

Transplantation of Chemical Compound-Induced Cells from Human Fibroblasts Improves Locomotor Recovery in a Spinal Cord Injury Rat Model

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Immature CiN Cells Are Not Fully Differentiated into Neuronal-like Cells
2.2. Transplantation of the Immature CiN Cells Tends to Improve Functional Recovery in Subacute SCI Rat Model
2.3. The Spinal Cord Injury Was Assessed by Performing MRI
2.4. Spinal Cord Area and Amounts of Myelin Sheath after SCI Tended to Be Larger at the Caudal +5 mm Site in the Immature CiN Cells Transplanted Group
2.5. Transplantation of the Immature CiN Cells Tends to Improve Motor Neuron Recovery and Reduce Inflammation in the Caudal +5 mm Site
2.6. Gene Expression of HGF and IL-1RN Was Enhanced in Immature CiN Cells
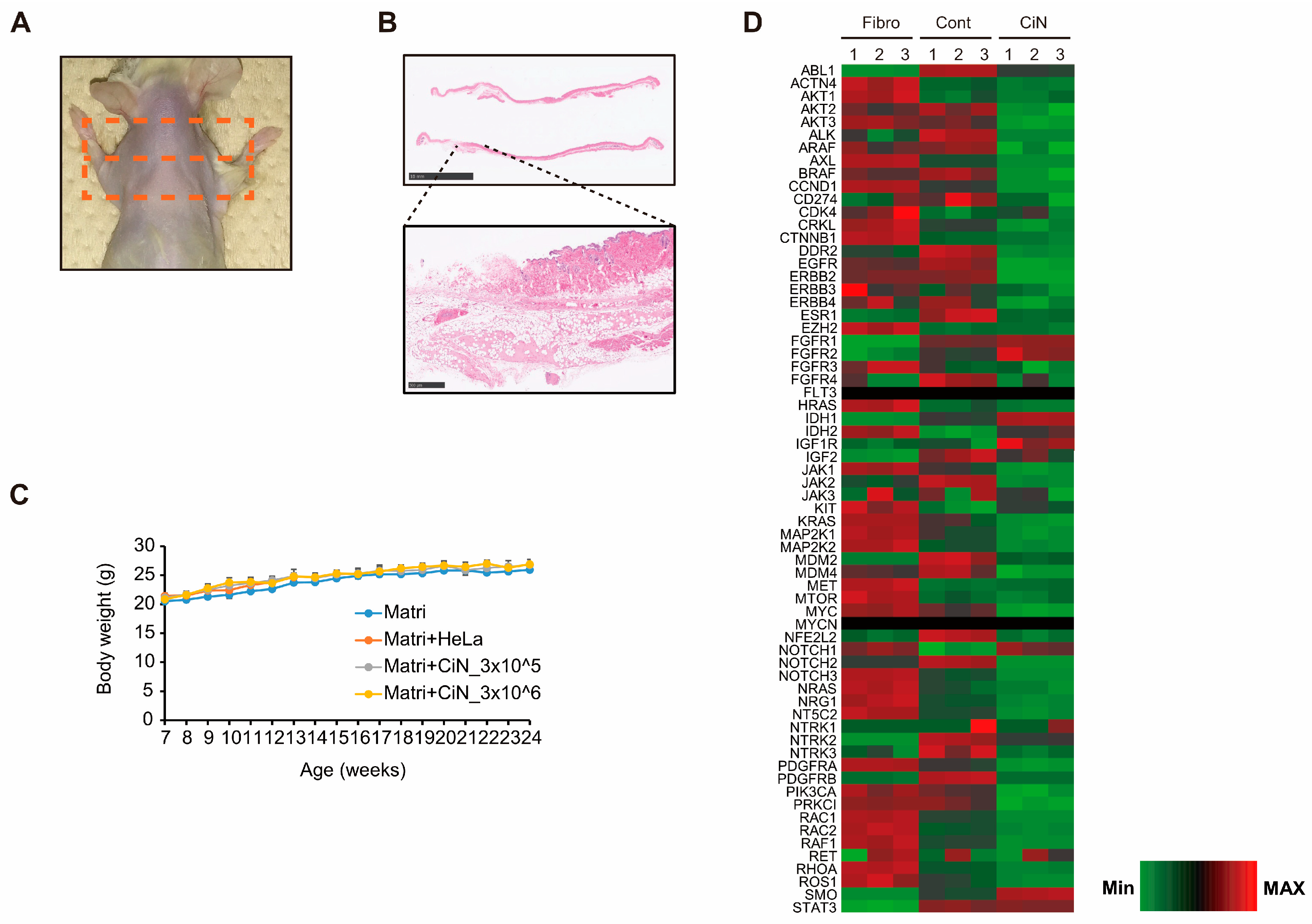
2.7. Transplantation of CiN Cells Did Not Induce Tumorigenesis
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Induction of Immature CiN Cells
4.2. Immunocytochemical Analysis
4.3. Animals
4.4. Contusive Spinal Cord Injury in Rats
4.5. Preparation of Cells and Transplantation
4.6. Basso, Beattie, and Bresnahan Test
4.7. CatWalk Analysis
4.8. Magnetic Resonance Imaging (MRI)
4.9. Histopathological Analysis
4.10. Immunohistochemical Analysis
4.11. RNA-Seq Analysis
4.12. Real-Time RT-PCR
4.13. Enzyme-Linked Immunosorbent Assay (ELISA)
4.14. Tumorigenicity Test
4.15. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ADC | Apparent diffusion coefficient |
| BBB test | Basso, Beattie, and Bresnahan test |
| Cont | Control |
| CiN | Chemical compound-induced neuronal-like cell |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| DTI | Diffusion tensor imaging |
| DWI | Diffusion-weighted image |
| EGF | Epidermal growth factor |
| FA | Fractional anisotropy |
| GFAP | Glial fibrillary acidic protein |
| HE | Hematoxylin and eosin |
| Iba1 | Ionized calcium binding adaptor molecule 1 |
| LFB | Luxol Fast Blue |
| MRI | Magnetic resonance imaging |
| NM | Neuronal medium |
| PBS | Phosphate buffered saline |
| RI | Regularity index |
| ROI | Region of interest |
| SCI | spinal cord injury |
| SEM | Standard error of the mean |
| SS | Swing speed |
| TubIII | βIII-Tubulin |
References
- Seif, M.; Gandini Wheeler-Kingshott, C.A.; Cohen-Adad, J.; Flanders, A.E.; Freund, P. Guidelines for the Conduct of Clinical Trials in Spinal Cord Injury: Neuroimaging Biomarkers. Spinal Cord 2019, 57, 717–728. [Google Scholar] [CrossRef] [PubMed]
