

A Novel Perspective on Neuronal Control of Anatomical Patterning, Remodeling, and Maintenance

Abstract

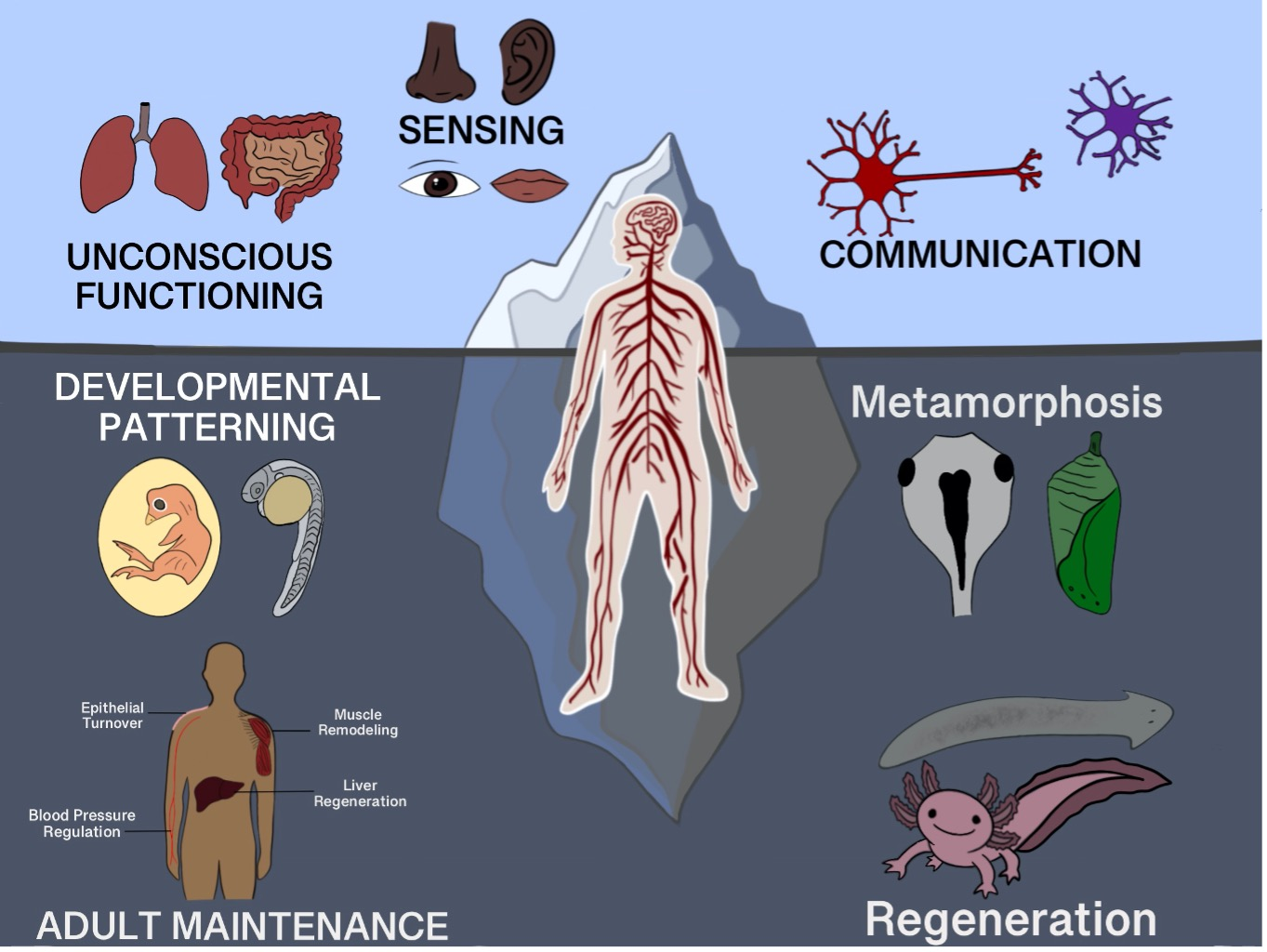{kind=link}
1. Introduction
2. How Do We Build an Organism? From Embryo to Juvenile to Adult Body Plans
2.1. Neural Ectoderm in Development
2.2. The CNS as a Regulator of Craniofacial and Anatomical Patterning
2.3. The Peripheral Nervous System’s Role in Developmental Patterning
3. How Do We Maintain Proper Patterning of the Adult Body Plan?
3.1. Neuronal Control of Organ System Function, Patterning, and Maintenance
3.2. Transformation toward an Adult Patterning
4. Resolution of Abnormal Body Plans
4.1. Regeneration
4.2. Non-Metamorphic Remodeling
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhat, N.; Kwon, H.-J.; Riley, B.B. A gene network that coordinates preplacodal competence and neural crest specification in zebrafish. Dev. Biol. 2013, 373, 107–117. [Google Scholar] [CrossRef]
- Grocott, T.; Tambalo, M.; Streit, A. The peripheral sensory nervous system in the vertebrate head: A gene regulatory perspective. Dev. Biol. 2012, 370, 3–23. [Google Scholar] [CrossRef]
- Patthey, C.; Gunhaga, L. Specification and regionalisation of the neural plate border. Eur. J. Neurosci. 2011, 34, 1516–1528. [Google Scholar] [CrossRef]
- Betters, E.; Liu, Y.; Kjaeldgaard, A.; Sundström, E.; García-Castro, M.I. Analysis of early human neural crest development. Dev. Biol. 2010, 344, 578–592. [Google Scholar] [CrossRef]
- Hernandez, M.R. The optic nerve head in glaucoma: Role of astrocytes in tissue remodeling. Prog. Retin. Eye Res. 2000, 19, 297–321. [Google Scholar] [CrossRef]
- Micera, A.; Lambiase, A.; Stampachiacchiere, B.; Bonini, S.; Bonini, S.; Levischaffer, F. Nerve growth factor and tissue repair remodeling: trkANGFR and p75NTR, two receptors one fate. Cytokine Growth Factor Rev. 2007, 18, 245–256. [Google Scholar] [CrossRef]
- Stocum, D.L. The role of peripheral nerves in urodele limb regeneration. Eur. J. Neurosci. 2011, 34, 908–916. [Google Scholar] [CrossRef]
- Adameyko, I.; Fried, K. The Nervous System Orchestrates and Integrates Craniofacial Development: A Review. Front. Physiol. 2016, 7, 49. [Google Scholar] [CrossRef]
- Fukai, T.; Takeda, A.; Uchinuma, E. Wound healing in denervated rat skin. Wound Repair Regen. 2005, 13, 175–180. [Google Scholar] [CrossRef]
- Mahmoud, A.I.; O’Meara, C.C.; Gemberling, M.; Zhao, L.; Bryant, D.M.; Zheng, R.; Gannon, J.B.; Cai, L.; Choi, W.-Y.; Egnaczyk, G.F.; et al. Nerves Regulate Cardiomyocyte Proliferation and Heart Regeneration. Dev. Cell 2015, 34, 387–399. [Google Scholar] [CrossRef]
- Copp, A.J.; Greene, N.D.E. Genetics and development of neural tube defects. J. Pathol. 2010, 220, 217–230. [Google Scholar] [CrossRef]
- Copp, A.J.; Greene, N.D.E.; Murdoch, J.N. The genetic basis of mammalian neurulation. Nat. Rev. Genet. 2003, 4, 784–793. [Google Scholar] [CrossRef]
- Copp, A.J.; Greene, N.D.E. Neural tube defects—Disorders of neurulation and related embryonic processes. WIREs Dev. Biol. 2013, 2, 213–227. [Google Scholar] [CrossRef]
- Engelhardt, D.M.; Martyr, C.A.; Niswander, L. Pathogenesis of neural tube defects: The regulation and disruption of cellular processes underlying neural tube closure. WIREs Mech. Dis. 2022, 14, e1559. [Google Scholar] [CrossRef]
- Greene, N.D.E.; Copp, A.J. Neural tube defects. Annu. Rev. Neurosci. 2014, 37, 221–242. [Google Scholar] [CrossRef]
- Harris, M.J.; Juriloff, D.M. Mouse mutants with neural tube closure defects and their role in understanding human neural tube defects. Birth Defects Res. Part A Clin. Mol. Teratol. 2007, 79, 187–210. [Google Scholar] [CrossRef]
- Lee, S.; Gleeson, J.G. Closing in on Mechanisms of Open Neural Tube Defects. Trends Neurosci. 2020, 43, 519–532. [Google Scholar] [CrossRef]
- Koontz, A.; Urrutia, H.A.; Bronner, M.E. Making a head: Neural crest and ectodermal placodes in cranial sensory development. Semin. Cell Dev. Biol. 2023, 138, 15–27. [Google Scholar] [CrossRef]
- Le Douarin, N.; Kalcheim, C. The Neural Crest, 2nd ed.; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- de Vellis, J.; Carpenter, E. General Development of the Nervous System. In Basic Neurochemistry: Molecular, Cellular and Medical Aspects, 6th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 1999. [Google Scholar]
- Weston, J.A. The Migration and Differentiation of Neural Crest Cells. In Advances in Morphogenesis; Abercrombie, M., Brachet, J., King, T.J., Eds.; Elsevier: Amsterdam, The Netherlands, 1970; Volume 8, pp. 41–114. [Google Scholar]
- Bronner, M.E.; Simões-Costa, M. The neural crest migrating into the 21st century. Curr. Top. Dev. Biol. 2016, 116, 115–134. [Google Scholar] [CrossRef]
- Bronner-Fraser, M.; Fraser, S.E. Cell lineage analysis reveals multipotency of some avian neural crest cells. Nature 1988, 335, 161–164. [Google Scholar] [CrossRef]
- Crane, J.F.; Trainor, P.A. Neural crest stem and progenitor cells. Annu. Rev. Cell Dev. Biol. 2006, 22, 267–286. [Google Scholar] [CrossRef] [PubMed]
- Hall, B.K. The Neural Crest and Neural Crest Cells in Vertebrate Development and Evolution, 2nd ed.; Springer Series in Solid-State Sciences Electron Transport in Compound Semiconductors; Springer: New York, NY, USA, 2009. [Google Scholar]
- Piacentino, M.L.; Li, Y.; Bronner, M.E. Epithelial-to-mesenchymal transition and different migration strategies as viewed from the neural crest. Curr. Opin. Cell Biol. 2020, 66, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Bronner, M.E. Neural crest lineage analysis: From past to future trajectory. Development 2020, 147, dev193193. [Google Scholar] [CrossRef] [PubMed]
- Harrow-Mortelliti, M.; Reddy, V.; Jimsheleishvili, G. Physiology, spinal cord. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Herrera-Rincon, C.; Pai, V.P.; Moran, K.M.; Lemire, J.M.; Levin, M. The brain is required for normal muscle and nerve patterning during early Xenopus development. Nat. Commun. 2017, 8, 587. [Google Scholar] [CrossRef] [PubMed]
- Popiela, H. In vivo limb tissue development in the absence of nerves: A quantitative study. Exp. Neurol. 1976, 53, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Anchan, R.M.; Drake, D.P.; Haines, C.F.; Gerwe, E.A.; LaMantia, A.S. Disruption of local retinoid-mediated gene expression accompanies abnormal development in the mammalian olfactory pathway. J. Comp. Neurol. 1997, 379, 171–184. [Google Scholar] [CrossRef]
- LaMantia, A.S.; Colbert, M.C.; Linney, E. Retinoic acid induction and regional differentiation prefigure olfactory pathway formation in the mammalian forebrain. Neuron 1993, 10, 1035–1048. [Google Scholar] [CrossRef]
