
Advances in Platelet Rich Plasma-Derived Extracellular Vesicles for Regenerative Medicine: A Systematic-Narrative Review
 , , , and
, , , and 
Abstract
1. Introduction
2. Material and Methods
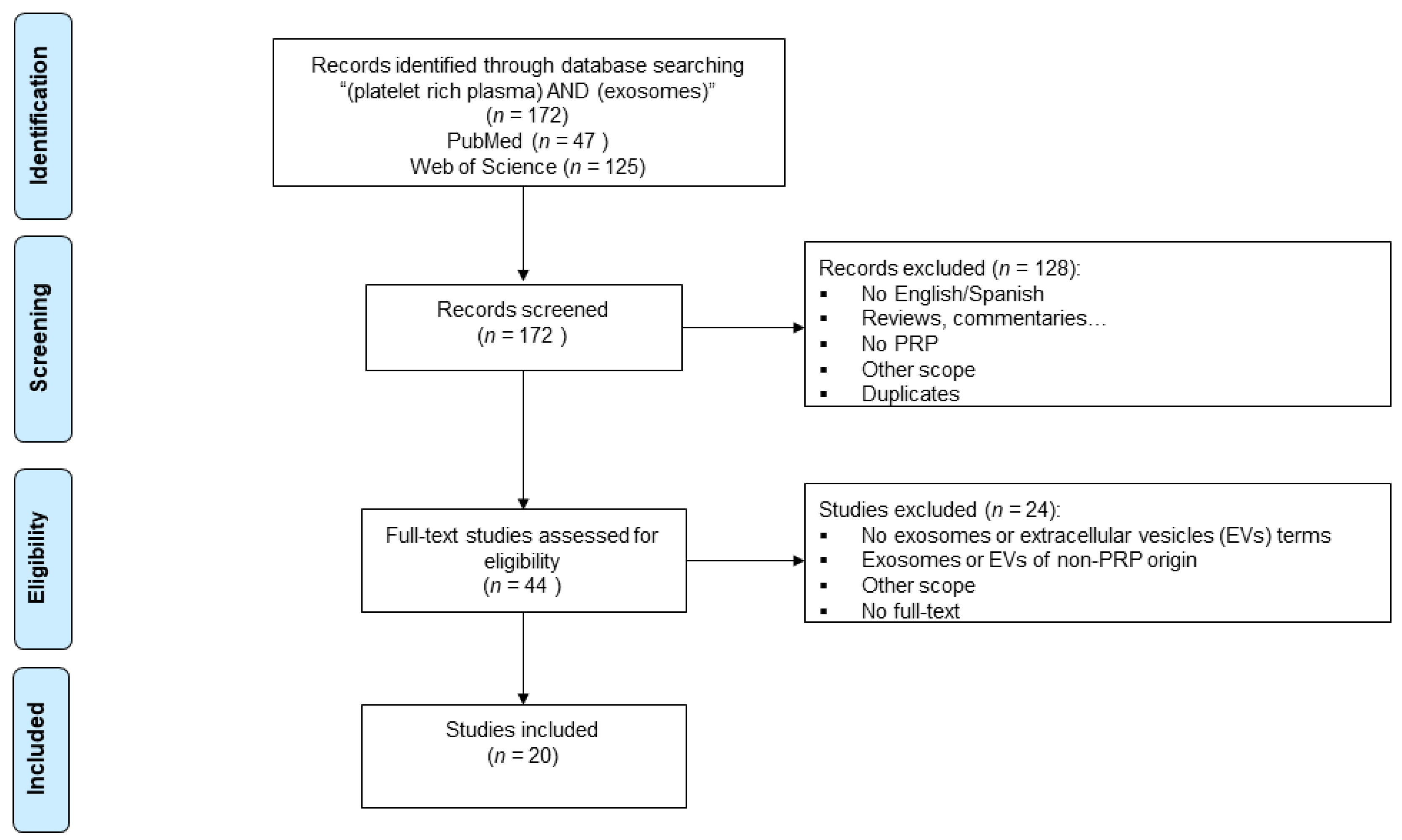
2.1. Literature Search
2.2. Data Extraction
2.3. Assessment of Reporting Quality and Risk of Bias
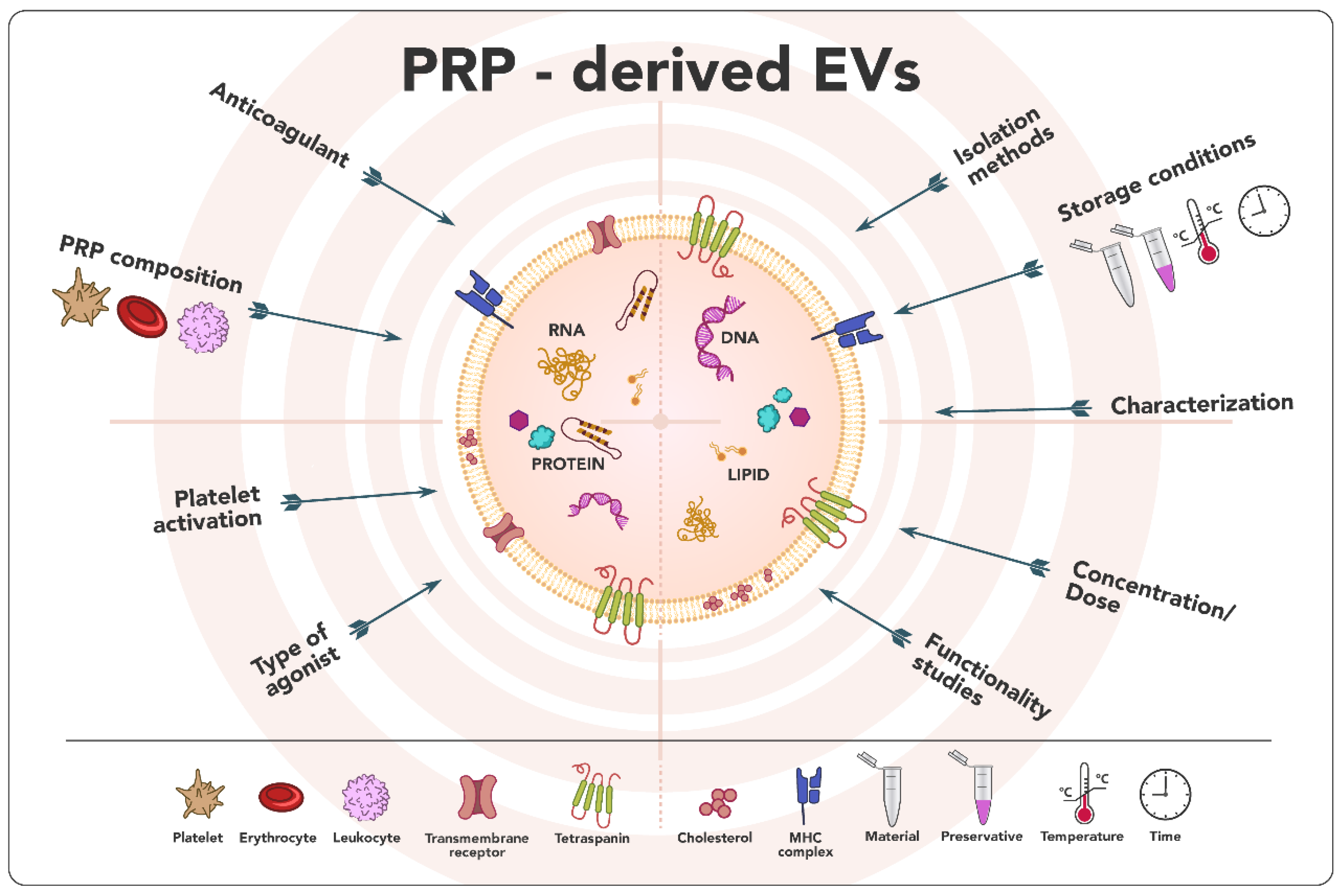
3. Results
3.1. Reporting Quality and Risk of Bias
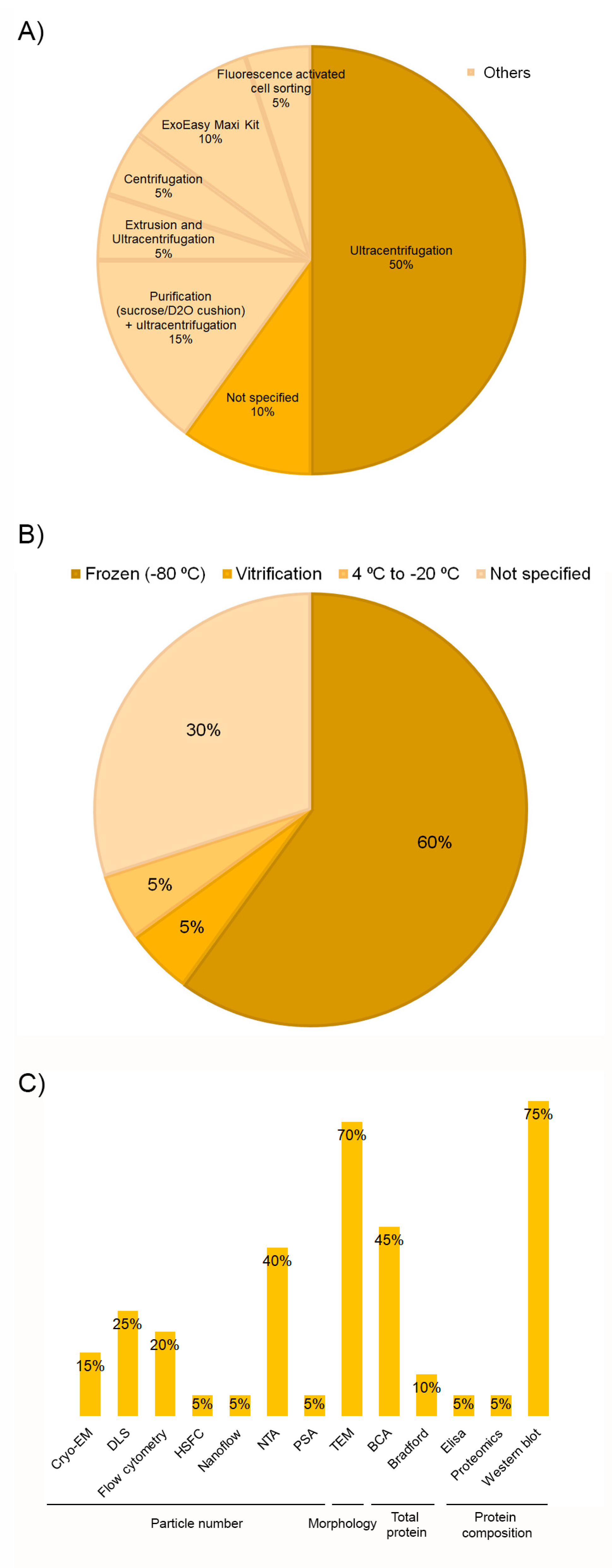
3.2. PRP Obtaining Process Affects Extracellular Vesicles Production
3.3. Extracellular Vesicles Obtaining Process and Storage Stability
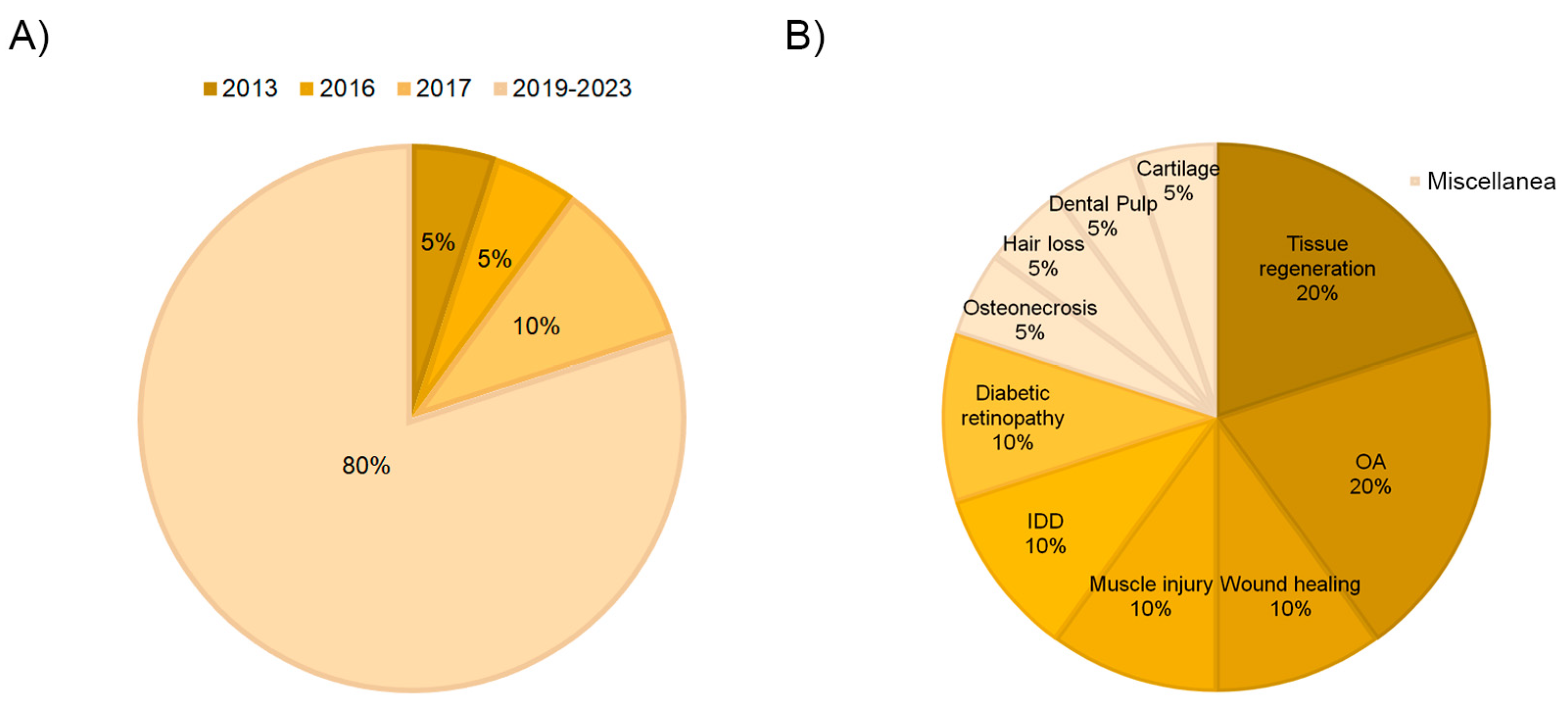
3.4. Fields of Application
3.5. Osteoarthritis
3.6. Tissue Regeneration
3.7. Diabetic Retinopathy
3.8. Intervertebral Disc Degeneration
3.9. Muscle Injury
3.10. Wound Healing
3.11. Miscellanea
3.11.1. Cartilage Regeneration
3.11.2. Dental Pulp Regeneration
3.11.3. Hair Loss
3.11.4. Osteonecrosis
4. Discussion and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Anitua, E. Plasma rich in growth factors: Preliminary results of use in the preparation of future sites for implants. Int. J. Oral Maxillofac. Implant. 1999, 14, 529–535. [Google Scholar]
- Marx, R.E.; Carlson, E.R.; Eichstaedt, R.M.; Schimmele, S.R.; Strauss, J.E.; Georgeff, K.R. Platelet-rich plasma: Growth factor enhancement for bone grafts. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 1998, 85, 638–646. [Google Scholar] [CrossRef]
- Anitua, E.; Troya, M.; Zalduendo, M.; Orive, G. Personalized plasma-based medicine to treat age-related diseases. Mater. Sci. Eng. C Mater. Biol. Appl. 2017, 74, 459–464. [Google Scholar] [CrossRef] [PubMed]
- DeLong, J.M.; Russell, R.P.; Mazzocca, A.D. Platelet-rich plasma: The PAW classification system. Arthroscopy 2012, 28, 998–1009. [Google Scholar] [CrossRef]
- Magalon, J.; Brandin, T.; Francois, P.; Degioanni, C.; De Maria, L.; Grimaud, F.; Veran, J.; Dignat-George, F.; Sabatier, F. Technical and biological review of authorized medical devices for platelets-rich plasma preparation in the field of regenerative medicine. Platelets 2021, 32, 200–208. [Google Scholar] [CrossRef]
- Acebes-Huerta, A.; Arias-Fernandez, T.; Bernardo, A.; Munoz-Turrillas, M.C.; Fernandez-Fuertes, J.; Seghatchian, J.; Gutierrez, L. Platelet-derived bio-products: Classification update, applications, concerns and new perspectives. Transfus. Apher. Sci. 2020, 59, 102716. [Google Scholar] [CrossRef] [PubMed]
- Anitua, E.; Padilla, S.; Prado, R.; Alkhraisat, M.H. Platelet-rich plasma: Are the obtaining methods, classification and clinical outcome always connected? Regen. Med. 2022, 17, 887–890. [Google Scholar] [CrossRef]
- Kon, E.; Di Matteo, B.; Delgado, D.; Cole, B.J.; Dorotei, A.; Dragoo, J.L.; Filardo, G.; Fortier, L.A.; Giuffrida, A.; Jo, C.H.; et al. Platelet-rich plasma for the treatment of knee osteoarthritis: An expert opinion and proposal for a novel classification and coding system. Expert. Opin. Biol. Ther. 2020, 20, 1447–1460. [Google Scholar] [CrossRef]
- van der Meijden, P.E.J.; Heemskerk, J.W.M. Platelet biology and functions: New concepts and clinical perspectives. Nat. Rev. Cardiol. 2019, 16, 166–179. [Google Scholar] [CrossRef]
- Sun, J.; Hu, Y.; Fu, Y.; Zou, D.; Lu, J.; Lyu, C. Emerging roles of platelet concentrates and platelet-derived extracellular vesicles in regenerative periodontology and implant dentistry. APL Bioeng. 2022, 6, 031503. [Google Scholar] [CrossRef] [PubMed]
- Anitua, E.; Troya, M.; Zalduendo, M.; Tejero, R.; Orive, G. Progress in the Use of Autologous Regenerative Platelet-based Therapies in Implant Dentistry. Curr. Pharm. Biotechnol. 2016, 17, 402–413. [Google Scholar] [CrossRef] [PubMed]
- Anitua, E.; Alkhraisat, M.H.; Orive, G. Perspectives and challenges in regenerative medicine using plasma rich in growth factors. J. Control. Release 2012, 157, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Anitua, E.; Troya, M.; Tierno, R.; Zalduendo, M.; Alkhraisat, M.H. The Effectiveness of Platelet-Rich Plasma as a Carrier of Stem Cells in Tissue Regeneration: A Systematic Review of Pre-Clinical Research. Cells Tissues Organs 2021, 210, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Piao, Y.; Liu, Q.; Yang, X. Platelet-rich plasma-derived extracellular vesicles: A superior alternative in regenerative medicine? Cell Prolif. 2021, 54, e13123. [Google Scholar] [CrossRef]
- Antich-Rossello, M.; Forteza-Genestra, M.A.; Monjo, M.; Ramis, J.M. Platelet-Derived Extracellular Vesicles for Regenerative Medicine. Int. J. Mol. Sci. 2021, 22, 8580. [Google Scholar] [CrossRef] [PubMed]
- Eustes, A.S.; Dayal, S. The Role of Platelet-Derived Extracellular Vesicles in Immune-Mediated Thrombosis. Int. J. Mol. Sci. 2022, 23, 7837. [Google Scholar] [CrossRef]
- Abels, E.R.; Breakefield, X.O. Introduction to Extracellular Vesicles: Biogenesis, RNA Cargo Selection, Content, Release, and Uptake. Cell. Mol. Neurobiol. 2016, 36, 301–312. [Google Scholar] [CrossRef]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Chargaff, E.; West, R. The biological significance of the thromboplastic protein of blood. J. Biol. Chem. 1946, 166, 189–197. [Google Scholar] [CrossRef]
- Wolf, P. The nature and significance of platelet products in human plasma. Br. J. Haematol. 1967, 13, 269–288. [Google Scholar] [CrossRef]
- Bahmani, L.; Ullah, M. Different Sourced Extracellular Vesicles and Their Potential Applications in Clinical Treatments. Cells 2022, 11, 1989. [Google Scholar] [CrossRef]
- Gasecka, A.; Nieuwland, R.; Siljander, P.R.M. Platelet-Derived Extracellular Vesicles. In Platelets; Academic Press: Cambridge, MA, USA, 2019; pp. 401–416. [Google Scholar]
- Palazzolo, S.; Canzonieri, V.; Rizzolio, F. The history of small extracellular vesicles and their implication in cancer drug resistance. Front. Oncol. 2022, 12, 948843. [Google Scholar] [CrossRef]