- Harvey, L.A. Physiotherapy Rehabilitation for People with Spinal Cord Injuries. J. Physiother. 2016, 62, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Tso, D.; McKinnon, R.D. Cell Replacement Therapy for Central Nervous System Diseases. Neural Regen. Res. 2015, 10, 1356–1358. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Uezono, N.; Yasui, T.; Nakashima, K. Neural Stem Cell Therapy Aiming at Better Functional Recovery after Spinal Cord Injury. Dev. Dyn. 2018, 247, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Zhang, X.; Zhai, J.; Xue, J. The Roles and Applications of Neural Stem Cells in Spinal Cord Injury Repair. Front. Bioeng. Biotechnol. 2022, 10, 966866. [Google Scholar] [CrossRef]
- Gazdic, M.; Volarevic, V.; Randall Harrell, C.; Fellabaum, C.; Jovicic, N.; Arsenijevic, N.; Stojkovic, M. Stem Cells Therapy for Spinal Cord Injury. Int. J. Mol. Sci. 2018, 19, 1039. [Google Scholar] [CrossRef]
- Ramotowski, C.; Qu, X.; Villa-Diaz, L.G. Progress in the Use of Induced Pluripotent Stem Cell-Derived Neural Cells for Traumatic Spinal Cord Injuries in Animal Populations: Meta-Analysis and Review. Stem Cells Transl. Med. 2019, 8, 681–693. [Google Scholar] [CrossRef]
- Yoshihara, M.; Oguchi, A.; Murakawa, Y. Genomic Instability of IPSCs and Challenges in Their Clinical Applications. Adv. Exp. Med. Biol. 2019, 1201, 23–47. [Google Scholar] [CrossRef]
- Masuda, S.; Miyagawa, S.; Fukushima, S.; Sougawa, N.; Okimoto, K.; Tada, C.; Saito, A.; Sawa, Y. Eliminating Residual IPS Cells for Safety in Clinical Application. Protein Cell 2015, 6, 469–471. [Google Scholar] [CrossRef][Green Version]
- Horisawa, K.; Suzuki, A. Direct Cell-Fate Conversion of Somatic Cells: Toward Regenerative Medicine and Industries. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2020, 96, 131–158. [Google Scholar] [CrossRef]
- Ladewig, J.; Koch, P.; Brüstle, O. Leveling Waddington: The Emergence of Direct Programming and the Loss of Cell Fate Hierarchies. Nat. Rev. Mol. Cell Biol. 2013, 14, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.L.; Weintraub, H.; Lassar, A.B. Expression of a Single Transfected CDNA Converts Fibroblasts to Myoblasts. Cell 1987, 51, 987–1000. [Google Scholar] [CrossRef]
- Yoo, A.S.; Sun, A.X.; Li, L.; Shcheglovitov, A.; Portmann, T.; Li, Y.; Lee-Messer, C.; Dolmetsch, R.E.; Tsien, R.W.; Crabtree, G.R. MicroRNA-Mediated Conversion of Human Fibroblasts to Neurons. Nature 2011, 476, 228–231. [Google Scholar] [CrossRef] [PubMed]
- Ladewig, J.; Mertens, J.; Kesavan, J.; Doerr, J.; Poppe, D.; Glaue, F.; Herms, S.; Wernet, P.; Kögler, G.; Müller, F.-J.; et al. Small Molecules Enable Highly Efficient Neuronal Conversion of Human Fibroblasts. Nat. Methods 2012, 9, 575–578. [Google Scholar] [CrossRef]
- Huangfu, D.; Maehr, R.; Guo, W.; Eijkelenboom, A.; Snitow, M.; Chen, A.E.; Melton, D.A. Induction of Pluripotent Stem Cells by Defined Factors Is Greatly Improved by Small-Molecule Compounds. Nat. Biotechnol. 2008, 26, 795–797. [Google Scholar] [CrossRef]