- LaMantia, A.-S. Why Does the Face Predict the Brain? Neural Crest Induction, Craniofacial Morphogenesis, and Neural Circuit Development. Front. Physiol. 2020, 11, 610970. [Google Scholar] [CrossRef]
- McGonnell, I.M.; McKay, I.J.; Graham, A. A Population of Caudally Migrating Cranial Neural Crest Cells: Functional and Evolutionary Implications. Dev. Biol. 2001, 236, 354–363. [Google Scholar] [CrossRef]
- Francis-West, P.; Ladher, R.; Barlow, A.; Graveson, A. Signalling interactions during facial development. Mech. Dev. 1998, 75, 3–28. [Google Scholar] [CrossRef]
- Lwigale, P.Y.; Conrad, G.W.; Bronner-Fraser, M. Graded potential of neural crest to form cornea, sensory neurons and cartilage along the rostrocaudal axis. Development 2004, 131, 1979–1991. [Google Scholar] [CrossRef] [PubMed]
- Serbedzija, G.N.; Bronner-Fraser, M.; Fraser, S.E. Vital dye analysis of cranial neural crest cell migration in the mouse embryo. Development 1992, 116, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Theveneau, E.; Mayor, R. Neural crest delamination and migration: From epithelium-to-mesenchyme transition to collective cell migration. Dev. Biol. 2012, 366, 34–54. [Google Scholar] [CrossRef]
- Aguiar, D.P.; Sghari, S.; Creuzet, S. The facial neural crest controls fore- and midbrain patterning by regulating Foxg1 expression through Smad1 activity. Development 2014, 141, 2494–2505. [Google Scholar] [CrossRef] [PubMed]
- Creuzet, S.E. Regulation of pre-otic brain development by the cephalic neural crest. Proc. Natl. Acad. Sci. USA 2009, 106, 15774–15779. [Google Scholar] [CrossRef]
- Creuzet, S.E.; Martinez, S.; Le Douarin, N.M. The cephalic neural crest exerts a critical effect on forebrain and midbrain development. Proc. Natl. Acad. Sci. USA 2006, 103, 14033–14038. [Google Scholar] [CrossRef]
- Marcucio, R.S.; Young, N.M.; Hu, D.; Hallgrimsson, B. Mechanisms that Underlie Co-variation of the Brain and Face. Genesis 2011, 49, 177–189. [Google Scholar] [CrossRef]
- Reid, S.N.; Ziermann, J.M.; Gondré-Lewis, M.C. Genetically induced abnormal cranial development in human trisomy 18 with holoprosencephaly: Comparisons with the normal tempo of osteogenic–neural development. J. Anat. 2015, 227, 21–33. [Google Scholar] [CrossRef]
- Richtsmeier, J.T.; Flaherty, K. Hand in glove: Brain and skull in development and dysmorphogenesis. Acta Neuropathol. 2013, 125, 469–489. [Google Scholar] [CrossRef]
- Nieman, B.J.; Blank, M.C.; Roman, B.B.; Henkelman, R.M.; Millen, K.J. If the skull fits: Magnetic resonance imaging and microcomputed tomography for combined analysis of brain and skull phenotypes in the mouse. Physiol. Genom. 2012, 44, 992–1002. [Google Scholar] [CrossRef]
- Olson, E.C.; Miller, R.L. Morphological Integration; University of Chicago Press: Chicago, IL, USA, 1999; 380p. [Google Scholar]
- Richtsmeier, J.T.; Aldridge, K.; DeLeon, V.B.; Panchal, J.; Kane, A.A.; Marsh, J.L.; Yan, P.; Cole, T.M. Phenotypic Integration of Neurocranium and Brain. J. Exp. Zool. B Mol. Dev. Evol. 2006, 306, 360–378. [Google Scholar] [CrossRef] [PubMed]
- Rightmire, G.P. Homo erectus and Middle Pleistocene hominins: Brain size, skull form, and species recognition. J. Hum. Evol. 2013, 65, 223–252. [Google Scholar] [CrossRef] [PubMed]
- Motch Perrine, S.M.; Stecko, T.; Neuberger, T.; Jabs, E.W.; Ryan, T.M.; Richtsmeier, J.T. Integration of Brain and Skull in Prenatal Mouse Models of Apert and Crouzon Syndromes. Front. Hum. Neurosci. 2017, 11, 369. [Google Scholar] [CrossRef]
- Knight, S.J.; Anderson, V.A.; Spencer-Smith, M.M.; Da Costa, A.C. Neurodevelopmental Outcomes in Infants and Children with Single-Suture Craniosynostosis: A Systematic Review. Dev. Neuropsychol. 2014, 39, 159–186. [Google Scholar] [CrossRef] [PubMed]
- Starr, J.R.; Collett, B.R.; Gaither, R.; Kapp-Simon, K.A.; Cradock, M.M.; Cunningham, M.L.; Speltz, M.L. Multicenter Study of Neurodevelopment in 3-Year-Old Children with and Without Single-Suture Craniosynostosis. Arch. Pediatr. Adolesc. Med. 2012, 166, 536–542. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Ma, L.; Yuan, Y.; Ye, X.; Montagne, A.; He, J.; Ho, T.-V.; Wu, Y.; Zhao, Z.; Sta Maria, N.; et al. Cranial Suture Regeneration Mitigates Skull and Neurocognitive Defects in Craniosynostosis. Cell 2021, 184, 243–256.e218. [Google Scholar] [CrossRef] [PubMed]
- Azoury, S.C.; Reddy, S.; Shukla, V.; Deng, C.-X. Fibroblast Growth Factor Receptor 2 (FGFR2) Mutation Related Syndromic Craniosynostosis. Int. J. Biol. Sci. 2017, 13, 1479–1488. [Google Scholar] [CrossRef]
- Lee, K.K.L.; Peskett, E.; Quinn, C.M.; Aiello, R.; Adeeva, L.; Moulding, D.A.; Stanier, P.; Pauws, E. Overexpression of Fgfr2c causes craniofacial bone hypoplasia and ameliorates craniosynostosis in the Crouzon mouse. Dis. Model. Mech. 2018, 11, dmm035311. [Google Scholar] [CrossRef]
- Martínez-Abadías, N.; Percival, C.; Aldridge, K.; Hill, C.A.; Ryan, T.; Sirivunnabood, S.; Wang, Y.; Jabs, E.W.; Richtsmeier, J.T. Beyond the closed suture in Apert syndrome mouse models: Evidence of primary effects of FGFR2 signaling on facial shape at birth. Dev. Dyn. 2010, 239, 3058–3071. [Google Scholar] [CrossRef]
- Motch Perrine, S.M.; Cole, T.M.; Martínez-Abadías, N.; Aldridge, K.; Jabs, E.W.; Richtsmeier, J.T. Craniofacial divergence by distinct prenatal growth patterns in Fgfr2 mutant mice. BMC Dev. Biol. 2014, 14, 8. [Google Scholar] [CrossRef]
- Pandey, R.K.; Bajpai, M.; Ali, A.; Gayan, S.; Singh, A. Mutational identification of fibroblast growth factor receptor 1 and fibroblast growth factor receptor 2 genes in craniosynostosis in Indian population. Indian J. Hum. Genet. 2013, 19, 449–453. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Posnick, J.C.; Armstrong, D.; Bite, U. Crouzon and Apert syndromes: Intracranial volume measurements before and after cranio-orbital reshaping in childhood. Plast. Reconstr. Surg. 1995, 96, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, S.; Li, J.; Yi, Z. The role of the fibroblast growth factor family in bone-related diseases. Chem. Biol. Drug Des. 2019, 94, 1740–1749. [Google Scholar] [CrossRef] [PubMed]
- Shaw, W. Global Strategies to Reduce the Health Care Burden of Craniofacial Anomalies: Report of WHO Meetings on International Collaborative Research on Craniofacial Anomalies. Cleft Palate-Craniofacial J. 2004, 41, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Folic Acid and Neural Tube Defects: Data & Statistics; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2018.