- Torreggiani, E.; Perut, F.; Roncuzzi, L.; Zini, N.; Baglio, S.R.; Baldini, N. Exosomes: Novel effectors of human platelet lysate activity. Eur. Cells Mater. 2014, 28, 137–151, discussion 151. [Google Scholar] [CrossRef] [PubMed]
- Momen-Heravi, F.; Getting, S.J.; Moschos, S.A. Extracellular vesicles and their nucleic acids for biomarker discovery. Pharmacol. Ther. 2018, 192, 170–187. [Google Scholar] [CrossRef] [PubMed]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef]
- Hessvik, N.P.; Llorente, A. Current knowledge on exosome biogenesis and release. Cell. Mol. Life Sci. 2018, 75, 193–208. [Google Scholar] [CrossRef]
- Ramirez-Garrastacho, M.; Bajo-Santos, C.; Line, A.; Martens-Uzunova, E.S.; de la Fuente, J.M.; Moros, M.; Soekmadji, C.; Tasken, K.A.; Llorente, A. Extracellular vesicles as a source of prostate cancer biomarkers in liquid biopsies: A decade of research. Br. J. Cancer 2022, 126, 331–350. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Li, S.; Zhang, G.; Xu, H.H.K.; Zhang, K.; Bai, Y. New frontiers of oral sciences: Focus on the source and biomedical application of extracellular vesicles. Front. Bioeng. Biotechnol. 2022, 10, 1023700. [Google Scholar] [CrossRef]
- Golbach, L.A.; Portelli, L.A.; Savelkoul, H.F.; Terwel, S.R.; Kuster, N.; de Vries, R.B.; Verburg-van Kemenade, B.M. Calcium homeostasis and low-frequency magnetic and electric field exposure: A systematic review and meta-analysis of in vitro studies. Environ. Int. 2016, 92–93, 695–706. [Google Scholar] [CrossRef]
- Liu, X.; Wang, L.; Ma, C.; Wang, G.; Zhang, Y.; Sun, S. Exosomes derived from platelet-rich plasma present a novel potential in alleviating knee osteoarthritis by promoting proliferation and inhibiting apoptosis of chondrocyte via Wnt/beta-catenin signaling pathway. J. Orthop. Surg. Res. 2019, 14, 470. [Google Scholar] [CrossRef]
- Otahal, A.; Kramer, K.; Kuten-Pella, O.; Weiss, R.; Stotter, C.; Lacza, Z.; Weber, V.; Nehrer, S.; De Luna, A. Characterization and Chondroprotective Effects of Extracellular Vesicles from Plasma- and Serum-Based Autologous Blood-Derived Products for Osteoarthritis Therapy. Front. Bioeng. Biotechnol. 2020, 8, 584050. [Google Scholar] [CrossRef] [PubMed]
- Otahal, A.; Kramer, K.; Kuten-Pella, O.; Moser, L.B.; Neubauer, M.; Lacza, Z.; Nehrer, S.; De Luna, A. Effects of Extracellular Vesicles from Blood-Derived Products on Osteoarthritic Chondrocytes within an Inflammation Model. Int. J. Mol. Sci. 2021, 22, 7224. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, X.; Chen, J.; Qian, D.; Gao, P.; Qin, T.; Jiang, T.; Yi, J.; Xu, T.; Huang, Y.; et al. Exosomes derived from platelet-rich plasma administration in site mediate cartilage protection in subtalar osteoarthritis. J. Nanobiotechnology 2022, 20, 56. [Google Scholar] [CrossRef] [PubMed]
- Rui, S.; Yuan, Y.; Du, C.; Song, P.; Chen, Y.; Wang, H.; Fan, Y.; Armstrong, D.G.; Deng, W.; Li, L. Comparison and Investigation of Exosomes Derived from Platelet-Rich Plasma Activated by Different Agonists. Cell Transplant. 2021, 30, 9636897211017833. [Google Scholar] [CrossRef]
- Saumell-Esnaola, M.; Delgado, D.; Garcia Del Cano, G.; Beitia, M.; Salles, J.; Gonzalez-Burguera, I.; Sanchez, P.; Lopez de Jesus, M.; Barrondo, S.; Sanchez, M. Isolation of Platelet-Derived Exosomes from Human Platelet-Rich Plasma: Biochemical and Morphological Characterization. Int. J. Mol. Sci. 2022, 23, 2861. [Google Scholar] [CrossRef]
- Stoner, S.A.; Duggan, E.; Condello, D.; Guerrero, A.; Turk, J.R.; Narayanan, P.K.; Nolan, J.P. High sensitivity flow cytometry of membrane vesicles. Cytom. Part A 2016, 89, 196–206. [Google Scholar] [CrossRef]
- Yuana, Y.; Koning, R.I.; Kuil, M.E.; Rensen, P.C.; Koster, A.J.; Bertina, R.M.; Osanto, S. Cryo-electron microscopy of extracellular vesicles in fresh plasma. J. Extracell. Vesicles 2013, 2, 21494. [Google Scholar] [CrossRef]
- Zhang, W.; Dong, X.; Wang, T.; Kong, Y. Exosomes derived from platelet-rich plasma mediate hyperglycemia-induced retinal endothelial injury via targeting the TLR4 signaling pathway. Exp. Eye Res. 2019, 189, 107813. [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, H.; Kong, Y. Exosomes derived from platelet-rich plasma activate YAP and promote the fibrogenic activity of Muller cells via the PI3K/Akt pathway. Exp. Eye Res. 2020, 193, 107973. [Google Scholar] [CrossRef]
- Dai, Z.; Xia, C.; Zhao, T.; Wang, H.; Tian, H.; Xu, O.; Zhu, X.; Zhang, J.; Chen, P. Platelet-derived extracellular vesicles ameliorate intervertebral disc degeneration by alleviating mitochondrial dysfunction. Mater. Today Bio 2023, 18, 100512. [Google Scholar] [CrossRef]
- Xu, J.; Xie, G.; Yang, W.; Wang, W.; Zuo, Z.; Wang, W. Platelet-rich plasma attenuates intervertebral disc degeneration via delivering miR-141-3p-containing exosomes. Cell Cycle 2021, 20, 1487–1499. [Google Scholar] [CrossRef] [PubMed]
- Catitti, G.; Cufaro, M.C.; De Bellis, D.; Cicalini, I.; Vespa, S.; Tonelli, F.; Miscia, G.; Secondi, L.; Simeone, P.; De Laurenzi, V.; et al. Extracellular Vesicles in Regenerative Processes Associated with Muscle Injury Recovery of Professional Athletes Undergoing Sub Maximal Strength Rehabilitation. Int. J. Mol. Sci. 2022, 23, 14913. [Google Scholar] [CrossRef] [PubMed]
- Iyer, S.R.; Scheiber, A.L.; Yarowsky, P.; Henn, R.F. 3rd.; Otsuru, S.; Lovering, R.M. Exosomes Isolated from Platelet-Rich Plasma and Mesenchymal Stem Cells Promote Recovery of Function After Muscle Injury. Am. J. Sports Med. 2020, 48, 2277–2286. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.C.; Tao, S.C.; Yin, W.J.; Qi, X.; Yuan, T.; Zhang, C.Q. Exosomes derived from platelet-rich plasma promote the re-epithelization of chronic cutaneous wounds via activation of YAP in a diabetic rat model. Theranostics 2017, 7, 81–96. [Google Scholar] [CrossRef]
- Xu, Y.; Lin, Z.; He, L.; Qu, Y.; Ouyang, L.; Han, Y.; Xu, C.; Duan, D. Platelet-Rich Plasma-Derived Exosomal USP15 Promotes Cutaneous Wound Healing via Deubiquitinating EIF4A1. Oxidative Med. Cell. Longev. 2021, 2021, 9674809. [Google Scholar] [CrossRef]
