
The Glycosaminoglycan Side Chains and Modular Core Proteins of Heparan Sulphate Proteoglycans and the Varied Ways They Provide Tissue Protection by Regulating Physiological Processes and Cellular Behaviour



Abstract
1. Introduction
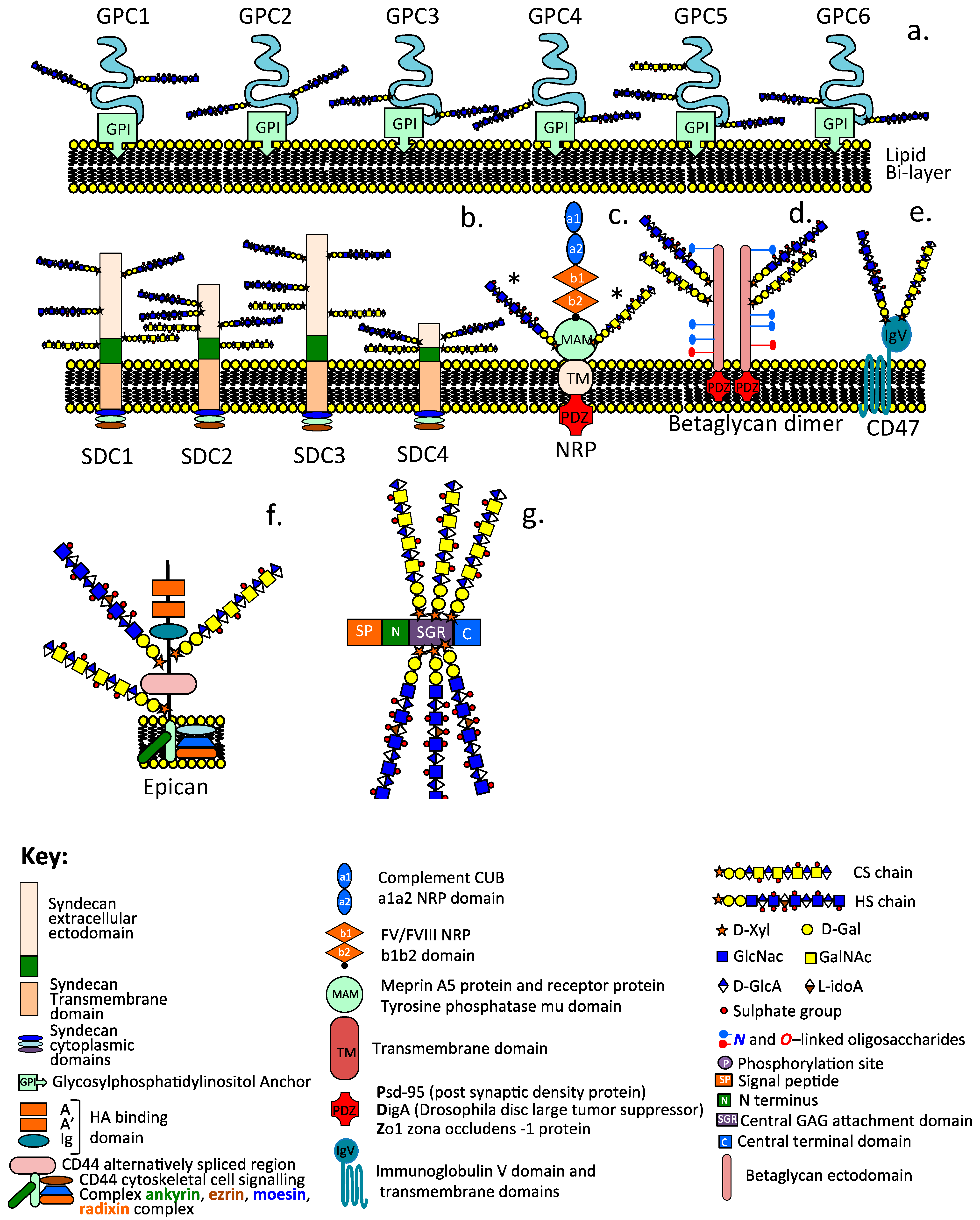
2. The Diversity of Cell Surface, Extracellular, and Cytoplasmic HS–PGs
2.1. Cytoplasmic HS–PGs
2.2. Cell-Associated HS–PGs
2.2.1. The Glypican Family
2.2.2. The Syndecan Family
2.3. Cell Surface Hybrid HS/CS Proteoglycan Co-Receptors
2.3.1. Betaglycan
2.3.2. Neuropilin
2.3.3. CD47
2.3.4. CD44
2.4. Functional HS/CS Dual Modification in PGs Controls Cellular Behavior
2.4.1. HS/CS Side Chains and Syndecan Core Proteins Promote Midkine and Pleiotrophin Binding and Tissue Growth
2.4.2. Co-Ordinated Actions of HS and CS in Neural Cell Regulation
2.4.3. HS Acts as a Molecular Switch over Cellular Behaviour in Specific Tissue Contexts
2.4.4. HS Directs Formation of Neural Networks through Axonal Guidance Proteins Slit and Robo Consistent with Its Roles in Neuritogenesis
3. Basement Membrane HS–PGs
3.1. Collagen XVIII
3.2. Agrin and Perlecan
3.3. Neurexins
3.4. Pikachurin
3.5. Eyes Shut
4. Regulatory Roles for HS–PGs in the Stem Cell Niche Determine Stem Cell Viability and Attainment of Pluripotent Migratory Stem Cell Lineages
5. HS Interactive Proteins
5.1. Perlecan

{kind=link}
{kind=link}
{kind=link}
| Domain I | Domain II | Domain III | Domain IV | Domain V |
|---|---|---|---|---|
| Laminin-1 | VLDL | FGF-7, 18 | Nidogen-1, 2 | Nidogen-1 |
| Collagen, IV, V, VI, XI | LDL | FGFBP | Fibronectin | Fibulin-2 |
| Fibronectin | Fibrillin-1 | WARP | Collagen IV | β1-integrin |
| PRELP, WARP | Wnt | Collagen VI | PDGF | α-DG |
| Fibrillin-1 | Tropoelastin | Fibulin-2 | FGF-7 | |
| Thrombospondin | Collagen VI | Endostatin | ||
| FGF1, 2, 7, 9, 10, 18 | Tropoelastin | ECM-1 | ||
| BMP-2, 4 | NG2/CSPG4 | Collagen VI | ||
| PDGF, VEGF, IL2 | Progranulin | |||
| Hh, Ang-3 | Acetyl Ch | |||
| Heparanase | α2β1 integrin | |||
| Activin A, HistoneH1 | Tropoelastin | |||
| G6b-B-R | NG2/CSPG4 |
5.2. Syndecan
| ECM Proteins | Proteases | Integrins Receptors | Morphogens Growth Factors | Cytokines Angiogenic Peptides | |
|---|---|---|---|---|---|
| Laminins | MMP-2, 7, 9 | αVβ3 | EphB4 | Activin Amphiregulin | GM–CSF |
| Fibronectin | ADAMTS-4 | αVβ5 | IGF1R | BMP-2, 4 HB-EGF | IL-2, 3, 4, 7, 12 |
| TSPs | MT1-MMP | α6β4 | FGFR | Chordin Neuroregulin | IFN |
| Collagens | Leucocyte elastase | α2β1 | ErbB2 | SHH FGF 1–23 | TNFα |
| Fibrin | Cathepsin G | α3β1 | CD148 | Frizzled proteins PDGF | C–C Chemokine |
| HB–GAM | Carboxypeptidase | α6β4 | L-Selectin | Wnts 1–3 GDNF | CXC |
| Tenascin | Thrombin | α4β1 | P-Selectin | VEGF | Angiostatin |
| Fibrillin | Plasmin | αMβ2 | E-Selectin | HGF | Endostatin |
| Tropoelastin | NCAM | TGFβ-1, TGFβ-2 | Endorepellin | ||
| PECAM | |||||
| EGFR | |||||
| VEGFR2 | |||||
| Molecule | Function, Ligands | Ref. |
|---|---|---|
| Anti-angiogenic agents | ||
| Angiostatin | 38 kDa plasmin fragment derived from plasminogen cleavage by urokinase/tPA, inhibits endothelial cell proliferation, angiogenesis | [337,338,339,340] |
| Endostatin | 20 kDa C-terminal fragment collagen XVIII, anti-angiogenic peptide | [340] |
| Restin | C-terminal fragment of XV collagen XV, anti-angiogenic peptide | |
| Cell adhesion molecules | ||
| L,E,P-Selectin | Cell adhesion leucocyte homing receptor (CD62) lectin-like sugar binding activity. Expressed by granulocytes, monocytes, lymphocytes, neutrophils | [341,342] |
| MAC-1 | Macrophage-1 antigen complement receptor (CR3) or CD11b | [343] |
| NCAM | Neurons, glia, skeletal muscle cell adhesion molecule (CD56) | [344,345,346] |
| PECAM-1 | Platelet endothelial cell adhesion molecule (CD31) on platelets, monocytes, neutrophils, T cells, endothelial cells promotes leukocyte transmigration during inflammation, angiogenesis, and integrin activation. | [347,348,349,350] |
| Chemokines | ||
| C-C | Induction of chemotaxis | [351] |
| CXC | Subfamily of the chemokine superfamily involved in leukocyte trafficking, recruitment, and activation | [352,353,354,355] |
| RANTES | RANTES is a prototypical T-cell-derived chemokine and potent inflammatory mediator that activates basophils and mast cells and attracts T cells and regulates CD8 T cell responses during chronic viral infection. | [356,357,358] |
| Cytokines | ||
| IL-2, 3, 4, 5, 7, 12 | Cytokines associated with innate immunity, trigger inflammation | [359,360,361,362] |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor, colony-stimulating factor 2 (CSF2), secreted by macrophages, T cells, mast cells, NK, endothelial cells, fibroblasts. | [363,364] |
| Interferon-γ | Signaling protein released by host cells due to viral infection | [365] |
| TNF-α | Cell-signaling protein in systemic inflammation and the acute phase response by activated macrophages; regulates immune cells. | [366,367] |
| G6b-B R | Inhibitory Megakaryocyte-Platelet Receptor G6b-B, regulated by HS, modulates tissue fibrosis modified by platelet function | [287] |
| PF4 | Interacts with HS inhibits AT-dependent thrombin and factor Xa. | [368] |
| Redox molecules | ||
| SOD | Enzyme converting superoxide free radical into O2 or H2O2 | [369] |
| ECM structural glycoproteins | ||
| Fibrin | Initial component of clot formation in wound repair | [370] |
| Fibronectin | High Mw glycoprotein, integrin binding, Cell attachment, ECM organization | [371,372] |
| Interstitial Collagens | ECM organization/stabilization provided by Type IV collagen stabilizes BM’s, HS binding in Collagen V, XI regulates collagen heterofibril formation, Collagen IV, V, VI, XI interact with perlecan. | [373,374,375,376,377,378] |
| Laminins | High Mw heterotrimeric ECM and basement membrane component | [379,380,381,382,383,384] |
| Tenascin | ECM glycoprotein, stabilizes perineuronal net formation in the CNS | [385,386] |
| TSP-1, 2 | Secreted glycoprotein family, anti-angiogenic, matricellular, multifunctional proteins in angiogenesis, apoptosis, TGF-b activation, immune regulation. | [387,388] |
| Vitronectin | Glycoprotein of hemopexin family found in serum, ECM and bone, binds to αVβ3 integrin to promote cell adhesion and cell spreading. | [389] |
| collagen V | Regulates collagen heterofibril architecture along with Coll XI | [390,391,392] |
| collagen XI | Interacts with pericellular perlecan and protects PCM | [376,393] |
| Histone H1 | H1 histones bind dynamically to chromatin in living cells and exchange rapidly between nucleosomes and may regulate transcription. | [394,395] |
| Growth factors | ||
| Amphiregulin | Amphiregullin is an EGF-like ligand that binds to EGFR and is an autocrine growth factor and mitogen for astrocytes, Schwann cells, and fibroblasts. | [396] |
| Betacellulin | Betacellulin is a member of the EGF growth factor family and an EGFR ligand. | [397,398] |
| Neuroregulin | EGF family member with diverse roles in neural development, Schwann cell and oligodendrocyte differentiation, binds to, and activates, the ErbB family of RTKs | [399] |
| Pleiotrophin | 18 kDa basic heparin-binding growth factor related to midkine, also known as neurite growth-promoting factor-1, or heparin-binding growth-associated molecule (HB–GAM) | [400,401,402] |
| Midkine | Heparin-binding growth factor, promotes cell proliferation, migration, angiogenesis, fibrinolysis. It is also known as neurite growth promoting factor-2. | [403,404,405] |
| FGF Family | 22 FGFs, FGF1–10 bind HS-inducing tyrosine kinase cell signaling | [406,407,408] |
| IGF II | Growth-promoting hormone synthesized in the liver | [409,410,411,412] |
| PDGF-AA | Potent mitogen for fibroblasts, SMCs, osteoblasts, tenocytes, and glial cells. Stored in the α-platelet granules and by SMCs, activated macrophages, and endothelial cells. Promotes angiogenesis, tissue remodelling, and PI3K-mediated cell differentiation. | [413] |
| VEGF-165, VEGF-189 | Stimulates formation of blood vessels and roles in bone formation, hematopoiesis, wound healing, and tissue development. | [240] |
| TGF-β1 TGF-β2 TGF-β3 | TGF-β is a multifunctional cytokine member of the TGF superfamily occurring as TGF-β1, β2, and β3 isoforms produced by white cell lineages. Key functions include the regulation of inflammatory processes, stem cell differentiation, T-cell regulation, and differentiation. Multifunctional homodimers interactive with Small Latent Complex (LCC), forming a Large Latent ECM Complex with LTBPs; requires proteolytic activation in situ; is anabolic and chondrogenic. HS potentiates TGF-β during wound contraction. | [414,415] |
| Activins | Interact with HS chains of cell surface and matrix HS–PGs. | [416] |
| Growth factor-binding proteins | ||
| IGFBP-3, 5 | Bind IGF-I, II, and cell surface proteins initiating outside-in cell signaling. Overexpressed in pulmonary disease, leading to excessive ECM deposition/fibrosis. | [417] |
| TGF-β BP | Latent ECM forms of TGF-β laid down in ECM as LTBP 1–4. | [418,419] |
| Follistatin | Activin-binding glycoprotein and widespread cellular distribution. Regulation/inactivation of TGF-β superfamily members, activin. | [420] |
| Proteases/protease Inhibitory proteins | ||
| AT | Heparin increases the affinity of AT for Factor IIa (Thrombin) and Factor Xa, and significantly increases AT’s inhibitory activity. HS inactivates ATs target enzymes, Thrombin, Factor Xa, and Factor IXa. | [421] |
| TFPI | TFPI, tissue factor pathway plasma Kunitz serine protease inhibitor anticoagulant, produced by endothelial cells, inhibits Factor VIIa, and Xa prevents tissue factor activation of coagulation cascade. | [422,423] |
| Factor Xa | Fondaparinux HS pentasaccharide specifically targets Factor Xa in clinical settings such as deep vein thrombosis and cardiac surgery. | [424,425,426,427] |
| Thrombin | HS anti-coagulant inhibits thrombin activity, preventing clots. | [428,429,430,431] |
| HNE | HS inhibits elastase activity through electrostatic interaction. | [432] |
| Cathepsin G | HS inhibits cathepsin-B through electrostatic interaction. | [433] |
| Chymase | Complexed by HS side chains of granular Mast cell Serglycin. | [67,434] |
| TIMP-3 | A and B β-strand N-terminal domain TIMP-3 hep-binding | [435,436,437] |
6. The Tissue-Protective Properties of HS–PGs
6.1. HS Interactions with Serpins Improves Their Inhibitory Efficiency and Tissue-Protective Properties
6.2. TIMP-3 GAG Interactions Are Tissue Protective
| Inhibitor | GAG | Functional Properties of HS-PG GAG Interactions | Ref. |
|---|---|---|---|
| α1-PI | HA | Protects HA from depolymerisation by ROS during inflammatory conditions/wound repair. | [440] |
| Bikunin ITI-L chain | HA | Transfer and covalent attachment of ITI H chains by a trans-esterification process catalyzed by TSG-6 and cross-links; stabilizes high Mw HA. | [446] |
| AT | HS (saccharide)5 GlcNAc 3-O-SO4 | Anti-coagulant inhibits thrombin, Factor Xa; IXa inhibits inflammation; angiogenesis aids in TBI and BBB repair, as well as neurocognitive recovery from TBI. | [462] |
| HCII | HS, DS 2-O-SO4, GlcA | Contributes to AT activity, inhibits Factor Xa, VIIIa, thrombin. | [455,463,464] |
| TFPI | HS, DS | Mediates TFPI-2/MSPI binding to Hep/DS to inhibit plasmin, trypsin, chymotrypsin, plasma kallikrein, Cathepsin G, and F VIIa. TFPI binds to cell surface HS–PGs. | [227,423,465,466,467,468] |
| PCI | HS/Hep | HS/Hep enhance the activity of PCI, accelerating the inhibition of α, γ-thrombin, APC, factor Xa, urokinase, and chymotrypsin. | [450,469] |
| Kallistatin | HS | Blocks VEGF, TNFα, HMBG1, Wnt, TGF-β, and EGF cell signaling. Inhibits OA, hypertension, heart and kidney disease, sepsis, influenza virus infection, tumor growth, and metastasis. | [451] |
| PAI-1 | HS/Hep | Hep/HS improves PAI-1 activity. Hep inhibits tPA synthesis matrix deposition of PAI-1 by human mesangial cells. Hep/HS enhances synthesis of two chain urokinase inhibitor PAI-1 form. | [77,470,471] |
| PN-1 | HS | Rapidly inhibits thrombin. Its physiological substrate also inhibits urokinase and plasmin, binding to ECM. HS–PGs accelerates inhibition of thrombin. It is anti-coagulant and anti-angiogenic. | [453,454,457,465,472] |
| SLPI | 12–14 unit Heparin oligos | Stoichiometric 1:1 binding of 12–14 Heparin oligosaccharide to SLPI accelerates SLPI inhibitory activity against proteinase-3, neutrophil elastase, cathepsin G, and mast cell tryptase/chymase. | [447] |
| TIMP-3 | Heparin, HS | TIMP-3 regulates the activity of MMPs, ADAMTS4, and ADAMTS5. | [459,473] |
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bermejo-Jambrina, M.; Eder, J.; Kaptein, T.M.; van Hamme, J.L.; Helgers, L.C.; Vlaming, K.E.; Brouwer, P.J.M.; van Nuenen, A.C.; Spaargaren, M.; de Bree, G.J.; et al. Infection and transmission of SARS-CoV-2 depend on heparan sulfate proteoglycans. EMBO J. 2021, 40, e106765. [Google Scholar] [CrossRef]
- Bishop, J.R.; Schuksz, M.; Esko, J.D. Heparan sulphate proteoglycans fine-tune mammalian physiology. Nature 2007, 446, 1030–1037. [Google Scholar] [CrossRef] [PubMed]
- Condomitti, G.; de Wit, J. Heparan Sulfate Proteoglycans as Emerging Players in Synaptic Specificity. Front. Mol. Neurosci. 2018, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- De Cat, B.; David, G. Developmental roles of the glypicans. Semin. Cell Dev. Biol. 2001, 12, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Dreyfuss, J.L.; Regatieri, C.V.; Jarrouge, T.R.; Cavalheiro, R.P.; Sampaio, L.O.; Nader, H.B. Heparan sulfate proteoglycans: Structure, protein interactions and cell signaling. An. Acad. Bras. Cienc. 2009, 81, 409–429. [Google Scholar] [CrossRef]
- Filmus, J. Glypicans in growth control and cancer. Glycobiology 2001, 11, 19R–23R. [Google Scholar] [CrossRef] [PubMed]
- Ghatak, S.; Maytin, E.V.; Mack, J.A.; Hascall, V.C.; Atanelishvili, I.; Moreno Rodriguez, R.; Markwald, R.R.; Misra, S. Roles of Proteoglycans and Glycosaminoglycans in Wound Healing and Fibrosis. Int. J. Cell Biol. 2015, 2015, 834893. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, C.; Selleck, S.B. Heparan sulfate proteoglycans at a glance. J. Cell Sci. 2007, 120, 1829–1832. [Google Scholar] [CrossRef] [PubMed]
- Kirn-Safran, C.; Farach-Carson, M.C.; Carson, D.D. Multifunctionality of extracellular and cell surface heparan sulfate proteoglycans. Cell. Mol. Life Sci. 2009, 66, 3421–3434. [Google Scholar] [CrossRef]
- Papy-Garcia, D.; Albanese, P. Heparan sulfate proteoglycans as key regulators of the mesenchymal niche of hematopoietic stem cells. Glycoconj. J. 2017, 34, 377–391. [Google Scholar] [CrossRef]
- Sarrazin, S.; Lamanna, W.C.; Esko, J.D. Heparan sulfate proteoglycans. Cold Spring Harb. Perspect. Biol. 2011, 3, a004952. [Google Scholar] [CrossRef] [PubMed]
- van Horssen, J.; Wesseling, P.; van den Heuvel, L.P.; de Waal, R.M.; Verbeek, M.M. Heparan sulphate proteoglycans in Alzheimer’s disease and amyloid-related disorders. Lancet Neurol. 2003, 2, 482–492. [Google Scholar] [CrossRef] [PubMed]
- Van Vactor, D.; Wall, D.P.; Johnson, K.G. Heparan sulfate proteoglycans and the emergence of neuronal connectivity. Curr. Opin. Neurobiol. 2006, 16, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Whitelock, J.; Melrose, J. Heparan sulfate proteoglycans in healthy and diseased systems. Wiley Interdiscip. Rev. Syst. Biol. Med. 2011, 3, 739–751. [Google Scholar] [CrossRef]
- Yamaguchi, Y. Heparan sulfate proteoglycans in the nervous system: Their diverse roles in neurogenesis, axon guidance, and synaptogenesis. Semin. Cell Dev. Biol. 2001, 12, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Kolset, S.O.; Tveit, H. Serglycin—Structure and biology. Cell. Mol. Life Sci. 2008, 65, 1073–1085. [Google Scholar] [CrossRef]
- Schick, B.; Gradowski, J.F.; San Antonio, J.D. Synthesis, secretion, and subcellular localization of serglycin proteoglycan in human endothelial cells. Blood 2001, 97, 449–458. [Google Scholar] [CrossRef]
- Nunes, Q.; Mournetas, V.; Lane, B.; Sutton, R.; Fernig, D.G.; Vasieva, O. The Heparin-Binding Protein Interactome in Pancreatic Diseases. Pancreatology 2013, 13, 598–604. [Google Scholar] [CrossRef]
