

Iron Load Toxicity in Medicine: From Molecular and Cellular Aspects to Clinical Implications


Abstract
1. Introduction
2. The Source of Iron Overload from Chronic Red Blood Cell Transfusions
3. Major Diseases of Transfusional Iron Overload
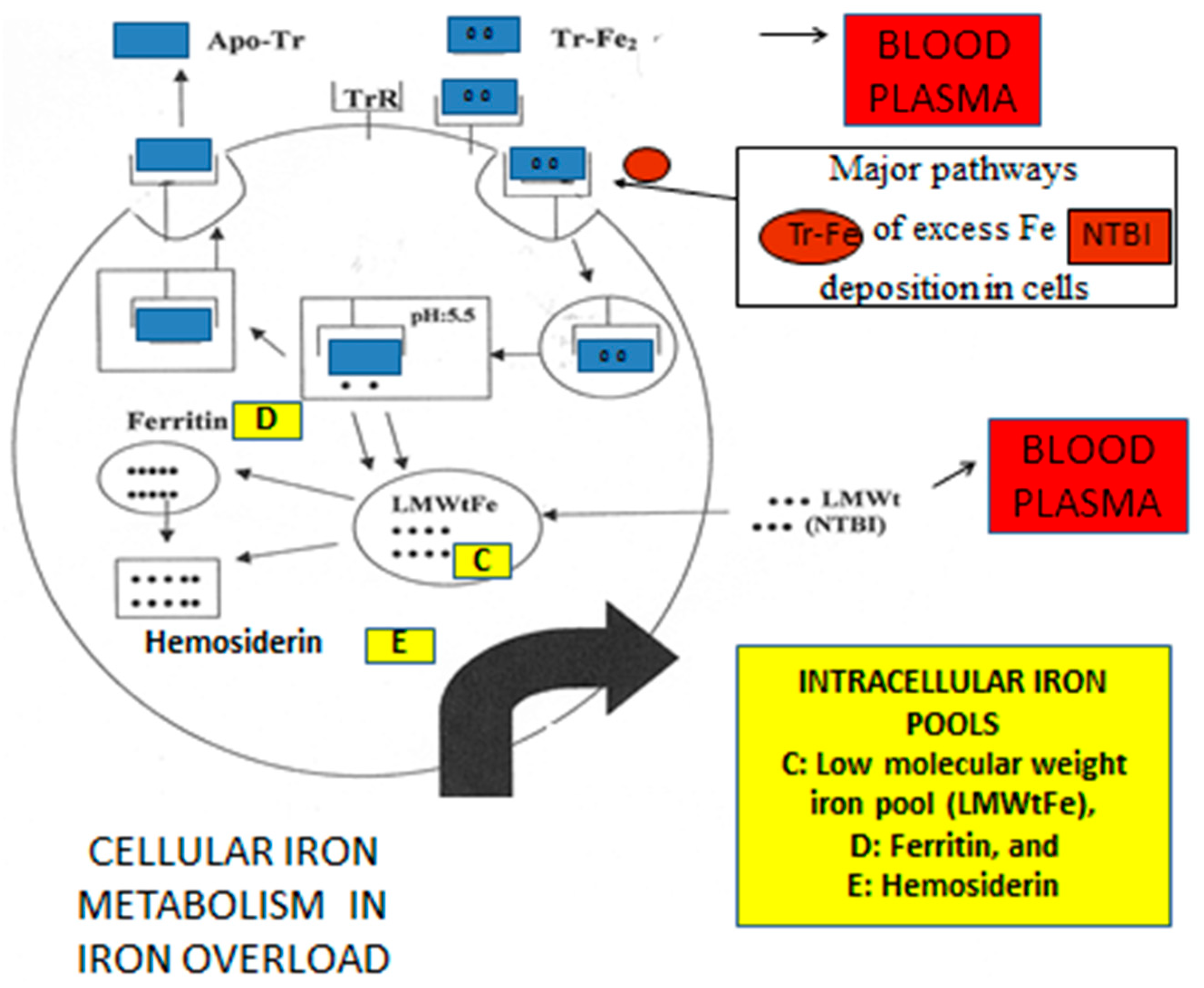
4. Iron Metabolism and Iron Toxicity in Chronically Transfused Patients

5. Diagnosis and Monitoring of Transfusional Iron Overload
6. Excess Iron Deposition and Toxicity in Non-Transfusional Iron Loading Conditions
7. Iron Toxicity from Labile Forms of Iron and Other Molecular Interactions
8. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| DF | deferoxamine |
| DFRA | deferasirox |
| DMT1 | divalent metal transported protein |
| EDTA | ethylenediaminetetraacetic acid |
| GPX4 | glutathione peroxidase |
| HCT | hematopoietic stem cell transplantation |
| Hb | hemoglobin |
| HPFH | high persistent fetal hemoglobin |
| L1 | deferiprone |
| LMWt | low molecular weight |
| MRI | magnetic resonance imaging |
| NBIA | neurodegeneration with brain iron accumulation |
| NTBI | non-transferrin bound iron |
| PKAN | pantothenate kinase 2-associated neurodegeneration |
| RBC | red blood cell |
| SCD | sickle cell disease |
| TI | thalassemia intermedia |
| TM | beta thalassemia major |
References
- Hokland, P.; Daar, S.; Khair, W.; Sheth, S.; Taher, A.T.; Torti, L.; Hantaweepant, C.; Rund, D. Thalassaemia—A global view. Br. J. Haematol. 2023, 201, 199–214. [Google Scholar] [CrossRef]
- Cattoni, A.; Capitoli, G.; Casagranda, S.; Corti, P.; Adavastro, M.; Molinaro, A.; Di Gennaro, F.; Bonanomi, S.; Biondi, A.; Galimberti, S.; et al. Iron Overload Following Hematopoietic Stem Cell Transplantation: Prevalence, Severity, and Management in Children and Adolescents with Malignant and Nonmalignant Diseases. Transplant. Cell Ther. 2023, 29, 271.e1–271.e12. [Google Scholar] [CrossRef] [PubMed]
- Tang, A.Y.; Zhou, M.; Maillis, A.N.; Lai, K.W.; Lane, P.A.; Snyder, A.B. Trends in blood transfusion, hydroxyurea use, and iron overload among children with sickle cell disease enrolled in Medicaid, 2004–2019. Pediatr. Blood Cancer 2023, 70, e30152. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Estcourt, L.J.; Doree, C.; Hopewell, S.; Vyas, P. Comparison of a restrictive versus liberal red cell transfusion policy for patients with myelodysplasia, aplastic anaemia, and other congenital bone marrow failure disorders. Cochrane Database Syst. Rev. 2015, 2015, CD011577. [Google Scholar] [CrossRef] [PubMed]
- Styczyński, J.; Słomka, A.; Łęcka, M.; Albrecht, K.; Romiszewski, M.; Pogorzała, M.; Kubicka, M.; Kuryło-Rafińska, B.; Tejza, B.; Gadomska, G.; et al. Soluble Hemojuvelin and Ferritin: Potential Prognostic Markers in Pediatric Hematopoietic Cell Transplantation. Cancers 2023, 15, 1041. [Google Scholar] [CrossRef]
- Iancu, T.C. Iron overload. Mol. Aspects Med. 1983, 6, 1–100. [Google Scholar] [CrossRef]