- Hill, C.; Jacobs, B.; Kennedy, L.; Rohde, S.; Zhou, B.; Baldwin, S.; Goudy, S. Cranial neural crest deletion of VEGFa causes cleft palate with aberrant vascular and bone development. Cell Tissue Res. 2015, 361, 711–722. [Google Scholar] [CrossRef]
- Jones, N.C.; Lynn, M.L.; Gaudenz, K.; Sakai, D.; Aoto, K.; Rey, J.-P.; Glynn, E.F.; Ellington, L.; Du, C.; Dixon, J.; et al. Prevention of the neurocristopathy Treacher Collins syndrome through inhibition of p53 function. Nat. Med. 2008, 14, 125–133. [Google Scholar] [CrossRef]
- Chong, H.J.; Young, N.M.; Hu, D.; Jeong, J.; McMahon, A.P.; Hallgrimsson, B.; Marcucio, R.S. Signaling by SHH rescues facial defects following blockade in the brain. Dev. Dyn. 2012, 241, 247–256. [Google Scholar] [CrossRef]
- Hu, D.; Marcucio, R.S. A SHH-responsive signaling center in the forebrain regulates craniofacial morphogenesis via the facial ectoderm. Development 2009, 136, 107–116. [Google Scholar] [CrossRef]
- Hu, D.; Young, N.M.; Xu, Q.; Jamniczky, H.; Green, R.M.; Mio, W.; Marcucio, R.S.; Hallgrimsson, B. Signals from the brain induce variation in avian facial shape. Dev. Dyn. 2015, 244, 1133–1143. [Google Scholar] [CrossRef]
- Corbisier de Meautsart, C.; Dyson, R.M.; Latter, J.L.; Berry, M.J.; Clifton, V.L.; Wright, I.M.R. Influence of sympathetic activity in the control of peripheral microvascular tone in preterm infants. Pediatr. Res. 2016, 80, 793–799. [Google Scholar] [CrossRef]
- Haraldsdottir, K.; Watson, A.M.; Beshish, A.G.; Pegelow, D.F.; Palta, M.; Tetri, L.H.; Brix, M.D.; Centanni, R.M.; Goss, K.N.; Eldridge, M.W. Heart rate recovery after maximal exercise is impaired in healthy young adults born preterm. Eur. J. Appl. Physiol. 2019, 119, 857–866. [Google Scholar] [CrossRef]
- Nino, G.; Govindan, R.B.; Al-Shargabi, T.; Metzler, M.; Massaro, A.N.; Perez, G.F.; McCarter, R.; Hunt, C.E.; du Plessis, A.J. Premature Infants Rehospitalized because of an Apparent Life-Threatening Event Had Distinctive Autonomic Developmental Trajectories. Am. J. Respir. Crit. Care Med. 2016, 194, 379–381. [Google Scholar] [CrossRef]
- Segar, J.L. Ontogeny of the arterial and cardiopulmonary baroreflex during fetal and postnatal life. Am. J. Physiol. 1997, 273, R457–R471. [Google Scholar] [CrossRef] [PubMed]
- Bain, J.R.; DeMatteo, C.; Gjertsen, D.; Packham, T.; Galea, V.; Harper, J.A. Limb Length Differences after Obstetrical Brachial Plexus Injury: A Growing Concern. Plast. Reconstr. Surg. 2012, 130, 558e–571e. [Google Scholar] [CrossRef]
- Mukouyama, Y.-S.; Shin, D.; Britsch, S.; Taniguchi, M.; Anderson, D.J. Sensory Nerves Determine the Pattern of Arterial Differentiation and Blood Vessel Branching in the Skin. Cell 2002, 109, 693–705. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, R.E.; Li, Z.; Zhang, Q.; Goh, B.C.; Li, Z.; Thorek, D.L.J.; Rajbhandari, L.; Brushart, T.M.; Minichiello, L.; Zhou, F.; et al. NGF-TrkA Signaling by Sensory Nerves Coordinates the Vascularization and Ossification of Developing Endochondral Bone. Cell Rep. 2016, 16, 2723–2735. [Google Scholar] [CrossRef] [PubMed]
- Knox, S.M.; Lombaert, I.M.A.; Reed, X.; Vitale-Cross, L.; Gutkind, J.S.; Hoffman, M.P. Parasympathetic innervation maintains epithelial progenitor cells during salivary organogenesis. Science 2010, 329, 1645–1647. [Google Scholar] [CrossRef]
- Hanzlíková, V.; Schiaffino, S. Studies on the effect of denervation in developing muscle. Z. Für Zellforsch. Mikrosk. Anat. 1973, 147, 75–85. [Google Scholar] [CrossRef]
- Hoffman, W.Y.M.D.; McCarthy, J.G.M.D. The Effects of Facial Nerve Ablation on Craniofacial Skeletal Development in Neonatal Rabbits. Plast. Reconstr. Surg. 1994, 93, 1236–1240. [Google Scholar] [CrossRef]
- Sinsel, N.K.; Opdebeeck, H.; Guelinckx, P.J. The Effect of Unilateral Partial Facial Paralysis and Muscle Ablation on Craniofacial Growth and Development: An Experimental Study in the Rabbit. Plast. Reconstr. Surg. 1998, 102, 1894–1912. [Google Scholar] [CrossRef]
- Cotrina, M.L.; González-Hoyuela, M.; Barbas, J.A.; Rodríguez-Tébar, A. Programmed Cell Death in the Developing Somites Is Promoted by Nerve Growth Factor via Its p75NTR Receptor. Dev. Biol. 2000, 228, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Hamburger, V.; Waugh, M. The Primary Development of the Skeleton in Nerveless and Poorly Innervated Limb Transplants of Chick Embryos. Physiol. Zool. 1940, 13, 367–382. [Google Scholar] [CrossRef]
- Brill, G.; Kahane, N.; Carmeli, C.; von Schack, D.; Barde, Y.A.; Kalcheim, C. Epithelial-mesenchymal conversion of dermatome progenitors requires neural tube-derived signals: Characterization of the role of Neurotrophin-3. Development 1995, 121, 2583–2594. [Google Scholar] [CrossRef]
- Larson, T.A.; Gordon, T.N.; Lau, H.E.; Parichy, D.M. Defective adult oligodendrocyte and Schwann cell development, pigment pattern, and craniofacial morphology in puma mutant zebrafish having an alpha tubulin mutation. Dev. Biol. 2010, 346, 296–309. [Google Scholar] [CrossRef]
- Blackiston, D.; Adams, D.S.; Lemire, J.M.; Lobikin, M.; Levin, M. Transmembrane potential of GlyCl-expressing instructor cells induces a neoplastic-like conversion of melanocytes via a serotonergic pathway. Dis. Models Mech. 2011, 4, 67–85. [Google Scholar] [CrossRef] [PubMed]
- Lobikin, M.; Lobo, D.; Blackiston, D.J.; Martyniuk, C.J.; Tkachenko, E.; Levin, M. Serotonergic regulation of melanocyte conversion: A bioelectrically regulated network for stochastic all-or-none hyperpigmentation. Sci. Signal. 2015, 8, ra99. [Google Scholar] [CrossRef] [PubMed]
- Lobikin, M.; Paré, J.-F.; Kaplan, D.L.; Levin, M. Selective depolarization of transmembrane potential alters muscle patterning and muscle cell localization in Xenopus laevis embryos. Int. J. Dev. Biol. 2015, 59, 303–311. [Google Scholar] [CrossRef]
- Sullivan, K.G.; Levin, M. Neurotransmitter signaling pathways required for normal development in Xenopus laevis embryos: A pharmacological survey screen. J. Anat. 2016, 229, 483–502. [Google Scholar] [CrossRef]
- Levin, M.; Buznikov, G.A.; Lauder, J.M. Of Minds and Embryos: Left-Right Asymmetry and the Serotonergic Controls of Pre-Neural Morphogenesis. Dev. Neurosci. 2006, 28, 171–185. [Google Scholar] [CrossRef]
- Melnikov, N.P.; Bolshakov, F.V.; Frolova, V.S.; Skorentseva, K.V.; Ereskovsky, A.V.; Saidova, A.A.; Lavrov, A.I. Tissue homeostasis in sponges: Quantitative analysis of cell proliferation and apoptosis. J. Exp. Zool. Part B Mol. Dev. Evol. 2022, 338, 360–381. [Google Scholar] [CrossRef]