- Dong, B.; Liu, X.; Li, J.; Wang, B.; Yin, J.; Zhang, H.; Liu, W. Berberine Encapsulated in Exosomes Derived from Platelet-Rich Plasma Promotes Chondrogenic Differentiation of the Bone Marrow Mesenchymal Stem Cells via the Wnt/β-Catenin Pathway. Biol. Pharm. Bull. 2022, 45, 1444–1451. [Google Scholar] [CrossRef]
- Bagio, D.A.; Julianto, I.; Margono, A.; Suprastiwi, E. Analysis of Thrombin-Activated Platelet-Derived Exosome (T-aPDE) Potential for Dental Pulp Regeneration: In-Vitro Study. Eur. J. Dent. 2023, 17, 173–182. [Google Scholar] [CrossRef]
- Nilforoushzadeh, M.A.; Aghdami, N.; Taghiabadi, E. Effects of Adipose-Derived Stem Cells and Platelet-Rich Plasma Exosomes on The Inductivity of Hair Dermal Papilla Cells. Cell J. 2021, 23, 576–583. [Google Scholar]
- Tao, S.C.; Yuan, T.; Rui, B.Y.; Zhu, Z.Z.; Guo, S.C.; Zhang, C.Q. Exosomes derived from human platelet-rich plasma prevent apoptosis induced by glucocorticoid-associated endoplasmic reticulum stress in rat osteonecrosis of the femoral head via the Akt/Bad/Bcl-2 signal pathway. Theranostics 2017, 7, 733–750. [Google Scholar] [CrossRef]
- Chahla, J.; Cinque, M.E.; Piuzzi, N.S.; Mannava, S.; Geeslin, A.G.; Murray, I.R.; Dornan, G.J.; Muschler, G.F.; LaPrade, R.F. A Call for Standardization in Platelet-Rich Plasma Preparation Protocols and Composition Reporting: A Systematic Review of the Clinical Orthopaedic Literature. J. Bone Jt. Surg. 2017, 99, 1769–1779. [Google Scholar] [CrossRef]
- Sundman, E.A.; Cole, B.J.; Fortier, L.A. Growth factor and catabolic cytokine concentrations are influenced by the cellular composition of platelet-rich plasma. Am. J. Sports Med. 2011, 39, 2135–2140. [Google Scholar] [CrossRef] [PubMed]
- Anitua, E.; Zalduendo, M.; Troya, M.; Padilla, S.; Orive, G. Leukocyte inclusion within a platelet rich plasma-derived fibrin scaffold stimulates a more pro-inflammatory environment and alters fibrin properties. PLoS ONE 2015, 10, e0121713. [Google Scholar] [CrossRef] [PubMed]
- Auber, M.; Svenningsen, P. An estimate of extracellular vesicle secretion rates of human blood cells. J. Extracell. Biol. 2022, 1, e46. [Google Scholar] [CrossRef]
- Alberro, A.; Iparraguirre, L.; Fernandes, A.; Otaegui, D. Extracellular Vesicles in Blood: Sources, Effects, and Applications. Int. J. Mol. Sci. 2021, 22, 8163. [Google Scholar] [CrossRef] [PubMed]
- Tissot, J.-D.; Canellini, G.; Rubin, O.; Angelillo-Scherrer, A.; Delobel, J.; Prudent, M.; Lion, N. Blood microvesicles: From proteomics to physiology. Transl. Proteom. 2013, 1, 38–52. [Google Scholar] [CrossRef]
- Bister, N.; Pistono, C.; Huremagic, B.; Jolkkonen, J.; Giugno, R.; Malm, T. Hypoxia and extracellular vesicles: A review on methods, vesicular cargo and functions. J. Extracell. Vesicles 2020, 10, e12002. [Google Scholar] [CrossRef]
- Brahmer, A.; Neuberger, E.; Esch-Heisser, L.; Haller, N.; Jorgensen, M.M.; Baek, R.; Mobius, W.; Simon, P.; Kramer-Albers, E.M. Platelets, endothelial cells and leukocytes contribute to the exercise-triggered release of extracellular vesicles into the circulation. J. Extracell. Vesicles 2019, 8, 1615820. [Google Scholar] [CrossRef]
- Milioli, M.; Ibáñez-Vea, M.; Sidoli, S.; Palmisano, G.; Careri, M.; Larsen, M.R. Quantitative proteomics analysis of platelet-derived microparticles reveals distinct protein signatures when stimulated by different physiological agonists. J. Proteom. 2015, 121, 56–66. [Google Scholar] [CrossRef]
- Shai, E.; Rosa, I.; Parguina, A.F.; Motahedeh, S.; Varon, D.; Garcia, A. Comparative analysis of platelet-derived microparticles reveals differences in their amount and proteome depending on the platelet stimulus. J. Proteom. 2012, 76, 287–296. [Google Scholar] [CrossRef]
- Suades, R.; Padro, T.; Vilahur, G.; Badimon, L. Platelet-released extracellular vesicles: The effects of thrombin activation. Cell. Mol. Life Sci. 2022, 79, 190. [Google Scholar] [CrossRef]
- Janouskova, O.; Herma, R.; Semeradtova, A.; Poustka, D.; Liegertova, M.; Hana Auer, M.; Maly, J. Conventional and Nonconventional Sources of Exosomes-Isolation Methods and Influence on Their Downstream Biomedical Application. Front. Mol. Biosci. 2022, 9, 846650. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.; Im, H.; Castro, C.M.; Breakefield, X.; Weissleder, R.; Lee, H. New Technologies for Analysis of Extracellular Vesicles. Chem. Rev. 2018, 118, 1917–1950. [Google Scholar] [CrossRef] [PubMed]
- Gelibter, S.; Marostica, G.; Mandelli, A.; Siciliani, S.; Podini, P.; Finardi, A.; Furlan, R. The impact of storage on extracellular vesicles: A systematic study. J. Extracell. Vesicles 2022, 11, e12162. [Google Scholar] [CrossRef]
- van de Wakker, S.I.; van Oudheusden, J.; Mol, E.A.; Roefs, M.T.; Zheng, W.; Gorgens, A.; El Andaloussi, S.; Sluijter, J.P.G.; Vader, P. Influence of short term storage conditions, concentration methodsand excipients on extracellular vesicle recovery and function. Eur. J. Pharm. Biopharm. 2022, 170, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Deville, S.; Berckmans, P.; Van Hoof, R.; Lambrichts, I.; Salvati, A.; Nelissen, I. Comparison of extracellular vesicle isolation and storage methods using high-sensitivity flow cytometry. PLoS ONE 2021, 16, e0245835. [Google Scholar] [CrossRef]
- Sivanantham, A.; Jin, Y. Impact of Storage Conditions on EV Integrity/Surface Markers and Cargos. Life 2022, 12, 697. [Google Scholar] [CrossRef]
- Evtushenko, E.G.; Bagrov, D.V.; Lazarev, V.N.; Livshits, M.A.; Khomyakova, E. Adsorption of extracellular vesicles onto the tube walls during storage in solution. PLoS ONE 2020, 15, e0243738. [Google Scholar] [CrossRef]
- Gorgens, A.; Corso, G.; Hagey, D.W.; Jawad Wiklander, R.; Gustafsson, M.O.; Felldin, U.; Lee, Y.; Bostancioglu, R.B.; Sork, H.; Liang, X.; et al. Identification of storage conditions stabilizing extracellular vesicles preparations. J. Extracell. Vesicles 2022, 11, e12238. [Google Scholar] [CrossRef]