- Bruzzese, A.; Martino, E.A.; Mendicino, F.; Lucia, E.; Olivito, V.; Bova, C.; Filippelli, G.; Capodanno, I.; Neri, A.; Morabito, F.; et al. Iron chelation therapy. Eur. J. Haematol. 2023, 110, 490–497. [Google Scholar] [CrossRef]
- Rai, P.; Okhomina, V.I.; Kang, G.; Martinez, H.R.; Hankins, J.S.; Joshi, V. Longitudinal effect of disease-modifying therapy on left ventricular diastolic function in children with sickle cell anemia. Am. J. Hematol. 2023, 98, 838–847. [Google Scholar] [CrossRef]
- Zurlo, M.; De Stefano, P.; Borgna-Pignatti, C.; Di Palma, A.; Melevendi, C.; Piga, A.; Di Gregorio, F.; Burattini, M.; Terzoli, S. Survival and causes of death in thalassaemia major. Lancet 1989, 2, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Adams, P.C. Epidemiology and diagnostic testing for hemochromatosis and iron overload. Int. J. Lab. Hematol. 2015, 37, 25–30. [Google Scholar] [CrossRef]
- Lehéricy, S.; Roze, E.; Goizet, C.; Mochel, F. MRI of neurodegeneration with brain iron accumulation. Curr. Opin. Neurol. 2020, 33, 462–473. [Google Scholar] [CrossRef] [PubMed]
- Kolnagou, A.; Fessas, C.; Papatryphonas, A.; Economides, C.; Kontoghiorghes, G.J. Prophylactic use of deferiprone (L1) and magnetic resonance imaging T2* or T2 for preventing heart disease in thalassaemia. Br. J. Haematol. 2004, 127, 360–361. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J.; Kontoghiorghe, C.N. Efficacy and safety of iron-chelation therapy with deferoxamine, deferiprone, and deferasirox for the treatment of iron-loaded patients with non-transfusion-dependent thalassemia syndromes. Drug Des. Dev. Ther. 2016, 10, 465–481. [Google Scholar] [CrossRef]
- Telfer, P.; Coen, P.G.; Christou, S.; Hadjigavriel, M.; Kolnakou, A.; Pangalou, E.; Pavlides, N.; Psiloines, M.; Simamonian, K.; Skordos, G.; et al. Survival of medically treated thalassemia patients in Cyprus. Trends and risk factors over the period 1980–2004. Haematologica 2006, 91, 1187–1192. [Google Scholar]
- Au, W.Y.; Lee, V.; Lau, C.W.; Yau, J.; Chan, D.; Chan, E.Y.T.; Cheung, W.W.W.; Ha, S.Y.; Kho, B.; Lee, C.Y.; et al. A synopsis of current care of thalassaemia major patients in Hong Kong. Hong Kong Med. J. 2011, 17, 261–266. [Google Scholar] [PubMed]
- Maggio, A.; Filosa, A.; Vitrano, A.; Aloj, G.; Kattamis, A.; Ceci, A.; Fucharoen, S.; Cianciulli, P.; Grady, R.W.; Prossomariti, L.; et al. Iron chelation therapy in thalassemia major: A systematic review with meta-analyses of 1520 patients included on randomized clinical trials. Blood Cells Mol. Dis. 2011, 47, 166–175. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J. The aim of iron chelation therapy in thalassaemia. Eur. J. Haematol. 2017, 99, 465–466. [Google Scholar] [CrossRef] [PubMed]