- Liu, D.; Kou, X.; Chen, C.; Liu, S.; Liu, Y.; Yu, W.; Yu, T.; Yang, R.; Wang, R.; Zhou, Y.; et al. Circulating apoptotic bodies maintain mesenchymal stem cell homeostasis and ameliorate osteopenia via transferring multiple cellular factors. Cell Res. 2018, 28, 918–933. [Google Scholar] [CrossRef]
- Rattan, S.I.S. Synthesis, modification and turnover of proteins during aging. In Protein Metabolism and Homeostasis in Aging; Tavernarakis, N., Ed.; Springer: Boston, MA, USA, 2010; pp. 1–13. [Google Scholar]
- Van Keymeulen, A.; Blanpain, C. Tracing epithelial stem cells during development, homeostasis, and repair. J. Cell Biol. 2012, 197, 575–584. [Google Scholar] [CrossRef]
- Krndija, D.; El Marjou, F.; Guirao, B.; Richon, S.; Leroy, O.; Bellaiche, Y.; Hannezo, E.; Matic Vignjevic, D. Active cell migration is critical for steady-state epithelial turnover in the gut. Science 2019, 365, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Leblond, C.P.; Stevens, C.E. The constant renewal of the intestinal epithelium in the albino rat. Anat. Rec. 1948, 100, 357–377. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Balachandra, S.; Ngo, S.; O’Brien, L.E. Feedback regulation of steady-state epithelial turnover and organ size. Nature 2017, 548, 588–591. [Google Scholar] [CrossRef]
- Peters, E.M.J.; Ericson, M.E.; Hosoi, J.; Seiffert, K.; Hordinsky, M.K.; Ansel, J.C.; Paus, R.; Scholzen, T.E. Neuropeptide control mechanisms in cutaneous biology: Physiological and clinical significance. J. Investig. Dermatol. 2006, 126, 1937–1947. [Google Scholar] [CrossRef] [PubMed]
- Shun, C.-T.; Chang, Y.-C.; Wu, H.-P.; Hsieh, S.-C.; Lin, W.-M.; Lin, Y.-H.; Tai, T.-Y.; Hsieh, S.-T. Skin denervation in type 2 diabetes: Correlations with diabetic duration and functional impairments. Brain 2004, 127, 1593–1605. [Google Scholar] [CrossRef] [PubMed]
- Skorvanek, M.; Bhatia, K.P. The Skin and Parkinson’s Disease: Review of Clinical, Diagnostic, and Therapeutic Issues. Mov. Disord. Clin. Pract. 2016, 4, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Faerman, I.; Faccio, E.; Calb, I.; Razumny, J.; Franco, N.; Dominguez, A.; Podestá, H.A. Autonomic neuropathy in the skin: A histological study of the sympathetic nerve fibres in diabetic anhidrosis. Diabetologia 1982, 22, 96–99. [Google Scholar] [CrossRef]
- Sun, P.-C.; Lin, H.-D.; Jao, S.-H.E.; Ku, Y.-C.; Chan, R.-C.; Cheng, C.-K. Relationship of skin temperature to sympathetic dysfunction in diabetic at-risk feet. Diabetes Res. Clin. Pract. 2006, 73, 41–46. [Google Scholar] [CrossRef]
- Zhifang, Z.; Yu, W.; Yishu, L.; Lulu, Z.; Jianbing, T.; Wenhua, H.; Biao, C. Lumbar sympathectomy regulates vascular cell turnover in rat hindfoot plantar skin. Clin. Hemorheol. Microcirc. 2017, 67, 149–157. [Google Scholar] [CrossRef]
- Cavin, J.-B.; Cuddihey, H.; MacNaughton, W.K.; Sharkey, K.A. Acute regulation of intestinal ion transport and permeability in response to luminal nutrients: The role of the enteric nervous system. Am. J. Physiol.-Gastrointest. Liver Physiol. 2020, 318, G254–G264. [Google Scholar] [CrossRef]
- You, X.-Y.; Zhang, H.-Y.; Han, X.; Wang, F.; Zhuang, P.-W.; Zhang, Y.-J. Intestinal Mucosal Barrier Is Regulated by Intestinal Tract Neuro-Immune Interplay. Front. Pharmacol. 2021, 12, 659716. [Google Scholar] [CrossRef] [PubMed]
- Kamimura, K.; Inoue, R.; Nagoya, T.; Sakai, N.; Goto, R.; Ko, M.; Niwa, Y.; Terai, S. Autonomic nervous system network and liver regeneration. World J. Gastroenterol. 2018, 24, 1616–1621. [Google Scholar] [CrossRef]
- Miller, B.M.; Oderberg, I.M.; Goessling, W. Hepatic Nervous System in Development, Regeneration, and Disease. Hepatology 2021, 74, 3513–3522. [Google Scholar] [CrossRef]
- Inoue, R.; Kamimura, K.; Nagoya, T.; Sakai, N.; Yokoo, T.; Goto, R.; Ogawa, K.; Shinagawa-Kobayashi, Y.; Watanabe-Mori, Y.; Sakamaki, A.; et al. Effect of a neural relay on liver regeneration in mice: Activation of serotonin release from the gastrointestinal tract. FEBS Open Bio 2018, 8, 449–460. [Google Scholar] [CrossRef]
- Kiba, T.; Tanaka, K.; Numata, K.; Hoshino, M.; Misugi, K.; Inoue, S. Ventromedial hypothalamic lesion-induced vagal hyperactivity stimulates rat pancreatic cell proliferation. Gastroenterology 1996, 110, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, T.; Liu, L. Hepatic branch vagotomy can block liver regeneration enhanced by ursodesoxycholic acid in 66% hepatectomized rats. Auton. Neurosci. 2002, 99, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Jiang, T.; Aji, T.; Aimulajiang, K.; Liu, Y.; Lv, G.; Wen, H. Netrin-1 promotes liver regeneration possibly by facilitating vagal nerve repair after partial hepatectomy in mice. Cell. Signal. 2022, 91, 110227. [Google Scholar] [CrossRef] [PubMed]
- Teratani, T.; Mikami, Y.; Nakamoto, N.; Suzuki, T.; Harada, Y.; Okabayashi, K.; Hagihara, Y.; Taniki, N.; Kohno, K.; Shibata, S.; et al. The liver–brain–gut neural arc maintains the Treg cell niche in the gut. Nature 2020, 585, 591–596. [Google Scholar] [CrossRef]
- Buhner, S.; Li, Q.; Vignali, S.; Barbara, G.; De Giorgio, R.; Stanghellini, V.; Cremon, C.; Zeller, F.; Langer, R.; Daniel, H.; et al. Activation of Human Enteric Neurons by Supernatants of Colonic Biopsy Specimens from Patients with Irritable Bowel Syndrome. Gastroenterology 2009, 137, 1425–1434. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, R.; Bai, T.; Xiang, X.; Qian, W.; Song, J.; Hou, X. EphrinB2/ephB2-mediated myenteric synaptic plasticity: Mechanisms underlying the persistent muscle hypercontractility and pain in postinfectious IBS. FASEB J. 2019, 33, 13644–13659. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, R.; Chen, Y.; Yang, P.; Bai, T.; Song, J.; Hou, X. EphrinB2/ephB2 activation facilitates colonic synaptic potentiation and plasticity contributing to long-term visceral hypersensitivity in irritable bowel syndrome. Life Sci. 2022, 295, 120419. [Google Scholar] [CrossRef] [PubMed]
- Drummond, M.J.; Dreyer, H.C.; Pennings, B.; Fry, C.S.; Dhanani, S.; Dillon, E.L.; Sheffield-Moore, M.; Volpi, E.; Rasmussen, B.B. Skeletal muscle protein anabolic response to resistance exercise and essential amino acids is delayed with aging. J. Appl. Physiol. 2008, 104, 1452–1461. [Google Scholar] [CrossRef] [PubMed]
- Hodges, P.W.; James, G.; Blomster, L.; Hall, L.; Schmid, A.; Shu, C.; Little, C.; Melrose, J. Multifidus Muscle Changes After Back Injury Are Characterized by Structural Remodeling of Muscle, Adipose and Connective Tissue, but Not Muscle Atrophy: Molecular and Morphological Evidence. Spine 2015, 40, 1057–1071. [Google Scholar] [CrossRef]
- Jeffery, P.K. Remodeling in asthma and chronic obstructive lung disease. Am. J. Respir. Crit. Care Med. 2001, 164, S28–S38. [Google Scholar] [CrossRef]