- Ori, A.; Wilkinson, M.C.; Fernig, D.G.A. Systems Biology Approach for the Investigation of the Heparin/Heparan Sulfate Interactome. J. Biol. Chem. 2011, 286, 19892–19904. [Google Scholar] [CrossRef]
- Turnbull, J.E. Heparan sulfate glycomics: Towards systems biology strategies. Biochem. Soc. Trans. 2010, 38, 1356–1360. [Google Scholar] [CrossRef]
- Vallet, S.; Berthollier, C.; Ricard-Blum, S. The glycosaminoglycan interactome 2.0. Am. J. Physiol. Cell Physiol. 2022, 322, C1271–C1278. [Google Scholar] [CrossRef]
- Monneau, Y.; Arenzana-Seisdedos, F.; Lortat-Jacob, H. The sweet spot: How GAGs help chemokines guide migrating cells. J. Leukoc. Biol. 2016, 99, 935–953. [Google Scholar] [CrossRef]
- Nunes, Q.; Su, D.; Brownridge, P.J.; Simpson, D.M.; Sun, C.; Li, Y.; Bui, T.P.; Zhang, X.; Huang, W.; Rigden, D.J.; et al. The Heparin-Binding Proteome in Normal Pancreas and Murine Experimental Acute Pancreatitis. PLoS ONE 2019, 14, e0217633. [Google Scholar] [CrossRef] [PubMed]
- Meneghetti, M.; Hughes, A.J.; Rudd, T.R.; Nader, H.B.; Powell, A.K.; Yates, E.A.; Lima, M.A. Heparan sulfate and heparin interactions with proteins. J. R. Soc. Interface 2015, 12, 0589. [Google Scholar] [CrossRef]
- Iozzo, R.V.; Schaefer, L. Proteoglycan form and function: A comprehensive nomenclature of proteoglycans. Matrix Biol. 2015, 42, 11–55. [Google Scholar] [CrossRef]
- Bang, M.; Owczarek, S. A matter of balance: Role of neurexin and neuroligin at the synapse. Neurochem. Res. 2013, 38, 1174–1189. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Tari, P.K.; She, K.; Haas, K. Neurexin-neuroligin cell adhesion complexes contribute to synaptotropic dendritogenesis via growth stabilization mechanisms in vivo. Neuron 2010, 67, 967–983. [Google Scholar] [CrossRef] [PubMed]
- Lisé, M.; El-Husseini, A. The neuroligin and neurexin families: From structure to function at the synapse. Cell. Mol. Life Sci. 2006, 63, 1833–1849. [Google Scholar] [CrossRef]
- Rudenko, G. Dynamic Control of Synaptic Adhesion and Organizing Molecules in Synaptic Plasticity. Neural Plast. 2017, 2017, 6526151. [Google Scholar] [CrossRef]
- Husain, N.; Pellikka, M.; Hong, H.; Klimentova, T.; Choe, K.M.; Clandinin, T.R.; Tepass, U. The agrin/perlecan-related protein eyes shut is essential for epithelial lumen formation in the Drosophila retina. Dev. Cell 2006, 11, 483–493. [Google Scholar] [CrossRef]
- Omori, Y.; Araki, F.; Chaya, T.; Kajimura, N.; Irie, S.; Terada, K.; Muranishi, Y.; Tsujii, T.; Ueno, S.; Koyasu, T.; et al. Presynaptic dystroglycan-pikachurin complex regulates the proper synaptic connection between retinal photoreceptor and bipolar cells. J. Neurosci. 2012, 32, 6126–6137. [Google Scholar] [CrossRef]
- Orlandi, C.; Omori, Y.; Wang, Y.; Cao, Y.; Ueno, A.; Roux, M.J.; Condomitti, G.; de Wit, J.; Kanagawa, M.; Furukawa, T.; et al. Transsynaptic Binding of Orphan Receptor GPR179 to Dystroglycan-Pikachurin Complex Is Essential for the Synaptic Organization of Photoreceptors. Cell Rep. 2018, 25, 130–145. [Google Scholar] [CrossRef] [PubMed]
- Govindraj, P.; West, L.; Koob, T.J.; Neame, P.; Doege, K.; Hassell, J.R. Isolation and identification of the major heparan sulfate proteoglycans in the developing bovine rib growth plate. J. Biol. Chem. 2002, 277, 19461–19469. [Google Scholar] [CrossRef]
- Kaur, S.; Kuznetsova, S.A.; Pendrak, M.L.; Sipes, J.M.; Romeo, M.J.; Li, Z.; Zhang, L.; Roberts, D.D. Heparan sulfate modification of the transmembrane receptor CD47 is necessary for inhibition of T cell receptor signaling by thrombospondin-1. J. Biol. Chem. 2011, 286, 14991–15002. [Google Scholar] [CrossRef] [PubMed]
- Shintani, Y.; Takashima, S.; Asano, Y.; Kato, H.; Liao, Y.; Yamazaki, S.; Tsukamoto, O.; Seguchi, O.; Yamamoto, H.; Fukushima, T.; et al. Glycosaminoglycan modification of neuropilin-1 modulates VEGFR2 signaling. EMBO J. 2006, 25, 3045–3055. [Google Scholar] [CrossRef] [PubMed]
- Andrechak, J.; Dooling, L.J.; Discher, D.E. The macrophage checkpoint CD47: SIRPα for recognition of ‘self’ cells: From clinical trials of blocking antibodies to mechanobiological fundamentals. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20180217. [Google Scholar] [CrossRef]
- Soto-Pantoja, D.; Kaur, S.; Roberts, D.D. CD47 signaling pathways controlling cellular differentiation and responses to stress. Crit. Rev. Biochem. Mol. Biol. 2015, 50, 212–230. [Google Scholar] [CrossRef]
- Mamluk, R.; Gechtman, Z.; Kutcher, M.E.; Gasiunas, N.; Gallagher, J.; Klagsbrun, M. Neuropilin-1 binds vascular endothelial growth factor 165, placenta growth factor-2, and heparin via its b1b2 domain. J. Biol. Chem. 2002, 277, 24818–24825. [Google Scholar] [CrossRef]
- Bilandzic, M.; Stenvers, K.L. Betaglycan: A multifunctional accessory. Mol. Cell Endocrinol. 2011, 339, 180–189. [Google Scholar] [CrossRef]
- Mythreye, K.; Blobe, G.C. Proteoglycan signaling co-receptors: Roles in cell adhesion, migration and invasion. Cell Signal 2009, 21, 1548–1558. [Google Scholar] [CrossRef]
- Bernard, D.; Smith, C.L.; Brûlé, E. A Tale of Two Proteins: Betaglycan, IGSF1, and the Continuing Search for the Inhibin B Receptor. Trends Endocrinol. Metab. 2020, 31, 37–45. [Google Scholar] [CrossRef]
- Boyd, F.; Cheifetz, S.; Andres, J.; Laiho, M.; Massagué, J. Transforming growth factor-beta receptors and binding proteoglycans. J. Cell Sci. 1990, 13, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Gray, P.C.; Bilezikjian, L.M.; Vale, W. Antagonism of activin by inhibin and inhibin receptors: A functional role for betaglycan-glycan. Mol. Cell Endocrinol. 2001, 180, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Henen, M.A.; Hinck, A.P. Structural biology of betaglycan and endoglin, membrane-bound co-receptors of the TGF-beta family. Exp. Biol. Med. 2019, 244, 1547–1558. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.; Gray, P.C.; Blount, A.L.; MacConell, L.A.; Wiater, E.; Bilezikjian, L.M.; Vale, W. Betaglycan binds inhibin and can mediate functional antagonism of activin signalling. Nature 2000, 404, 411–414. [Google Scholar] [CrossRef]
- Massagué, J.; Andres, J.; Attisano, L.; Cheifetz, S.; López-Casillas, F.; Ohtsuki, M.; Wrana, J.L. TGF-beta receptors. Mol. Reprod. Dev. 1992, 32, 99–104. [Google Scholar] [CrossRef]
- Miyazono, K. TGF-beta receptors and signal transduction. Int. J. Hematol. 1997, 65, 97–104. [Google Scholar] [CrossRef]
- Sandbrink, R.; Masters, C.L.; Beyreuther, K. APP gene family. Alternative splicing generates functionally related isoforms. Ann. N. Y. Acad. Sci. 1996, 777, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Segarini, P. TGF-beta receptors. Ciba Found. Symp. 1991, 157, 29–50. [Google Scholar]
- Lin, X. Functions of heparan sulfate proteoglycans in cell signaling during development. Development 2004, 131, 6009–6021. [Google Scholar] [CrossRef]
- Ohtake, Y.; Saito, A.; Li, S. Diverse functions of protein tyrosine phosphatase σ in the nervous and immune systems. Exp. Neurol. 2018, 302, 196–204. [Google Scholar] [CrossRef]
- Schwartz, N.; Domowicz, M.S. Proteoglycans in brain development and pathogenesis. FEBS Lett. 2018, 592, 3791–3805. [Google Scholar] [CrossRef]
- Liang, Y.; Annan, R.S.; Carr, S.A.; Popp, S.; Mevissen, M.; Margolis, R.K.; Margolis, R.U. Mammalian homologues of the Drosophila slit protein are ligands of the heparan sulfate proteoglycan glypican-1 in brain. J. Biol. Chem. 1999, 274, 17885–17892. [Google Scholar] [CrossRef] [PubMed]
- Ronca, F.; Andersen, J.S.; Paech, V.; Margolis, R.U. Characterization of Slit protein interactions with glypican-1. J. Biol. Chem. 2001, 276, 29141–29147. [Google Scholar] [CrossRef]
- Zhang, F.; Ronca, F.; Linhardt, R.J.; Margolis, R.U. Structural determinants of heparan sulfate interactions with Slit proteins. Biochem. Biophys. Res. Commun. 2004, 317, 352–357. [Google Scholar] [CrossRef]
- Mega, A.; Hartmark Nilsen, M.; Leiss, L.W.; Tobin, N.P.; Miletic, H.; Sleire, L.; Strell, C.; Nelander, S.; Krona, C.; Hägerstrand, D.; et al. Astrocytes enhance glioblastoma growth. Glia 2020, 68, 316–327. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Attarha, S.; Weishaupt, H.; Edqvist, P.H.; Swartling, F.J.; Bergqvist, M.; Siebzehnrubl, F.A.; Smits, A.; Pontén, F.; Tchougounova, E. Serglycin as a potential biomarker for glioma: Association of serglycin expression, extent of mast cell recruitment and glioblastoma progression. Oncotarget 2017, 8, 24815–24827. [Google Scholar] [CrossRef]
- Abrink, M.; Grujic, M.; Pejler, G. Serglycin is essential for maturation of mast cell secretory granule. J. Biol. Chem. 2004, 279, 40897–40905. [Google Scholar] [CrossRef] [PubMed]
- Elliott, J.; Miller, C.L.; Pohajdak, B.; Talbot, D.; Helgason, C.D.; Bleackley, R.C.; Paetkau, V. Induction of a proteoglycan core protein mRNA in mouse T lymphocytes. Mol. Immunol. 1993, 30, 749–754. [Google Scholar] [CrossRef] [PubMed]
- Schick, B.; Ho, H.C.; Brodbeck, K.C.; Wrigley, C.W.; Klimas, J. Serglycin proteoglycan expression and synthesis in embryonic stem cells. Biochim. Biophys. Acta 2003, 1593, 259–267. [Google Scholar] [CrossRef]
- Zernichow, L.; Abrink, M.; Hallgren, J.; Grujic, M.; Pejler, G.; Kolset, S.O. Serglycin is the major secreted proteoglycan in macrophages and has a role in the regulation of macrophage tumor necrosis factor-alpha secretion in response to lipopolysaccharide. J. Biol. Chem. 2006, 281, 26792–26801. [Google Scholar] [CrossRef] [PubMed]
- D’Ascola, A.; Scuruchi, M.; Avenoso, A.; Bruschetta, G.; Campo, S.; Mandraffino, G.; Campo, G.M. Serglycin is involved in inflammatory response in articular mouse chondrocytes. Biochem. Biophys. Res. Commun. 2018, 499, 506–512. [Google Scholar] [CrossRef] [PubMed]
- Gruber, H.; Hanley, E.N., Jr. Expression of serglycin in human disc is increased in degenerated discs and up-regulated in vitro by exposure to IL-1ß or TNF-α. Biotech. Histochem. 2018, 93, 109–117. [Google Scholar] [CrossRef]
- Scuruchi, M.; D’Ascola, A.; Avenoso, A.; Mandraffino, G.G.; Campo, S.S.; Campo, G.M. Serglycin as part of IL-1β induced inflammation in human chondrocytes. Arch. Biochem. Biophys. 2019, 669, 80–86. [Google Scholar] [CrossRef]
- Kolset, S.; Pejler, G. Serglycin: A structural and functional chameleon with wide impact on immune cells. J. Immunol. 2011, 187, 4927–4933. [Google Scholar] [CrossRef] [PubMed]
- Henningsson, F.; Hergeth, S.; Cortelius, R.; Abrink, M.; Pejler, G. A role for serglycin proteoglycan in granular retention and processing of mast cell secretory granule components. FEBS J. 2006, 273, 4901–4912. [Google Scholar] [CrossRef]
- Pejler, G.; Abrink, M.; Ringvall, M.; Wernersson, S. Mast cell proteases. Adv. Immunol. 2007, 95, 167–255. [Google Scholar] [CrossRef]
- Stevens, R.; Adachi, R. Protease-proteoglycan complexes of mouse and human mast cells and importance of their beta-tryptase-heparin complexes in inflammation and innate immunity. Immunol. Rev. 2007, 217, 155–167. [Google Scholar] [CrossRef]
- Filmus, J.; Capurro, M.; Rast, J. Glypicans. Genome Biol. 2008, 9, 224. [Google Scholar] [CrossRef]
- Filmus, J.; Selleck, S.B. Glypicans: Proteoglycans with a surprise. J. Clin. Investig. 2001, 108, 497–501. [Google Scholar] [CrossRef]
- Gao, W.; Mitchell, H. The role of glypican-3 in regulating Wnt in hepatocellular carcinomas. Cancer Rep. 2011, 1, 14–19. [Google Scholar]
- Gao, W.; Xu, Y.; Liu, J.; Ho, M. Epitope mapping by a Wnt-blocking antibody: Evidence of the Wnt binding domain in heparan sulfate. Sci. Rep. 2016, 6, 26245. [Google Scholar] [CrossRef]
- Matsuda, K.; Maruyama, H.; Guo, F.; Kleeff, J.; Itakura, J.; Matsumoto, Y.; Lander, A.D.; Korc, M. Glypican-1 is overexpressed in human breast cancer and modulates the mitogenic effects of multiple heparin-binding growth factors in breast cancer cells. Cancer Res. 2001, 61, 5562–5569. [Google Scholar] [PubMed]
- Qiao, D.; Yang, X.; Meyer, K.; Friedl, A. Glypican-1 regulates anaphase promoting complex/cyclosome substrates and cell cycle progression in endothelial cells. Mol. Biol. Cell 2008, 19, 2789–2801. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bosse, K.; Raman, P.; Zhu, Z.; Lane, M.; Martinez, D.; Heitzeneder, S.; Rathi, K.S.; Kendsersky, N.M.; Randall, M.; Donovan, L.; et al. Identification of GPC2 as an Oncoprotein and Candidate Immunotherapeutic Target in High-Risk Neuroblastoma. Cancer Cell 2017, 32, 295–309.e212. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Fu, H.; Hewitt, S.M.; Dimitrov, D.S.; Ho, M. Therapeutically targeting glypican-2 via single-domain antibody-based chimeric antigen receptors and immunotoxins in neuroblastoma. Proc. Nat. Acad. Sci. USA 2017, 114, E6623–E6631. [Google Scholar] [CrossRef] [PubMed]
- Afratis, N.; Nikitovic, D.; Multhaupt, H.A.; Theocharis, A.D.; Couchman, J.R.; Karamanos, N.K. Syndecans—Key regulators of cell signaling and biological functions. FEBS J. 2017, 284, 27–41. [Google Scholar] [CrossRef]
- Gopal, S. Syndecans in inflammation at a glance. Front. Immunol. 2020, 11, 227. [Google Scholar]
- Xian, X.; Gopal, S.; Couchman, J.R. Syndecans as receptors and organizers of the extracellular matrix. Cell Tissue Res. 2010, 339, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Couchman, J. Syndecans: Proteoglycan regulators of cell-surface microdomains? Nat. Rev. Mol. Cell Biol. 2003, 4, 926–937. [Google Scholar] [CrossRef]
- Couchman, J.R.; Gopal, S.; Lim, H.C.; Norgaard, S.; Multhaupt, H.A. Fell-Muir Lecture: Syndecans: From peripheral coreceptors to mainstream regulators of cell behaviour. Int. J. Exp. Pathol. 2015, 96, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Deepa, S.; Yamada, S.; Zako, M.; Goldberger, O.; Sugahara, K. Chondroitin sulfate chains on syndecan-1 and syndecan-4 from normal murine mammary gland epithelial cells are structurally and functionally distinct and cooperate with heparan sulfate chains to bind growth factors. A novel function to control binding of midkine, pleiotrophin, and basic fibroblast growth factor. J. Biol. Chem. 2004, 279, 37368–37376. [Google Scholar] [PubMed]
- Jenkins, L.; Horst, B.; Lancaster, C.L.; Mythreye, K. Dually modified transmembrane proteoglycans in development and disease. Cytokine Growth Factor. Rev. 2018, 39, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Andres, J.; DeFalcis, D.; Noda, M.; Massagué, J. Binding of two growth factor families to separate domains of the proteoglycan betaglycan. J. Biol. Chem. 1992, 267, 5927–5930. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, L.; Singh, P.; Varadaraj, A.; Lee, N.Y.; Shah, S.; Flores, H.V.; O’Connell, K.; Mythreye, K. Altering the Proteoglycan State of Transforming Growth Factor β Type III Receptor (TβRIII)/Betaglycan Modulates Canonical Wnt/β-Catenin Signaling. J. Biol. Chem. 2016, 291, 25716–25728. [Google Scholar] [CrossRef] [PubMed]
- Barclay, A.; Van den Berg, T.K. The interaction between signal regulatory protein alpha (SIRPα) and CD47: Structure, function, and therapeutic target. Annu. Rev. Immunol. 2014, 32, 25–50. [Google Scholar] [CrossRef]
- Murata, Y.; Kotani, T.; Ohnishi, H.; Matozaki, T. The CD47-SIRPα signalling system: Its physiological roles and therapeutic application. J. Biochem. 2014, 155, 335–344. [Google Scholar] [CrossRef]
- Bauer, E.; Qin, Y.; Miller, T.W.; Bandle, R.W.; Csanyi, G.; Pagano, P.J.; Bauer, P.M.; Schnermann, J.; Roberts, D.D.; Isenberg, J.S. Thrombospondin-1 supports blood pressure by limiting eNOS activation and endothelial-dependent vasorelaxation. Cardiovasc. Res. 2010, 88, 471–481. [Google Scholar] [CrossRef]
- Matlung, H.; Szilagyi, K.; Barclay, N.A.; van den Berg, T.K. The CD47-SIRPα signaling axis as an innate immune checkpoint in cancer. Immunol. Rev. 2017, 276, 145–164. [Google Scholar] [CrossRef]
- Weiskopf, K. Cancer immunotherapy targeting the CD47/SIRPα axis. Eur. J. Cancer 2017, 76, 100–109. [Google Scholar] [CrossRef]
- Zhang, H.; Li, F.; Yang, Y.; Chen, J.; Hu, X. SIRP/CD47 signaling in neurological disorders. Brain Res. 2015, 1623, 74–80. [Google Scholar] [CrossRef]