- Szatanek, R.; Baj-Krzyworzeka, M.; Zimoch, J.; Lekka, M.; Siedlar, M.; Baran, J. The Methods of Choice for Extracellular Vesicles (EVs) Characterization. Int. J. Mol. Sci. 2017, 18, 1153. [Google Scholar] [CrossRef]
- Imanbekova, M.; Suarasan, S.; Lu, Y.; Jurchuk, S.; Wachsmann-Hogiu, S. Recent advances in optical label-free characterization of extracellular vesicles. Nanophotonics 2022, 11, 2827–2863. [Google Scholar] [CrossRef]
- Campos-Silva, C.; Suarez, H.; Jara-Acevedo, R.; Linares-Espinos, E.; Martinez-Pineiro, L.; Yanez-Mo, M.; Vales-Gomez, M. High sensitivity detection of extracellular vesicles immune-captured from urine by conventional flow cytometry. Sci. Rep. 2019, 9, 2042. [Google Scholar] [CrossRef]
- Anitua, E.; Zalduendo, M.; Troya, M.; Tierno, R.; Alkhraisat, M.H. The inclusion of leukocytes into platelet rich plasma reduces scaffold stability and hinders extracellular matrix remodelling. Ann. Anat.-Anat. Anz. 2022, 240, 151853. [Google Scholar] [CrossRef] [PubMed]
- Amiri, A.; Bagherifar, R.; Ansari Dezfouli, E.; Kiaie, S.H.; Jafari, R.; Ramezani, R. Exosomes as bio-inspired nanocarriers for RNA delivery: Preparation and applications. J. Transl. Med. 2022, 20, 125. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Zhao, T.; Song, N.; Pan, K.; Yang, Y.; Zhu, X.; Chen, P.; Zhang, J.; Xia, C. Platelets and platelet extracellular vesicles in drug delivery therapy: A review of the current status and future prospects. Front. Pharmacol. 2022, 13, 1026386. [Google Scholar] [CrossRef]
- Xu, K.; Liu, Q.; Wu, K.; Liu, L.; Zhao, M.; Yang, H.; Wang, X.; Wang, W. Extracellular vesicles as potential biomarkers and therapeutic approaches in autoimmune diseases. J. Transl. Med. 2020, 18, 432. [Google Scholar] [CrossRef]
- Wu, W.C.; Song, S.J.; Zhang, Y.; Li, X. Role of Extracellular Vesicles in Autoimmune Pathogenesis. Front. Immunol. 2020, 11, 579043. [Google Scholar] [CrossRef] [PubMed]
- Jansen, F.; Nickenig, G.; Werner, N. Extracellular Vesicles in Cardiovascular Disease: Potential Applications in Diagnosis, Prognosis, and Epidemiology. Circ. Res. 2017, 120, 1649–1657. [Google Scholar] [CrossRef]
- Mallia, A.; Gianazza, E.; Zoanni, B.; Brioschi, M.; Barbieri, S.S.; Banfi, C. Proteomics of Extracellular Vesicles: Update on Their Composition, Biological Roles and Potential Use as Diagnostic Tools in Atherosclerotic Cardiovascular Diseases. Diagnostics 2020, 10, 843. [Google Scholar] [CrossRef]
- Patel, S.; Guo, M.K.; Abdul Samad, M.; Howe, K.L. Extracellular vesicles as biomarkers and modulators of atherosclerosis pathogenesis. Front. Cardiovasc. Med. 2023, 10, 1202187. [Google Scholar] [CrossRef]
- Valencia, J.; Ferreira, M.; Merino-Torres, J.F.; Marcilla, A.; Soriano, J.M. The Potential Roles of Extracellular Vesicles as Biomarkers for Parkinson’s Disease: A Systematic Review. Int. J. Mol. Sci. 2022, 23, 11508. [Google Scholar] [CrossRef]
- Vassileff, N.; Cheng, L.; Hill, A.F. Extracellular vesicles—Propagators of neuropathology and sources of potential biomarkers and therapeutics for neurodegenerative diseases. J. Cell. Sci. 2020, 133, jcs243139. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Wang, Y.; Zheng, J.C. Extracellular vesicles, from the pathogenesis to the therapy of neurodegenerative diseases. Transl. Neurodegener. 2022, 11, 53. [Google Scholar] [CrossRef] [PubMed]
- Vozel, D.; Bozic, D.; Jeran, M.; Jan, Z.; Pajnic, M.; Paden, L.; Steiner, N.; Kralj-Iglic, V.; Battelino, S. Autologous Platelet- and Extracellular Vesicle-Rich Plasma Is an Effective Treatment Modality for Chronic Postoperative Temporal Bone Cavity Inflammation: Randomized Controlled Clinical Trial. Front. Bioeng. Biotechnol. 2021, 9, 677541. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ref. | Field of Application | Study Type | Experimental System | Comparison Groups | [Exosomes] | Methods | Summary Results |
|---|---|---|---|---|---|---|---|
| Liu et al., 2019 [31] | Osteoarthritis | In vitro | Primary rabbit chondrocytes | Group I: PBS Group II: IL-1β Group III: IL-1β + PRP-exos Group IV: IL-1β + PRP-As | 5, 50 µg/mL | Elisa, proliferation, apoptosis, migration, scratch wound assay, western blotting | PRP-Exos and PRP-As both inhibited TNF-α release. PRP-Exos could significantly decrease the apoptotic rate of OA chondrocytes compared with PRP-As. PRP-Exos promoted proliferation and migration to a larger extent than PRP-As. The potential mechanism might be through activation of the Wnt/β-catenin signaling pathway. β-catenin, RUNX2, and Wnt5a typical increase in IL-1β-treated chondrocytes could accordingly reverse by both PRP-Exos and PRP-As, although the former performed better than the latter. In vivo, both PRP-Exos and PRP-As prevented the progression of OA, the effect of PRP-Exos being significatively more effective. |
| In vivo | Rabbit knee OA model | Group I: Control Group II: OA Group III: OA + PRP-Exos Group IV: OA + PRP-As | 100 µg/mL | Histological analysis, immunohistochemistry, the OARSI score system | |||
| Otahal et al., 2020 [32] | Osteoarthritis | In vitro | Human chondrocytes from OA cartilage | Group I: CPRP Group II: HypACT Group III: CPRP-EVs Group IV: HypACT-EVs Group V: 10% FCS (Control) | hypACT EV: 1.42 × 109 ± 2.12 × 106 EVs per well CPRP EV:1.42 × 109 ± 2.95 × 107 EVs per well | Western blot, RT-qPCR, ELISA | The total EV concentration was significantly lower in hypACT than in CPRP. Around 84% of total EVs originated from platelets in hypACT, whereas platelet EVs constituted a 44% proportion of total EVs in CPRP. The clotting for hypACT obtention might be responsible for such differences between plasma (PRP) and serum (hypACT) derivatives. EV treatment of OA patient-derived chondrocytes enhanced expression of both anabolic (Type II collagen, SOX9, and ACAN) and catabolic (MMP3) chondroprotective markers, compared to full blood products. HypACT EVs prevented Type I collagen expression. CPRP EVs might provide beneficial anti-inflammatory effects in acute OA, whereas hypACT EVs might be more suitable to drive chondrogenesis and cartilage regeneration in chronic OA. |