- Xu, Y.; Shi, Y.; Ding, S. A Chemical Approach to Stem-Cell Biology and Regenerative Medicine. Nature 2008, 453, 338–344. [Google Scholar] [CrossRef]
- Pennarossa, G.; Maffei, S.; Campagnol, M.; Tarantini, L.; Gandolfi, F.; Brevini, T.A. Brief Demethylation Step Allows the Conversion of Adult Human Skin Fibroblasts into Insulin-Secreting Cells. Proc. Natl. Acad. Sci. USA 2013, 110, 8948–8953. [Google Scholar] [CrossRef]
- Takeda, Y.; Harada, Y.; Yoshikawa, T.; Dai, P. Chemical Compound-Based Direct Reprogramming for Future Clinical Applications. Biosci. Rep. 2018, 38, BSR20171650. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Hu, W.; Qiu, B.; Zhao, J.; Yu, Y.; Guan, W.; Wang, M.; Yang, W.; Pei, G. Generation of Neural Progenitor Cells by Chemical Cocktails and Hypoxia. Cell Res. 2014, 24, 665–679. [Google Scholar] [CrossRef]
- Hu, W.; Qiu, B.; Guan, W.; Wang, Q.; Wang, M.; Li, W.; Gao, L.; Shen, L.; Huang, Y.; Xie, G.; et al. Direct Conversion of Normal and Alzheimer’s Disease Human Fibroblasts into Neuronal Cells by Small Molecules. Cell Stem Cell 2015, 17, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zuo, X.; Jing, J.; Ma, Y.; Wang, J.; Liu, D.; Zhu, J.; Du, X.; Xiong, L.; Du, Y.; et al. Small-Molecule-Driven Direct Reprogramming of Mouse Fibroblasts into Functional Neurons. Cell Stem Cell 2015, 17, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Hou, P.; Li, Y.; Zhang, X.; Liu, C.; Guan, J.; Li, H.; Zhao, T.; Ye, J.; Yang, W.; Liu, K.; et al. Pluripotent Stem Cells Induced from Mouse Somatic Cells by Small-Molecule Compounds. Science 2013, 341, 651–654. [Google Scholar] [CrossRef] [PubMed]
- Dai, P.; Harada, Y.; Takamatsu, T. Highly Efficient Direct Conversion of Human Fibroblasts to Neuronal Cells by Chemical Compounds. J. Clin. Biochem. Nutr. 2015, 56, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, T.; Nagoshi, N.; Okano, H.; Nakamura, M. A Narrative Review of Advances in Neural Precursor Cell Transplantation Therapies for Spinal Cord Injury. Neurospine 2022, 19, 935–945. [Google Scholar] [CrossRef]
- Volarevic, V.; Erceg, S.; Bhattacharya, S.S.; Stojkovic, P.; Horner, P.; Stojkovic, M. Stem Cell-Based Therapy for Spinal Cord Injury. Cell Transplant. 2013, 22, 1309–1323. [Google Scholar] [CrossRef]
- Nagoshi, N.; Tsuji, O.; Nakamura, M.; Okano, H. Cell Therapy for Spinal Cord Injury Using Induced Pluripotent Stem Cells. Regen. Ther. 2019, 11, 75–80. [Google Scholar] [CrossRef]
- Fujiyoshi, K.; Konomi, T.; Yamada, M.; Hikishima, K.; Tsuji, O.; Komaki, Y.; Momoshima, S.; Toyama, Y.; Nakamura, M.; Okano, H. Diffusion Tensor Imaging and Tractography of the Spinal Cord: From Experimental Studies to Clinical Application. Exp. Neurol. 2013, 242, 74–82. [Google Scholar] [CrossRef]