- Oldenborg, P. CD47: A Cell Surface Glycoprotein Which Regulates Multiple Functions of Hematopoietic Cells in Health and Disease. ISRN Haematol. 2013, 2013, 614619. [Google Scholar] [CrossRef]
- Gholamin, S.; Mitra, S.S.; Feroze, A.H.; Liu, J.; Kahn, S.A.; Zhang, M.; Esparza, R.; Richard, C.; Ramaswamy, V.; Remke, M.; et al. Disrupting the CD47-SIRPα anti-phagocytic axis by a humanized anti-CD47 antibody is an efficacious treatment for malignant pediatric brain tumors. Sci. Transl. Med. 2017, 9, eaaf2968. [Google Scholar] [CrossRef]
- Li, Z.; Li, Y.; Gao, J.; Fu, Y.; Hua, P.; Jing, Y.; Cai, M.; Wang, H.; Tong, T. The role of CD47-SIRPα immune checkpoint in tumor immune evasion and innate immunotherapy. Life Sci. 2021, 273, 119150. [Google Scholar] [CrossRef]
- Zhang, W.; Huang, Q.; Xiao, W.; Zhao, Y.; Pi, J.; Xu, H.; Zhao, H.; Xu, J.; Evans, C.E.; Jin, H. Advances in Anti-Tumor Treatments Targeting the CD47/SIRPα Axis. Front. Immunol. 2020, 11, 18. [Google Scholar] [CrossRef]
- Milstone, L.; Hough-Monroe, L.; Kugelman, L.C.; Bender, J.R.; Haggerty, J.G. Epican, a heparan/chondroitin sulfate proteoglycan form of CD44, mediates cell-cell adhesion. J. Cell Sci. 1994, 107, 3183–3190. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.; Bouchard, T.; St. John, T.; Wayner, E.; Carter, W.G. Human keratinocytes express a new CD44 core protein (CD44E) as a heparan-sulfate intrinsic membrane proteoglycan with additional exons. J. Cell Biol. 1991, 113, 207–221. [Google Scholar] [CrossRef]
- Kugelman, L.; Ganguly, S.; Haggerty, J.G.; Weissman, S.M.; Milstone, L.M. The core protein of epican, a heparan sulfate proteoglycan on keratinocytes, is an alternative form of CD44. J. Investig. Dermatol. 1992, 99, 886–891. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Misra, S.; Hascall, V.C.; Markwald, R.R.; Ghatak, S. Interactions between Hyaluronan and Its Receptors (CD44, RHAMM) Regulate the Activities of Inflammation and Cancer. Front. Immunol. 2015, 6, 201. [Google Scholar] [CrossRef] [PubMed]
- Jackson, D.; Schenker, T.; Waibel, R.; Bell, J.I.; Stahel, R.A. Expression of alternatively spliced forms of the CD44 extracellular-matrix receptor on human lung carcinomas. Int. J. Cancer Suppl. 1994, 8, 110–115. [Google Scholar] [CrossRef]
- Jackson, D.; Bell, J.I.; Dickinson, R.; Timans, J.; Shields, J.; Whittle, N. Proteoglycan forms of the lymphocyte homing receptor CD44 are alternatively spliced variants containing the v3 exon. J. Cell Biol. 1995, 128, 673–685. [Google Scholar] [CrossRef]
- Naor, D.; Sionov, R.V.; Ish-Shalom, D. CD44: Structure, function, and association with the malignant process. Adv. Cancer Res. 1997, 71, 241–319. [Google Scholar]
- Zhou, J.; Haggerty, J.G.; Milstone, L.M. Growth and differentiation regulate CD44 expression on human keratinocytes. In Vitro Cell Dev. Biol. Anim. 1999, 35, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Dzwonek, J.; Wilczynski, G.M. CD44: Molecular interactions, signaling and functions in the nervous system. Front. Cell. Neurosci. 2015, 9, 175. [Google Scholar] [CrossRef]
- Goodison, S.; Urquidi, V.; Tarin, D. CD44 cell adhesion molecules. Mol. Pathol. 1999, 52, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Mair, D.; Ames, H.M.; Li, R. Mechanisms of invasion and motility of high-grade gliomas in the brain. Mol. Biol. Cell 2018, 29, 2509–2515. [Google Scholar] [CrossRef]
- Mooney, K.; Choy, W.; Sidhu, S.; Pelargos, P.; Bui, T.T.; Voth, B.; Barnette, N.; Yang, I. The role of CD44 in glioblastoma multiforme. J. Clin. Neurosci. 2016, 34, 1–5. [Google Scholar] [CrossRef]
- Melrose, J. Hyaluronan hydrates and compartmentalises the CNS/PNS extracellular matrix and provides niche environments conducive to the optimisation of neuronal activity. J. Neurochem. 2023, 166, 637–653. [Google Scholar] [CrossRef] [PubMed]
- Peters, A.; Sherman, L.S. Diverse Roles for Hyaluronan and Hyaluronan Receptors in the Developing and Adult Nervous System. Int. J. Mol. Sci. 2020, 21, 5988. [Google Scholar] [CrossRef]
- Preston, M.; Sherman, L.S. Neural stem cell niches: Roles for the hyaluronan-based extracellular matrix. Front. Biosci. 2011, 3, 1165–1179. [Google Scholar] [CrossRef] [PubMed]
- Raber, J.; Olsen, R.H.; Su, W.; Foster, S.; Xing, R.; Acevedo, S.F.; Sherman, L.S. CD44 is required for spatial memory retention and sensorimotor functions. Behav. Brain Res. 2014, 275, 146–149. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Ikeuchi, T.; Nara, K.; Rhodes, C.S.; Zhang, P.; Chiba, Y.; Kazuno, S.; Miura, Y.; Ago, T.; Arikawa-Hirasawa, E.; et al. Perlecan regulates pericyte dynamics in the maintenance and repair of the blood-brain barrier. J. Cell Biol. 2019, 218, 3506–3525. [Google Scholar] [CrossRef] [PubMed]
- Ahrens, T.; Bang-Christensen, S.R.; Jørgensen, A.M.; Løppke, C.; Spliid, C.B.; Sand, N.T.; Clausen, T.M.; Salanti, A.; Agerbæk, M.Ø. The Role of Proteoglycans in Cancer Metastasis and Circulating Tumor Cell Analysis. Front. Cell Dev. Biol. 2020, 8, 749. [Google Scholar] [CrossRef]
- Okamoto, I.; Kawano, Y.; Tsuiki, H.; Sasaki, J.; Nakao, M.; Matsumoto, M.; Suga, M.; Ando, M.; Nakajima, M.; Saya, H. CD44 cleavage induced by a membrane-associated metalloprotease plays a critical role in tumor cell migration. Oncogene 1999, 18, 1435–1446. [Google Scholar] [CrossRef] [PubMed]
- Bourguignon, L.; Zhu, D.; Zhu, H. CD44 isoform-cytoskeleton interaction in oncogenic signaling and tumor progression. Front. Biosci. 1998, 3, 637–649. [Google Scholar] [CrossRef]
- Lander, A.; Stipp, C.S.; Ivins, J.K. The glypican family of heparan sulfate proteoglycans: Major cell-surface proteoglycans of the developing nervous system. Perspect. Dev. Neurobiol. 1996, 3, 347–358. [Google Scholar]
- Yu, P.; Pearson, C.S.; Geller, H.M. Flexible Roles for Proteoglycan Sulfation and Receptor Signaling. Trends Neurosci. 2018, 41, 47–61. [Google Scholar] [CrossRef]
- Morimoto-Tomita, M.; Uchimura, K.; Werb, Z.; Hemmerich, S.; Rosen, S.D. Cloning and characterization of two extracellular heparin-degrading endosulfatases in mice and humans. J. Biol. Chem. 2002, 277, 49175–49185. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Omori, Y.; Katoh, K.; Kondo, M.; Kanagawa, M.; Miyata, K.; Funabiki, K.; Koyasu, T.; Kajimura, N.; Miyoshi, T.; et al. Pikachurin, a dystroglycan ligand, is essential for photoreceptor ribbon synapse formation. Nat. Neurosci. 2008, 11, 923–931. [Google Scholar] [CrossRef]
- Aricescu, A.; McKinnell, I.W.; Halfter, W.; Stoker, A.W. Heparan sulfate proteoglycans are ligands for receptor protein tyrosine phosphatase sigma. Mol. Cell Biol. 2002, 22, 1881–1892. [Google Scholar] [CrossRef]
- Fisher, D.; Xing, B.; Dill, J.; Li, H.; Hoang, H.H.; Zhao, Z.; Yang, X.L.; Bachoo, R.; Cannon, S.; Longo, F.M.; et al. Leukocyte common antigen-related phosphatase is a functional receptor for chondroitin sulfate proteoglycan axon growth inhibitors. J. Neurosci. 2011, 31, 14051–14066. [Google Scholar] [CrossRef]
- Shen, Y.; Tenney, A.P.; Busch, S.A.; Horn, K.P.; Cuascut, F.X.; Liu, K.; He, Z.; Silver, J.; Flanagan, J.G. PTPσ is a receptor for chondroitin sulfate proteoglycan, an inhibitor of neural regeneration. Science 2009, 326, 592–596. [Google Scholar] [CrossRef] [PubMed]
- Dickendesher, T.L.; Baldwin, K.T.; Mironova, Y.A.; Koriyama, Y.; Raiker, S.J.; Askew, K.L.; Wood, A.; Geoffroy, C.G.; Zheng, B.; Liepmann, C.D.; et al. NgR1 and NgR3 are receptors for chondroitin sulfate proteoglycans. Nat. Neurosci. 2012, 15, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Hayes, A.; Melrose, J. Neural Tissue Homeostasis and Repair Is Regulated via CS and DS Proteoglycan Motifs. Front. Cell Dev. Biol. 2021, 9, 696640. [Google Scholar] [CrossRef]
- Melrose, J.; Hayes, A.J.; Bix, G. The CNS/PNS Extracellular Matrix Provides Instructive Guidance Cues to Neural Cells and Neuroregulatory Proteins in Neural Development and Repair. Int. J. Mol. Sci. 2021, 22, 5583. [Google Scholar] [CrossRef]
- Coles, C.H.; Shen, Y.; Tenney, A.P.; Siebold, C.; Sutton, G.C.; Lu, W.; Gallagher, J.T.; Jones, E.Y.; Flanagan, J.G.; Aricescu, A.R. Proteoglycan-specific molecular switch for RPTPsigma clustering and neuronal extension. Science 2011, 332, 484–488. [Google Scholar] [CrossRef] [PubMed]
- Alcalde, I.; Sánchez-Fernández, C.; Del Olmo-Aguado, S.; Martín, C.; Olmiere, C.; Artime, E.; Quirós, L.M.; Merayo-Lloves, J. Synthetic Heparan Sulfate Mimetic Polymer Enhances Corneal Nerve Regeneration and Wound Healing after Experimental Laser Ablation Injury in Mice. Polymers 2022, 14, 4921. [Google Scholar] [CrossRef]
- Cruz, L.; Tellman, T.V.; Farach-Carson, M.C. Flipping the Molecular Switch: Influence of Perlecan and Its Modi-fiers in the Tumor Microenvironment. Adv. Exp. Med. Biol. 2020, 1245, 133–146. [Google Scholar]
- Elgundi, Z.; Papanicolaou, M.; Major, G.; Cox, T.R.; Melrose, J.; Whitelock, J.M.; Farrugia, B.L. Cancer Metastasis: The Role of the Extracellular Matrix and the Heparan Sulfate Proteoglycan Perlecan. Front. Oncol. 2020, 17, 1482. [Google Scholar] [CrossRef]
- Steeg, P. Tumor metastasis: Mechanistic insights and clinical challenges. Nat. Med. 2006, 12, 895–904. [Google Scholar] [CrossRef]
- Gao, Q.; Chen, C.Y.; Zong, C.; Wang, S.; Ramiah, A.; Prabhakar, P.; Morris, L.C.; Boons, G.J.; Moremen, K.W.; Prestegard, J.H. Structural Aspects of Heparan Sulfate Binding to Robo1-Ig1-2. ACS Chem. Biol. 2016, 11, 3106–3113. [Google Scholar] [CrossRef]
- Liang, Y.; Haring, M.; Roughley, P.J.; Margolis, R.K.; Margolis, R.U. Glypican and biglycan in the nuclei of neurons and glioma cells: Presence of functional nuclear localization signals and dynamic changes in glypican during the cell cycle. J. Cell Biol. 1997, 139, 851–864. [Google Scholar] [CrossRef] [PubMed]
- Roper, J.; Williamson, R.C.; Bass, M.D. Syndecan and integrin interactomes: Large complexes in small spaces. Curr. Opin. Struct. Biol. 2012, 22, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Russell, S.; Bashaw, G.J. Axon guidance pathways and the control of gene expression. Dev. Dyn. 2018, 247, 571–580. [Google Scholar] [CrossRef]
- Bashaw, G.; Kidd, T.; Murray, D.; Pawson, T.; Goodman, C.S. Repulsive axon guidance: Abelson and Enabled play opposing roles downstream of the roundabout receptor. Cell 2000, 101, 703–715. [Google Scholar] [CrossRef]
- Zang, Y.; Chaudhari, K.; Bashaw, G.J. New insights into the molecular mechanisms of axon guidance receptor regulation and signaling. Curr. Top. Dev. Biol. 2021, 142, 147–196. [Google Scholar] [PubMed]
- Muragaki, Y.; Abe, N.; Ninomiya, Y.; Olsen, B.R.; Ooshima, A. The human alpha 1(XV) collagen chain contains a large amino-terminal non-triple helical domain with a tandem repeat structure and homology to alpha 1(XVIII) collagen. J. Biol. Chem. 1994, 269, 4042–4046. [Google Scholar] [CrossRef]
- Saarela, J.; Rehn, M.; Oikarinen, A.; Autio-Harmainen, H.; Pihlajaniemi, T. The short and long forms of type XVIII collagen show clear tissue specificities in their expression and location in basement membrane zones in humans. Am. J. Pathol. 1998, 153, 611–626. [Google Scholar] [CrossRef]
- van Horssen, J.; Wilhelmus, M.M.; Heljasvaara, R.; Pihlajaniemi, T.; Wesseling, P.; de Waal, R.M.; Verbeek, M.M. Collagen XVIII: A novel heparan sulfate proteoglycan associated with vascular amyloid depositions and senile plaques in Alzheimer’s disease brains. Brain Pathol. 2002, 12, 456–462. [Google Scholar] [CrossRef]
- Heljasvaara, R.; Aikio, M.; Ruotsalainen, H.; Pihlajaniemi, T. Collagen XVIII in tissue homeostasis and dysregulation—Lessons learned from model organisms and human patients. Matrix Biol. 2017, 57–58, 55–75. [Google Scholar] [CrossRef]
- Almutairi, M.; Gong, C.; Xu, Y.G.; Chang, Y.; Shi, H. Factors controlling permeability of the blood-brain barrier. Cell Mol. Life Sci. 2016, 73, 57–77. [Google Scholar] [CrossRef]
- Tidow, H.; Mattle, D.; Nissen, P. Structural and biophysical characterisation of agrin laminin G3 domain constructs. Protein Eng. Des. Sel. 2011, 24, 219–224. [Google Scholar] [CrossRef]
- Witzemann, V. Development of the neuromuscular junction. Cell Tissue Res. 2006, 326, 263–271. [Google Scholar] [CrossRef]
- Zoeller, J.; Whitelock, J.M.; Iozzo, R.V. Perlecan regulates developmental angiogenesis by modulating the VEGF-VEGFR2 axis. Matrix Biol. 2009, 28, 284–291. [Google Scholar] [CrossRef]
- Whitelock, J.M.; Melrose, J.; Iozzo, R.V. Diverse cell signaling events modulated by perlecan. Biochemistry 2008, 47, 11174–11183. [Google Scholar] [CrossRef]
- Cole, G.J.; Halfter, W. Agrin: An extracellular matrix heparan sulfate proteoglycan involved in cell interactions and synaptogenesis. Perspect. Dev. Neurobiol. 1996, 3, 359–371. [Google Scholar] [PubMed]
- Groffen, A.J.; Ruegg, M.A.; Dijkman, H.; van de Velden, T.J.; Buskens, C.A.; van den Born, J.; Assmann, K.J.; Monnens, L.A.; Veerkamp, J.H.; van den Heuvel, L.P. Agrin is a major heparan sulfate proteoglycan in the human glomerular basement membrane. J. Histochem. Cytochem. 1998, 46, 19–27. [Google Scholar] [CrossRef]
- Halfter, W.; Schurer, B.; Yip, J.; Yip, L.; Tsen, G.; Lee, J.A.; Cole, G.J. Distribution and substrate properties of agrin, a heparan sulfate proteoglycan of developing axonal pathways. J. Comp. Neurol. 1997, 383, 1–17. [Google Scholar] [CrossRef]
- Hausser, H.; Ruegg, M.A.; Brenner, R.E.; Ksiazek, I. Agrin is highly expressed by chondrocytes and is required for normal growth. Histochem. Cell Biol. 2007, 127, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Hayes, A.; Melrose, J. 3D distribution of perlecan within intervertebral disc chondrons suggests novel regulatory roles for this multifunctional modular heparan sulphate proteoglycan. Eur. Cells Mater. 2021, 41, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Hayes, A.; Whitelock, J.; Melrose, J. Regulation of FGF-2, FGF-18 and Transcription Factor Activity by Perlecan in the Maturational Development of Transitional Rudiment and Growth Plate Cartilages and in the Maintenance of Permanent Cartilage Homeostasis. Int. J. Mol. Sci. 2022, 23, 1934. [Google Scholar] [CrossRef]
- Melrose, J.; Smith, S.; Cake, M.; Read, R.; Whitelock, J. Perlecan displays variable spatial and temporal immunolocalisation patterns in the articular and growth plate cartilages of the ovine stifle joint. Histochem. Cell Biol. 2005, 123, 561–571. [Google Scholar] [CrossRef] [PubMed]
- Melrose, J.; Smith, S.; Cake, M.; Read, R.; Whitelock, J. Comparative spatial and temporal localisation of perlecan, aggrecan and type I, II and IV collagen in the ovine meniscus: An ageing study. Histochem. Cell Biol. 2005, 124, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Melrose, J.; Roughley, P.; Knox, S.; Smith, S.; Lord, M.; Whitelock, J. The structure, location, and function of perlecan, a prominent pericellular proteoglycan of fetal, postnatal, and mature hyaline cartilages. J. Biol. Chem. 2006, 281, 36905–36914. [Google Scholar] [CrossRef]
- Eldridge, S.; Nalesso, G.; Ismail, H.; Vicente-Greco, K.; Kabouridis, P.; Ramachandran, M.; Niemeier, A.; Herz, J.; Pitzalis, C.; Perretti, M.; et al. Agrin mediates chondrocyte homeostasis and requires both LRP4 and α-dystroglycan to enhance cartilage formation in vitro and in vivo. Ann. Rheum. Dis. 2016, 75, 1228–1235. [Google Scholar] [CrossRef]
- Smith, S.; Whitelock, J.M.; Iozzo, R.V.; Little, C.B.; Melrose, J. Topographical variation in the distributions of versican, aggrecan and perlecan in the foetal human spine reflects their diverse functional roles in spinal development. Histochem. Cell Biol. 2009, 132, 491–503. [Google Scholar] [CrossRef]
- Smith, S.; Shu, C.; Melrose, J. Comparative immunolocalisation of perlecan with collagen II and aggrecan in human foetal, newborn and adult ovine joint tissues demonstrates perlecan as an early developmental chondrogenic marker. Histochem. Cell Biol. 2010, 134, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Eldridge, S.; Barawi, A.; Wang, H.; Roelofs, A.J.; Kaneva, M.; Guan, Z.; Lydon, H.; Thomas, B.L.; Thorup, A.S.; Fernandez, B.F.; et al. Agrin induces long-term osteochondral regeneration by supporting repair morphogenesis. Sci. Transl. Med. 2020, 12, eaax9086. [Google Scholar] [CrossRef]