- Zaglia, T.; Milan, G.; Franzoso, M.; Bertaggia, E.; Pianca, N.; Piasentini, E.; Voltarelli, V.A.; Chiavegato, D.; Brum, P.C.; Glass, D.J.; et al. Cardiac sympathetic neurons provide trophic signal to the heart via β2-adrenoceptor-dependent regulation of proteolysis. Cardiovasc. Res. 2013, 97, 240–250. [Google Scholar] [CrossRef]
- Chambers, J.D.; Bornstein, J.C.; Thomas, E.A. Insights into mechanisms of intestinal segmentation in guinea pigs: A combined computational modeling and in vitro study. Am. J. Physiol.-Gastrointest. Liver Physiol. 2008, 295, G534–G541. [Google Scholar] [CrossRef]
- Fairfax, S.T.; Padilla, J.; Vianna, L.C.; Davis, M.J.; Fadel, P.J. Spontaneous bursts of muscle sympathetic nerve activity decrease leg vascular conductance in resting humans. Am. J. Physiol.-Heart Circ. Physiol. 2013, 304, H759–H766. [Google Scholar] [CrossRef]
- Holwerda, S.W.; Restaino, R.M.; Fadel, P.J. Adrenergic and non-adrenergic control of active skeletal muscle blood flow: Implications for blood pressure regulation during exercise. Auton. Neurosci. 2015, 188, 24–31. [Google Scholar] [CrossRef]
- Nausch, L.W.M.; Bonev, A.D.; Heppner, T.J.; Tallini, Y.; Kotlikoff, M.I.; Nelson, M.T. Sympathetic nerve stimulation induces local endothelial Ca2+ signals to oppose vasoconstriction of mouse mesenteric arteries. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, H594–H602. [Google Scholar] [CrossRef]
- Shoemaker, J.K.; Badrov, M.B.; Al-Khazraji, B.K.; Jackson, D.N. Neural control of vascular function in skeletal muscle. In Comprehensive Physiology; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2015; pp. 303–329. [Google Scholar]
- Hedner, J.; Sellgren, J.; Ejnell, H.; Wallin, G. High and fluctuating muscle nerve sympathetic activity in the sleep apnea syndrome: A pathogenetic mechanism in the development of hypertension? In Control of Breathing During Sleep and Anesthesia; Karczewski, W.A., Grieb, P., Kulesza, J., Bonsignore, G., Eds.; Springer: Boston, MA, USA, 1988; pp. 55–59. [Google Scholar]
- Jacob, D.W.; Harper, J.L.; Ivie, C.L.; Ott, E.P.; Limberg, J.K. Sex differences in the vascular response to sympathetic activation during acute hypoxaemia. Exp. Physiol. 2021, 106, 1689–1698. [Google Scholar] [CrossRef]
- Dalise, S.; Azzollini, V.; Chisari, C. Brain and Muscle: How Central Nervous System Disorders Can Modify the Skeletal Muscle. Diagnostics 2020, 10, 1047. [Google Scholar] [CrossRef] [PubMed]
- Falk, D.J.; Galatas, T.; Todd, A.G.; Soto, E.P.; Harris, A.B.; Notterpek, L. Locomotor and skeletal muscle abnormalities in trembler J neuropathic mice. Muscle Nerve 2018, 57, 664–671. [Google Scholar] [CrossRef]
- Garner, D.J.P.; Widrick, J.J. Cross-bridge mechanisms of muscle weakness in multiple sclerosis. Muscle Nerve 2003, 27, 456–464. [Google Scholar] [CrossRef]
- Wens, I.; Dalgas, U.; Vandenabeele, F.; Krekels, M.; Grevendonk, L.; Eijnde, B.O. Multiple Sclerosis Affects Skeletal Muscle Characteristics. PLoS ONE 2014, 9, e108158. [Google Scholar] [CrossRef]
- Willingham, T.B.; Backus, D.; McCully, K.K. Muscle Dysfunction and Walking Impairment in Women with Multiple Sclerosis. Int. J. MS Care 2019, 21, 249–256. [Google Scholar] [CrossRef]
- Nozoe, M.; Kanai, M.; Kubo, H.; Takeuchi, Y.; Kobayashi, M.; Yamamoto, M.; Furuichi, A.; Yamazaki, M.; Shimada, S.; Mase, K. Efficacy of neuromuscular electrical stimulation for preventing quadriceps muscle wasting in patients with moderate or severe acute stroke: A pilot study. NeuroRehabilitation 2017, 41, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Scherbakov, N.; von Haehling, S.; Anker, S.D.; Dirnagl, U.; Doehner, W. Stroke induced Sarcopenia: Muscle wasting and disability after stroke. Int. J. Cardiol. 2013, 170, 89–94. [Google Scholar] [CrossRef]
- Nozoe, M.; Kubo, H.; Kanai, M.; Yamamoto, M.; Shimada, S.; Mase, K. Peripheral motor nerve conduction abnormality, muscle strength, and muscle wasting in patients with acute stroke: A pilot study. J. Clin. Neurosci. 2020, 75, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Braun, T.P.; Zhu, X.; Szumowski, M.; Scott, G.D.; Grossberg, A.J.; Levasseur, P.R.; Graham, K.; Khan, S.; Damaraju, S.; Colmers, W.F.; et al. Central nervous system inflammation induces muscle atrophy via activation of the hypothalamic–pituitary–adrenal axis. J. Exp. Med. 2011, 208, 2449–2463. [Google Scholar] [CrossRef]
- Kim, A.R.; Pavlenko, T.A.; Katargina, L.A.; Chesnokova, N.B.; Ugrumov, M.V. Biochemical and Functional Changes in the Eye as a Manifestation of Systemic Degeneration of the Nervous System in Parkinsonism. Acta Naturae 2018, 10, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Lopatina, E.V.; Penniyaynen, V.A.; Tsyrline, V.A. Impact of norepinephrine and selective β1-adrenoceptor blockers on the growth of retinal tissue explants. Bull. Exp. Biol. Med. 2012, 153, 48–50. [Google Scholar] [CrossRef]
- Li, Y.-K.; Yang, J.-M.; Huang, Y.-B.; Ren, D.-D.; Chi, F.-L. Shrinkage of ipsilateral taste buds and hyperplasia of contralateral taste buds following chorda tympani nerve transection. Neural Regen. Res. 2015, 10, 989–995. [Google Scholar] [CrossRef]
- Castillo-Azofeifa, D.; Losacco, J.T.; Salcedo, E.; Golden, E.J.; Finger, T.E.; Barlow, L.A. Sonic hedgehog from both nerves and epithelium is a key trophic factor for taste bud maintenance. Development 2017, 144, 3054–3065. [Google Scholar] [CrossRef] [PubMed]
- Nagato, T.; Matsumoto, K.; Tanioka, H.; Kodama, J.; Toh, H. Effect of denervation on morphogenesis of the rat fungiform papilla. Acta Anat. 1995, 153, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Fife, K.; Herd, R.; Lalondrelle, S.; Plummer, R.; Strong, A.; Jones, S.; Lear, J.T. Managing adverse events associated with vismodegib in the treatment of basal cell carcinoma. Future Oncol. 2017, 13, 175–184. [Google Scholar] [CrossRef]
- Kumari, A.; Ermilov, A.N.; Grachtchouk, M.; Dlugosz, A.A.; Allen, B.L.; Bradley, R.M.; Mistretta, C.M. Recovery of taste organs and sensory function after severe loss from Hedgehog/Smoothened inhibition with cancer drug sonidegib. Proc. Natl. Acad. Sci. USA 2017, 114, E10369–E10378. [Google Scholar] [CrossRef]
- Magnon, C.; Hall, S.J.; Lin, J.; Xue, X.; Gerber, L.; Freedland, S.J.; Frenette, P.S. Autonomic Nerve Development Contributes to Prostate Cancer Progression. Science 2013, 341, 1236361. [Google Scholar] [CrossRef]
- Schwartz, E.S.; Christianson, J.A.; Chen, X.; La, J.-H.; Davis, B.M.; Albers, K.M.; Gebhart, G.F. Synergistic Role of TRPV1 and TRPA1 in Pancreatic Pain and Inflammation. Gastroenterology 2011, 140, 1283–1291.e1282. [Google Scholar] [CrossRef]