- Chang, Y.; Jung, T.D.; Yoo, D.S.; Hyun, J.K. Diffusion Tensor Imaging and Fiber Tractography of Patients with Cervical Spinal Cord Injury. J. Neurotrauma 2010, 27, 2033–2040. [Google Scholar] [CrossRef]
- Ito, S.; Nagoshi, N.; Tsuji, O.; Shibata, S.; Shinozaki, M.; Kawabata, S.; Kojima, K.; Yasutake, K.; Hirokawa, T.; Matsumoto, M.; et al. LOTUS Inhibits Neuronal Apoptosis and Promotes Tract Regeneration in Contusive Spinal Cord Injury Model Mice. eNeuro 2018, 5, e0303-18.2018. [Google Scholar] [CrossRef]
- Hara, M.; Kobayakawa, K.; Ohkawa, Y.; Kumamaru, H.; Yokota, K.; Saito, T.; Kijima, K.; Yoshizaki, S.; Harimaya, K.; Nakashima, Y.; et al. Interaction of Reactive Astrocytes with Type i Collagen Induces Astrocytic Scar Formation through the Integrin-N-Cadherin Pathway after Spinal Cord Injury. Nat. Med. 2017, 23, 818–828. [Google Scholar] [CrossRef] [PubMed]
- Nagoshi, N.; Tsuji, O.; Kitamura, K.; Suda, K.; Maeda, T.; Yato, Y.; Abe, T.; Hayata, D.; Matsumoto, M.; Okano, H.; et al. Phase I/II Study of Intrathecal Administration of Recombinant Human Hepatocyte Growth Factor in Patients with Acute Spinal Cord Injury: A Double-Blind, Randomized Clinical Trial of Safety and Efficacy. J. Neurotrauma 2020, 37, 1752–1758. [Google Scholar] [CrossRef]
- Kitamura, K.; Fujiyoshi, K.; Yamane, J.-I.; Toyota, F.; Hikishima, K.; Nomura, T.; Funakoshi, H.; Nakamura, T.; Aoki, M.; Toyama, Y.; et al. Human Hepatocyte Growth Factor Promotes Functional Recovery in Primates after Spinal Cord Injury. PLoS ONE 2011, 6, e27706. [Google Scholar] [CrossRef]
- Kitamura, K.; Iwanami, A.; Nakamura, M.; Yamane, J.; Watanabe, K.; Suzuki, Y.; Miyazawa, D.; Shibata, S.; Funakoshi, H.; Miyatake, S.; et al. Hepatocyte Growth Factor Promotes Endogenous Repair and Functional Recovery after Spinal Cord Injury. J. Neurosci. Res. 2007, 85, 2332–2342. [Google Scholar] [CrossRef]
- Fischer, I.; Dulin, J.N.; Lane, M.A. Transplanting Neural Progenitor Cells to Restore Connectivity after Spinal Cord Injury. Nat. Rev. Neurosci. 2020, 21, 366–383. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Iwata, M.; Hamano, M.; Eguchi, R.; Takeshita, J.I.; Yamanishi, Y. Small Compound-Based Direct Cell Conversion with Combinatorial Optimization of Pathway Regulations. Bioinformatics 2022, 38, ii99–ii105. [Google Scholar] [CrossRef] [PubMed]
- Guan, J.; Wang, G.; Wang, J.; Zhang, Z.; Fu, Y.; Cheng, L.; Meng, G.; Lyu, Y.; Zhu, J.; Li, Y.; et al. Chemical Reprogramming of Human Somatic Cells to Pluripotent Stem Cells. Nature 2022, 605, 325–331. [Google Scholar] [CrossRef]
- Liuyang, S.; Wang, G.; Wang, Y.; He, H.; Lyu, Y.; Cheng, L.; Yang, Z.; Guan, J.; Fu, Y.; Zhu, J.; et al. Highly Efficient and Rapid Generation of Human Pluripotent Stem Cells by Chemical Reprogramming. Cell Stem Cell 2023, 30, 450–459. [Google Scholar] [CrossRef]
