
Tyrosine Kinase Inhibitor Profiling Using Multiple Forskolin-Responsive Reporter Cells

 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Isolation of Multiple Forskolin Reporter Cell Clones
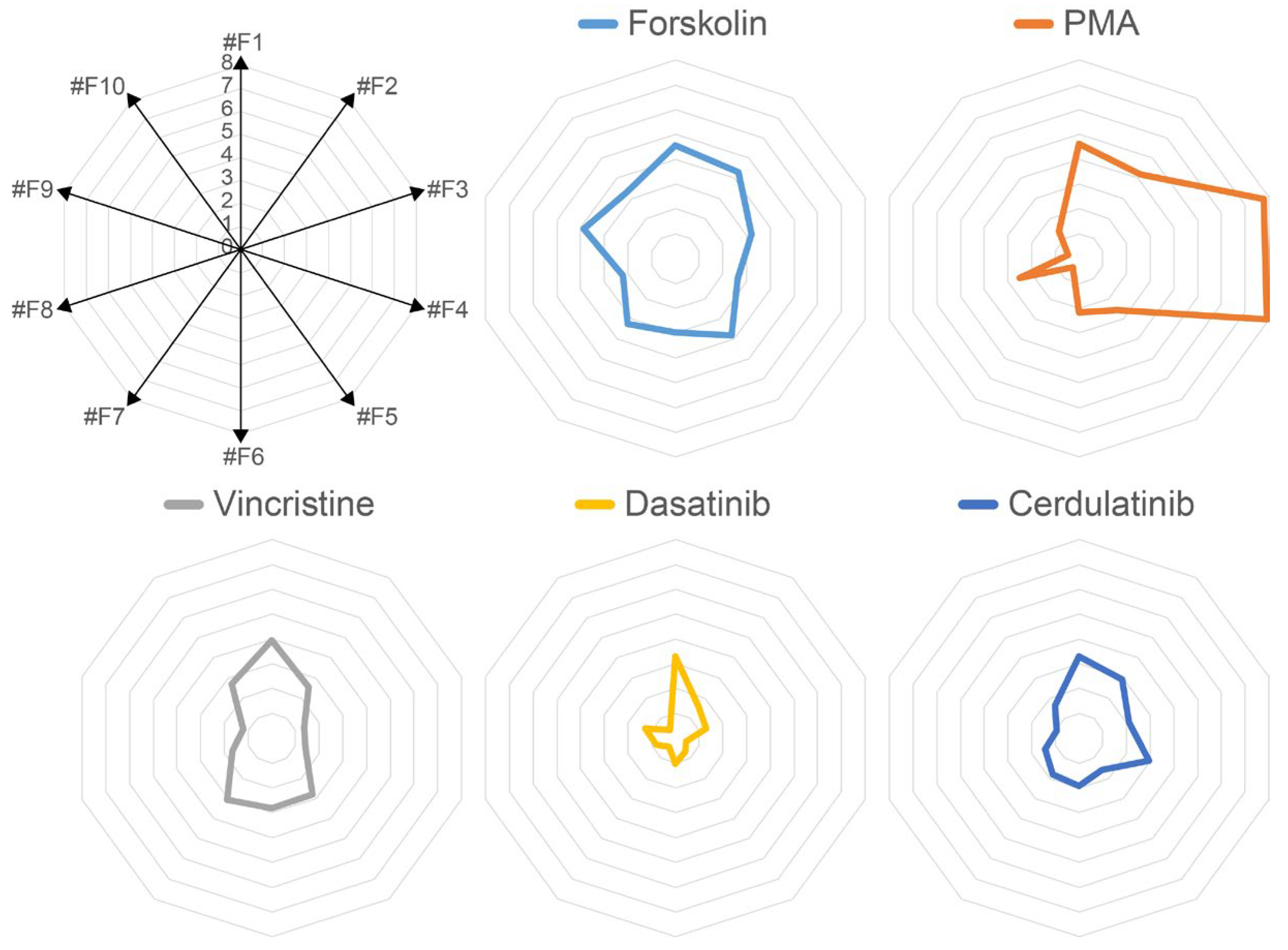
2.2. Each Forskolin-Responsive Clone Has a Unique Reaction Pattern for Multiple Compounds
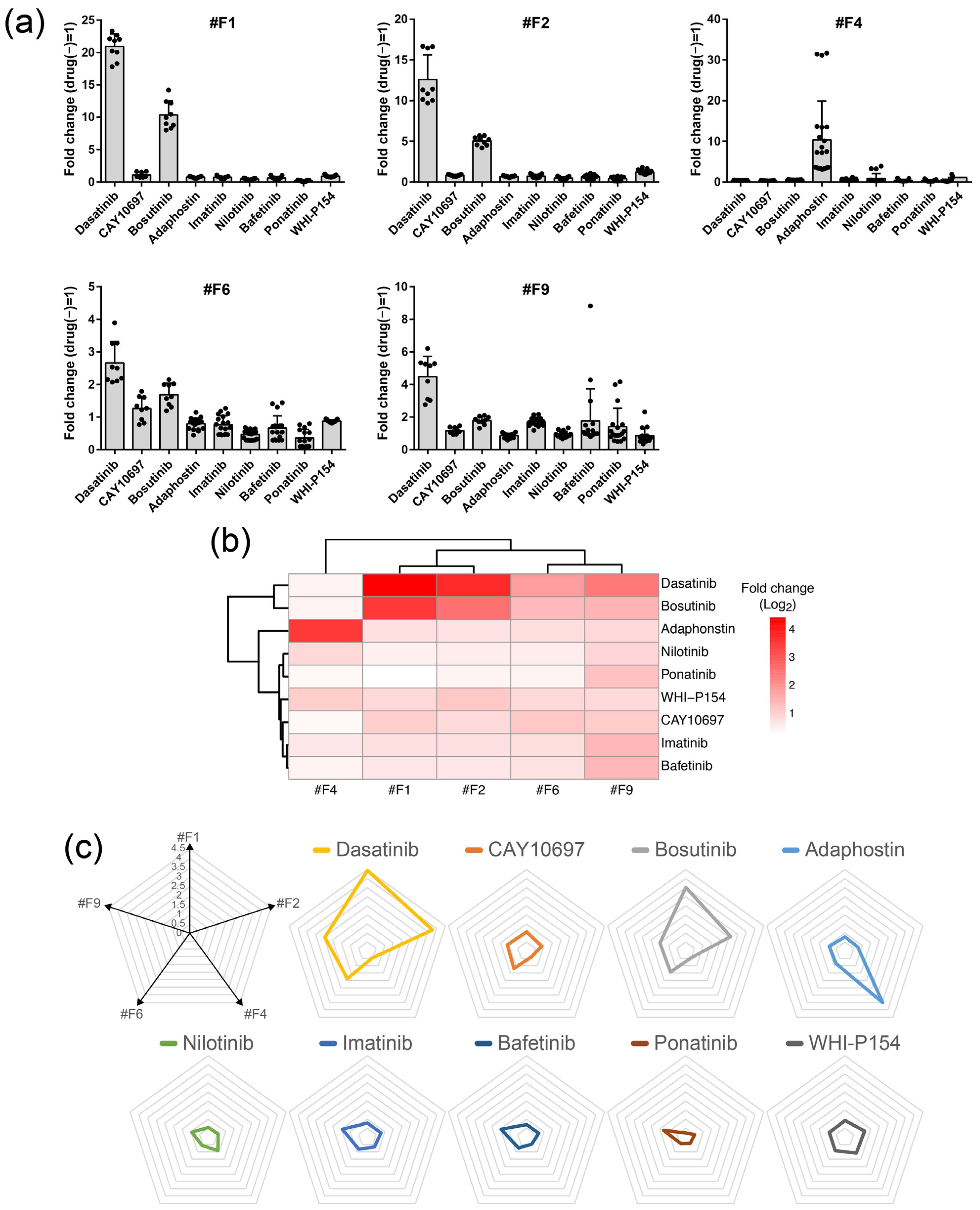
2.3. Compound Profiling Using Multiple Forskolin Reporter Cells
2.4. Evaluation of Other Tyrosine Kinase Inhibitors
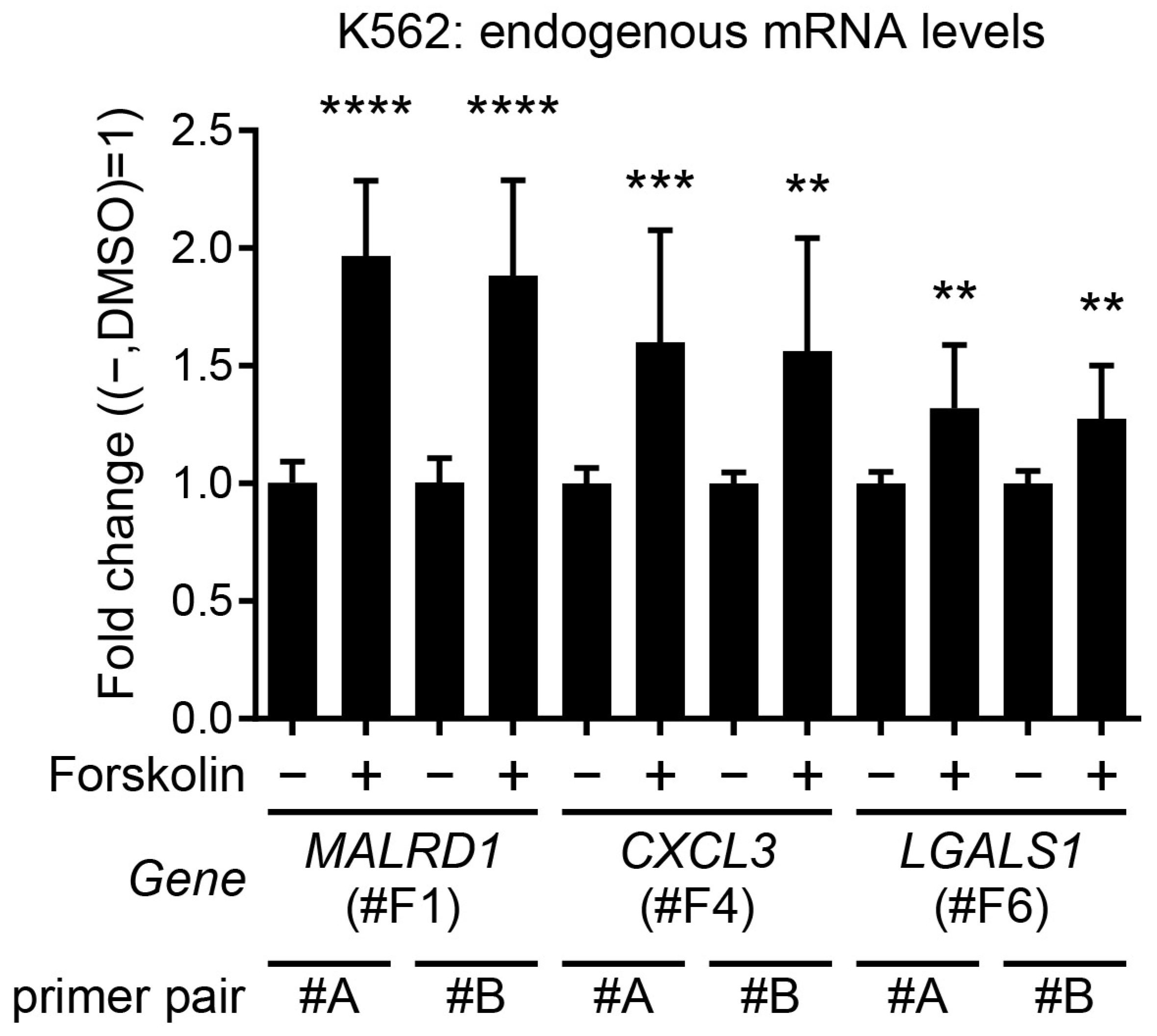
2.5. Identification of Forskolin-Responsive Genes by Splinkerette PCR or 5′Rapid Amplification of cDNA Ends (5′RACE)
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Establishment of Reporter Cells Responsive to Forskolin
4.3. Luciferase Assay
4.4. Hierarchical Clustering Analysis
4.5. 5′RACE
4.6. Real-Time PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ishikawa, K.; Kobayashi, Y.; Wakabayashi, Y.; Watanabe, S.; Semba, K. A highly sensitive trap vector system for isolating reporter cells and identification of responsive genes. Biol. Methods Protoc. 2018, 3, bpy003. [Google Scholar] [CrossRef] [PubMed]
- Ivics, Z.; Li, M.A.; Mates, L.; Boeke, J.D.; Nagy, A.; Bradley, A.; Izsvak, Z. Transposon-mediated genome manipulation in vertebrates. Nat. Methods 2009, 6, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, K.; Tamamura, S.; Semba, K.; Watanabe, S. Establishment of reporter cells that respond to glucocorticoids by a transposon-mediated promoter-trapping system. Eur. J. Pharm. Sci. 2021, 162, 105819. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, K.; Tamamura, S.; Takahashi, N.; Takagi, M.; Semba, K.; Watanabe, S. Isolation of Reporter Cells That Respond to Vitamin A and/or D Using a piggyBac Transposon Promoter-Trapping Vector System. Int. J. Mol. Sci. 2022, 23, 9366. [Google Scholar] [CrossRef]