- Garcia, J.; McCarthy, H.S.; Kuiper, J.H.; Melrose, J.; Roberts, S. Perlecan in the Natural and Cell Therapy Repair of Human Adult Articular Cartilage: Can Modifications in This Proteoglycan Be a Novel Therapeutic Approach? Biomolecules 2021, 11, 92. [Google Scholar] [CrossRef]
- Hayes, A.; Farrugia, B.L.; Biose, I.J.; Bix, G.J.; Melrose, J. Perlecan, A Multi-Functional, Cell-Instructive, Matrix-Stabilizing Proteoglycan With Roles in Tissue Development Has Relevance to Connective Tissue Repair and Regeneration. Front. Cell Dev. Biol. 2022, 10, 856261. [Google Scholar] [CrossRef]
- Hayes, A.; Hughes, C.E.; Smith, S.M.; Caterson, B.; Little, C.B.; Melrose, J. The CS Sulfation Motifs 4C3, 7D4, 3B3 [−]; and Perlecan Identify Stem Cell Populations and Their Niches, Activated Progenitor Cells and Transitional Areas of Tissue Development in the Fetal Human Elbow. Stem Cells Dev. 2016, 25, 836–847. [Google Scholar] [CrossRef]
- Kerever, A.; Schnack, J.; Vellinga, D.; Ichikawa, N.; Moon, C.; Arikawa-Hirasawa, E.; Efird, J.T.; Mercier, F. Novel extracellular matrix structures in the neural stem cell niche capture the neurogenic factor fibroblast growth factor 2 from the extracellular milieu. Stem Cells 2007, 25, 2146–2157. [Google Scholar] [CrossRef] [PubMed]
- Kerever, A.; Mercier, F.; Nonaka, R.; de Vega, S.; Oda, Y.; Zalc, B.; Okada, Y.; Hattori, N.; Yamada, Y.; Arikawa-Hirasawa, E. Perlecan is required for FGF-2 signaling in the neural stem cell niche. Stem Cell Res. 2014, 12, 492–505. [Google Scholar] [CrossRef] [PubMed]
- Melrose, J. Fractone Stem Cell Niche Components Provide Intuitive Clues in the Design of New Therapeutic Procedures/Biomatrices for Neural Repair. Int. J. Mol. Sci. 2022, 23, 5148. [Google Scholar] [CrossRef]
- Smith, S.; Melrose, J. Perlecan Delineates Stem Cell Niches in Human Foetal Hip, Knee and Elbow Cartilage Rudiments and Has Potential Roles in the Regulation of Stem Cell Differentiation. J. Stem Cell. Res. Dev. Ther. 2016, 5, 118–126. [Google Scholar] [CrossRef]
- Chakraborty, S.; Njah, K.; Pobbati, A.V.; Lim, Y.B.; Raju, A.; Lakshmanan, M.; Tergaonkar, V.; Lim, C.T.; Hong, W. Agrin as a Mechanotransduction Signal Regulating YAP through the Hippo Pathway. Cell Rep. 2017, 18, 2464–2479. [Google Scholar] [CrossRef]
- Xiong, W.; Mei, L. Agrin to YAP in Cancer and Neuromuscular Junctions. Trends Cancer 2017, 3, 247–248. [Google Scholar] [CrossRef] [PubMed]
- Münch, J.; Abdelilah-Seyfried, S. Sensing and Responding of Cardiomyocytes to Changes of Tissue Stiffness in the Diseased Heart. Front. Cell Dev. Biol. 2021, 9, 642840. [Google Scholar] [CrossRef]
- Jing, X.; Liu, B.; Deng, S.; Du, J.; She, Q. Agrin Yes-associated Protein Promotes the Proliferation of Epicardial Cells. J. Cardiovasc. Pharmacol. 2021, 77, 94–99. [Google Scholar] [CrossRef]
- Pandya, M.; Gopinathan, G.; Tillberg, C.; Wang, J.; Luan, X.; Diekwisch, T.G.H. The Hippo Pathway Effectors YAP/TAZ Are Essential for Mineralized Tissue Homeostasis in the Alveolar Bone/Periodontal Complex. J. Dev. Biol. 2022, 10, 14. [Google Scholar] [CrossRef]
- Gilbert, S.; Bonnet, C.S.; Blain, E.J. Mechanical Cues: Bidirectional Reciprocity in the Extracellular Matrix Drives Mechano-Signalling in Articular Cartilage. Int. J. Mol. Sci. 2021, 22, 13595. [Google Scholar] [CrossRef] [PubMed]
- Vincent, T.; McClurg, O.; Troeberg, L. The Extracellular Matrix of Articular Cartilage Controls the Bioavailability of Pericellular Matrix-Bound Growth Factors to Drive Tissue Homeostasis and Repair. Int. J. Mol. Sci. 2022, 23, 6003. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Li, Y.; Wang, M.; Zhao, S.; Zhao, Z.; Fang, J. Mechanotransduction pathways in the regulation of cartilage chondrocyte homoeostasis. J. Cell Mol. Med. 2020, 24, 5408–5419. [Google Scholar] [CrossRef] [PubMed]
- Guilak, F.; Hayes, A.J.; Melrose, J. Perlecan In Pericellular Mechanosensory Cell-Matrix Communication, Extracellular Matrix Stabilisation and Mechanoregulation of Load-Bearing Connective Tissues. Int. J. Mol. Sci. 2021, 22, 2716. [Google Scholar] [CrossRef] [PubMed]
- Noborn, F.; Sterky, F.H. Role of neurexin heparan sulfate in the molecular assembly of synapses—Expanding the neurexin code? FEBS J. 2023, 290, 252–265. [Google Scholar] [CrossRef]
- Roppongi, R.; Dhume, S.H.; Padmanabhan, N.; Silwal, P.; Zahra, N.; Karimi, B.; Bomkamp, C.; Patil, C.S.; Champagne-Jorgensen, K.; Twilley, R.E.; et al. LRRTMs Organize Synapses through Differential Engagement of Neurexin and PTPσ. Neuron 2020, 106, 701. [Google Scholar] [CrossRef]
- Kim, J.; Wulschner, L.E.G.; Oh, W.C.; Ko, J. Trans-synaptic mechanisms orchestrated by mammalian synaptic cell adhesion molecules. Bioessays 2022, 44, e2200134. [Google Scholar] [CrossRef]
- Uchigashima, M.; Hayashi, Y.; Futai, K. Regulation of Presynaptic Release Machinery by Cell Adhesion Molecules. Adv. Neurobiol. 2023, 33, 333–356. [Google Scholar]
- Südhof, T. Synaptic Neurexin Complexes: A Molecular Code for the Logic of Neural Circuits. Cell 2017, 171, 745–769. [Google Scholar] [CrossRef]
- Zhang, P.; Lu, H.; Peixoto, R.T.; Pines, M.K.; Ge, Y.; Oku, S.; Siddiqui, T.J.; Xie, Y.; Wu, W.; Archer-Hartmann, S.; et al. Heparan Sulfate Organizes Neuronal Synapses through Neurexin Partnerships. Cell 2018, 174, 1450–1464.e23. [Google Scholar] [CrossRef]
- Cuttler, K.; Hassan, M.; Carr, J.; Cloete, R.; Bardien, S. Emerging evidence implicating a role for neurexins in neurodegenerative and neuropsychiatric disorders. Open Biol. 2021, 11, 210091. [Google Scholar] [CrossRef]
- Cao, X.; Tabuchi, K. Functions of synapse adhesion molecules neurexin/neuroligins and neurodevelopmental disorders. Neurosci. Res. 2017, 116, 3–9. [Google Scholar] [CrossRef]
- Südhof, T. Neuroligins and neurexins link synaptic function to cognitive disease. Nature 2008, 455, 903–911. [Google Scholar] [CrossRef]
- Gauthier, J.; Siddiqui, T.J.; Huashan, P.; Yokomaku, D.; Hamdan, F.F.; Champagne, N.; Lapointe, M.; Spiegelman, D.; Noreau, A.; Lafrenière, R.G.; et al. Truncating mutations in NRXN2 and NRXN1 in autism spectrum disorders and schizophrenia. Hum. Genet. 2011, 130, 563–573. [Google Scholar] [CrossRef]
- Uchigashima, M.; Cheung, A.; Futai, K. Neuroligin-3: A Circuit-Specific Synapse Organizer That Shapes Normal Function and Autism Spectrum Disorder-Associated Dysfunction. Front. Mol. Neurosci. 2021, 14, 749164. [Google Scholar] [CrossRef] [PubMed]
- Oguro, K.; Shimazaki, K.; Yokota, H.; Onuki, Y.; Murashima, Y.; Kawai, K.; Muramatsu, S.I. Global brain delivery of neuroligin 2 gene ameliorates seizures in a mouse model of epilepsy. J. Gene Med. 2022, 24, e3402. [Google Scholar] [CrossRef] [PubMed]
- Blundell, J.; Tabuchi, K.; Bolliger, M.F.; Blaiss, C.A.; Brose, N.; Liu, X.; Südhof, T.C.; Powell, C.M. Increased anxiety-like behavior in mice lacking the inhibitory synapse cell adhesion molecule neuroligin 2. Genes Brain Behav. 2009, 8, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Maćkowiak, M.; Mordalska, P.; Wędzony, K. Neuroligins, synapse balance and neuropsychiatric disorders. Pharmacol. Rep. 2014, 66, 830–835. [Google Scholar] [CrossRef]
- Trobiani, L.; Meringolo, M.; Diamanti, T.; Bourne, Y.; Marchot, P.; Martella, G.; Dini, L.; Pisani, A.; De Jaco, A.; Bonsi, P. The neuroligins and the synaptic pathway in Autism Spectrum Disorder. Neurosci. Biobehav. Rev. 2020, 119, 37–51. [Google Scholar] [CrossRef]
- Lai, E.; Nakayama, H.; Miyazaki, T.; Nakazawa, T.; Tabuchi, K.; Hashimoto, K.; Watanabe, M.; Kano, M. An Autism-Associated Neuroligin-3 Mutation Affects Developmental Synapse Elimination in the Cerebellum. Front. Neural Circuits 2021, 15, 676891. [Google Scholar] [CrossRef]
- Wang, J.; Gong, J.; Li, L.; Chen, Y.; Liu, L.; Gu, H.; Luo, X.; Hou, F.; Zhang, J.; Song, R. Neurexin gene family variants as risk factors for autism spectrum disorder. Autism Res. 2018, 11, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Kanagawa, M.; Omori, Y.; Sato, S.; Kobayashi, K.; Miyagoe-Suzuki, Y.; Takeda, S.; Endo, T.; Furukawa, T.; Toda, T. Post-translational maturation of dystroglycan is necessary for pikachurin binding and ribbon synaptic localization. J. Biol. Chem. 2010, 285, 31208–31216. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Li, J.; Zhang, Z.; Yu, M. Pikachurin interaction with dystroglycan is diminished by defective O-mannosyl glycosylation in congenital muscular dystrophy models and rescued by LARGE overexpression. Neurosci. Lett. 2011, 489, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Noborn, F.; Nilsson, J.; Larson, G. Site-specific glycosylation of proteoglycans: A revisited frontier in proteoglycan research. Matrix Biol. 2022, 12, 289–306. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Townes-Anderson, E. Cell specific post-translational processing of pikachurin, a protein involved in retinal synaptogenesis. PLoS ONE 2012, 7, e50552. [Google Scholar] [CrossRef]
- McGuigan, D.; Heon, E.; Cideciyan, A.V.; Ratnapriya, R.; Lu, M.; Sumaroka, A.; Roman, A.J.; Batmanabane, V.; Garafalo, A.V.; Stone, E.M.; et al. EYS Mutations Causing Autosomal Recessive Retinitis Pigmentosa: Changes of Retinal Structure and Function with Disease Progression. Genes 2017, 8, 178. [Google Scholar] [CrossRef]
- Alfano, G.; Kruczek, P.M.; Shah, A.Z.; Kramarz, B.; Jeffery, G.; Zelhof, A.C.; Bhattacharya, S.S. EYS Is a Protein Associated with the Ciliary Axoneme in Rods and Cones. PLoS ONE 2016, 11, e0166397. [Google Scholar] [CrossRef]
- Garcia-Delgado, A.; Valdes-Sanchez, L.; Morillo-Sanchez, M.J.; Ponte-Zuñiga, B.; Diaz-Corrales, F.J.; de la Cerda, B. Dissecting the role of EYS in retinal degeneration: Clinical and molecular aspects and its implications for future therapy. Orphanet J. Rare Dis. 2021, 16, 222. [Google Scholar] [CrossRef]
- Korpetinou, A.; Skandalis, S.S.; Labropoulou, V.T.; Smirlaki, G.; Noulas, A.; Karamanos, N.K.; Theocharis, A.D. Serglycin: At the crossroad of inflammation and malignancy. Front. Oncol. 2014, 3, 327. [Google Scholar] [CrossRef]
- Fransson, L. Glypicans. Int. J. Biochem. Cell Biol. 2003, 35, 125–129. [Google Scholar] [CrossRef]
- Aikawa, T.; Whipple, C.A.; Lopez, M.E.; Gunn, J.; Young, A.; Lander, A.D.; Korc, M. Glypican-1 modulates the angiogenic and metastatic potential of human and mouse cancer cells. J. Clin. Investig. 2008, 118, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Bloechlinger, S.; Karchewski, L.A.; Woolf, C.J. Dynamic changes in glypican-1 expression in dorsal root ganglion neurons after peripheral and central axonal injury. Eur. J. Neurosci. 2004, 19, 1119–1132. [Google Scholar] [CrossRef] [PubMed]
- Kleeff, J.; Ishiwata, T.; Kumbasar, A.; Friess, H.; Büchler, M.W.; Lander, A.D.; Korc, M. The cell-surface heparan sulfate proteoglycan glypican-1 regulates growth factor action in pancreatic carcinoma cells and is overexpressed in human pancreatic cancer. J. Clin. Investig. 1998, 102, 1662–1673. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Ho, M. Role of glypican-1 in regulating multiple cellular signaling pathways. Am. J. Physiol. Cell Physiol. 2021, 321, C846–C858. [Google Scholar] [CrossRef] [PubMed]
- Awad, W.; Logan, D.T.; Mani, K. GPC1 (glypican 1). Atlas Genet. Cytogenet. Oncol. Haematol. 2014, 7, 461–463. [Google Scholar] [CrossRef][Green Version]
- Li, N.; Gao, W.; Zhang, Y.F.; Ho, M. Glypicans as Cancer Therapeutic Targets. Trends Cancer 2018, 4, 741–754. [Google Scholar] [CrossRef]
- Ivins, J.; Litwack, E.D.; Kumbasar, A.; Stipp, C.S.; Lander, A.D. Cerebroglycan, a developmentally regulated cell-surface heparan sulfate proteoglycan, is expressed on developing axons and growth cones. Dev. Biol. 1997, 184, 320–332. [Google Scholar] [CrossRef]
- Stipp, C.; Litwack, E.D.; Lander, A.D. Cerebroglycan: An integral membrane heparan sulfate proteoglycan that is unique to the developing nervous system and expressed specifically during neuronal differentiation. J. Cell Biol. 1994, 124, 149–160. [Google Scholar] [CrossRef]
- Kurosawa, N.; Chen, G.Y.; Kadomatsu, K.; Ikematsu, S.; Sakuma, S.; Muramatsu, T. Glypican-2 binds to midkine: The role of glypican-2 in neuronal cell adhesion and neurite outgrowth. Glycoconj. J. 2001, 18, 499–507. [Google Scholar] [CrossRef]
- Filmus, J.; Capurro, M. The role of glypicans in Hedgehog signaling. Matrix Biol. 2014, 35, 248–252. [Google Scholar] [CrossRef]
- Filmus, J.; Song, H.; Shi, W.; Duenas Gonzalez, A.; Kaya, M.; Cano-Gauci, D. Glypican-3 is a novel inhibitor of insulin-like growth factor signaling. Medicina 1999, 59, 546. [Google Scholar]
- Grisaru, S.; Cano-Gauci, D.; Tee, J.; Filmus, J.; Rosenblum, N.D. Glypican-3 modulates BMP- and FGF-mediated effects during renal branching morphogenesis. Dev. Biol. 2001, 231, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wierbowski, B.M.; Salic, A. Hedgehog pathway modulation by glypican 3-conjugated heparan sulfate. J. Cell Sci. 2022, 135, jcs259297. [Google Scholar] [CrossRef] [PubMed]
- Tumova, S.; Woods, A.; Couchman, J.R. Heparan sulfate proteoglycans on the cell surface: Versatile coordinators of cellular functions. Int. J. Biochem. Cell Biol. 2000, 32, 269–288. [Google Scholar] [CrossRef] [PubMed]
- Galli, A.; Roure, A.; Zeller, R.; Dono, R. Glypican 4 modulates FGF signalling and regulates dorsoventral forebrain patterning in Xenopus embryos. Development 2003, 130, 4919–4929. [Google Scholar] [CrossRef]
- Saunders, S.; Paine-Saunders, S.; Lander, A.D. Expression of the cell surface proteoglycan glypican-5 is developmentally regulated in kidney, limb, and brain. Dev. Biol. 1997, 190, 78–93. [Google Scholar] [CrossRef]
- Melleby, A.; Strand, M.E.; Romaine, A.; Herum, K.M.; Skrbic, B.; Dahl, C.P.; Sjaastad, I.; Fiane, A.E.; Filmus, J.; Christensen, G.; et al. The Heparan Sulfate Proteoglycan Glypican-6 Is Upregulated in the Failing Heart, and Regulates Cardiomyocyte Growth through ERK1/2 Signaling. PLoS ONE 2016, 11, e0165079. [Google Scholar] [CrossRef]
- Agere, S.; Kim, E.Y.; Akhtar, N.; Ahmed, S. Syndecans in chronic inflammatory and autoimmune diseases: Pathological insights and therapeutic opportunities. J. Cell Physiol. 2018, 233, 6346–6358. [Google Scholar] [CrossRef]
- Baba, F.; Swartz, K.; van Buren, R.; Eickhoff, J.; Zhang, Y.; Wolberg, W.; Friedl, A. Syndecan-1 and syndecan-4 are overexpressed in an estrogen receptor-negative, highly proliferative breast carcinoma subtype. Breast Cancer Res. Treat. 2006, 98, 91–98. [Google Scholar] [CrossRef]
- Gotte, M.; Joussen, A.M.; Klein, C.; Andre, P.; Wagner, D.D.; Hinkes, M.T.; Kirchhof, B.; Adamis, A.P.; Bernfield, M. Role of syndecan-1 in leukocyteendothelial interactions in the ocular vasculature. Investig. Ophthalmol. Vis. Sci. 2002, 43, 1135–1141. [Google Scholar]
- Stepp, M.; Gibson, H.E.; Gala, P.H.; Iglesia, D.D.; Pajoohesh-Ganji, A.; Pal-Ghosh, S.; Brown, M.; Aquino, C.; Schwartz, A.M.; Goldberger, O.; et al. Defects in keratinocyte activation during wound healing in the syndecan-1-deficient mouse. J. Cell Sci. 2002, 115, 4517–4531. [Google Scholar] [CrossRef]
- Zong, F.; Fthenou, E.; Wolmer, N.; Hollosi, P.; Kovalszky, I.; Szilak, L.; Mogler, C.; Nilsonne, G.; Tzanakakis, G.; Dobra, K. Syndecan-1 and FGF-2, but not FGF receptor-1, share a common transport route and co-localize with heparanase in the nuclei of mesenchymal tumor cells. PLoS ONE 2009, 4, e7346. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.; Hermanson, S.; Ekker, S.C. Syndecan-2 is essential for angiogenic sprouting during zebrafish development. Blood 2004, 103, 1710–1719. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Klass, C.; Woods, A. Syndecan-2 regulates transforming growth factor-beta signaling. J. Biol. Chem. 2004, 279, 15715–15718. [Google Scholar] [CrossRef]
- Cornelison, D.; Wilcox-Adelman, S.A.; Goetinck, P.F.; Rauvala, H.; Rapraeger, A.C.; Olwin, B.B. Essential and separable roles for syndecan-3 and syndecan-4 in skeletal muscle development and regeneration. Genes Dev. 2004, 18, 2231–2236. [Google Scholar] [CrossRef] [PubMed]