- Cairo, G.; Bernuzzi, F.; Recalcati, S. A precious metal: Iron, an essential nutrient for all cells. Genes Nutr. 2006, 1, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Prudent, M.; D’Alessandro, A. Red blood cell storage lesion: Causes and potential clinical consequences. Blood Transfus. 2019, 17, 27–52. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J.; Kontoghiorghe, C.N. Iron and Chelation in Biochemistry and Medicine: New Approaches to Controlling Iron Metabolism and Treating Related Diseases. Cells 2020, 9, 1456. [Google Scholar] [CrossRef] [PubMed]
- Galaris, D.; Pantopoulos, K. Oxidative Stress and Iron Homeostasis: Mechanistic and Health Aspects. Crit. Rev. Clin. Lab. Sci. 2008, 45, 1–23. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C.; Cross, C.E. Free radicals, antioxidants and human disease: Where are we now? J. Lab. Clin. Med. 1992, 119, 598–620. [Google Scholar]
- Fibach, E. The Redox Balance and Membrane Shedding in RBC Production, Maturation, and Senescence. Front. Physiol. 2021, 12, 604738. [Google Scholar] [CrossRef]
- Asaro, R.J.; Zhu, Q.; Cabrales, P. Erythrocyte Aging, Protection via Vesiculation: An Analysis Methodology via Oscillatory Flow. Front. Physiol. 2018, 9, 1607. [Google Scholar] [CrossRef] [PubMed]
- Alaarg, A.; Schiffelers, R.M.; van Solinge, W.W.; van Wijk, R. Red blood cell vesiculation in hereditary hemolytic anemia. Front. Physiol. 2013, 4, 365. [Google Scholar] [CrossRef]
- Tzounakas, V.L.; Anastasiadi, A.T.; Lekka, M.E.; Papageorgiou, E.G.; Stamoulis, K.; Papassideri, I.S.; Kriebardis, A.G.; Antonelou, M.H. Deciphering the Relationship Between Free and Vesicular Hemoglobin in Stored Red Blood Cell Units. Front. Physiol. 2022, 13, 840995. [Google Scholar] [CrossRef] [PubMed]
- Tolosano, E.; Altruda, F. Hemopexin: Structure, Function, and Regulation. DNA Cell Boil. 2002, 21, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Shih, A.W.; McFarlane, A.; Verhovsek, M. Haptoglobin testing in hemolysis: Measurement and interpretation. Am. J. Hematol. 2014, 89, 443–447. [Google Scholar] [CrossRef]
- Weatherall, D.J.; Clegg, J.B. Inherited haemoglobin disorders: An increasing global health problem. Bull. World Health Organ. 2001, 79, 704–712. [Google Scholar]
- Anonymous. Community control of hereditary anaemias: Memorandum from a WHO meeting. Bull. World Health Organ. 1983, 61, 63–80. [Google Scholar]
- Teawtrakul, N.; Chansung, K.; Sirijerachai, C.; Wanitpongpun, C.; Thepsuthammarat, K. The impact and disease burden of thalassemia in Thailand: A population-based study in 2010. J. Med. Assoc. Thail. 2012, 95, 95. [Google Scholar]
- Pippard, M.J.; Callender, S.T.; Warner, G.T.; Weatherall, D.J. Iron absorption and loading in beta-thalassaemia intermedia. Lancet 1979, 2, 819–821. [Google Scholar] [CrossRef]