- Schwartz, E.S.; La, J.-H.; Scheff, N.N.; Davis, B.M.; Albers, K.M.; Gebhart, G.F. TRPV1 and TRPA1 Antagonists Prevent the Transition of Acute to Chronic Inflammation and Pain in Chronic Pancreatitis. J. Neurosci. 2013, 33, 5603–5611. [Google Scholar] [CrossRef] [PubMed]
- Stopczynski, R.E.; Normolle, D.P.; Hartman, D.J.; Ying, H.; DeBerry, J.J.; Bielefeldt, K.; Rhim, A.D.; DePinho, R.A.; Albers, K.M.; Davis, B.M. Neuroplastic changes occur early in the development of pancreatic ductal adenocarcinoma. Cancer Res. 2014, 74, 1718–1727. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.-M.; Hayakawa, Y.; Kodama, Y.; Muthupalani, S.; Westphalen, C.B.; Andersen, G.T.; Flatberg, A.; Johannessen, H.; Friedman, R.A.; Renz, B.W.; et al. Denervation suppresses gastric tumorigenesis. Sci. Transl. Med. 2014, 6, 250ra115. [Google Scholar] [CrossRef] [PubMed]
- Saloman, J.L.; Albers, K.M.; Rhim, A.D.; Davis, B.M. Can Stopping Nerves, Stop Cancer? Trends Neurosci. 2016, 39, 880–889. [Google Scholar] [CrossRef]
- Fernandes, J.; VijayRaghavan, K. The development of indirect flight muscle innervation in Drosophila melanogaster. Development 1993, 118, 215–227. [Google Scholar] [CrossRef]
- Fernandes, J.J.; Keshishian, H. Nerve-muscle interactions during flight muscle development in Drosophila. Development 1998, 125, 1769–1779. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Toral, M.; Siefert, M.; Conway, D.; Dorr, M.; Fernandes, J. dHb9 expressing larval motor neurons persist through metamorphosis to innervate adult-specific muscle targets and function in Drosophila eclosion. Dev. Neurobiol. 2016, 76, 1387–1416. [Google Scholar] [CrossRef]
- Hegstrom, C.D.; Riddiford, L.M.; Truman, J.W. Steroid and Neuronal Regulation of Ecdysone Receptor Expression during Metamorphosis of Muscle in the Moth, Manduca sexta. J. Neurosci. 1998, 18, 1786–1794. [Google Scholar] [CrossRef]
- Kempf, S.C.; Page, L.R. Anti-Tubulin Labeling Reveals Ampullary Neuron Ciliary Bundles in Opisthobranch Larvae and a New Putative Neural Structure Associated with the Apical Ganglion. Biol. Bull. 2005, 208, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Marois, R.; Carew, T.J. The gastropod nervous system in metamorphosis. J. Neurobiol. 1990, 21, 1053–1071. [Google Scholar] [CrossRef] [PubMed]
- Hadfield, M.; Meleshkevitch, E.; Boudko, D. The apical sensory organ of a gastropod veliger is a receptor for settlement cues. Biol. Bull. 2000, 198, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Page, L.R. Comparative structure of the larval apical sensory organ in gastropods and hypotheses about function and developmental evolution. Invertebr. Reprod. Dev. 2002, 41, 193–200. [Google Scholar] [CrossRef]
- Caicci, F.; Zaniolo, G.; Burighel, P.; Degasperi, V.; Gasparini, F.; Manni, L. Differentiation of papillae and rostral sensory neurons in the larva of the ascidian Botryllus schlosseri (Tunicata). J. Comp. Neurol. 2010, 518, 547–566. [Google Scholar] [CrossRef]
- Kimura, Y.; Yoshida, M.; Morisawa, M. Interaction between noradrenaline or adrenaline and the β1-adrenergic receptor in the nervous system triggers early metamorphosis of larvae in the ascidian, Ciona savignyi. Dev. Biol. 2003, 258, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Nakayama-Ishimura, A.; Chambon, J.-P.; Horie, T.; Satoh, N.; Sasakura, Y. Delineating metamorphic pathways in the ascidian Ciona intestinalis. Dev. Biol. 2009, 326, 357–367. [Google Scholar] [CrossRef]
- Ryan, K.; Lu, Z.; Meinertzhagen, I.A. The peripheral nervous system of the ascidian tadpole larva: Types of neurons and their synaptic networks. J. Comp. Neurol. 2018, 526, 583–608. [Google Scholar] [CrossRef] [PubMed]
- Zega, G.; Pennati, R.; Groppelli, S.; Sotgia, C.; Bernardi, F.D. Dopamine and serotonin modulate the onset of metamorphosis in the ascidian Phallusia mammillata. Dev. Biol. 2005, 282, 246–256. [Google Scholar] [CrossRef]
- Costlow, J.D. The effect of eyestalk extirpation on larval development of the mud crab, Rhithropanopeus harrisii (Gould). Gen. Comp. Endocrinol. 1966, 7, 255–274. [Google Scholar] [CrossRef]
- Dan, S.; Kaneko, T.; Takeshima, S.; Ashidate, M.; Hamasaki, K. Eyestalk ablation affects larval morphogenesis in the swimming crab Portunus trituberculatus during metamorphosis into megalopae. Sex. Early Dev. Aquat. Org. 2014, 1, 57–73. [Google Scholar] [CrossRef]
- Denver, R.J. Chapter seven—Neuroendocrinology of amphibian metamorphosis. In Current Topics in Developmental Biology; Shi, Y.-B., Ed.; Academic Press: Cambridge, MA, USA, 2013; Volume 103, pp. 195–227. [Google Scholar]
- Paul, B.; Sterner, Z.R.; Buchholz, D.R.; Shi, Y.-B.; Sachs, L.M. Thyroid and Corticosteroid Signaling in Amphibian Metamorphosis. Cells 2022, 11, 1595. [Google Scholar] [CrossRef] [PubMed]
- Ruthsatz, K.; Dausmann, K.H.; Drees, C.; Becker, L.I.; Hartmann, L.; Reese, J.; Reinhardt, S.; Robinson, T.; Sabatino, N.M.; Peck, M.A.; et al. Altered thyroid hormone levels affect the capacity for temperature-induced developmental plasticity in larvae of Rana temporaria and Xenopus laevis. J. Therm. Biol. 2020, 90, 102599. [Google Scholar] [CrossRef] [PubMed]
- Shibata, Y.; Wen, L.; Okada, M.; Shi, Y.-B. Organ-Specific Requirements for Thyroid Hormone Receptor Ensure Temporal Coordination of Tissue-Specific Transformations and Completion of Xenopus Metamorphosis. Thyroid 2020, 30, 300–313. [Google Scholar] [CrossRef] [PubMed]
- Dietz, F.R. Effect of peripheral nerve on limb development. J. Orthop. Res. 1987, 5, 576–585. [Google Scholar] [CrossRef]
- Marsh-Armstrong, N.; Cai, L.; Brown, D.D. Thyroid hormone controls the development of connections between the spinal cord and limbs during Xenopus laevis metamorphosis. Proc. Natl. Acad. Sci. USA 2004, 101, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Graf, W.; Baker, R. Neuronal adaptation accompanying metamorphosis in the flatfish. J. Neurobiol. 1990, 21, 1136–1152. [Google Scholar] [CrossRef]
- Kavouras, M.; Malandrakis, E.E.; Blom, E.; Tsilika, K.; Danis, T.; Panagiotaki, P.; Exadactylos, A. Malpigmentation of Common Sole (Solea solea) during Metamorphosis Is Associated with Differential Synaptic-Related Gene Expression. Animals 2021, 11, 2273. [Google Scholar] [CrossRef] [PubMed]
- Budi, E.H.; Patterson, L.B.; Parichy, D.M. Post-Embryonic Nerve-Associated Precursors to Adult Pigment Cells: Genetic Requirements and Dynamics of Morphogenesis and Differentiation. PLoS Genet. 2011, 7, e1002044. [Google Scholar] [CrossRef]