- Kaksonen, M.; Pavlov, I.; Voikar, V.; Lauri, S.E.; Hienola, A.; Riekki, R.; Lakso, M.; Taira, T.; Rauvala, H. Syndecan-3-deficient mice exhibit enhanced LTP and impaired hippocampus-dependent memory. Mol. Cell Neurosci. 2002, 21, 158–172. [Google Scholar] [CrossRef] [PubMed]
- Tinholt, M.; Stavik, B.; Louch, W.; Carlson, C.R.; Sletten, M.; Ruf, W.; Skretting, G.; Sandset, P.M.; Iversen, N. Syndecan-3 and TFPI colocalize on the surface of endothelial-, smooth muscle-, and cancer cells. PLoS ONE 2015, 10, e0117404. [Google Scholar] [CrossRef]
- Echtermeyer, F.; Bertrand, J.; Dreier, R.; Meinecke, I.; Neugebauer, K.; Fuerst, M.; Lee, Y.J.; Song, Y.W.; Herzog, C.; Theilmeier, G.; et al. Syndecan-4 regulates ADAMTS-5 activation and cartilage breakdown in osteoarthritis. Nat. Med. 2009, 15, 1072–1076. [Google Scholar] [CrossRef]
- Echtermeyer, F.; Betrand, J.; Meinecke, I.; Neugebauer, K.; Herzog, C.; Lee, Y.J.; Song, Y.W.; Dreier, R.; Pap, T. Syndecan-4 regulates cartilage degradation in osteoarthritis. Ann. Rheum. Dis. 2010, 69 (Suppl. 2), A23–A24. [Google Scholar] [CrossRef]
- Mathiesen, S.B.; Lunde, M.; Aronsen, J.M.; Romaine, A.; Kaupang, A.; Martinsen, M.; de Souza, G.A.; Nyman, T.A.; Sjaastad, I.; Christensen, G.; et al. The cardiac syndecan-4 interactome reveals a role for syndecan-4 in nuclear translocation of muscle LIM protein (MLP). J. Biol. Chem. 2019, 294, 8717–8731. [Google Scholar] [CrossRef]
- Wilcox-Adelman, S.; Denhez, F.; Goetinck, P.F. Syndecan-4 modulates focal adhesion kinase phosphorylation. J. Biol. Chem. 2002, 277, 32970–32977. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, J.; Partovian, C.; Sellke, F.W.; Simons, M. Syndecan-4 modulates basic fibroblast growth factor 2 signaling in vivo. Am. J. Physiol. Heart Circ. Physiol. 2003, 284, H2078–H2082. [Google Scholar] [CrossRef] [PubMed]
- Andres, J.L.; Stanley, K.; Cheifetz, S.; Massagué, J. Membrane-anchored and soluble forms of betaglycan, a polymorphic proteoglycan that binds transforming growth factor-beta. J. Cell Biol. 1989, 6, 3137–3145. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, I.; da Luz, F.A.C.; Uehara, I.A.; Silva, M.J.B. Cell-adhesion molecules and their soluble forms: Promising predictors of “tumor progression” and relapse in leukemia. Tumour Biol. 2018, 40, 1010428318811525. [Google Scholar] [CrossRef]
- Henke, C.; Roongta, U.; Mickelson, D.J.; Knutson, J.R.; McCarthy, J.B. CD44-related chondroitin sulfate proteoglycan, a cell surface receptor implicated with tumor cell invasion, mediates endothelial cell migration on fibrinogen and invasion into a fibrin matrix. J. Clin. Investig. 1996, 97, 2541–2552. [Google Scholar] [CrossRef]
- McDonald, B.; Kubes, P. Interactions between CD44 and Hyaluronan in Leukocyte Trafficking. Front. Immunol. 2015, 6, 68. [Google Scholar] [CrossRef] [PubMed]
- Painter, C.; Clausen, T.M.; Park, J.; Vasquez, G.J.; Liu, J.; Gordts, P.L.; Esko, J.D. Molecular Interaction of Neuropilin-1 and 3-O-sulfated Heparan Sulfate Modulates Angiogenesis-dependent Tumor Growth. FASEB J. 2020, 34, 1. [Google Scholar] [CrossRef]
- Thacker, B.; Seamen, E.; Lawrence, R.; Parker, M.W.; Xu, Y.; Liu, J.; Vander Kooi, C.W.; Esko, J.D. Expanding the 3-O-Sulfate Proteome—Enhanced Binding of Neuropilin-1 to 3-O-Sulfated Heparan Sulfate Modulates Its Activity. ACS Chem. Biol. 2016, 11, 971–980. [Google Scholar] [CrossRef]
- Uniewicz, K.; Ori, A.; Ahmed, Y.A.; Yates, E.A.; Fernig, D.G. Characterisation of the interaction of neuropilin-1 with heparin and a heparan sulfate mimetic library of heparin-derived sugars. PeerJ 2014, 2, e461. [Google Scholar] [CrossRef]
- Vempati, P.; Popel, A.S.; Mac Gabhann, F. Extracellular regulation of VEGF: Isoforms, proteolysis, and vascular patterning. Cytokine Growth Factor. Rev. 2014, 25, 1–19. [Google Scholar] [CrossRef]
- West, D.; Rees, C.G.; Duchesne, L.; Patey, S.J.; Terry, C.J.; Turnbull, J.E.; Delehedde, M.; Heegaard, C.W.; Allain, F.; Vanpouille, C.; et al. Interactions of multiple heparin binding growth factors with neuropilin-1 and potentiation of the activity of fibroblast growth factor-2. J. Biol. Chem. 2005, 280, 13457–13464. [Google Scholar] [CrossRef]
- Halfter, W.; Dong, S.; Schurer, B.; Cole, G.J. Collagen XVIII is a basement membrane heparan sulfate proteoglycan. J. Biol. Chem. 1998, 273, 25404–25412. [Google Scholar] [CrossRef]
- Kaur, I.; Ruskamo, S.; Koivunen, J.; Heljasvaara, R.; Lackman, J.J.; Izzi, V.; Petäjä-Repo, U.E.; Kursula, P.; Pihlajaniemi, T. The N-terminal domain of unknown function (DUF959) in collagen XVIII is intrinsically disordered and highly O-glycosylated. Biochem. J. 2018, 475, 3577–3593. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, H.; Watanabe, N.; Hirose, M.; Sun, X.; Atarashi, K.; Kimura, T.; Shikata, K.; Matsuda, M.; Ogawa, D.; Heljasvaara, R.; et al. Collagen XVIII, a basement membrane heparan sulfate proteoglycan, interacts with L-selectin and monocyte chemoattractant protein-1. J. Biol. Chem. 2003, 278, 13069–13076. [Google Scholar] [CrossRef]
- Marneros, A.G.; Olsen, B.R. Physiological role of collagen XVIII and endostatin. FASEB J. 2005, 19, 716–728. [Google Scholar] [CrossRef]
- Seppinen, L.; Pihlajaniemi, T. The multiple functions of collagen XVIII in development and disease. Matrix Biol. 2011, 30, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Groffen, A.J.; Buskens, C.A.; van Kuppevelt, T.H.; Veerkamp, J.H.; Monnens, L.A.; van den Heuvel, L.P. Primary structure and high expression of human agrin in basement membranes of adult lung and kidney. Eur. J. Biochem. 1998, 254, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Hayes, A.; Melrose, J. What Are the Potential Roles of Nuclear Perlecan and Other Heparan Sulphate Proteoglycans in the Normal and Malignant Phenotype. Int. J. Mol. Sci. 2021, 22, 4415. [Google Scholar] [CrossRef] [PubMed]
- Lord, M.; Chuang, C.Y.; Melrose, J.; Davies, M.J.; Iozzo, R.V.; Whitelock, J.M. The role of vascular-derived perlecan in modulating cell adhesion, proliferation and growth factor signaling. Matrix Biol. 2014, 35, 112–122. [Google Scholar] [CrossRef]
- Melrose, J. Perlecan, a modular instructive proteoglycan with diverse functional properties. Int. J. Biochem. Cell Biol. 2020, 128, 105849. [Google Scholar] [CrossRef]
- Trout, A.; Kahle, M.P.; Roberts, J.M.; Marcelo, A.; de Hoog, L.; Boychuk, J.A.; Grupke, S.L.; Berretta, A.; Gowing, E.K.; Boychuk, C.R.; et al. Perlecan Domain-V Enhances Neurogenic Brain Repair After Stroke in Mice. Transl. Stroke Res. 2021, 12, 72–86. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yu, M.; Shang, X.; Nguyen, M.H.H.; Balakrishnan, S.; Sager, R.; Hu, H. Eyes shut homolog (EYS) interacts with matriglycan of O mannosyl glycans whose deficiency results in EYS mislocalization and degeneration of photoreceptors. Sci. Rep. 2020, 10, 7795. [Google Scholar] [CrossRef]
- Mikami, T.; Kitagawa, H. Sulfated glycosaminoglycans: Their distinct roles in stem cell biology. Glycoconj. J. 2017, 34, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Oikari, L.; Okolicsanyi, R.K.; Qin, A.; Yu, C.; Griffiths, L.R.; Haupt, L.M. Cell surface heparan sulfate proteoglycans as novel markers of human neural stem cell fate determination. Stem Cell Res. 2016, 16, 92–104. [Google Scholar] [CrossRef]
- Okolicsanyi, R.; Griffiths, L.R.; Haupt, L.M. Mesenchymal stem cells, neural lineage potential, heparan sulfate proteoglycans and the matrix. Dev. Biol. 2014, 388, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Patel, N.G.; Nicholson, E.D.; Weiss, R.J. Spatiotemporal diversity and regulation of glycosaminoglycans in cell homeostasis and human disease. Am. J. Physiol. Cell Physiol. 2022, 322, C849–C864. [Google Scholar] [CrossRef]
- Motta, J.; Hassan, H.; Ibrahim, S.A. Revisiting the Syndecans: Master Signaling Regulators with Prognostic and Targetable Therapeutic Values in Breast Carcinoma. Cancers 2023, 15, 1794. [Google Scholar] [CrossRef]
- Hassan, N.; Efing, J.; Kiesel, L.; Bendas, G.; Götte, M. The Tissue Factor Pathway in Cancer: Overview and Role of Heparan Sulfate Proteoglycans. Cancers 2023, 15, 1524. [Google Scholar] [CrossRef]
- Ozsan McMillan, I.; Li, J.P.; Wang, L. Heparan sulfate proteoglycan in Alzheimer’s disease: Aberrant expression and functions in molecular pathways related to amyloid-β metabolism. Am. J. Physiol. Cell Physiol. 2023, 324, C893–C909. [Google Scholar] [CrossRef]
- Schultheis, N.; Becker, R.; Berhanu, G.; Kapral, A.; Roseman, M.; Shah, S.; Connell, A.; Selleck, S. Regulation of autophagy, lipid metabolism, and neurodegenerative pathology by heparan sulfate proteoglycans. Front. Genet. 2023, 13, 1012706. [Google Scholar] [CrossRef]
- Hayes, A.; Melrose, J. HS, an Ancient Molecular Recognition and Information Storage Glycosaminoglycan, Equips HS-Proteoglycans with Diverse Matrix and Cell-Interactive Properties Operative in Tissue Development and Tissue Function in Health and Disease. Int. J. Mol. Sci. 2023, 24, 1148. [Google Scholar] [CrossRef]
- Colin-Pierre, C.; El Baraka, O.; Danoux, L.; Bardey, V.; André, V.; Ramont, L.; Brézillon, S. Regulation of stem cell fate by HSPGs: Implication in hair follicle cycling. NPJ Regen. Med. 2022, 7, 77. [Google Scholar] [CrossRef] [PubMed]
- Hayashida, K.; Aquino, R.S.; Park, P.W. Coreceptor functions of cell surface heparan sulfate proteoglycans. Am. J. Physiol. Cell Physiol. 2022, 322, C896–C912. [Google Scholar] [CrossRef] [PubMed]
- Mashima, R.; Okuyama, T.; Ohira, M. Physiology and Pathophysiology of Heparan Sulfate in Animal Models: Its Biosynthesis and Degradation. Int. J. Mol. Sci. 2022, 23, 1963. [Google Scholar] [CrossRef] [PubMed]
- Vlodavsky, I.; Barash, U.; Nguyen, H.M.; Yang, S.M.; Ilan, N. Biology of the Heparanase-Heparan Sulfate Axis and Its Role in Disease Pathogenesis. Semin. Thromb. Hemost. 2021, 47, 240–253. [Google Scholar] [CrossRef]
- Lamanna, W.C.; Kalus, I.; Padva, M.; Baldwin, R.J.; Merry, C.L.; Dierks, T. The heparanome—The enigma of encoding and decoding heparan sulfate sulfation. J. Biotechnol. 2007, 129, 290–307. [Google Scholar] [CrossRef]
- Ori, A.; Wilkinson, M.C.; Fernig, D.G. The heparanome and regulation of cell function: Structures, functions and challenges. Front. Biosci. 2008, 13, 4309–4338. [Google Scholar] [CrossRef]
- Garzon, J.I.; Deng, L.; Murray, D.; Shapira, S.; Petrey, D.; Honig, B. A computational interactome and functional annotation for the human proteome. Elife 2016, 5, e18715. [Google Scholar] [CrossRef]
- Perez, S.; Makshakova, O.; Angulo, J.; Bedini, E.; Bisio, A.; de Paz, J.L.; Fadda, E.; Guerrini, M.; Hricovini, M.; Hricovini, M.; et al. Glycosaminoglycans: What Remains To Be Deciphered? JACS Au 2023, 3, 628–656. [Google Scholar] [CrossRef]
- Takahashi, I.; Yamada, S.; Nata, K. Effects of heparan sulfate proteoglycan syndecan-4 on the insulin secretory response in a mouse pancreatic β-cell line, MIN6. Mol. Cell Endocrinol. 2018, 470, 142–150. [Google Scholar] [CrossRef]
- Melrose, J.; Isaacs, M.D.; Smith, S.M.; Hughes, C.E.; Little, C.B.; Caterson, B.; Hayes, A.J. Chondroitin sulphate and heparan sulphate sulphation motifs and their proteoglycans are involved in articular cartilage formation during human foetal knee joint development. Histochem. Cell Biol. 2012, 138, 461–475. [Google Scholar] [CrossRef]
- Melrose, J.; Smith, S.; Whitelock, J. Perlecan immunolocalizes to perichondrial vessels and canals in human fetal cartilaginous primordia in early vascular and matrix remodeling events associated with diarthrodial joint development. J. Histochem. Cytochem. 2004, 52, 1405–1413. [Google Scholar] [CrossRef]
- Melrose, J.; Shu, C.; Whitelock, J.M.; Lord, M.S. The cartilage extracellular matrix as a transient developmental scaffold for growth plate maturation. Matrix Biol. 2016, 52–54, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Chen, S.; Pei, Y.A.; Pei, M. Impact of perlecan, a core component of basement membrane, on regeneration of cartilaginous tissues. Acta Biomater. 2021, 135, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Xie, W.Q.; Xiao, W.F.; Li, H.Z.; Naranmandakh, S.; Bruyere, O.; Reginster, J.Y.; Li, Y.S. Perlecan: Roles in osteoarthritis and potential treating target. Life Sci. 2023, 312, 121190. [Google Scholar] [CrossRef] [PubMed]
- Thompson, W.; Modla, S.; Grindel, B.J.; Czymmek, K.J.; Kirn-Safran, C.B.; Wang, L.; Duncan, R.L.; Farach-Carson, M.C. Perlecan/Hspg2 deficiency alters the pericellular space of the lacunocanalicular system surrounding osteocytic processes in cortical bone. J. Bone Min. Miner. Res. 2011, 26, 618–629. [Google Scholar] [CrossRef]
- Chakravarti, S.; Enzo, E.; Rocha Monteiro de Barros, M.; Maffezzoni, M.B.R.; Pellegrini, G. Genetic Disorders of the Extracellular Matrix: From Cell and Gene Therapy to Future Applications in Regenerative Medicine. Annu. Rev. Genom. Hum. Genet. 2022, 23, 193–222. [Google Scholar] [CrossRef]
- Arikawa-Hirasawa, E. Impact of the heparan sulfate proteoglycan perlecan on human disease and health. Am. J. Physiol. Cell Physiol. 2022, 322, C1117–C1122. [Google Scholar] [CrossRef]
- Xie, C.; Wang, Y.; Wang, J.; Xu, Y.; Liu, H.; Guo, J.; Zhu, L. Perlecan Improves Blood Spinal Cord Barrier Repair Through the Integrin β1/ROCK/MLC Pathway After Spinal Cord Injury. Mol. Neurobiol. 2023, 60, 51–67. [Google Scholar] [CrossRef]
- Lavorgna, T.; Gressett, T.E.; Chastain, W.H.; Bix, G.J. Perlecan: A review of its role in neurologic and musculoskeletal disease. Front. Physiol. 2023, 14, 1189731. [Google Scholar] [CrossRef]
- Snow, A.; Cummings, J.A.; Lake, T. The Unifying Hypothesis of Alzheimer’s Disease: Heparan Sulfate Proteoglycans/Glycosaminoglycans Are Key as First Hypothesized Over 30 Years Ago. Front. Aging Neurosci. 2021, 13, 710683. [Google Scholar] [CrossRef]
- Farach-Carson, M.; Carson, D.D. Perlecan—A multifunctional extracellular proteoglycan scaffold. Glycobiology 2007, 17, 897–905. [Google Scholar] [CrossRef] [PubMed]
- Hayes, A.; Gibson, M.A.; Shu, C.; Melrose, J. Confocal microscopy demonstrates association of LTBP-2 in fibrillin-1 microfibrils and colocalisation with perlecan in the disc cell pericellular matrix. Tissue Cell 2014, 46, 185–197. [Google Scholar] [CrossRef]
- Hayes, A.; Shu, C.C.; Lord, M.S.; Little, C.B.; Whitelock, J.M.; Melrose, J. Pericellular colocalisation and interactive properties of type VI collagen and perlecan in the intervertebral disc. Eur. Cells Mater. 2016, 32, 40–57. [Google Scholar] [CrossRef] [PubMed]
- Mongiat, M.; Taylor, K.; Otto, J.; Aho, S.; Uitto, J.; Whitelock, J.M.; Iozzo, R.V. The protein core of the proteoglycan perlecan binds specifically to fibroblast growth factor-7. J. Biol. Chem. 2000, 275, 7095–7100. [Google Scholar] [CrossRef]
- Tang, F.; Lord, M.S.; Stallcup, W.B.; Whitelock, J.M. Cell surface chondroitin sulphate proteoglycan 4 (CSPG4) binds to the basement membrane heparan sulphate proteoglycan, perlecan, and is involved in cell adhesion. J. Biochem. 2018, 163, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Vögtle, T.; Sharma, S.; Mori, J.; Nagy, Z.; Semeniak, D.; Scandola, C.; Geer, M.J.; Smith, C.W.; Lane, J.; Pollack, S.; et al. Heparan sulfates are critical regulators of the inhibitory megakaryocyte-platelet receptor G6b-B. Elife 2019, 8, e46840. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.; Fuki, I.V. Cell-surface heparan sulfate proteoglycans: Dynamic molecules mediating ligand catabolism. Curr. Opin. Lipidol. 1997, 8, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Stepp, M.; Pal-Ghosh, S.; Tadvalkar, G.; Pajoohesh-Ganji, A. Syndecan-1 and Its Expanding List of Contacts. Adv. Wound Care 2015, 4, 235–249. [Google Scholar] [CrossRef]
- Zhang, L. Glycosaminoglycan (GAG) biosynthesis and GAG-binding proteins. Prog. Mol. Biol. Transl. Sci. 2010, 93, 1–17. [Google Scholar]
- Melrose, J. Glycosaminoglycans in wound healing. Bone Tissue Regen. Insights 2016, 7, BTRI.S38670. [Google Scholar] [CrossRef]
- Bachy, S.; Letourneur, F.; Rousselle, P. Syndecan-1 interaction with the LG4/5 domain in laminin-332 is essential for keratinocyte migration. J. Cell Physiol. 2008, 214, 238–249. [Google Scholar] [CrossRef] [PubMed]