- Pootrakul, P.; Kitcharoen, K.; Yansukon, P.; Wasi, P.; Fucharoen, S.; Charoenlarp, P.; Brittenham, G.; Pippard, M.J.; Finch, C.A. The effect of erythroid hyperplasia on iron balance. Blood 1988, 71, 1124–1129. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J.; Spyrou, A.; Kolnagou, A. Iron Chelation Therapy in Hereditary Hemochromatosis and Thalassemia Intermedia: Regulatory and Non Regulatory Mechanisms of Increased Iron Absorption. Hemoglobin 2010, 34, 251–264. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J.; Kolnagou, A. Molecular Factors and Mechanisms Affecting Iron and Other Metal Excretion or Absorption in Health and Disease. The Role of Natural and Synthetic Chelators. Curr. Med. Chem. 2005, 12, 2695–2709. [Google Scholar] [CrossRef] [PubMed]
- Galanello, R.; Cao, A. Relationship between Genotype and Phenotype: Thalassemia Intermediaa. Ann. N. Y. Acad. Sci. 1998, 850, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Borgna-Pignatti, C. Modern treatment of thalassaemia intermedia. Br. J. Haematol. 2007, 138, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Singer, S.T. Variable Clinical Phenotypes of α-Thalassemia Syndromes. Sci. World J. 2008, 9, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Kolnagou, A.; Kleanthous, M.; Kontoghiorghes, G.J. Benefits and Risks in Polypathology and Polypharmacotherapy Challenges in the Era of the Transition of Thalassaemia from a Fatal to a Chronic or Curable Disease. Front. Biosci. 2022, 14, 18. [Google Scholar] [CrossRef]
- Inamoto, Y.; Lee, S.J. Late effects of blood and marrow transplantation. Haematologica 2017, 102, 614–625. [Google Scholar] [CrossRef] [PubMed]
- Gratwohl, A.; Pasquini, M.C.; Aljurf, M.; Atsuta, Y.; Baldomero, H.; Foeken, L.; Gratwohl, M.; Bouzas, L.F.; Confer, D.; Frauendorfer, K.; et al. Worldwide Network for Blood and Marrow Transplantation (WBMT). One million haemopoietic stem-cell transplants: A retrospective observational study. Lancet Haematol. 2015, 2, e91–e100. [Google Scholar] [CrossRef]
- Shenoy, S.; Angelucci, E.; Arnold, S.D.; Baker, K.S.; Bhatia, M.; Bresters, D.; Dietz, A.C.; De La Fuente, J.; Duncan, C.; Gaziev, J.; et al. Current Results and Future Research Priorities in Late Effects after Hematopoietic Stem Cell Transplantation for Children with Sickle Cell Disease and Thalassemia: A Consensus Statement from the Second Pediatric Blood and Marrow Transplant Consortium International Conference on Late Effects after Pediatric Hematopoietic Stem Cell Transplantation. Biol. Blood Marrow Transplant. 2017, 23, 552–561. [Google Scholar] [CrossRef]
- Germing, U.; Schroeder, T.; Kaivers, J.; Kündgen, A.; Kobbe, G.; Gattermann, N. Novel therapies in low- and high-risk myelodysplastic syndrome. Expert Rev. Hematol. 2019, 12, 893–908. [Google Scholar] [CrossRef]
- Ricci, A.; Di Betto, G.; Bergamini, E.; Buzzetti, E.; Corradini, E.; Ventura, P. Iron Metabolism in the Disorders of Heme Biosynthesis. Metabolites 2022, 12, 819. [Google Scholar] [CrossRef]
- Gao, J.; Zhou, Q.; Wu, D.; Chen, L. Mitochondrial iron metabolism and its role in diseases. Clin. Chim. Acta 2021, 513, 6–12. [Google Scholar] [CrossRef]
- Girard, S.; Genevieve, F.; Rault, E.; Fenneteau, O.; Lesesve, J.-F. When Ring Sideroblasts on Bone Marrow Smears Are Inconsistent with the Diagnosis of Myelodysplastic Neoplasms. Diagnostics 2022, 12, 1752. [Google Scholar] [CrossRef]