- Asakawa, K.; Suster, M.L.; Mizusawa, K.; Nagayoshi, S.; Kotani, T.; Urasaki, A.; Kishimoto, Y.; Hibi, M.; Kawakami, K. Genetic dissection of neural circuits by Tol2 transposon-mediated Gal4 gene and enhancer trapping in zebrafish. Proc. Natl. Acad. Sci. USA 2008, 105, 1255–1260. [Google Scholar] [CrossRef]
- Ogura, E.; Okuda, Y.; Kondoh, H.; Kamachi, Y. Adaptation of GAL4 activators for GAL4 enhancer trapping in zebrafish. Dev. Dyn. 2009, 238, 641–655. [Google Scholar] [CrossRef]
- Valent, P. Standard treatment of Ph+ CML in 2010: How, when and where not to use what BCR/ABL1 kinase inhibitor? Eur. J. Clin. Investig. 2010, 40, 918–931. [Google Scholar] [CrossRef]
- Leonetti, F.; Stefanachi, A.; Nicolotti, O.; Catto, M.; Pisani, L.; Cellamare, S.; Carotti, A. BCR-ABL inhibitors in chronic myeloid leukemia: Process chemistry and biochemical profile. Curr. Med. Chem. 2011, 18, 2943–2959. [Google Scholar] [CrossRef]
- Uitdehaag, J.C.; de Roos, J.A.; van Doornmalen, A.M.; Prinsen, M.B.; de Man, J.; Tanizawa, Y.; Kawase, Y.; Yoshino, K.; Buijsman, R.C.; Zaman, G.J. Comparison of the cancer gene targeting and biochemical selectivities of all targeted kinase inhibitors approved for clinical use. PLoS ONE 2014, 9, e92146. [Google Scholar] [CrossRef]
- Rossari, F.; Minutolo, F.; Orciuolo, E. Past, present, and future of Bcr-Abl inhibitors: From chemical development to clinical efficacy. J. Hematol. Oncol. 2018, 11, 84. [Google Scholar] [CrossRef]