- Bernfield, M.; Sanderson, R.D. Syndecan, a developmentally regulated cell surface proteoglycan that binds extracellular matrix and growth factors. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1990, 327, 171–186. [Google Scholar] [PubMed]
- Carulli, S.; Beck, K.; Dayan, G.; Boulesteix, S.; Lortat-Jacob, H.; Rousselle, P. Cell surface proteoglycans syndecan-1 and -4 bind overlapping but distinct sites in laminin α3 LG45 protein domain. J. Biol. Chem. 2012, 287, 12204–12216. [Google Scholar] [CrossRef] [PubMed]
- Kennelly, T.; Li, Y.; Cao, Y.; Qwarnstrom, E.E.; Geoghegan, M. Distinct Binding Interactions of α5β1-Integrin and Proteoglycans with Fibronectin. Biophys. J. 2019, 117, 688–695. [Google Scholar] [CrossRef]
- Kinnunen, T.; Raulo, E.; Nolo, R.; Maccarana, M.; Lindahl, U.; Rauvala, H. Neurite outgrowth in brain neurons induced by heparin-binding growth-associated molecule (HB-GAM) depends on the specific interaction of HB-GAM with heparan sulfate at the cell surface. J. Biol. Chem. 1996, 271, 2243–2248. [Google Scholar] [CrossRef]
- Kwon, M.; Kim, Y.; Choi, Y.; Kim, S.H.; Park, S.; Han, I.; Kang, D.H.; Oh, E.S. The extracellular domain of syndecan-2 regulates the interaction of HCT116 human colon carcinoma cells with fibronectin. Biochem. Biophys. Res. Commun. 2013, 431, 415–420. [Google Scholar] [CrossRef]
- Mahalingam, Y.; Gallagher, J.T.; Couchman, J.R. Cellular adhesion responses to the heparin-binding (HepII) domain of fibronectin require heparan sulfate with specific properties. J. Biol. Chem. 2007, 282, 3221–3230. [Google Scholar] [CrossRef]
- Matsuura, H.; Momota, Y.; Murata, K.; Matsushima, H.; Suzuki, N.; Nomizu, M.; Shinkai, H.; Utani, A. Localization of the laminin alpha4 chain in the skin and identification of a heparin-dependent cell adhesion site within the laminin alpha4 chain C-terminal LG4 module. J. Investig. Dermatol. 2004, 122, 614–620. [Google Scholar] [CrossRef]
- Naganuma, H.; Satoh, E.; Asahara, T.; Amagasaki, K.; Watanabe, A.; Satoh, H.; Kuroda, K.; Zhang, L.; Nukui, H. Quantification of thrombospondin-1 secretion and expression of alphavbeta3 and alpha3beta1 integrins and syndecan-1 as cell-surface receptors for thrombospondin-1 in malignant glioma cells. J. Neurooncol. 2004, 70, 309–317. [Google Scholar] [CrossRef]
- Nanki, N.; Fujita, J.; Yang, Y.; Hojo, S.; Bandoh, S.; Yamaji, Y.; Ishida, T. Expression of oncofetal fibronectin and syndecan-1 mRNA in 18 human lung cancer cell lines. Tumour Biol. 2001, 22, 390–396. [Google Scholar] [CrossRef] [PubMed]
- Nunes, S.; Outeiro-Bernstein, M.A.; Juliano, L.; Vardiero, F.; Nader, H.B.; Woods, A.; Legrand, C.; Morandi, V. Syndecan-4 contributes to endothelial tubulogenesis through interactions with two motifs inside the pro-angiogenic N-terminal domain of thrombospondin-1. J. Cell Physiol. 2008, 214, 828–837. [Google Scholar] [CrossRef]
- Raulo, E.; Chernousov, M.A.; Carey, D.J.; Nolo, R.; Rauvala, H. Isolation of a neuronal cell surface receptor of heparin binding growth-associated molecule (HB-GAM). Identification as N-syndecan (syndecan-3). J. Biol. Chem. 1994, 269, 12999–13004. [Google Scholar] [CrossRef] [PubMed]
- Rousselle, P.; Letourneur, F. Mysterious tasks of tyrosines in syndecan-1 cytoplasmic tail. ScientificWorldJournal 2009, 9, 629–632. [Google Scholar] [CrossRef]
- Santas, A.; Bahler, C.; Peterson, J.A.; Filla, M.S.; Kaufman, P.L.; Tamm, E.R.; Johnson, D.H.; Peters, D.M. Effect of heparin II domain of fibronectin on aqueous outflow in cultured anterior segments of human eyes. Investig. Ophthalmol. Vis. Sci. 2003, 44, 4796–4804. [Google Scholar] [CrossRef] [PubMed]
- Saoncella, S.; Echtermeyer, F.; Denhez, F.; Nowlen, J.K.; Mosher, D.F.; Robinson, S.D.; Hynes, R.O.; Goetinck, P.F. Syndecan-4 signals cooperatively with integrins in a Rho-dependent manner in the assembly of focal adhesions and actin stress fibers. Proc. Natl. Acad. Sci. USA 1999, 96, 2805–2810. [Google Scholar] [CrossRef]
- Tumova, S.; Woods, A.; Couchman, J.R. Heparan sulfate chains from glypican and syndecans bind the Hep II domain of fibronectin similarly despite minor structural differences. J. Biol. Chem. 2000, 275, 9410–9417. [Google Scholar] [CrossRef]
- Utani, A.; Nomizu, M.; Matsuura, H.; Kato, K.; Kobayashi, T.; Takeda, U.; Aota, S.; Nielsen, P.K.; Shinkai, H. A unique sequence of the laminin alpha 3 G domain binds to heparin and promotes cell adhesion through syndecan-2 and -4. J. Biol. Chem. 2001, 276, 28779–28788. [Google Scholar] [CrossRef]
- Vicente, C.; Ricci, R.; Nader, H.B.; Toma, L. Syndecan-2 is upregulated in colorectal cancer cells through interactions with extracellular matrix produced by stromal fibroblasts. BMC Cell Biol. 2013, 14, 25. [Google Scholar] [CrossRef]
- Wang, J.; Markova, D.; Anderson, D.G.; Zheng, Z.; Shapiro, I.M.; Risbud, M.V. TNF-α and IL-1β promote a disintegrin-like and metalloprotease with thrombospondin type I motif-5-mediated aggrecan degradation through syndecan-4 in intervertebral disc. J. Biol. Chem. 2011, 286, 39738–39749. [Google Scholar] [CrossRef]
- Wang, Z.; Telci, D.; Griffin, M. Importance of syndecan-4 and syndecan -2 in osteoblast cell adhesion and survival mediated by a tissue transglutaminase-fibronectin complex. Exp. Cell Res. 2011, 317, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Whiteford, J.; Behrends, V.; Kirby, H.; Kusche-Gullberg, M.; Muramatsu, T.; Couchman, J.R. Syndecans promote integrin-mediated adhesion of mesenchymal cells in two distinct pathways. Exp. Cell Res. 2007, 313, 3902–3913. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, F.; Suzuki, N.; Kadoya, Y.; Utani, A.; Nakatsuka, H.; Nishi, N.; Haruki, M.; Kleinman, H.K.; Nomizu, M. Bifunctional peptides derived from homologous loop regions in the laminin alpha chain LG4 modules interact with both alpha 2 beta 1 integrin and syndecan-2. Biochemistry 2005, 44, 9581–9589. [Google Scholar] [CrossRef]
- Koyama, E.; Leatherman, J.L.; Shimazu, A.; Nah, H.D.; Pacifici, M. Syndecan-3, tenascin-C, and the development of cartilaginous skeletal elements and joints in chick limbs. Dev. Dyn. 1995, 203, 152–162. [Google Scholar] [CrossRef]
- Ricard-Blum, S.; Couchman, J.R. Conformations, interactions and functions of intrinsically disordered syndecans. Biochem. Soc. Trans. 2023, 51, 1083–1096. [Google Scholar] [CrossRef]
- Saito, Y.; Imazeki, H.; Miura, S.; Yoshimura, T.; Okutsu, H.; Harada, Y.; Ohwaki, T.; Nagao, O.; Kamiya, S.; Hayashi, R.; et al. A peptide derived from tenascin-C induces beta1 integrin activation through syndecan-4. J. Biol. Chem. 2007, 382, 34929–34937. [Google Scholar] [CrossRef]
- Salmivirta, M.; Elenius, K.; Vainio, S.; Hofer, U.; Chiquet-Ehrismann, R.; Thesleff, I.; Jalkanen, M. Syndecan from embryonic tooth mesenchyme binds tenascin. J. Biol. Chem. 1991, 266, 7733–7739. [Google Scholar] [CrossRef] [PubMed]
- Thesleff, I.; Vainio, S.; Salmivirta, M.; Jalkanen, M. Syndecan and tenascin: Induction during early tooth morphogenesis and possible interactions. Cell Differ. Dev. 1990, 32, 383–389. [Google Scholar] [CrossRef]
- Thesleff, I.; Vaahtokari, A.; Vainio, S.; Jowett, A. Molecular mechanisms of cell and tissue interactions during early tooth development. Anat. Rec. 1996, 245, 151–161. [Google Scholar] [CrossRef]
- Conklin, M.; Gangnon, R.E.; Sprague, B.L.; Van Gemert, L.; Hampton, J.M.; Eliceiri, K.W.; Bredfeldt, J.S.; Liu, Y.; Surachaicharn, N.; Newcomb, P.A.; et al. Collagen Alignment as a Predictor of Recurrence after Ductal Carcinoma In Situ. Cancer Epidemiol. Biomark. Prev. 2018, 27, 138–145. [Google Scholar] [CrossRef]
- Corless, C.; Mendoza, A.; Collins, T.; Lawler, J. Colocalization of thrombospondin and syndecan during murine development. Dev. Dyn. 1992, 193, 346–358. [Google Scholar] [CrossRef] [PubMed]
- Delehedde, M.; Lyon, M.; Sergeant, N.; Rahmoune, H.; Fernig, D.G. Proteoglycans: Pericellular and cell surface multireceptors that integrate external stimuli in the mammary gland. J. Mammary Gland Biol. Neoplasia 2001, 6, 253–273. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Litwack, E.D.; Stanley, M.J.; Langford, J.K.; Lander, A.D.; Sanderson, R.D. Heparan sulfate proteoglycans as adhesive and anti-invasive molecules. Syndecans and glypican have distinct functions. J. Biol. Chem. 1998, 273, 22825–22832. [Google Scholar] [CrossRef]
- Sanderson, R.; Lalor, P.; Bernfield, M. B lymphocytes express and lose syndecan at specific stages of differentiation. Cell Regul. 1989, 1, 27–35. [Google Scholar] [CrossRef]
- Barbosa, G.; Biancardi, M.F.; Carvalho, H.F. Heparan sulfate fine-tunes stromal-epithelial communication in the prostate gland. Dev. Dyn. 2021, 250, 618–628. [Google Scholar] [CrossRef] [PubMed]
- Bartolini, B.; Caravà, E.; Caon, I.; Parnigoni, A.; Moretto, P.; Passi, A.; Vigetti, D.; Viola, M.; Karousou, E. Heparan Sulfate in the Tumor Microenvironment. Adv. Exp. Med. Biol. 2020, 1245, 147–161. [Google Scholar] [PubMed]
- Haeger, S.; Yang, Y.; Schmidt, E.P. Heparan Sulfate in the Developing, Healthy, and Injured Lung. Am. J. Respir. Cell Mol. Biol. 2016, 55, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Lanzi, C.; Cassinelli, G. Heparan Sulfate Mimetics in Cancer Therapy: The Challenge to Define Structural Determinants and the Relevance of Targets for Optimal Activity. Molecules 2018, 23, 2915. [Google Scholar] [CrossRef]
- Park, H.; Kim, M.; Kim, H.J.; Lee, Y.; Seo, Y.; Pham, C.D.; Lee, J.; Byun, S.J.; Kwon, M.H. Heparan sulfate proteoglycans (HSPGs) and chondroitin sulfate proteoglycans (CSPGs) function as endocytic receptors for an internalizing anti-nucleic acid antibody. Sci. Rep. 2017, 7, 14373. [Google Scholar] [CrossRef]
- Park, P.; Shukla, D. Role of heparan sulfate in ocular diseases. Exp. Eye Res. 2013, 110, 1–9. [Google Scholar] [CrossRef]
- Patel, V.; Pineda, D.L.; Hoffman, M.P. The function of heparan sulfate during branching morphogenesis. Matrix Biol. 2017, 57–58, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, J.; Powell, A.; Guimond, S. Heparan sulfate: Decoding a dynamic multifunctional cell regulator. Trends Cell Biol. 2001, 11, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, J.; Miller, R.L.; Ahmed, Y.; Puvirajesinghe, T.M.; Guimond, S.E. Glycomics profiling of heparan sulfate structure and activity. Methods Enzym. Enzymol. 2010, 480, 65–85. [Google Scholar]
- Xiong, A.; Kundu, S.; Forsberg-Nilsson, K. Heparan sulfate in the regulation of neural differentiation and glioma development. FEBS J. 2014, 281, 4993–5008. [Google Scholar] [CrossRef]
- Xu, D.; Esko, J.D. Demystifying heparan sulfate-protein interactions. Annu. Rev. Biochem. 2014, 83, 129–157. [Google Scholar] [CrossRef]
- Xu, Z.; Chen, S.; Feng, D.; Liu, Y.; Wang, Q.; Gao, T.; Liu, Z.; Zhang, Y.; Chen, J.; Qiu, L. Biological role of heparan sulfate in osteogenesis: A review. Carbohydr. Polym. 2021, 272, 118490. [Google Scholar] [CrossRef]
- Barrett, C.D.; Moore, H.B.; Banerjee, A.; Silliman, C.C.; Moore, E.E.; Yaffe, M.B. Human neutrophil elastase mediates fibrinolysis shutdown through competitive degradation of plasminogen and generation of angiostatin. J. Trauma Acute Care Surg. 2017, 83, 1053–1061. [Google Scholar] [CrossRef]
- Doll, J.A.; Soff, G.A. Angiostatin. Cancer Treat. Res. 2005, 126, 175–204. [Google Scholar]
- Li, T.; Kang, G.; Wang, T.; Huang, H. Tumor angiogenesis and anti-angiogenic gene therapy for cancer. Oncol. Lett. 2018, 16, 687–702. [Google Scholar] [CrossRef]
- Yadav, L.; Puri, N.; Rastogi, V.; Satpute, P.; Sharma, V. Tumour Angiogenesis and Angiogenic Inhibitors: A Review. J. Clin. Diagn. Res. 2015, 9, XE01–XE05. [Google Scholar] [CrossRef]
- Farrugia, B.L.; Lord, M.S.; Melrose, J.; Whitelock, J.M. The Role of Heparan Sulfate in Inflammation, and the Development of Biomimetics as Anti-Inflammatory Strategies. J. Histochem. Cytochem. 2018, 66, 321–336. [Google Scholar] [CrossRef] [PubMed]
- Lanzi, C.; Zaffaroni, N.; Cassinelli, G. Targeting Heparan Sulfate Proteoglycans and their Modifying Enzymes to Enhance Anticancer Chemotherapy Efficacy and Overcome Drug Resistance. Curr. Med. Chem. 2017, 24, 2860–2886. [Google Scholar] [CrossRef] [PubMed]
- Rosetti, F.; Mayadas, T.N. The many faces of Mac-1 in autoimmune disease. Immunol. Rev. 2016, 269, 175–193. [Google Scholar] [CrossRef] [PubMed]
- Goridis, C.; Brunet, J.F. NCAM: Structural diversity, function and regulation of expression. Semin. Cell Biol. 1992, 3, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.P. Cell adhesion molecules of the immunoglobulin supergene family and their role in malignant transformation and progression to metastatic disease. Cancer Metastasis Rev. 1991, 10, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Weledji, E.P.; Assob, J.C. The ubiquitous neural cell adhesion molecule (N-CAM). Ann. Med. Surg. 2014, 3, 77–81. [Google Scholar] [CrossRef]
- Bousarghin, L.; Hubert, P.; Franzen, E.; Jacobs, N.; Boniver, J.; Delvenne, P. Human papillomavirus 16 virus-like particles use heparan sulfates to bind dendritic cells and colocalize with langerin in Langerhans cells. J. Gen. Virol. 2005, 86, 1297–1305. [Google Scholar] [CrossRef]
- Escribano-Romero, E.; Jimenez-Clavero, M.A.; Gomes, P.; García-Ranea, J.A.; Ley, V. Heparan sulphate mediates swine vesicular disease virus attachment to the host cell. J. Gen. Virol. 2004, 85, 653–663. [Google Scholar] [CrossRef][Green Version]
- Hilgard, P.; Stockert, R. Heparan sulfate proteoglycans initiate dengue virus infection of hepatocytes. Hepatology 2000, 32, 1069–1077. [Google Scholar] [CrossRef]
- Olenina, L.; Kuzmina, T.I.; Sobolev, B.N.; Kuraeva, T.E.; Kolesanova, E.F.; Archakov, A.I. Identification of glycosaminoglycan-binding sites within hepatitis C virus envelope glycoprotein E2. J. Viral Hepat. 2005, 12, 584–593. [Google Scholar] [CrossRef]
- Qiu, J.; Handa, A.; Kirby, M.; Brown, K.E. The interaction of heparin sulfate and adeno-associated virus 2. Virology 2000, 269, 137–147. [Google Scholar] [CrossRef][Green Version]
- Cabrero-de Las Heras, S.; Martinez-Balibrea, E. CXC family of chemokines as prognostic or predictive biomarkers and possible drug targets in colorectal cancer. World J. Gastroenterol. 2018, 24, 4738–4749. [Google Scholar] [CrossRef] [PubMed]
- Proudfoot, A.E. Chemokines and Glycosaminoglycans. Front. Immunol. 2015, 6, 246. [Google Scholar] [CrossRef] [PubMed]
- Susek, K.H.; Karvouni, M.; Alici, E.; Lundqvist, A. The Role of CXC Chemokine Receptors 1-4 on Immune Cells in the Tumor Microenvironment. Front. Immunol. 2018, 9, 2159. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Cao, H.B.; Li, W.J.; Zhao, L. The CXCL12 (SDF-1)/CXCR4 chemokine axis: Oncogenic properties, molecular targeting, and synthetic and natural product CXCR4 inhibitors for cancer therapy. Chin. J. Nat. Med. 2018, 16, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Graziano, F.M.; Cook, E.B.; Stahl, J.L. Cytokines, chemokines, RANTES, and eotaxin. Allergy Asthma Proc. 1999, 20, 141–146. [Google Scholar] [CrossRef]
- Kalinkovich, A.; Weisman, Z.; Bentwich, Z. Chemokines and chemokine receptors: Role in HIV infection. Immunol. Lett. 1999, 68, 281–287. [Google Scholar] [CrossRef]
- Zhang, L.; Redington, A.E.; Holgate, S.T. RANTES: A novel mediator of allergic inflammation? Clin. Exp. Allergy 1994, 24, 899–904. [Google Scholar] [CrossRef]
- Gupta, S.C.; Kunnumakkara, A.B.; Aggarwal, S.; Aggarwal, B.B. Inflammation, a Double-Edge Sword for Cancer and Other Age-Related Diseases. Front. Immunol. 2018, 9, 2160. [Google Scholar] [CrossRef]
- Jupe, S.; Ray, K.; Roca, C.D.; Varusai, T.; Shamovsky, V.; Stein, L.; D’Eustachio, P.; Hermjakob, H. Interleukins and their signaling pathways in the Reactome biological pathway database. J. Allergy Clin. Immunol. 2018, 141, 1411–1416. [Google Scholar] [CrossRef]
- Ribatti, D. Interleukins as modulators of angiogenesis and anti-angiogenesis in tumors. Cytokine 2019, 118, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.; Bhar, S.; Devi, C.S. A review on interleukins: The key manipulators in rheumatoid arthritis. Mod. Rheumatol. 2017, 27, 723–746. [Google Scholar] [CrossRef] [PubMed]