| Otahal et al., 2021 [33] | Osteoarthritis | In vitro | Human primary OA chondrocytes. Human primary monocytes and THP-1 were differentiated and activated into M1 macrophages. Coculture. | Group I: CPRP Group II: HypACT Group III: CPRP-EVs Group IV: HypACT-EVs Group V: 10% FCS (Control) | hypACT EV: 1.91 × 108 ± 1.09 × 108 EVs CPRP EV: 1.91 × 108 ± 1.65 × 108 EVs of resuspended EV pellets added to 2 mL co-culture medium | RT-qPCR, ELISA | Mode sizes were similar between EVs from CPRP and hypACT. Higher concentrations of EVs were obtained from CPRP compared to hypACT. EV treatment was associated with a reduction in proinflammatory cytokine levels compared to blood product treatment. EV treatment increased Type II collagen expression while lowering TNF-α and IL-1β secretion. This could indicate either that the EV treatment replicates the effect of the complete blood product or that the EV concentration from blood products was too low to elicit therapeutically relevant changes. This did not exclude the possibility that EVs are therapeutically relevant agents in blood product therapy. |
| Zhang et al., 2022 [34] | Subtalar Osteoarthritis | In vitro | mBMSCs and murine chondrocytes | Group I: Control Group II: Gel Group III: Exo-gel | The conditioned medium from Exo-Gel (200 μg Exo incorporated in 100 μL thermossensitive Gel) | Uptake of Exo, viability, cell proliferation, migration assay, RTqPCR, chondrocyte apoptosis, cell fluorescence analysis, Western blot. | PRP-Exo incorporated into gel (Exo-gel) could release and promote the proliferation and migration of mBMSCs and chondrocytes, enhance the chondrogenic differentiation of mBMSCs, and inhibit inflammation-induced chondrocyte degeneration. PRP-Exo had positive regulation of mBMSCs function and played a protective role against IL-1β-induced apoptosis and degeneration of chondrocytes in vivo. Application of this system allowed retention of EVs (Exo) in the ankle compared to Exo-only injection and maintained the local concentration of Exo. Together, Exo-Gel had effective effects on suppressing cartilage degeneration and the development of STOA. |
| In vivo | STOA model | Group I: PBS Group II: Gel Group III: Exo Group IV: Exo-Gel | 4 µg/2 µL (2000 µg/mL) | Histomorpholgical detection, immunohistochemical analysis, TUNEL staining | |||
| Rui et al., 2021 [35] | Tissue regeneration | In vitro | HUVECs | Group I: PRP-AS Group II: PRP-Exos + saline Group III: PRP-Exos + thrombin Group IV: PRP-Exos + Ca2+ Group V: PRP-Exos + mixture | 50 µg/mL | Exosomes internalization, cell proliferation, migration, matrigel tube formation, Western blot | The size of calcium-activated PRP-Exos was larger than the thrombin-activated group and the mixture-activated group. PRP activated by the mixture released the highest concentration of exosomes. All the activation methods were suitable for PRP-Exos isolation, calcium gluconate alone was found to be weaker. The protein levels of VEGF, PDGFBB, bFGF, TGF-β in the mixture-activated PRP-Exos group were higher than in other groups. PRP-Exos yielded after thrombin and calcium gluconate together were found to contain more cytokines than the other groups. PRP-Exos harvested after activation by thrombin and calcium gluconate together could more significantly promote HUVECs proliferation, migration, and formation of vessel-like via the AKT ERK signal pathway, compared with other groups. |
| Saumell-Esnaola et al., 2022 [36] | Tissue regeneration | In vitro | N/A | Group I: PLT Group II: PLT-Ca2+ Group III: PLT-Exos Group IV: PLT-Exos-Ca2+ Group V: PPP Group VI: PRP | N/A | Semiquantitative measurement of 105 analyte levels, including cytokines, chemokines, growth factors, and other soluble proteins | The calcium activation of PRP promoted the release of highly purified platelet-derived exosomes, showing a concordant size and morphology and the absence of contaminants from other cellular compartments. Calcium was proved to alter the cytokine cargo expression profile of PLT-Exos-Ca2+ in relation to exosomes isolated from non-activated platelets. Although PRP calcium activation promoted exosome release, its net contribution to the total PRP might be minimal, in view of the low yield of exosome protein content relative to the total in PRP. |
| Stoner et al., 2016 [37] | Tissue regeneration | In vitro | N/A | N/A | N/A | HSFC/VFC | Treatment of platelet-rich plasma with calcium ionophore resulted in an increase in the fraction of annexin V and CD61-positive EVs. Vesicle flow cytometry using fluorescence-based detection of EVs had the potential to realize the potential of cell-derived membrane vesicles as functional biomarkers for a variety of applications. |
| Yuana et al., 2013 [38] | Tissue regeneration | In vitro | N/A | Group I: PRP Group II: PPP Group III: PFP Group IV: PRP-EVs Group V: PPP-EVs Group VI: PFP-EVs | N/A | cryo-EM/cryo-ET | EVs isolated from fresh plasma were highly heterogeneous in morphology and size. EV constituted only a very small fraction of all particles present in fresh plasma isolated from the blood of fasted healthy volunteers, whereas the majority were lipoprotein particles. Cryo-EM was a powerful technique that enabled the characterization of EV in fresh plasma. |
| Zhang et al., 2019 [39] | Diabetic retinopathy | In vitro | HRECs | Group I: ctrl Group II: ctrl-PRP-Exos Group III: DM-PRP-Exos TLR4 assay: Group I: without PRP-Exos (Ctrl) Group II: with normal PRP-Exos Group III: HG-PRP-Exos Group IV: HG-PRP-Exos + TAK-242 CXCL10 assay: Group I: without PRP-Exos (Ctrl) Group II: HG-PRP-Exos Group III: HG-PRP-Exos + anti-CXCL10 | 500 μg PRP-Exos (RNA concentration) suspended in 500 μL PBS (1000 µg/mL) | Immunohistochemical measurements, western blot, ROS | The level of circulating PRP-Exos was greatly elevated in diabetic rats. PRP-Exos mediated hyperglycemia-induced retina endothelia injury via TLR4 signaling pathway. PRP-Exos can activate the TLR4 pathway by promoting an increase in TLR4 and its downstream proteins in vitro and in vivo. CXCL10 was a major effector of PRP-Exo-derived retinal endothelial damage; thus, blocking PRP-Exo-derived CXCL10 may be a novel therapeutic approach for DR. |