- Rix, U.; Colinge, J.; Blatt, K.; Gridling, M.; Remsing Rix, L.L.; Parapatics, K.; Cerny-Reiterer, S.; Burkard, T.R.; Jäger, U.; Melo, J.V.; et al. A target-disease network model of second-generation BCR-ABL inhibitor action in Ph+ ALL. PLoS ONE 2013, 8, e77155. [Google Scholar] [CrossRef] [PubMed]
- Mow, B.M.; Chandra, J.; Svingen, P.A.; Hallgren, C.G.; Weisberg, E.; Kottke, T.J.; Narayanan, V.L.; Litzow, M.R.; Griffin, J.D.; Sausville, E.A.; et al. Effects of the Bcr/abl kinase inhibitors STI571 and adaphostin (NSC 680410) on chronic myelogenous leukemia cells in vitro. Blood 2002, 99, 664–671. [Google Scholar] [CrossRef] [PubMed]
- Orsolic, N.; Golemovic, M.; Quintás-Cardama, A.; Scappini, B.; Manshouri, T.; Chandra, J.; Basic, I.; Giles, F.; Kantarjian, H.; Verstovsek, S. Adaphostin has significant and selective activity against chronic and acute myeloid leukemia cells. Cancer Sci. 2006, 97, 952–960. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.X.; Frey, M.R.; Kohli, R. The Role of FGF19 and MALRD1 in Enterohepatic Bile Acid Signaling. Front. Endocrinol. 2021, 12, 799648. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, L.; Li, H.; Ge, C.; Zhao, F.; Tian, H.; Chen, T.; Jiang, G.; Xie, H.; Cui, Y.; et al. CXCL3 contributes to CD133(+) CSCs maintenance and forms a positive feedback regulation loop with CD133 in HCC via Erk1/2 phosphorylation. Sci. Rep. 2016, 6, 27426. [Google Scholar] [CrossRef]
- Cheng, Y.; Ma, X.L.; Wei, Y.Q.; Wei, X.W. Potential roles and targeted therapy of the CXCLs/CXCR2 axis in cancer and inflammatory diseases. Biochim. Biophys. Acta Rev. Cancer 2019, 1871, 289–312. [Google Scholar] [CrossRef]
- Kusuyama, J.; Komorizono, A.; Bandow, K.; Ohnishi, T.; Matsuguchi, T. CXCL3 positively regulates adipogenic differentiation. J. Lipid Res. 2016, 57, 1806–1820. [Google Scholar] [CrossRef]