- Becher, B.; Tugues, S.; Greter, M. GM-CSF: From Growth Factor to Central Mediator of Tissue Inflammation. Immunity 2016, 45, 963–973. [Google Scholar] [CrossRef]
- Wicks, I.P.; Roberts, A.W. Targeting GM-CSF in inflammatory diseases. Nat. Rev. Rheumatol. 2016, 12, 37–48. [Google Scholar] [CrossRef]
- Pestka, S.; Krause, C.D.; Walter, M.R. Interferons, interferon-like cytokines, and their receptors. Immunol. Rev. 2004, 202, 8–32. [Google Scholar] [CrossRef] [PubMed]
- Eskay, R.L.; Grino, M.; Chen, H.T. Interleukins, signal transduction, and the immune system-mediated stress response. Adv. Exp. Med. Biol. 1990, 274, 331–343. [Google Scholar] [PubMed]
- Scheringa, M.; Marquet, R.L. TNF: A brief review with emphasis on its antitumor activity. Biotherapy 1990, 2, 275–281. [Google Scholar] [CrossRef]
- Fiore, M.; Kakkar, V.V. Platelet factor 4 neutralizes heparan sulfate-enhanced antithrombin inactivation of factor Xa by preventing interaction(s) of enzyme with polysaccharide. Biochem. Biophys. Res. Commun. 2003, 311, 71–76. [Google Scholar] [CrossRef]
- Kaur, N.; Sharma, A.K.; Shakeel, A.; Kumar, V.; Singh, A.; Gupta, A.; Suhag, D.; Rajput, S.K.; Mukherjee, M. Therapeutic Implications of Superoxide Dismutase And Its Importance in Kinase Drug Discovery. Curr. Top. Med. Chem. 2017, 17, 2495–2508. [Google Scholar] [CrossRef]
- Weisel, J.W.; Litvinov, R.I. Fibrin Formation, Structure and Properties. Subcell. Biochem. 2017, 82, 405–456. [Google Scholar] [CrossRef]
- Maurer, L.M.; Ma, W.; Mosher, D.F. Dynamic structure of plasma fibronectin. Crit. Rev. Biochem. Mol. Biol. 2015, 51, 213–227. [Google Scholar] [CrossRef]
- Zollinger, A.J.; Smith, M.L. Fibronectin, the extracellular glue. Matrix Biol. 2017, 60–61, 27–37. [Google Scholar] [CrossRef]
- Birk, D.E.; Trelstad, R.L. Extracellular compartments in tendon morphogenesis: Collagen fibril, bundle, and macroaggregate formation. J. Cell Biol. 1986, 103, 231–240. [Google Scholar] [PubMed]
- Haramoto, T.; Makino, H.; Ikeda, S.; Wieslander, J.; Ota, Z. Ultrastructural localization of the three major basement membrane components—Type IV collagen, heparan sulfate proteoglycan and laminin—In human membranous glomerulonephritis. Am. J. Nephrol. 1994, 14, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Keene, D.R.; Oxford, J.T.; Morris, N.P. Ultrastructural localization of collagen types II, IX, and XI in the growth plate of human rib and fetal bovine epiphyseal cartilage: Type XI collagen is restricted to thin fibrils. J. Histochem. Cytochem. 1995, 43, 967–979. [Google Scholar]
- McDougal, O.; Warner, L.R.; Mallory, C.; Oxford, J.T. Predicted Structure and Binding Motifs of Collagen α1(XI). GSTF Int. J. Bioinforma Biotechnol. 2011, 1, 43–48. [Google Scholar]
- Thom, J.R.; Morris, N.P. Biosynthesis and proteolytic processing of type XI collagen in embryonic chick sterna. J. Biol. Chem. 1991, 266, 7262–7269. [Google Scholar]
- Wenstrup, R.J.; Florer, J.B.; Brunskill, E.W.; Bell, S.M.; Chervoneva, I.; Birk, D.E. Type V collagen controls the initiation of collagen fibril assembly. J. Biol. Chem. 2004, 279, 53331–53337. [Google Scholar] [CrossRef]
- Engvall, E.; Wewer, U.M. Domains of laminin. J. Cell Biochem. 1996, 61, 493–501. [Google Scholar] [PubMed]
- Hunt, G. The role of laminin in cancer invasion and metastasis. Exp. Cell Biol. 1989, 57, 165–176. [Google Scholar] [PubMed]
- Kleinman, H.K.; Weeks, B.S. Laminin: Structure, functions and receptors. Curr. Opin. Cell Biol. 1989, 1, 964–967. [Google Scholar] [CrossRef] [PubMed]
- Mercurio, A.M. Laminin: Multiple forms, multiple receptors. Curr. Opin. Cell Biol. 1990, 2, 845–849. [Google Scholar] [PubMed]
- Timpl, R.; Brown, J.C. The laminins. Matrix Biol. 1994, 14, 275–281. [Google Scholar] [PubMed]
- Yamada, M.; Sekiguchi, K. Molecular Basis of Laminin-Integrin Interactions. Curr. Top. Membr. 2015, 76, 197–229. [Google Scholar] [CrossRef]
- Faissner, A.; Roll, L.; Theocharidis, U. Tenascin-C in the matrisome of neural stem and progenitor cells. Mol. Cell Neurosci. 2017, 81, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Midwood, K.S.; Chiquet, M.; Tucker, R.P.; Orend, G. Tenascin-C at a glance. J. Cell Sci. 2016, 129, 4321–4327. [Google Scholar] [CrossRef]
- Huang, T.; Sun, L.; Yuan, X.; Qiu, H. Thrombospondin-1 is a multifaceted player in tumor progression. Oncotarget 2017, 8, 84546–84558. [Google Scholar] [CrossRef] [PubMed]
- Stenina-Adognravi, O.; Plow, E.F. Thrombospondin-4 in tissue remodeling. Matrix Biol. 2019, 75, 300–3131. [Google Scholar]
- Schvartz, I.; Seger, D.; Shaltiel, S. Vitronectin. Int. J. Biochem. Cell Biol. 1999, 31, 539–544. [Google Scholar] [CrossRef]
- Delacoux, F.; Fichard, A.; Geourjon, C.; Garrone, R.; Ruggiero, F. Molecular features of the collagen V heparin binding site. J. Biol. Chem. 1998, 273, 15069–15076. [Google Scholar] [CrossRef]
- Delacoux, F.; Fichard, A.; Cogne, S.; Garrone, R.; Ruggiero, F. Unraveling the amino acid sequence crucial for heparin binding to collagen V. J. Biol. Chem. 2000, 275, 29377–29382. [Google Scholar] [CrossRef]
- LeBaron, R.; Höök, A.; Esko, J.D.; Gay, S.; Höök, M. Binding of heparan sulfate to type V collagen. A mechanism of cell-substrate adhesion. J. Biol. Chem. 1989, 264, 7950–7956. [Google Scholar] [CrossRef]
- Vaughan-Thomas, A.; Young, R.D.; Phillips, A.C.; Duance, V.C. Characterization of type XI collagen-glycosaminoglycan interactions. J. Biol. Chem. 2001, 276, 5303–5309. [Google Scholar] [CrossRef] [PubMed]
- Henriquez, J.; Casar, J.C.; Fuentealba, L.; Carey, D.J.; Brandan, E. Extracellular matrix histone H1 binds to perlecan, is present in regenerating skeletal muscle and stimulates myoblast proliferation. J. Cell Sci. 2002, 115, 2041–2051. [Google Scholar] [CrossRef] [PubMed]
- Watson, K.; Gooderham, N.J.; Davies, D.S.; Edwards, R.J. Nucleosomes bind to cell surface proteoglycans. J. Biol. Chem. 1999, 274, 21707–21713. [Google Scholar] [CrossRef]
- Berasain, C.; Avila, M.A. Amphiregulin. Semin. Cell Dev. Biol. 2014, 28, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Dahlhoff, M.; Wolf, E.; Schneider, M.R. The ABC of BTC: Structural properties and biological roles of betacellulin. Semin. Cell Dev. Biol. 2014, 28, 42–48. [Google Scholar] [CrossRef]
- Singh, B.; Carpenter, G.; Coffey, R.J. EGF receptor ligands: Recent advances. F1000Research 2016, 5, F1000 Faculty Rev-2270. [Google Scholar] [CrossRef]
- Birchmeier, C.; Bennett, D.L. Neuregulin/ErbB Signaling in Developmental Myelin Formation and Nerve Repair. Curr. Top. Dev. Biol. 2016, 116, 45–64. [Google Scholar] [CrossRef]
- Gonzalez-Castillo, C.; Ortuno-Sahagun, D.; Guzman-Brambila, C.; Pallas, M.; Rojas-Mayorquin, A.E. Pleiotrophin as a central nervous system neuromodulator, evidences from the hippocampus. Front. Cell. Neurosci. 2014, 8, 443. [Google Scholar] [CrossRef]
- Papadimitriou, E.; Pantazaka, E.; Castana, P.; Tsalios, T.; Polyzos, A.; Beis, D. Pleiotrophin and its receptor protein tyrosine phosphatase beta/zeta as regulators of angiogenesis and cancer. Biochim. Biophys. Acta 2016, 1866, 252–265. [Google Scholar] [CrossRef] [PubMed]
- Sorrelle, N.; Dominguez, A.T.A.; Brekken, R.A. From top to bottom: Midkine and pleiotrophin as emerging players in immune regulation. J. Leukoc. Biol. 2017, 102, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Kadomatsu, K.; Kishida, S.; Tsubota, S. The heparin-binding growth factor midkine: The biological activities and candidate receptors. J. Biochem. 2013, 153, 511–521. [Google Scholar] [CrossRef]
- Liedert, A.; Schinke, T.; Ignatius, A.; Amling, M. The role of midkine in skeletal remodelling. Br. J. Pharmacol. 2014, 171, 870–878. [Google Scholar] [CrossRef]
- Winkler, C.; Yao, S. The midkine family of growth factors: Diverse roles in nervous system formation and maintenance. Br. J. Pharmacol. 2014, 171, 905–912. [Google Scholar] [CrossRef]
- Beenken, A.; Mohammadi, M. The FGF family: Biology, pathophysiology and therapy. Nat. Rev. Drug Discov. 2009, 8, 235–253. [Google Scholar] [CrossRef] [PubMed]
- Katoh, M. Therapeutics Targeting FGF Signaling Network in Human Diseases. Trends Pharmacol. Sci. 2016, 37, 1081–1096. [Google Scholar] [CrossRef]
- Zhang, X.; Ibrahimi, O.A.; Olsen, S.K.; Umemori, H.; Mohammadi, M.; Ornitz, D.M. Receptor specificity of the fibroblast growth factor family. The complete mammalian FGF family. J. Biol. Chem. 2006, 281, 15694–15700. [Google Scholar] [CrossRef] [PubMed]
- Cohick, W.S.; Clemmons, D.R. The insulin-like growth factors. Annu. Rev. Physiol. 1993, 55, 131–153. [Google Scholar] [CrossRef]
- Furstenberger, G.; Senn, H.J. Insulin-like growth factors and cancer. Lancet Oncol. 2002, 3, 298–302. [Google Scholar] [CrossRef]
- Kessler, S.M.; Haybaeck, J.; Kiemer, A.K. Insulin-Like Growth Factor 2—The Oncogene and its Accomplices. Curr. Pharm. Des. 2016, 22, 5948–5961. [Google Scholar] [CrossRef]
- O’Dell, S.D.; Day, I.N. Insulin-like growth factor II (IGF-II). Int. J. Biochem. Cell Biol. 1998, 30, 767–771. [Google Scholar] [CrossRef] [PubMed]
- Fredriksson, L.; Li, H.; Eriksson, U. The PDGF family: Four gene products form five dimeric isoforms. Cytokine Growth Factor. Rev. 2004, 15, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Colak, S.; Ten Dijke, P. Targeting TGF-beta Signaling in Cancer. Trends Cancer 2017, 3, 56–71. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Chen, S.; Zeng, J. TGFbeta signaling: A complex role in tumorigenesis (Review). Mol. Med. Rep. 2018, 17, 699–704. [Google Scholar] [CrossRef]
- Billings, P.; Bizzaro, C.; Yang, E.; Chung, J.; Mundy, C.; Pacifici, M. Human and mouse activin genes: Divergent expression of activin A protein variants and identification of a novel heparan sulfate-binding domain in activin B. PLoS ONE 2020, 15, e0229254. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Zhou, Y.; Ying, K.; Ruan, W. IGFBP, a novel target of lung cancer? Clin. Chim. Acta 2017, 466, 172–177. [Google Scholar] [CrossRef]
- Meng, X.M.; Nikolic-Paterson, D.J.; Lan, H.Y. TGF-beta: The master regulator of fibrosis. Nat. Rev. Nephrol. 2016, 12, 325–338. [Google Scholar] [CrossRef]
- Oklu, R.; Hesketh, R. The latent transforming growth factor beta binding protein (LTBP) family. Biochem. J. 2000, 352 Pt 3, 601–610. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, K.; Han, B.; Xu, Z.; Gao, X. The emerging role of follistatin under stresses and its implications in diseases. Gene 2018, 639, 111–116. [Google Scholar] [CrossRef]
- Quinsey, N.S.; Greedy, A.L.; Bottomley, S.P.; Whisstock, J.C.; Pike, R.N. Antithrombin: In control of coagulation. Int. J. Biochem. Cell Biol. 2004, 36, 386–389. [Google Scholar] [CrossRef]
- Mast, A.E. Tissue Factor Pathway Inhibitor: Multiple Anticoagulant Activities for a Single Protein. Arter. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.P.; Ellery, P.E.; Maroney, S.A.; Mast, A.E. Biology of tissue factor pathway inhibitor. Blood 2014, 123, 2934–2943. [Google Scholar] [CrossRef] [PubMed]
- Ansell, J. Factor Xa or thrombin: Is factor Xa a better target? J. Thromb. Haemost. 2007, 5 (Suppl. 1), 60–64. [Google Scholar] [CrossRef]
- Borensztajn, K.; Peppelenbosch, M.P.; Spek, C.A. Factor Xa: At the crossroads between coagulation and signaling in physiology and disease. Trends Mol. Med. 2008, 14, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Daubie, V.; Pochet, R.; Houard, S.; Philippart, P. Tissue factor: A mini-review. J. Tissue Eng. Regen. Med. 2007, 1, 161–169. [Google Scholar] [CrossRef]
- Jackson, C.M. Factor X. Prog. Hemost. Thromb. 1984, 7, 55–109. [Google Scholar]
- Franchini, M.; Mannucci, P.M. Thrombin and cancer: From molecular basis to therapeutic implications. Semin. Thromb. Hemost. 2012, 38, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Olson, S.T.; Bjork, I. Regulation of thrombin activity by antithrombin and heparin. Semin. Thromb. Hemost. 1994, 20, 373–409. [Google Scholar] [CrossRef]
- Siller-Matula, J.M.; Schwameis, M.; Blann, A.; Mannhalter, C.; Jilma, B. Thrombin as a multi-functional enzyme. Focus on in vitro and in vivo effects. Thromb. Haemost. 2011, 106, 1020–1033. [Google Scholar] [CrossRef]
- Strukova, S.M. Thrombin as a regulator of inflammation and reparative processes in tissues. Biochemistry 2001, 66, 8–18. [Google Scholar]
- Lerman, I.; Hammes, S.R. Neutrophil elastase in the tumor microenvironment. Steroids 2018, 133, 96–101. [Google Scholar] [CrossRef]
- Alatrash, G.; Garber, H.R.; Zhang, M.; Sukhumalchandra, P.; Qiu, Y.; Jakher, H.; Perakis, A.A.; Becker, L.; Yoo, S.Y.; Dwyer, K.C.; et al. Cathepsin G is broadly expressed in acute myeloid leukemia and is an effective immunotherapeutic target. Leukemia 2017, 31, 234–237. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, R.; Sali, A.; Ghildyal, N.; Karplus, M.; Stevens, R.L. Packaging of proteases and proteoglycans in the granules of mast cells and other hematopoietic cells. A cluster of histidines on mouse mast cell protease 7 regulates its binding to heparin serglycin proteoglycans. J. Biol. Chem. 1995, 270, 19524–19531. [Google Scholar] [CrossRef]
- Rother, S.; Samsonov, S.A.; Hofmann, T.; Blaszkiewicz, J.; Köhling, S.; Moeller, S.; Schnabelrauch, M.; Rademann, J.; Kalkhof, S.; von Bergen, M.; et al. Structural and Functional Insights Into the Interaction of Sulfated Glycosaminoglycans with Tissue Inhibitor of metalloproteinase-3—A Possible Regulatory Role on Extracellular Matrix Homeostasis. Acta Biomater. 2016, 45, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Troeberg, L.; Lazenbatt, C.; Anower-E.-Khuda, M.F.; Freeman, C.; Federov, O.; Habuchi, H.; Habuchi, O.; Kimata, K.; Nagase, H. Sulfated Glycosaminoglycans Control the Extracellular Trafficking and the Activity of the Metalloprotease Inhibitor TIMP-3. Chem. Biol. 2014, 21, 1300–1309. [Google Scholar] [CrossRef]
- Yu, W.; Yu, S.; Meng, Q.; Brew, K.; Woessner, J.F., Jr. TIMP-3 Binds to Sulfated Glycosaminoglycans of the Extracellular Matrix. J. Biol. Chem. 2000, 275, 31226–31232. [Google Scholar] [CrossRef] [PubMed]
- Huntington, J. Mechanisms of glycosaminoglycan activation of the serpins in hemostasis. J. Thromb. Haemost. 2003, 1, 1535–1549. [Google Scholar] [CrossRef] [PubMed]
- Tollefsen, D. Vascular dermatan sulfate and heparin cofactor II. Prog. Mol. Biol. Transl. Sci. 2010, 93, 351–372. [Google Scholar] [PubMed]
- Hutadilok, N.; Ghosh, P.; Brooks, P.M. Binding of haptoglobin, inter-alpha-trypsin inhibitor, and alpha 1 proteinase inhibitor to synovial fluid hyaluronate and the influence of these proteins on its degradation by oxygen derived free radicals. Ann. Rheum. Dis. 1988, 47, 377–385. [Google Scholar] [CrossRef]
- Kotla, N.; Bonam, S.R.; Rasala, S.; Wankar, J.; Bohara, R.A.; Bayry, J.; Rochev, Y.; Pandit, A. Recent advances and prospects of hyaluronan as a multifunctional therapeutic system. J. Control. Release 2021, 336, 598–620. [Google Scholar] [CrossRef]
- Lierova, A.; Kasparova, J.; Filipova, A.; Cizkova, J.; Pekarova, L.; Korecka, L.; Mannova, N.; Bilkova, Z.; Sinkorova, Z. Hyaluronic Acid: Known for Almost a Century, but Still in Vogue. Pharmaceutics 2022, 14, 838. [Google Scholar] [CrossRef] [PubMed]
- Hayes, A.; Melrose, J. Glycosaminoglycan and Proteoglycan Biotherapeutics in Articular Cartilage Protection and Repair Strategies: Novel Approaches to Visco-supplementation in Orthobiologics. Adv. Ther. 2019, 2, 1900034. [Google Scholar] [CrossRef]
- Kim, Y.; Guilak, F. Engineering Hyaluronic Acid for the Development of New Treatment Strategies for Osteoarthritis. Int. J. Mol. Sci. 2022, 23, 8662. [Google Scholar] [CrossRef]
- Agarwal, G.; Agiwal, S.; Srivastava, A. Hyaluronic acid containing scaffolds ameliorate stem cell function for tissue repair and regeneration. Int. J. Biol. Macromol. 2020, 165, 388–401. [Google Scholar] [CrossRef] [PubMed]
- Lord, M.; Melrose, J.; Day, A.J.; Whitelock, J.M. The Inter-α-Trypsin Inhibitor Family: Versatile Molecules in Biology and Pathology. J. Histochem. Cytochem. 2020, 68, 907–927. [Google Scholar] [CrossRef] [PubMed]