- Kitamura, K.; Iwanami, A.; Iwai, H.; Toyama, Y. Therapeutic Time Window and Preclinical Efficacy of Intrathecal Administration of Recombinant Human Hepatocyte Growth Factor for Acute Spinal Cord Injury. J. Spine Res. 2016, 7, 934–939. [Google Scholar]
- Yates, A.G.; Jogia, T.; Gillespie, E.R.; Couch, Y.; Ruitenberg, M.J.; Anthony, D.C. Acute IL-1RA Treatment Suppresses the Peripheral and Central Inflammatory Response to Spinal Cord Injury. J. Neuroinflamm. 2021, 18, 15. [Google Scholar] [CrossRef]
- Takeuchi, H.; Natsume, A.; Wakabayashi, T.; Aoshima, C.; Shimato, S.; Ito, M.; Ishii, J.; Maeda, Y.; Hara, M.; Kim, S.U.; et al. Intravenously Transplanted Human Neural Stem Cells Migrate to the Injured Spinal Cord in Adult Mice in an SDF-1- and HGF-Dependent Manner. Neurosci. Lett. 2007, 426, 69–74. [Google Scholar] [CrossRef]
- Coulson-Thomas, V.J.; Lauer, M.E.; Soleman, S.; Zhao, C.; Hascall, V.C.; Day, A.J.; Fawcett, J.W. Tumor Necrosis Factor-Stimulated Gene-6 (TSG-6) Is Constitutively Expressed in Adult Central Nervous System (CNS) and Associated with Astrocyte-Mediated Glial Scar Formation Following Spinal Cord Injury. J. Biol. Chem. 2016, 291, 19939–19952. [Google Scholar] [CrossRef] [PubMed]
- Buss, A.; Pech, K.; Kakulas, B.A.; Martin, D.; Schoenen, J.; Noth, J.; Brook, G.A. TGF-Β1 and TGF-Β2 Expression after Traumatic Human Spinal Cord Injury. Spinal Cord 2008, 46, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Rust, R.; Kaiser, J. Insights into the Dual Role of Inflammation after Spinal Cord Injury. J. Neurosci. 2017, 37, 4658–4660. [Google Scholar] [CrossRef]
- Hegarty, S.V.; O’Keeffe, G.W.; Sullivan, A.M. BMP-Smad 1/5/8 Signalling in the Development of the Nervous System. Prog. Neurobiol. 2013, 109, 28–41. [Google Scholar] [CrossRef] [PubMed]
- Mehler, M.F.; Mabie, P.C.; Zhang, D.; Kessler, J.A. Bone Morphogenetic Proteins in the Nervous System. Trends Neurosci. 1997, 20, 309–317. [Google Scholar] [CrossRef]
- Jeong, S.R.; Kwon, M.J.; Lee, H.G.; Joe, E.H.; Lee, J.H.; Kim, S.S.; Suh-Kim, H.; Kim, B.G. Hepatocyte Growth Factor Reduces Astrocytic Scar Formation and Promotes Axonal Growth beyond Glial Scars after Spinal Cord Injury. Exp. Neurol. 2012, 233, 312–322. [Google Scholar] [CrossRef]
- Huang, C.; Tu, W.; Fu, Y.; Wang, J.; Xie, X. Chemical-Induced Cardiac Reprogramming in Vivo. Cell Res. 2018, 28, 686–689. [Google Scholar] [CrossRef]
- Desole, C.; Gallo, S.; Vitacolonna, A.; Montarolo, F.; Bertolotto, A.; Vivien, D.; Comoglio, P.; Crepaldi, T. HGF and MET: From Brain Development to Neurological Disorders. Front. Cell Dev. Biol. 2021, 9, 683609. [Google Scholar] [CrossRef]