- Chen, H.Y.; Liu, Q.; Salter, A.M.; Lomax, M.A. Synergism between cAMP and PPARγ Signalling in the Initiation of UCP1 Gene Expression in HIB1B Brown Adipocytes. PPAR Res. 2013, 2013, 476049. [Google Scholar] [CrossRef]
- Nio-Kobayashi, J.; Hashiba, K.; Sano, M.; Okuda, K.; Duncan, W.C.; Iwanaga, T. Expression Profiles and Possible Roles of Galectins in the Corpus Luteum. Trends Glycosci. Glycotechnol. 2016, 162, E71–E77. [Google Scholar] [CrossRef]
- Casarini, L.; Lispi, M.; Longobardi, S.; Milosa, F.; La Marca, A.; Tagliasacchi, D.; Pignatti, E.; Simoni, M. LH and hCG action on the same receptor results in quantitatively and qualitatively different intracellular signalling. PLoS ONE 2012, 7, e46682. [Google Scholar] [CrossRef]
- Li, J.; Rix, U.; Fang, B.; Bai, Y.; Edwards, A.; Colinge, J.; Bennett, K.L.; Gao, J.; Song, L.; Eschrich, S.; et al. A chemical and phosphoproteomic characterization of dasatinib action in lung cancer. Nat. Chem. Biol. 2010, 6, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Karwaciak, I.; Sałkowska, A.; Kara, K.; Sobalska-Kwapis, M.; Walczak-Drzewiecka, A.; Pułaski, Ł.; Strapagiel, D.; Dastych, J.; Ratajewski, M. SIRT2 Contributes to the Resistance of Melanoma Cells to the Multikinase Inhibitor Dasatinib. Cancers 2019, 11, 673. [Google Scholar] [CrossRef] [PubMed]
- Rix, U.; Hantschel, O.; Dürnberger, G.; Remsing Rix, L.L.; Planyavsky, M.; Fernbach, N.V.; Kaupe, I.; Bennett, K.L.; Valent, P.; Colinge, J.; et al. Chemical proteomic profiles of the BCR-ABL inhibitors imatinib, nilotinib, and dasatinib reveal novel kinase and nonkinase targets. Blood 2007, 110, 4055–4063. [Google Scholar] [CrossRef] [PubMed]