- Fath, M.; Wu, X.; Hileman, R.E.; Linhardt, R.J.; Kashem, M.A.; Nelson, R.M.; Wright, C.D.; Abraham, W.M. Interaction of secretory leukocyte protease inhibitor with heparin inhibits proteases involved in asthma. J. Biol. Chem. 1998, 273, 13563–13569. [Google Scholar] [CrossRef]
- Walter, M.; Plotnick, M.; Schechter, N.M. Inhibition of human mast cell chymase by secretory leukocyte proteinase inhibitor: Enhancement of the interaction by heparin. Arch. Biochem. Biophys. 1996, 327, 81–88. [Google Scholar] [CrossRef]
- Neese, L.; Pratt, C.W.; Church, F.C. Modulation of protein C inhibitor activity. Blood Coagul. Fibrinolysis 1994, 5, 737–746. [Google Scholar] [CrossRef]
- Pratt, C.; Church, F.C. Heparin binding to protein C inhibitor. J. Biol. Chem. 1992, 267, 8789–8794. [Google Scholar] [CrossRef]
- Chao, J.; Bledsoe, G.; Chao, L. Protective Role of Kallistatin in Vascular and Organ Injury. Hypertension 2016, 68, 533–541. [Google Scholar] [CrossRef]
- Bhakuni, T.; Ali, M.F.; Ahmad, I.; Bano, S.; Ansari, S.; Jairajpuri, M.A. Role of heparin and non heparin binding serpins in coagulation and angiogenesis: A complex interplay. Arch. Biochem. Biophys. 2016, 604, 128–142. [Google Scholar] [CrossRef] [PubMed]
- Aymonnier, K.; Kawecki, C.; Venisse, L.; Boulaftali, Y.; Christophe, O.D.; Lenting, P.J.; Arocas, V.; de Raucourt, E.; Denis, C.V.; Bouton, M.C. Targeting protease nexin-1, a natural anticoagulant serpin, to control bleeding and improve hemostasis in hemophilia. Blood 2019, 134, 1632–1644. [Google Scholar] [CrossRef] [PubMed]
- Selbonne, S.; Azibani, F.; Iatmanen, S.; Boulaftali, Y.; Richard, B.; Jandrot-Perrus, M.; Bouton, M.C.; Arocas, V. In vitro and in vivo antiangiogenic properties of the serpin protease nexin-1. Mol. Cell Biol. 2012, 32, 1496–1505. [Google Scholar] [CrossRef] [PubMed]
- Rein, C.; Desai, U.R.; Church, F.C. Serpin-glycosaminoglycan interactions. Methods Enzym. Enzymol. 2011, 501, 105–137. [Google Scholar]
- Monard, D. SERPINE2/Protease Nexin-1 in vivo multiple functions: Does the puzzle make sense? Semin. Cell Dev. Biol. 2017, 62, 160–169. [Google Scholar] [CrossRef]
- Donovan, F.; Vaughan, P.J.; Cunningham, D.D. Regulation of protease nexin-1 target protease specificity by collagen type IV. J. Biol. Chem. 1994, 269, 17199–17205. [Google Scholar] [CrossRef]
- Ruiz-Gómez, G.; Vogel, S.; Möller, S.; Pisabarro, M.T.; Hempel, U. Glycosaminoglycans influence enzyme activity of MMP2 and MMP2/TIMP3 complex formation—Insights at cellular and molecular level. Sci. Rep. 2019, 9, 4905. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Lee, K.B.; Linhardt, R.J. SPR Biosensor Probing the Interactions between TIMP-3 and Heparin/GAGs. Biosensors 2015, 5, 500–512. [Google Scholar] [CrossRef] [PubMed]
- Schnabelrauch, M.; Scharnweber, D.; Schiller, J. Sulfated glycosaminoglycans as promising artificial extracellular matrix components to improve the regeneration of tissues. Curr. Med. Chem. 2013, 20, 2501–2523. [Google Scholar] [CrossRef]
- Wayne, G.; Deng, S.J.; Amour, A.; Borman, S.; Matico, R.; Carter, H.L.; Murphy, G. TIMP-3 inhibition of ADAMTS-4 (Aggrecanase-1) is modulated by interactions between aggrecan and the C-terminal domain of ADAMTS-4. J. Biol. Chem. 2007, 289, 20991–20998. [Google Scholar] [CrossRef] [PubMed]
- El Saadani, M.; Ahmed, S.M.; Jacovides, C.; Lopez, A.; Johnson, V.E.; Kaplan, L.J.; Schwab, C.W.; Smith, D.H.; Pascual, J.L. Antithrombin III ameliorates post-traumatic brain injury cerebral leukocyte mobilization enhancing recovery of blood brain barrier integrity. J. Trauma Acute Care Surg. 2021, 90, 274–280. [Google Scholar] [CrossRef]
- Petzelbauer, E.; Seiffert, D.; Beckmann, R.; Pusch, B.; Geiger, M.; Binder, B.R. Modulation of heparin cofactor II activity by glycosaminoglycans and adhesive glycoproteins. Thromb. Res. 1992, 66, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Verhamme, I.; Bock, P.E.; Jackson, C.M. The preferred pathway of glycosaminoglycan-accelerated inactivation of thrombin by heparin cofactor II. J. Biol. Chem. 2004, 279, 9785–9795. [Google Scholar] [CrossRef]
- Ho, G.; Broze GJJr Schwartz, A.L. Role of heparan sulfate proteoglycans in the uptake and degradation of tissue factor pathway inhibitor-coagulation factor Xa complexes. J. Biol. Chem. 1997, 272, 16838–16844. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Stack, S.M.; Lakka, S.S.; Khan, A.J.; Woodley, D.T.; Rao, J.S.; Rao, C.N. Matrix localization of tissue factor pathway inhibitor-2/matrix-associated serine protease inhibitor (TFPI-2/MSPI) involves arginine-mediated ionic interactions with heparin and dermatan sulfate: Heparin accelerates the activity of TFPI-2/MSPI toward plasmin. Arch. Biochem. Biophys. 1999, 370, 112–118. [Google Scholar]
- Mine, S.; Yamazaki, T.; Miyata, T.; Hara, S.; Kato, H. Structural mechanism for heparin-binding of the third Kunitz domain of human tissue factor pathway inhibitor. Biochemistry 2002, 41, 78–85. [Google Scholar] [CrossRef]
- Xu, X.; Takano, R.; Nagai, Y.; Yanagida, T.; Kamei, K.; Kato, H.; Kamikubo, Y.; Nakahara, Y.; Kumeda, K.; Hara, S. Effect of heparin chain length on the interaction with tissue factor pathway inhibitor (TFPI). Int. J. Biol. Macromol. 2002, 30, 151–160. [Google Scholar] [CrossRef]
- Li, W.; Huntington, J.A. The heparin binding site of protein C inhibitor is protease-dependent. J. Biol. Chem. 2008, 283, 36039–36045. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, H.; Keijer, J.; Preissner, K.T.; Gebbink, R.K.; Pannekoek, H. Functional interaction of plasminogen activator inhibitor type 1 (PAI-1) and heparin. Biochemistry 1991, 30, 1021–1028. [Google Scholar] [CrossRef] [PubMed]
- Hagège, J.; Delarue, F.; Peraldi, M.N.; Sraer, J.D.; Rondeau, E. Heparin selectively inhibits synthesis of tissue type plasminogen activator and matrix deposition of plasminogen activator inhibitor 1 by human mesangial cells. Lab. Investig. 1994, 71, 828–837. [Google Scholar] [PubMed]
- Rovelli, G.; Stone, S.R.; Guidolin, A.; Sommer, J.; Monard, D. Characterization of the heparin-binding site of glia-derived nexin/protease nexin-1. Biochemistry 1992, 31, 3542–3549. [Google Scholar] [CrossRef] [PubMed]
- Qiao, M.; Lin, L.; Xia, K.; Li, J.; Zhang, X.; Linhardt, R.J. Recent advances in biotechnology for heparin and heparan sulfate analysis. Talanta 2020, 219, 121270. [Google Scholar] [CrossRef]
- Allen, N.; Bennett, M.L.; Foo, L.C.; Wang, G.X.; Chakraborty, C.; Smith, S.J.; Barres, B.A. Astrocyte glypicans 4 and 6 promote formation of excitatory synapses via GluA1 AMPA receptors. Nature 2012, 486, 410–414. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.; O’Shea, T.M.; Burda, J.E.; Ao, Y.; Barlatey, S.L.; Bernstein, A.M.; Kim, J.H.; James, N.D.; Rogers, A.; Kato, B.; et al. Required growth facilitators propel axon regeneration across complete spinal cord injury. Nature 2018, 561, 396–400. [Google Scholar] [CrossRef]
- Buzanska, L.; Zychowicz, M.; Kinsner-Ovaskainen, A. Bioengineering of the Human Neural Stem Cell Niche: A Regulatory Environment for Cell Fate and Potential Target for Neurotoxicity. Results Probl. Cell Differ. 2018, 66, 207–230. [Google Scholar]
- Fawcett, J.; Kwok, J.C.F. Proteoglycan Sulphation in the Function of the Mature Central Nervous System. Front. Integr. Neurosci. 2022, 16, 895493. [Google Scholar] [CrossRef]
- Hayes, A.; Melrose, J. Electro-Stimulation, a Promising Therapeutic Treatment Modality for Tissue Repair: Emerging Roles of Sulfated Glycosaminoglycans as Electro-Regulatory Mediators of Intrinsic Repair Processes. Adv. Ther. 2020, 3, 2000151. [Google Scholar] [CrossRef]



| Member | Characteristics | Distribution/Function | Ref. |
|---|---|---|---|
| Intracellular | |||
| Serglycin | Intracellular granule Heparin, HS, and CS–PG, 17.6 kDa core protein, GAGs are attached to central 16 amino acid SGR domain in SRGN | Expressed by haematopoietic cells, neutrophils, monocytes, lymphocytes, platelets, macrophages, mast cells, megakaryocytes, chondrocytes, endothelial, and embryonic stem cells. Forms inactive protease complexes in intracellular granules | [62,63,64,65,199] |
| Cell-associated | |||
| The Glypicans | GPI anchored HS–PG endocytic co-receptors | Cell surface co-receptors for GFs, ECM proteins, proteases, protease inhibitors, Wnt signaling. | [4,6,69,70,200] |
| Glypican-1 | 64 kDa core protein | GPC-1 is expressed in embryonic CNS and skeletal systems and in adult kidney and skeletal muscle. HS side chains facilitate interactions with Wnt/β-catenin, Hh, FGFs, IGFs, VEGF, TGF-β. | [69,201,202,203,204,205,206] |
| Glypican-2 cerebroglycan | 57 kDa core protein | Preferentially expressed in neural tissues, binds Midkine and regulates neural network development. | [207,208,209] |
| Glypican-3 | 69 kDa core protein | Roles in mesoderm and intestine, binds FGF2, regulates tissue development, neural network assembly, modulates BMP/FGF mediated renal branching, tissue morphogenesis, Hh signaling. | [4,69,70,116,210,211,212,213,214] |
| Glypican-4 K-glypican | 57 kDa core protein | Found in blood vessels, kidney, brain, promotes CNS development, expressed in the ventricular neuroprogenitor stem cells, committed neurons do not express GPC4, expressed by cortical, hippocampal, cerebellar astrocytes, roles in synapse formation/fore-brain development. | [215] |
| Glypican-5 | 84 kDa core protein, a CS chain may modulate HS-GF GPC-5 interactions | Developmentally regulated in kidney, limbs, and brain. Controls cell growth and differentiation. | [216] |
| Glypican-6 | 65 kDa core protein | Regulates ERK1/2 signaling neurotransmitter receptor localization in postsynaptic membrane, roles in presynapse assembly and action, broad distribution in fetal kidney, adult ovary, liver, gall bladder. | [217] |
| Syndecans | Single pass TM PGs containing 3–5 CS/HS chains | Co-receptors of FGFs, VEGFs, TGFβ, AT, Fibronectin | [77,80,218] |
| Syndecan-1 | 33 kDa core protein | Found in skin, liver, kidney, lung, and small intestine. | [219,220,221,222] |
| Syndecan-2 fibroglycan | 23 kDa core protein | Found in brain, heart, skeletal muscle, liver, kidney, and lung; regulates TGFβ signaling, angiogenic sprouting. | [223,224] |
| Syndecan-3 N-syndecan | 43 kDa core protein | Found in brain, heart, skin, and skeletal muscle. Colocalizes with TFPI on surface of endothelial and SMCs. | [225,226,227] |
| Syndecan-4 Ryudocan Amphiglycan | 22 kDa core protein | Found in brain, liver, kidney, lung, heart, skeletal muscle, skin, and pancreas. Regulates FGF-2, FAK, ADAMTS-4 activation, and insulin secretion in pancreas. | [225,228,229,230,231,232] |
| Betaglycan TGF-β-R III | 300 kDa Cell-surface CS/HS co-receptor, binds TGF-β via core protein, and FGF-2 via HS. | Multifunctional co-receptor binds TGF-β2, InhA, BMP-9, BMP-10, and FGF-2. MMPs and plasmin shedses release a soluble ecto-domain that antagonizes cell-bound betaglycan activity. | [39,43,44,84,233] |
| CD44 Epican | Epican is a HS isoform. 80–250 kDa TM HS substituted CD44 HA receptor. | Interacts with osteopontin, collagens, and MMPs; stabilizes/remodebrain ECM and generates focal Interacts with osteopontin, collagens, MMPs, stabilises/remodels ECM. Attaches neural cells to brain ECM, generates focal adhesion complexes/traction forces during cell migration and spreading. Entraps HA and water in brain, preserving ionic gradients and neural cell niches. | [96,97,98,104,105,234,235,236] |
| CD47 | Haematopoietic, 50 kDa HS/CS TM high affinity TSP-1 receptor of CD47-SIRPα system. Protects cells from degradation by macrophages. | HS on CD47 inhibits T cell receptor signaling with TSP-1. Regulates cell migration, proliferation, and vascular cell survival in innate/adaptive immune regulation. TSP-1 inhibits eNOS activation/NO production. | [87,89,90,93,94] |
| NRP-1 | 120 kDa single span TM co-receptor, has roles in tumor vascularization in brain, prostate, breast, colon, and lung cancer. Is a SARS-CoV-2 receptor in COVID-19 disease. | Soluble NRP-1 antagonizes cellular NRP-1 activity in axonal guidance, angiogenesis, cell survival, migration, and tumor invasion. Binds HS-binding GFs, VEGF 165, and placenta growth factor; modulates VEGF/VEGFR2 signaling. | [35,38,237,238,239,240,241] |
| Neurexins α, β, γ | α, β, γ isoforms interact with neuroligin and LRRTM-2, 4 to stabilize synapse and neural-signal transduction. | Provides synaptic stabilization and neuronal plasticity in neural networks. Nrxn dysregulation is implicated in neuro-degenerative cognitive disorders. | [29,181,183] |
| Extracellular matrix | |||
| Collagen XVIII | Homotrimeric multiplexin (300 kDa) with 3 HS chains, in long, medium, and short isoforms; multiple collagenous and NC-domains, 20 kDa C-terminal anti-angiogenic endostatin peptide. | Interacts with laminin, perlecan, fibulin, and nidogen, forming network formations that stabilize basement membranes. LamG and TSP-1 protein interactive domains, and a homologous Frizzled receptor cysteine rich domain, have Wnt inhibitory activity. | [9,139,140,242,243,244,245,246] |
| Agrin | ~220 kDa core protein; HS/CS increases size to 400 kDa. Multiple Lam G and EGF repeats, central Ser–Thr-rich domain contains HS. | Component of cartilage, roles in synaptic organization, and NMJ assembly facilitating regulation of neuro-muscular control and mechanotransduction. | [9,149,155,166,247] |
| Perlecan | 467 kDa core protein with five functional domains. Domain I has 3 HS/CS/KS chains, domain V also has GAG. Domain IV has 23 Ig repeats, Domain V has multiple LamG/EGF repeats and antiangiogenic endorepellin peptide, domain V promotes BBB and angiogenic repair | Ubiquitous HS, or HS/CS PG, sequesters acetylcholinesterase in NMJ. Protects chondrocyte viability. Promotes vascularization, 20 kDa domain V fragment inhibits angiogenesis, and stabilises BM, BBB, BVs, and stem cell niche. HS chains sequester GFs to promote cell proliferation/differentiation, tissue repair, and Domain II LDLR transports Wnt and ShH in tissue development. Reduces SMC proliferation and atherosclerotic plaque deposition. | [112,144,150,151,154,160,163,248,249,250,251] |
| Eyes-shut | Modular 250–350 kDa retinal PG related to agrin/perlecan; central Ser–Thr module binds HS chains; has multiple EGF and LamG modules. | Organizational roles in the assembly and function of interphotoreceptor ECM in the retinal epithelium, as well as stabilization of ciliary axoneme and ribbon synapse in rods and cones. Vital roles in the photoreceptor ECM stabilization and visual acuity. | [30,252] |
| Pikachurin | Modular 110 kDa photoreceptor HSPG | Interacts with αDG, stabilises photoreceptor | [31,32,119] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farrugia, B.L.; Melrose, J. The Glycosaminoglycan Side Chains and Modular Core Proteins of Heparan Sulphate Proteoglycans and the Varied Ways They Provide Tissue Protection by Regulating Physiological Processes and Cellular Behaviour. Int. J. Mol. Sci. 2023, 24, 14101. https://doi.org/10.3390/ijms241814101
Farrugia BL, Melrose J. The Glycosaminoglycan Side Chains and Modular Core Proteins of Heparan Sulphate Proteoglycans and the Varied Ways They Provide Tissue Protection by Regulating Physiological Processes and Cellular Behaviour. International Journal of Molecular Sciences. 2023; 24(18):14101. https://doi.org/10.3390/ijms241814101
Chicago/Turabian StyleFarrugia, Brooke L., and James Melrose. 2023. "The Glycosaminoglycan Side Chains and Modular Core Proteins of Heparan Sulphate Proteoglycans and the Varied Ways They Provide Tissue Protection by Regulating Physiological Processes and Cellular Behaviour" International Journal of Molecular Sciences 24, no. 18: 14101. https://doi.org/10.3390/ijms241814101
APA StyleFarrugia, B. L., & Melrose, J. (2023). The Glycosaminoglycan Side Chains and Modular Core Proteins of Heparan Sulphate Proteoglycans and the Varied Ways They Provide Tissue Protection by Regulating Physiological Processes and Cellular Behaviour. International Journal of Molecular Sciences, 24(18), 14101. https://doi.org/10.3390/ijms241814101