- Yang, B.; Zhang, F.; Cheng, F.; Ying, L.; Wang, C.; Shi, K.; Wang, J.; Xia, K.; Gong, Z.; Huang, X.; et al. Strategies and Prospects of Effective Neural Circuits Reconstruction after Spinal Cord Injury. Cell Death Dis. 2020, 11, 439. [Google Scholar] [CrossRef]
- Basso, D.M.; Beattie, M.S.; Bresnahan, J.C. A Sensitive and Reliable Locomotor Rating Scale for Open Field Testing in Rats. J. Neurotrauma 1995, 12, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Hamers, F.P.T.; Lankhorst, A.J.; Van Laar, T.J.; Veldhuis, W.B.; Gispen, W.H. Automated Quantitative Gait Analysis during Overground Locomotion in the Rat: Its Application to Spinal Cord Contusion and Transection Injuries. J. Neurotrauma 2001, 18, 187–201. [Google Scholar] [CrossRef] [PubMed]
- Konomi, T.; Fujiyoshi, K.; Hikishima, K.; Komaki, Y.; Tsuji, O.; Okano, H.J.; Toyama, Y.; Okano, H.; Nakamura, M. Conditions for Quantitative Evaluation of Injured Spinal Cord by in Vivo Diffusion Tensor Imaging and Tractography: Preclinical Longitudinal Study in Common Marmosets. Neuroimage 2012, 63, 1841–1853. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Benner, T. Diffusion Toolkit: A Software Package for Diffusion Imaging Data Processing and Tractography. Proc. Intl. Soc. Mag. Reson. Med. 2007, 15, 3720. [Google Scholar]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurahashi, T.; Nishime, C.; Nishinaka, E.; Komaki, Y.; Seki, F.; Urano, K.; Harada, Y.; Yoshikawa, T.; Dai, P. Transplantation of Chemical Compound-Induced Cells from Human Fibroblasts Improves Locomotor Recovery in a Spinal Cord Injury Rat Model. Int. J. Mol. Sci. 2023, 24, 13853. https://doi.org/10.3390/ijms241813853
Kurahashi T, Nishime C, Nishinaka E, Komaki Y, Seki F, Urano K, Harada Y, Yoshikawa T, Dai P. Transplantation of Chemical Compound-Induced Cells from Human Fibroblasts Improves Locomotor Recovery in a Spinal Cord Injury Rat Model. International Journal of Molecular Sciences. 2023; 24(18):13853. https://doi.org/10.3390/ijms241813853
Chicago/Turabian StyleKurahashi, Toshihiro, Chiyoko Nishime, Eiko Nishinaka, Yuji Komaki, Fumiko Seki, Koji Urano, Yoshinori Harada, Toshikazu Yoshikawa, and Ping Dai. 2023. "Transplantation of Chemical Compound-Induced Cells from Human Fibroblasts Improves Locomotor Recovery in a Spinal Cord Injury Rat Model" International Journal of Molecular Sciences 24, no. 18: 13853. https://doi.org/10.3390/ijms241813853
APA StyleKurahashi, T., Nishime, C., Nishinaka, E., Komaki, Y., Seki, F., Urano, K., Harada, Y., Yoshikawa, T., & Dai, P. (2023). Transplantation of Chemical Compound-Induced Cells from Human Fibroblasts Improves Locomotor Recovery in a Spinal Cord Injury Rat Model. International Journal of Molecular Sciences, 24(18), 13853. https://doi.org/10.3390/ijms241813853