| In vivo | Diabetes mellitus induced in rats | Group I: normal controls were treated with vehicle (PBS) Group II: DM (diabetic rats were treated with vehicle) Group III: DM + TAK. | NS | Immunohistochemical measurements, western blot, ROS, measurement of blood-retinal barrier (BRB) breakdown | |||
| Zhang et al., 2020 [40] | Diabetic retinopathy/Retinal fibrogenesis | In vitro | hMCs | Group I: Nor-PRP-Exos Group II: DM-PRP-Exos Group III: Nor-PL (control) Group IV: DM-PL (control) | 500 μg PRP-Exos (RNA concentration) suspended in 500 μL PBS (1000 µg/mL) | Internalization, cell proliferation, migration, Elisa, Western blot, Immunohistochemical analysis | The concentration of exosomes was increased in the DM-PRP-Exos group compared with the Nor-PRP-Exos group. Analysis of the exosome cargo showed that the levels of PDGF, bFGF, and TGF-β were elevated in DM-PRP-Exos compared with those in PL or Nor-PRP-Exos. DM-PRP-Exos significantly increased the proliferative and migratory ability of hMCs and the expression of CTGF and fibronectin. DM-PRP-Exos contributed to activating YAP and promoting the fibrogenic activity of Müller cells via the PI3K/Akt pathway. |
| Dai et al., 2023 [41] | Intervertebral disc degeneration | In vitro | NP cells from rats | Group I: Normal control Group II: H2O2 Group III: PLTs (H2O2 + 100 μg/mL platelets) Group IV: PEVs (H2O2 + 100 μg/mL PEVs) | 100 μg/mL | Cytotoxicity and proliferation, cytokine detection, ROS detection, apoptosis, SA-β-gal staining, PEVs uptake and mitochondrial localization, qPCR, western blot, immunofluorescence staining, mitochondrial function, ATP detection, lactic acid detection | A significant increase in IL-1β, IL-6, and TNFα production in platelets after thrombin addition was detected. PEVs did not release these pro-inflammatory factors. PEVs inhibited apoptosis and senescence in NP cells. PEVs could restore impaired mitochondrial function, reduce oxidative stress, and restore cell metabolism by regulating the SIRT1-peroxisome PGC1α-TFAM pathway. In rat models, PEVs retarded the progression of IVDD by reducing oxidative stress and inflammatory response. |
| In vivo | A rat IVDD model | Group I: NC group Group II: Puncture + vehicle (PBS) Group III: Puncture + PLTs Group IV: Puncture + PEVs | 100 μg/mL | Micro-CT and MRI, histological analysis, immunofluorescence staining, systemic toxicity | |||
| Xu et al., 2021 [42] | Intervertebral disc degeneration | In vitro | NP cells and HEK293T cells | Group I: Control Group II: H2O2 Group III: H2O2 + PRP-exo Group IV: H2O2 + NAC | 50 µg/mL | RTqPCR, Western blot, Elisa, ROS, cell apoptosis, cell proliferation and viability, Dual-luciferase reporter gene system assay, RNA pull-down assay | Both PRP-exo and NAC suppressed ROS generation and pro-inflammatory cytokines in NP cells treated with H2O2. PRP-exo and NAC ameliorated H2O2- induced cell apoptosis in NP cells. PRP-exo reversed the detrimental effects of H2O2 treatment on NP cells. The Keap1-Nrf2 pathway could be activated by PRP-exo in the NP cells. PRP-exo delivered miR-141-3p to degrade Keap1, leading to the release of Nrf2 from the Keap1-Nrf2 complex, which further translocated from cytoplasm to nucleus to exert its anti-oxidant effects, resulting in the attenuation of IVD degeneration. |
| Catitti et al., 2022 [43] | Muscle injury | In vitro | N/A | PRP from athletes recovering from injuries | N/A | EV Protein Cargo Detection by Label-Free Proteomics | Leukocyte-derived EVs were the most represented EV subpopulation within PRP-EVs, followed by platelet-derived EVs and EVs from endothelium. There were 105 proteins identified, mostly classified in the “defense and immunity” biological function and related to “vesicle-mediated transport” and “wound healing”. |
| Iyer et al., 2020 [44] | Muscle injury | In vivo | Rat bone marrow–derived MSCs/Animal model of muscle strain injury | Group I: Saline Group II: PRP exosomes Group III: MSC exosomes | 1 × 108 exosomes resuspended in 50 µL of PBS | Recovery of muscle function, histological assay to detect myogenesis, RT-PCR | Both PRP-exos and MSC-exos accelerated recovery of contractile function over the saline-treated group. A significant increase in CNFs was observed with both types of exosome groups by day 15. Muscles treated with PRP-exos had increased expression of Myogenin gene. Exosomes derived from PRP or MSCs could facilitate recovery after a muscle strain injury in a small-animal model. |
| Guo et al., 2017 [45] | Wound healing | In vitro | HMEC-1 and primary dermal fibroblasts treated with Y-27632 2HCl, or transfected with shYAP or S127A | Group I: PRP-AS Group II: PRP-Exos. | 5, 50 µg/mL | Cell proliferation, PRP-Exos-loaded sodium alginate hidrogel, PRP-Exos release, cell migration, capillary-like construction activity, YAP nuclear localization, gene expression analysis, and western blotting | PRP-Exos increased the proliferation and migration of HMEC-1 cells and fibroblasts and tube formation to a greater extent than PRP. The wounds treated with PRP-Exos closed significantly faster. Higher blood vessel area, blood vessel number, and longer neo-epithelium in the PRP-Exos-treated defects were observed. Exosomes released by PRP may contribute to PRP-induced angiogenesis through activation of Erk and Akt signaling pathways, and PRP-induced re-epithelialization may be triggered by activation of YAP. |