- Gogol-Doring, A.; Ammar, I.; Gupta, S.; Bunse, M.; Miskey, C.; Chen, W.; Uckert, W.; Schulz, T.F.; Izsvak, Z.; Ivics, Z. Genome-wide Profiling Reveals Remarkable Parallels between Insertion Site Selection Properties of the MLV Retrovirus and the piggyBac Transposon in Primary Human CD4(+) T Cells. Mol. Ther. 2016, 24, 592–606. [Google Scholar] [CrossRef] [PubMed]
- Asakawa, K.; Kawakami, K. Targeted gene expression by the Gal4-UAS system in zebrafish. Dev. Growth Differ. 2008, 50, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Yusa, K.; Zhou, L.; Li, M.A.; Bradley, A.; Craig, N.L. A hyperactive piggyBac transposase for mammalian applications. Proc. Natl. Acad. Sci. USA 2011, 108, 1531–1536. [Google Scholar] [CrossRef]
- Devon, R.S.; Porteous, D.J.; Brookes, A.J. Splinkerettes—Improved vectorettes for greater efficiency in PCR walking. Nucleic Acids Res. 1995, 23, 1644–1645. [Google Scholar] [CrossRef]
- Potter, C.J.; Luo, L. Splinkerette PCR for mapping transposable elements in Drosophila. PLoS ONE 2010, 5, e10168. [Google Scholar] [CrossRef]
- Siebring-van Olst, E.; Vermeulen, C.; de Menezes, R.X.; Howell, M.; Smit, E.F.; van Beusechem, V.W. Affordable luciferase reporter assay for cell-based high-throughput screening. J. Biomol. Screen. 2013, 18, 453–461. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 15 June 2023).
- Kolde, R. Pheatmap: Pretty Heatmaps. R Package Version 1.0.12. 2019. Available online: https://CRAN.R-project.org/package=pheatmap (accessed on 15 June 2023).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clone# | Frame | 1 Predicted Response Gene | Method |
|---|---|---|---|
| #F1 | A | MALRD1 | Splinkerette PCR |
| #F2 | A | 2 N.D. | - |