- Dooley, C.M.; Mongera, A.; Walderich, B.; Nüsslein-Volhard, C. On the embryonic origin of adult melanophores: The role of ErbB and Kit signalling in establishing melanophore stem cells in zebrafish. Development 2013, 140, 1003–1013. [Google Scholar] [CrossRef]
- Singh, A.P.; Dinwiddie, A.; Mahalwar, P.; Schach, U.; Linker, C.; Irion, U.; Nüsslein-Volhard, C. Pigment Cell Progenitors in Zebrafish Remain Multipotent through Metamorphosis. Dev. Cell 2016, 38, 316–330. [Google Scholar] [CrossRef] [PubMed]
- Buchholz, D.R. More similar than you think: Frog metamorphosis as a model of human perinatal endocrinology. Dev. Biol. 2015, 408, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Faunes, F.; Larraín, J. Conservation in the involvement of heterochronic genes and hormones during developmental transitions. Dev. Biol. 2016, 416, 3–17. [Google Scholar] [CrossRef]
- Fisher, D.A.; Dussault, J.H.; Sack, J.; Chopra, I.J. Ontogenesis of Hypothalamic-Pituitary-Thyroid Function and Metabolism in Man, Sheep, and Rat. In Proceedings of the 1976 Laurentian Hormone Conference, Mont Tremblant, QC, Canada, 21 November 1997; Greep, R.O., Ed.; Academic Press: Boston, MA, USA, 1977; Volume 33, pp. 59–116. [Google Scholar]
- Hadj-Sahraoui, N.; Seugnet, I.; Ghorbel, M.T.; Demeneix, B. Hypothyroidism prolongs mitotic activity in the post-natal mouse brain. Neurosci. Lett. 2000, 280, 79–82. [Google Scholar] [CrossRef]
- Polak, M.; Le Gac, I.; Vuillard, E.; Guibourdenche, J.; Leger, J.; Toubert, M.E.; Madec, A.M.; Oury, J.F.; Czernichow, P.; Luton, D. Fetal and neonatal thyroid function in relation to maternal Graves’ disease. Best Pract. Res. Clin. Endocrinol. Metab. 2004, 18, 289–302. [Google Scholar] [CrossRef]
- Elks, C.E.; Perry, J.R.B.; Sulem, P.; Chasman, D.I.; Franceschini, N.; He, C.; Lunetta, K.L.; Visser, J.A.; Byrne, E.M.; Cousminer, D.L.; et al. Thirty new loci for age at menarche identified by a meta-analysis of genome-wide association studies. Nat. Genet. 2010, 42, 1077–1085. [Google Scholar] [CrossRef] [PubMed]
- Gundermann, D.G.; Martínez, J.; De Kervor, G.; González-Pinto, K.; Larraín, J.; Faunes, F. Overexpression of Lin28a delays Xenopus metamorphosis and down-regulates albumin independently of its translational regulation domain. Dev. Dyn. 2019, 248, 969–978. [Google Scholar] [CrossRef] [PubMed]
- Ong, K.K.; Elks, C.E.; Li, S.; Zhao, J.H.; Luan, J.A.; Andersen, L.B.; Bingham, S.A.; Brage, S.; Smith, G.D.; Ekelund, U.; et al. Genetic variation in LIN28B is associated with the timing of puberty. Nat. Genet. 2009, 41, 729–733. [Google Scholar] [CrossRef] [PubMed]
- Carr, M.J.; Toma, J.S.; Johnston, A.P.W.; Steadman, P.E.; Yuzwa, S.A.; Mahmud, N.; Frankland, P.W.; Kaplan, D.R.; Miller, F.D. Mesenchymal Precursor Cells in Adult Nerves Contribute to Mammalian Tissue Repair and Regeneration. Cell Stem Cell 2019, 24, 240–256.e249. [Google Scholar] [CrossRef]
- Stocum, D.L. Mechanisms of urodele limb regeneration. Regeneration 2017, 4, 159–200. [Google Scholar] [CrossRef]
- Aztekin, C. Tissues and Cell Types of Appendage Regeneration: A Detailed Look at the Wound Epidermis and Its Specialized Forms. Front. Physiol. 2021, 12, 771040. [Google Scholar] [CrossRef] [PubMed]
- Morgan, T.H. Experimental studies of the regeneration of Planaria maculata. Arch. Entwickelungsmechanik Org. 1898, 7, 364–397. [Google Scholar] [CrossRef]
- Oviedo, N.J.; Morokuma, J.; Walentek, P.; Kema, I.P.; Gu, M.B.; Ahn, J.-M.; Hwang, J.S.; Gojobori, T.; Levin, M. Long-range Neural and Gap Junction Protein-mediated Cues Control Polarity During Planarian Regeneration. Dev. Biol. 2010, 339, 188–199. [Google Scholar] [CrossRef]
- Pietak, A.; Bischof, J.; LaPalme, J.; Morokuma, J.; Levin, M. Neural control of body-plan axis in regenerating planaria. PLoS Comput. Biol. 2019, 15, e1006904. [Google Scholar] [CrossRef] [PubMed]
- Arnold, C.P.; Benham-Pyle, B.W.; Lange, J.J.; Wood, C.J.; Sánchez Alvarado, A. Wnt and TGFβ coordinate growth and patterning to regulate size-dependent behaviour. Nature 2019, 572, 655–659. [Google Scholar] [CrossRef]
- Sarkar, A.; Mukundan, N.; Sowndarya, S.; Dubey, V.K.; Babu, R.; Lakshmanan, V.; Rangiah, K.; Panicker, M.M.; Palakodeti, D.; Subramanian, S.P.; et al. Serotonin is essential for eye regeneration in planaria Schmidtea mediterranea. FEBS Lett. 2019, 593, 3198–3209. [Google Scholar] [CrossRef]
- Lambert, F.M.; Cardoit, L.; Courty, E.; Bougerol, M.; Thoby-Brisson, M.; Simmers, J.; Tostivint, H.; Le Ray, D. Functional limb muscle innervation prior to cholinergic transmitter specification during early metamorphosis in Xenopus. eLife 2018, 7, e30693. [Google Scholar] [CrossRef]
- Kumar, A.; Godwin, J.W.; Gates, P.B.; Garza-Garcia, A.A.; Brockes, J.P. Molecular Basis for the Nerve Dependence of Limb Regeneration in an Adult Vertebrate. Science 2007, 318, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Nevill, G.; Brockes, J.P.; Forge, A. A comparative study of gland cells implicated in the nerve dependence of salamander limb regeneration. J. Anat. 2010, 217, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Singer, M. A Theory of Trophic Nervous Control of Amphibian Limb Regeneration, Including A Re-Evaluation of Quantitative Nerve Requirements; North-Holland Publ. Co.: Amsterdam, The Netherlands, 1965; pp. 20–32. [Google Scholar]
- Bryant, D.M.; Sousounis, K.; Farkas, J.E.; Bryant, S.; Thao, N.; Guzikowski, A.R.; Monaghan, J.R.; Levin, M.; Whited, J.L. Repeated removal of developing limb buds permanently reduces appendage size in the highly-regenerative axolotl. Dev. Biol. 2017, 424, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Farkas, J.E.; Freitas, P.D.; Bryant, D.M.; Whited, J.L.; Monaghan, J.R. Neuregulin-1 signaling is essential for nerve-dependent axolotl limb regeneration. Development 2016, 143, 2724–2731. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.E.; Salhotra, A.; Robertson, K.S.; Ransom, R.C.; Foster, D.S.; Shah, H.N.; Quarto, N.; Wan, D.C.; Longaker, M.T. Skeletal Stem Cell-Schwann Cell Circuitry in Mandibular Repair. Cell Rep. 2019, 28, 2757–2766.e2755. [Google Scholar] [CrossRef] [PubMed]
- Muneoka, K.; Holler-Dinsmore, G.; Bryant, S.V. Intrinsic control of regenerative loss in Xenopus laevis limbs. J. Exp. Zool. 1986, 240, 47–54. [Google Scholar] [CrossRef]
- Filoni, S.; Paglialunga, L. Effect of denervation on hindlimb regeneration in Xenopus laevis larvae. Differentiation 1990, 43, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Gaze, R.M. Regeneration of the Optic Nerve in Xenopus Lævis. Q. J. Exp. Physiol. Cogn. Med. Sci. 1959, 44, 290–308. [Google Scholar] [CrossRef] [PubMed]