| In vivo | Diabetic rat model | Group I: Control Group II: SAH Group III: PRP Group IV: PRP-Exos | It was calculated on the basis of the determined PRP-AS protein concentration and mixed with SAH | Wound size, microCT, histological analysis, IHC and IF methods | |||
| Xu et al., 2021 [46] | Wound healing | In vitro | HaCaT | Group I: Control (PBS) Group II: PRP Group III: PRP-Exos. | N.S. | Internalization of exosomes, cell proliferation, wound healing, migration, cell cycle analysis, Western blot, qPCR | PRP-Exo treatment enhanced in vitro keratinocyte responses. PRP-Exo treatment was superior to PRP treatment as a means of enhancing wound healing associated with increased USP15 in wound tissues relative to PBS or PRP administration. |
| In vivo | Murine Cutaneous Wound Model | Group I: PBS Group II: PRP-Exos Group III: siRNA-NC Group IV: siRNA-USP15 Group V: siRNAUSP15+ PRP-Exos | PRP-exos: 100 μg PRP-Exos in 100 μL PBS (1000 µg/mL); siRNA-USP15 + PRP-Exos: 10μmol/L | Wound healing, histological analysis, and Immunohistochemical Staining | |||
| Dong et al., 2022 [47] | Cartilage regeneration | In vitro | BMSCs from rats | Group I: PRP-exos Group II: Ber Group III: Exos-Ber Group IV: Control (PBS or medium) | 5, 25 and 50 µg/mL | Proliferation, migration, chondrogenesis | PRP-exos or Exos-Ber at 25 μg/mL led to significantly enhanced proliferation of BMSCs. PRP-exos, Ber, and Exos-Ber could significantly promote the chondrogenic differentiation of BMSCs. The highest upregulation was observed with Exos-Ber. PRP-exos and Exos-Ber could significantly promote the migration of BMSCs. The proteins Collagen II, SOX9, and ACAN were upregulated in BMSCs exposed to Ber, PRP-exos, or Exos-Ber. The greatest upregulation was observed in cells treated with Exos-Ber. Exos-Ber could promote BMSCs chondrogenic differentiation via the Wnt/β-catenin signaling pathway. |
| Bagio et al., 2023 [48] | Dental Pulp Regeneration | In vitro | hDPSCs | Group I: no FBS Group II: 10% FBS Group III: 10% PRP-T Group IV: 0.5% T-aPDE Group V: 1% T-aPDE Group VI: 5% T-aPDE. | 0.5%/1%/5% | Viability assay/Migration/VEGF-A Expression | The 5% T-aPDE group could produce hDPSCs with a superior mean viability rate compared with the other conditioned media culture groups, and it had the best migration activity of all the groups. The VEGF-A amount of hDPSCs escalated more significantly 24 to 72 h after they were cultured in 5% T-aPDE. |
| Nilforoushzadeh et al., 2021 [49] | Hair loss | In vitro | DPCs and ORSCs | Group I: Control (serum-free culture medium) Group II: hASCs-Exo Group III: PRP-Exo | 25, 50 and 100 µg/mL | Internalization of exosomes, cell scratch, proliferation, SEM, RT-PCR | Exosomes could enter the DPCs cytoplasm and be localized in the perinuclear area. The culture of human DPCs with ASC-Exo exhibited significantly increased migration, proliferation, and hair inductivity compared to other experimental groups. When 100 μg/mL ASCs-Exo was compared to the same concentration of PRP-Exo, the former significantly promoted DP proliferation and migration, as well as overregulation of ALP, Versican, and α-SMA proteins. |
| Tao et al., 2017 [50] | ONFH | In vitro | MC3T3-E1 cells, HMEC-1 and primary BMSCs/DEX-treated in vitro cell model | Group I: Control Group II: DEX Group III: DEX + PRP-exos Group IV: PRP-Exos | 50 µg/mL | Cell viability, cell proliferation, tube formation, migration, apoptosis, osteogenesis, Western blot. | PRP-Exos played proliferative and anti-apoptotic roles against GC-ER induced-stress in vitro and in vivo. The co-administration with PRP-Exos could rescue angiogenesis in vitro and had a notable protective effect on blood vessels in vivo. PRP-Exos could prevent the inhibition of osteogenesis observed in BMSCs and MC3T3-E1 cells caused by DEX in vitro and could rescue the expression of osteogenesis-related proteins. PRP-Exos showed to be effective in preventing GC-induced ONFH in rats. The stimulatory effects of PRP-Exos on anti-apoptosis mainly resulted from the activation of the Akt/Bad/Bcl-2 signal pathway. In bone cells, PRP-Exos rescued the osteogenic protein expression level through the Wnt/β-catenin signaling pathway. |
| In vivo | MPS-treated in vivo rat model | Group I: Control group Group II: MPS group Group III: PRP-Exos group | 100 μg exosomes (dissolved in 200 μL of PBS) (500 µg/mL) | Micro CT (angiography and osteogenesis), TUNEL assay and Ki67 immunostaining |
| Article | PRP Origin | Anticoagulant | Leukocytes | Activator | EVs/Exosomes |
|---|---|---|---|---|---|
| Liu et al., 2019 [31] | Rabbit PRP | ACD-A | No | No | PRP-derived exosomes |
| Otahal et al., 2020 [32] | Human CPRP | Trisodium Citrate | Low numbers of monocytes (20 per µL) | No | CPRP-EVs |
| Otahal et al., 2021 [33] | Human CPRP | Trisodium Citrate | NS | No | CPRP-EVs |
| Zhang et al., 2022 [34] | Human PRP | ACD-A | NS | CaCl2 and thrombin | PRP-derived exosomes |
| Rui et al., 2021 [35] | Human PRP | Citrate glucose | No | Thrombin/Calcium gluconate/Mixture of both | PRP-derived exosomes |
| Saumell-Esnaola et al., 2022 [36] | Human PRP (13-00-11) | Sodium citrate | No | CaCl2 | PRP Platelets derived exosomes |
| Stoner et al., 2016 [37] | Rat PRP | Sodium citrate | NS | Ionophore A23187 | EVs |
| Yuana et al., 2013 [38] | Human PRP | Sodium citrate and EDTA | NS | No | PRP-EVs |
| Zhang et al., 2019 [39] | Rat PRP | Sodium citrate | NS | NS *,** | PRP-derived exosomes |
| Zhang et al., 2020 [40] | Rat PRP (from diabetic rats and from normal rats) | Sodium citrate | NS | NS * | PRP-derived exosomes |
| Dai et al., 2023 [41] | Human PRP | ACD-A | NS | w/o and w/Thrombin | Platelet-derived EVs |
| Xu et al., 2021 [42] | NS | NS | NS | NS | PRP-derived exosomes |
| Catitti et al., 2022 [43] | Human PRP | ACD-A | No | NS | PRP-EVs |
| Iyer et al., 2020 [44] | Rat PRP (Arthrex ACP Double Syringe System) | NS | No | Sonication | PRP-derived exosomes |
| Guo et al., 2017 [45] | Human PRP | ACD-A | No | NS * | PRP Platelets derived exosomes |
| Xu et al., 2021 [46] | Mice PRP | NS | Yes | NS | PRP-derived exosomes |