| #F4 | B | CXCL3 | 5′RACE |
| #F6 | B | LGALS1 | 5′RACE |
| #F9 | C | N.D. | - |
| Reagent | Manufacturer | Product Number | CAS Number |
|---|---|---|---|
| Forskolin | Wako, Saitama, Japan | 063-02193 | 66575-29-9 |
| PMA (Phorbol 12-myristate 13-acetate) | Cayman Chemical, Ann Arbor, MI, USA | 10008014 | 16561-29-8 |
| Vincristine Sulfate | Wako | 220-02301 | 2068-78-2 |
| Dasatinib | Wako | ST-7591 | 302962-49-8 |
| Cerdulatinib | Wako | 20076 | 1198300-79-6 |
| Bosutinib | Selleck | S1014 | 380843-75-4 |
| CAY10697 | Cayman Chemical | CAY10697 | 302964-08-5 |
| Adaphostin | Sigma-Aldrich, St. Louis, MO, USA | SML1110 | 241127-58-2 |
| Imatinib | Cayman Chemical | 13139 | 152459-95-5 |
| Nilotinib | Cayman Chemical | 10010422 | 641571-10-0 |
| Bafetinib | Selleck | S1369 | 859212-16-1 |
| Ponatinib | Selleck | S1490 | 943319-70-8 |
| WHI-P154 | Selleck | S2867 | 211555-04-3 |
| Temperature | Time | Repeat |
|---|---|---|
| 94 °C | 2 min | 1 |
| 98 °C | 10 s | 30 |
| 60 °C | 30 s | |
| 68 °C | 2 min | |
| 68 °C | 5 min | 1 |
| 15 °C | ∞ | - |
| Purpose | Primer Sequence (5′ to 3′) |
|---|---|
| Reverse transcription | TGGCCAGTCTATCAGTAAC (GSP1) |
| 5′RACE 1st PCR | GGCCACGCGTCGACTAGTACGGGGGGGGGGGGGGGG (AAP) |
| AGCTTTGATGTCTTGCAGGGAATC (GSP2) | |
| 5′RACE 2nd PCR | GGCCACGCGTCGACTAGTAC (AUAP) |
| GTTGTTCAAGCCTCTCCAGTCTTG (GSP3) | |
| Amplifying pBluesciptII SK(−) for subcloning | AGAGGCTTGAACAACCTTATCGATACCGTCGACCTCG |
| AGTCGACGCGTGGCCCTTGATATCGAATTCCTGCAGC | |
| Colony PCR for subcloned plasmid | GGAAACAGCTATGACCATG |
| GGACTGTACCTACACTCCCAATTG |
| Gene, Primer Pair# | Primer Sequence (5′ to 3′) |
|---|---|
| MALRD1, #A | CTCACATCACAGTTGCAGTCTTG |
| GACAAAGGCACAGTTACCACTTC | |