- Slater, P.G.; Palacios, M.; Larraín, J. Xenopus, a Model to Study Wound Healing and Regeneration: Experimental Approaches. Cold Spring Harb. Protoc. 2021, 2021, pdb.top100966. [Google Scholar] [CrossRef]
- Taniguchi, Y.; Sugiura, T.; Tazaki, A.; Watanabe, K.; Mochii, M. Spinal cord is required for proper regeneration of the tail in Xenopus tadpoles. Dev. Growth Differ. 2008, 50, 109–120. [Google Scholar] [CrossRef]
- Mondia, J.P.; Levin, M.; Omenetto, F.G.; Orendorff, R.D.; Branch, M.R.; Adams, D.S. Long-Distance Signals Are Required for Morphogenesis of the Regenerating Xenopus Tadpole Tail, as Shown by Femtosecond-Laser Ablation. PLoS ONE 2011, 6, e24953. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, J.; Tochinai, S. Functional regeneration of the olfactory bulb requires reconnection to the olfactory nerve in Xenopus larvae. Dev. Growth Differ. 2006, 48, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Satoh, A.; Ide, H.; Tamura, K. Nerve-dependent and -independent events in blastema formation during Xenopus froglet limb regeneration. Dev. Biol. 2005, 286, 361–375. [Google Scholar] [CrossRef]
- Yokoyama, H.; Maruoka, T.; Ochi, H.; Aruga, A.; Ohgo, S.; Ogino, H.; Tamura, K. Different Requirement for Wnt/β-Catenin Signaling in Limb Regeneration of Larval and Adult Xenopus. PLoS ONE 2011, 6, e21721. [Google Scholar] [CrossRef] [PubMed]
- Mitogawa, K.; Makanae, A.; Satoh, A. Hyperinnervation improves Xenopus laevis limb regeneration. Dev. Biol. 2018, 433, 276–286. [Google Scholar] [CrossRef]
- Geraudie, J.; Singer, M. Relation between nerve fiber number and pectoral fin regeneration in the teleost. J. Exp. Zool. 1977, 199, 1–8. [Google Scholar] [CrossRef]
- Simões, M.G.; Bensimon-Brito, A.; Fonseca, M.; Farinho, A.; Valério, F.; Sousa, S.; Afonso, N.; Kumar, A.; Jacinto, A. Denervation impairs regeneration of amputated zebrafish fins. BMC Dev. Biol. 2014, 14, 49. [Google Scholar] [CrossRef]
- Johnston, A.P.W.; Yuzwa, S.A.; Carr, M.J.; Mahmud, N.; Storer, M.A.; Krause, M.P.; Jones, K.; Paul, S.; Kaplan, D.R.; Miller, F.D. Dedifferentiated Schwann Cell Precursors Secreting Paracrine Factors Are Required for Regeneration of the Mammalian Digit Tip. Cell Stem Cell 2016, 19, 433–448. [Google Scholar] [CrossRef]
- Rinkevich, Y.; Montoro, D.T.; Muhonen, E.; Walmsley, G.G.; Lo, D.; Hasegawa, M.; Januszyk, M.; Connolly, A.J.; Weissman, I.L.; Longaker, M.T. Clonal analysis reveals nerve-dependent and independent roles on mammalian hind limb tissue maintenance and regeneration. Proc. Natl. Acad. Sci. USA 2014, 111, 9846–9851. [Google Scholar] [CrossRef]
- Pinet, K.; McLaughlin, K.A. Mechanisms of physiological tissue remodeling in animals: Manipulating tissue, organ, and organism morphology. Dev. Biol. 2019, 451, 134–145. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Adams, D.S.; Levin, M. Normalized shape and location of perturbed craniofacial structures in the Xenopus tadpole reveal an innate ability to achieve correct morphology. Dev. Dyn. 2012, 241, 863–878. [Google Scholar] [CrossRef]
- Govind, C.K.; Pearce, J. Remodeling of nerves during claw reversal in adult snapping shrimps. J. Comp. Neurol. 1988, 268, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Young, R.E.; Wong, A.; Pearce, J.; Govind, C.K. Neural factors influence the degeneration of muscle fibers in the chelae of snapping shrimps. Mol. Chem. Neuropathol. 1996, 28, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Suttie, J.M. Experimental Manipulation of the Neural Control of Antler Growth. In Horns, Pronghorns, and Antlers: Evolution, Morphology, Physiology, and Social Significance; Bubenik, G.A., Bubenik, A.B., Eds.; Springer: New York, NY, USA, 1990; pp. 359–370. [Google Scholar]
- Boettiger, A.; Ermentrout, B.; Oster, G. The neural origins of shell structure and pattern in aquatic mollusks. Proc. Natl. Acad. Sci. USA 2009, 106, 6837–6842. [Google Scholar] [CrossRef] [PubMed]
- Jonz, M.G.; Fearon, I.M.; Nurse, C.A. Neuroepithelial oxygen chemoreceptors of the zebrafish gill. J. Physiol. 2004, 560, 737–752. [Google Scholar] [CrossRef] [PubMed]
- Sollid, J.; Nilsson, G.E. Plasticity of respiratory structures—Adaptive remodeling of fish gills induced by ambient oxygen and temperature. Respir. Physiol. Neurobiol. 2006, 154, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Long, H.; Ahmed, M.; Ackermann, P.; Stark, A.; Li, J. Neuropeptide Y innervation during fracture healing and remodeling. A study of angulated tibial fractures in the rat. Acta Orthop. 2010, 81, 639–646. [Google Scholar] [CrossRef]
- Allison, S.J.; Baldock, P.A.; Herzog, H. The control of bone remodeling by neuropeptide Y receptors. Peptides 2007, 28, 320–325. [Google Scholar] [CrossRef]
- Qin, Y.-Y.; Huang, X.-R.; Zhang, J.; Wu, W.; Chen, J.; Wan, S.; Yu, X.-Y.; Lan, H.-Y. Neuropeptide Y attenuates cardiac remodeling and deterioration of function following myocardial infarction. Mol. Ther. 2022, 30, 881–897. [Google Scholar] [CrossRef]
- Basson, M.D.; Burney, R.E. Defective wound healing in patients with paraplegia and quadriplegia. Surg. Gynecol. Obstet. 1982, 155, 9–12. [Google Scholar] [PubMed]
- Engin, C.; Demirkan, F.; Ayhan, S.; Atabay, K.; Baran, N.K. Delayed effect of denervation on wound contraction in rat skin. Plast. Reconstr. Surg. 1996, 98, 1063–1067. [Google Scholar] [CrossRef]
- Kiya, K.; Kubo, T. Neurovascular interactions in skin wound healing. Neurochem. Int. 2019, 125, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Liu, D.; Ying, J.; Yao, Z.; Hou, Q.; Wang, H.; Qi, F.; Luan, W.; Jiang, H. Denervation Affected Skin Wound Healing in a Modified Rat Model. Int. J. Low. Extrem. Wounds 2022, 22, 1–13. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jones, E.; McLaughlin, K.A. A Novel Perspective on Neuronal Control of Anatomical Patterning, Remodeling, and Maintenance. Int. J. Mol. Sci. 2023, 24, 13358. https://doi.org/10.3390/ijms241713358
Jones E, McLaughlin KA. A Novel Perspective on Neuronal Control of Anatomical Patterning, Remodeling, and Maintenance. International Journal of Molecular Sciences. 2023; 24(17):13358. https://doi.org/10.3390/ijms241713358
Chicago/Turabian StyleJones, Emilie, and Kelly A. McLaughlin. 2023. "A Novel Perspective on Neuronal Control of Anatomical Patterning, Remodeling, and Maintenance" International Journal of Molecular Sciences 24, no. 17: 13358. https://doi.org/10.3390/ijms241713358
APA StyleJones, E., & McLaughlin, K. A. (2023). A Novel Perspective on Neuronal Control of Anatomical Patterning, Remodeling, and Maintenance. International Journal of Molecular Sciences, 24(17), 13358. https://doi.org/10.3390/ijms241713358