| Dong et al., 2022 [47] | Rat PRP | ACD-A | No | Sonication | PRP-derived exosomes |
| Bagio et al., 2023 [48] | Human PRP | NS | Yes | Thrombin | T-aPDE |
| Nilforoushzadeh et al., 2021 [49] | Human PRP (from cord blood) | NS | NS | NS | PRP-derived exosomes |
| Tao et al., 2017 [50] | Human PRP | ACD-A | No | NS * | PRP-derived exosomes |
| Article | Isolation | EVs Storage | |||
|---|---|---|---|---|---|
| Type of Tube | Buffer | Temperature | Time | ||
| Liu et al., 2019 [31] | ExoEasy Maxi kit (Qiagen) | N.S. | PBS | Frozen (−80 °C) | N.S. |
| Otahal et al., 2020 [32] | Ultracentrifugation | N.S. | PBS | Frozen (−80 °C) | N.S. |
| Otahal et al., 2021 [33] | Ultracentrifugation | N.S. | PBS | Frozen (−80 °C) | N.S. |
| Zhang et al., 2022 [34] | Ultrafiltration/purification with 30% sucrose D2O cushion and ultracentrifugation | Ultra-clearTM tube (Beckman Coulter) | PBS | Frozen (−80 °C) | N.S. |
| Rui et al., 2021 [35] | Gradient ultracentrifugation | N.S. | PBS containing protease inhibitor cocktail and phosphataseinhibitor cocktail | Frozen (−80 °C) | N.S. |
| Saumell-Esnaola et al., 2022 [36] | Differential ultracentrifugation | N.S. | PBS | Frozen (−80 °C) | N.S. |
| Stoner et al., 2016 [37] | N.S. | N.S. | N.S. | N.S. | N.S. |
| Yuana et al., 2013 [38] | Ultracentrifugation | N.S. | HEPES buffer | Vitrification | N.S. |
| Zhang et al., 2019 [39] | Ultracentrifugation | N.S. | PBS | Frozen (−80 °C) | N.S. |
| Zhang et al., 2020 [40] | Ultracentrifugation | N.S. | PBS | Frozen (−80 °C) | N.S. |
| Dai et al., 2023 [41] | Extrusion and ultracentrifugation | N.S. | PBS + EDTA + PGE1 | N.S. | Test stability 0, 1, 3, 5, and 7 days (in vitro) |
| Xu et al., 2021 [42] | N.S. | N.S. | N.S. | N.S. | N.S. |
| Catitti et al., 2022 [43] | Fluorescence activated cell sorting | N.S. | N.S. | N.S. | N.S. |
| Iyer et al., 2020 [44] | Ultracentrifugation | Ultra-carbon fiber tube | PBS | N.S. | N.S. |
| Guo et al., 2017 [45] | Ultrafiltration/purification with 30% sucrose D2O cushion and ultracentrifugation | Ultra-clearTM tube (Beckman Coulter) | PBS | Frozen (−80 °C) | N.S. |
| Xu et al., 2021 [46] | Ultracentrifugation | N.S. | PBS | N.S. | N.S. |
| Dong et al., 2022 [47] | ExoEasy Maxi kit (Qiagen) | N.S. | PBS | Frozen (−80 °C) | N.S. |
| Bagio et al., 2023 [48] | Centrifugation | N.S. | PBS | 4 °C to −20 °C | 7 days |
| Nilforoushzadeh et al., 2021 [49] | Ultracentrifugation | N.S. | PBS | Frozen (−80 °C) | N.S. |
| Tao et al., 2017 [50] | Ultrafiltration/purification with 30% sucrose D2O cushion and ultracentrifugation | Ultra-clearTM tube (Beckman Coulter) | PBS | Frozen (−80 °C) | N.S. |
| Article | Markers | Size | ||
|---|---|---|---|---|
| Cat. 1 (Transmembrane or GPI-Anchored Proteins) | Cat. 2 (Cytosolic Proteins) | Cat. 3 (Purity Control) | ||
| Liu et al., 2019 [31] | ✓ (CD9, CD63, CD81) | ✓ (TSG101) | ✕ | 145.6 ± 50.4 nm |
| Otahal et al., 2020 [32] | ✓ (CD14, CD41, CD9, CD63, CD235a) | ✓ (ALIX) | ✓ (APOA1, APOB100) | *** |
| Otahal et al., 2021 [33] | ✓ (CD9, CD63) | ✓ (ALIX) | ✓ (APOA1, APOB100/48) | *** |
| Zhang et al., 2022 [34] | ✓ (CD9, CD63, CD81) | ✕ | ✕ | Ranging from 40 to 140 nm |
| Rui et al., 2021 [35] | ✓ (CD9, CD41, CD63, CD81) | ✓ (Flotillin, TSG101) | ✓ * (Calnexin) | Ranging from 30 to 150 nm |
| Saumell-Esnaola et al., 2022 [36] | ✓ (CD9, CD63, CD81, LAMP1) | ✓ (β-actin, Caveolin-1, Flotillin, HSP70) | ✓ (APOA1) | Ranging from 20 to 40 nm |
| Stoner et al., 2016 [37] | ✓ (CD61) | ✕ | ✕ | *** |
| Yuana et al., 2013 [38] | ✕ | ✕ | ✕ | Ranging from 25 to 260 nm |
| Zhang et al., 2019 [39] | ✓ (CD9, CD61, CD63, CD81) | ✕ | ✕ | N.S. |
| Zhang et al., 2020 [40] | ✓ (CD9, CD61, CD63, CD81) | ✓ (TSG101) | ✕ | Peak sizes of 84 nm and 95 nm |
| Dai et al., 2023 [41] | ✓ (CD9, CD41, CD63) | ✕ | ✕ | 195.6 nm |
| Xu et al., 2021 [42] | ✓ (CD81) | ✓ (TSG101) | ✕ | N.S. |
| Catitti et al., 2022 [43] | ✓ ** | ✓ ** | ✓ ** | N.S. |
| Iyer et al., 2020 [44] | ✓ (CD9, CD63, CD81) | ✕ | ✕ | Ranging from 30 to 200 nm |
| Guo et al., 2017 [45] | ✓ (CD9, CD41, CD63, CD81) | ✕ | ✕ | Ranging from 40 to 100 nm |
| Xu et al., 2021 [46] | ✓ (CD9) | ✓ (TSG101) | ✕ | Ranging from 40 to 100 nm |
| Dong et al., 2022 [47] | ✓ (CD9, CD63, CD81) | ✓ (ALIX) | ✕ | 145.6 ± 50.4 nm |
| Bagio et al., 2023 [48] | ✓ (CD63, CD81) | ✕ | ✕ | Ranging from 44 to 127 nm |
| Nilforoushzadeh et al., 2021 [49] | ✓ (CD9, CD63, CD81) | ✓ (TSG101) | ✓ * (Calnexin) | Ranging from 50 to 150 nm |
| Tao et al., 2017 [50] | ✓ (CD9, CD41, CD63, CD81) | ✓ (TSG101) | ✓ * (Calnexin) | Ranging from 30 to 100 nm |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anitua, E.; Troya, M.; Falcon-Pérez, J.M.; López-Sarrio, S.; González, E.; Alkhraisat, M.H. Advances in Platelet Rich Plasma-Derived Extracellular Vesicles for Regenerative Medicine: A Systematic-Narrative Review. Int. J. Mol. Sci. 2023, 24, 13043. https://doi.org/10.3390/ijms241713043
Anitua E, Troya M, Falcon-Pérez JM, López-Sarrio S, González E, Alkhraisat MH. Advances in Platelet Rich Plasma-Derived Extracellular Vesicles for Regenerative Medicine: A Systematic-Narrative Review. International Journal of Molecular Sciences. 2023; 24(17):13043. https://doi.org/10.3390/ijms241713043
Chicago/Turabian StyleAnitua, Eduardo, María Troya, Juan Manuel Falcon-Pérez, Silvia López-Sarrio, Esperanza González, and Mohammad H. Alkhraisat. 2023. "Advances in Platelet Rich Plasma-Derived Extracellular Vesicles for Regenerative Medicine: A Systematic-Narrative Review" International Journal of Molecular Sciences 24, no. 17: 13043. https://doi.org/10.3390/ijms241713043
APA StyleAnitua, E., Troya, M., Falcon-Pérez, J. M., López-Sarrio, S., González, E., & Alkhraisat, M. H. (2023). Advances in Platelet Rich Plasma-Derived Extracellular Vesicles for Regenerative Medicine: A Systematic-Narrative Review. International Journal of Molecular Sciences, 24(17), 13043. https://doi.org/10.3390/ijms241713043