| MALRD1, #B | ACAGCCTGCAGTCTTACTCAAG |
| TGCTGGGAAGGATTTGGAAGTAG | |
| CXCL3, #A | GAAGTCATAGCCACACTCAAGAATG |
| CGCTGATAAGCTTCTTACTTCTCTC | |
| CXCL3, #B | GTCATAGCCACACTCAAGAATG |
| GCTTCTTACTTCTCTCCTGTCAG | |
| LGALS1, #A | CTGGACTCAATCATGGCTTGTG |
| TTGCTGTCTTTGCCCAGGTTC | |
| LGALS1, #B | CTGGGCAAAGACAGCAACAAC |
| ACACCTCTGCAACACTTCCAG | |
| ACTB | GGCGGCAACACCATGTACCCT |
| AGGGGCCGGACTCGTCATACT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasahara, Y.; Tamamura, S.; Hiyama, G.; Takagi, M.; Nakamichi, K.; Doi, Y.; Semba, K.; Watanabe, S.; Ishikawa, K. Tyrosine Kinase Inhibitor Profiling Using Multiple Forskolin-Responsive Reporter Cells. Int. J. Mol. Sci. 2023, 24, 13863. https://doi.org/10.3390/ijms241813863
Kasahara Y, Tamamura S, Hiyama G, Takagi M, Nakamichi K, Doi Y, Semba K, Watanabe S, Ishikawa K. Tyrosine Kinase Inhibitor Profiling Using Multiple Forskolin-Responsive Reporter Cells. International Journal of Molecular Sciences. 2023; 24(18):13863. https://doi.org/10.3390/ijms241813863
Chicago/Turabian StyleKasahara, Yamato, Sakura Tamamura, Gen Hiyama, Motoki Takagi, Kazuya Nakamichi, Yuta Doi, Kentaro Semba, Shinya Watanabe, and Kosuke Ishikawa. 2023. "Tyrosine Kinase Inhibitor Profiling Using Multiple Forskolin-Responsive Reporter Cells" International Journal of Molecular Sciences 24, no. 18: 13863. https://doi.org/10.3390/ijms241813863
APA StyleKasahara, Y., Tamamura, S., Hiyama, G., Takagi, M., Nakamichi, K., Doi, Y., Semba, K., Watanabe, S., & Ishikawa, K. (2023). Tyrosine Kinase Inhibitor Profiling Using Multiple Forskolin-Responsive Reporter Cells. International Journal of Molecular Sciences, 24(18), 13863. https://doi.org/10.3390/ijms241813863