 | 8.70 ± 0.23 | 8.42 ± 0.14 | 0.34 |
| Total SATFA | 61.05 ± 1.60 | 54.94 ± 1.67 | 0.02 | 55.26 ± 1.64 | 49.35 ± 0.54 | 0.001 |
| 16:1n-9 (Palmitoleic acid) | 0.32 ± 0.02 | 0.31 ± 0.02 | 0.87 | 0.35 ± 0.07 | 0.29 ± 0.02 | 0.47 |
| 18:1n-9 (Oleic acid) | 16.08 ± 0.24 | 15.37 ± 0.27 | 0.07 | 16.06 ± 0.37 | 14.38 ± 0.16 | 0.001 |
| 22:1n-9 (Erucic acid) | 0.14 ± 0.01 | 0.15 ± 0.01 | 0.07 | 0.15 ± 0.01 | 0.17 ± 0.01 | 0.15 |
| 24:1n-9 (Nervonic acid) | 7.19 ± 0.20 | 7.06 ± 0.12 | 0.56 | 6.83 ± 0.21 | 6.67 ± 0.19 | 0.54 |
| Total MUFA | 23.72 ± 0.35 | 22.89 ± 0.33 | 0.11 | 23.39 ± 0.58 | 21.50 ± 0.29 | 0.01 |
| 18:2n-6 (Linoleic acid) | 4.76 ± 0.31 | 5.74 ± 0.30 | 0.06 | 5.41 ± 0.31 | 6.49 ± 0.22 | 0.007 |
| 18:3n-6 (γ-linolenic acid) | 0.07 ± 0.01 | 0.09 ± 0.01 | 0.02 | 0.08 ± 0.01 | 0.10 ± 0.00 | 0.28 |
| 18:3n-3 (α-linolenic acid) | 0.04 ± 0.01 | 0.07 ± 0.01 | 0.005 | 0.05 ± 0.01 | 0.09 ± 0.01 | 0.007 |
| 20:3n-6 (Eicosatrienoic acid) | 0.75 ± 0.09 | 0.94 ± 0.08 | 0.12 | 0.93 ± 0.08 | 1.17 ± 0.07 | 0.02 |
| 20:4n-6 (Arachidonic acid) | 5.55 ± 0.77 | 8.61 ± 0.89 | 0.01 | 8.41 ± 0.94 | 11.75 ± 0.35 | 0.001 |
| 20:5n-3 (Eicosapentaenoic acid) | 0.49 ± 0.11 | 0.84 ± 0.12 | 0.06 | 0.96 ± 0.27 | 1.19 ± 0.18 | 0.52 |
| 22:6n-3 Docosahexaenoic acid) | 2.48 ± 0.40 | 4.20 ± 0.49 | 0.02 | 3.76 ± 0.48 | 5.96 ± 0.35 | 0.001 |
| Total PUFA | 15.17 ± 1.7 | 22.05 ± 1.88 | 0.02 | 21.30 ± 2.02 | 29.02 ± 0.74 | 0.001 |
| Total n-3 PUFA | 4.05 ± 0.70 | 6.67 ± 0.74 | 0.02 | 6.47 ± 0.94 | 9.51 ± 0.58 | 0.009 |
| Total n-6 PUFA | 11.12 ± 1.15 | 15.38 ± 1.18 | 0.02 | 14.83 ± 1.22 | 19.51 ± 0.49 | 0.001 |
| n-3/n-6 ratio | 0.32 ± 0.03 | 0.41 ± 0.02 | 0.02 | 0.40 ± 0.04 | 0.49 ± 0.03 | 0.11 |
| PUFA/SATFA | 0.27 ± 0.04 | 0.43 ± 0.06 | 0.01 | 0.41 ± 0.05 | 0.59 ± 0.02 | 0.001 |
| Controls N = 39 | MCI N = 20 | AD N = 19 | |
|---|---|---|---|
| Age (years) | |||
| Mean ± S.D | 75.75 ± 9.95 | 74.60 ± 8.74 | 76.85 ± 10.96 |
| Range | 46–90 | 58–89 | 46–89 |
| Gender | |||
| Male | 12 | 7 | 4 |
| Female | 28 | 13 | 15 |
| MMSE (0–30) | |||
| Mean ± S.D | 28.56 ± 1.33 a | 26.25 ± 2.33 b | 21.0 ± 3.86 c |
| Range | 25–30 | 21–29 | 14–26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhillon, V.S.; Thomas, P.; Lee, S.L.; Deo, P.; Fenech, M. Red Blood Cell Fatty Acid Profiles Are Significantly Altered in South Australian Mild Cognitive Impairment and Alzheimer’s Disease Cases Compared to Matched Controls. Int. J. Mol. Sci. 2023, 24, 14164. https://doi.org/10.3390/ijms241814164
Dhillon VS, Thomas P, Lee SL, Deo P, Fenech M. Red Blood Cell Fatty Acid Profiles Are Significantly Altered in South Australian Mild Cognitive Impairment and Alzheimer’s Disease Cases Compared to Matched Controls. International Journal of Molecular Sciences. 2023; 24(18):14164. https://doi.org/10.3390/ijms241814164
Chicago/Turabian StyleDhillon, Varinderpal S., Philip Thomas, Sau L. Lee, Permal Deo, and Michael Fenech. 2023. "Red Blood Cell Fatty Acid Profiles Are Significantly Altered in South Australian Mild Cognitive Impairment and Alzheimer’s Disease Cases Compared to Matched Controls" International Journal of Molecular Sciences 24, no. 18: 14164. https://doi.org/10.3390/ijms241814164
APA StyleDhillon, V. S., Thomas, P., Lee, S. L., Deo, P., & Fenech, M. (2023). Red Blood Cell Fatty Acid Profiles Are Significantly Altered in South Australian Mild Cognitive Impairment and Alzheimer’s Disease Cases Compared to Matched Controls. International Journal of Molecular Sciences, 24(18), 14164. https://doi.org/10.3390/ijms241814164