- Brissot, P.; Troadec, M.-B.; Loréal, O.; Brissot, E. Pathophysiology and classification of iron overload diseases; update 2018. Transfus. Clin. Biol. 2019, 26, 80–88. [Google Scholar] [CrossRef]
- Camaschella, C.; Nai, A. Ineffective erythropoiesis and regulation of iron status in iron loading anaemias. Br. J. Haematol. 2016, 172, 512–523. [Google Scholar] [CrossRef]
- Verma, I.C. Burden of genetic disorders in India. Indian J. Pediatr. 2000, 67, 893–898. [Google Scholar] [CrossRef]
- Gozzelino, R.; Arosio, P. Iron Homeostasis in Health and Disease. Int. J. Mol. Sci. 2016, 17, 130. [Google Scholar] [CrossRef]
- Anderson, G.J.; Frazer, D.M. Current understanding of iron homeostasis. Am. J. Clin. Nutr. 2017, 106 (Suppl. 6), 1559S–1566S. [Google Scholar] [CrossRef]
- Andrews, N.C. Closing the iron gate. N. Engl. J. Med. 2012, 366, 376–377. [Google Scholar] [CrossRef]
- Kim, A.; Nemeth, E. New insights into iron regulation and erythropoiesis. Curr. Opin. Hematol. 2015, 22, 199–205. [Google Scholar] [CrossRef]
- Galaris, D.; Barbouti, A.; Pantopoulos, K. Iron homeostasis and oxidative stress: An intimate relationship. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2019, 1866, 118535. [Google Scholar] [CrossRef]
- Sargent, P.J.; Farnaud, S.; Evans, R.W. Structure/Function Overview of Proteins Involved in Iron Storage and Transport. Curr. Med. Chem. 2005, 12, 2683–2693. [Google Scholar] [CrossRef]
- Theil, E.C. Ferritin: The Protein Nanocage and Iron Biomineral in Health and in Disease. Inorg. Chem. 2013, 52, 12223–12233. [Google Scholar] [CrossRef]
- Iancu, T.C. Ferritin and hemosiderin in pathological tissues. Electron Microsc. Rev. 1992, 5, 209–229. [Google Scholar] [CrossRef]
- Denisov, E.T.; Afanas’Ev, I.B. Oxidation and Antioxidants in Organic Chemistry and Biology; CRC Press: Boca Raton, FL, USA; Taylor and Francis group: Abingdon, UK, 2005. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J. Iron Chelation in Biochemistry and Medicine; Free radicals, oxidant stress and drug action; Rice-Evans, C., Ed.; Rechelieu Press: London, UK, 1987; pp. 277–303. [Google Scholar]
- Duračková, Z. Some current insights into oxidative stress. Physiol. Res. 2010, 59, 459–469. [Google Scholar] [CrossRef]
- Nakamura, T.; Naguro, I.; Ichijo, H. Iron homeostasis and iron-regulated ROS in cell death, senescence and human diseases. Biochim. Biophys. Acta (BBA) Gen. Subj. 2019, 1863, 1398–1409. [Google Scholar] [CrossRef]
- Shah, S.V.; Rajapurkar, M.M. The Role of Labile Iron in Kidney Disease and Treatment with Chelation. Hemoglobin 2009, 33, 378–385. [Google Scholar] [CrossRef]
- Reeder, B.J.; Wilson, M.T. Hemoglobin and Myoglobin Associated Oxidative Stress: From Molecular Mechanisms to Disease States. Curr. Med. Chem. 2005, 12, 2741–2751. [Google Scholar] [CrossRef]
- Reeder, B.J.; Strader, M.B.; Alayash, A.I.; Di Domenico, F.; Barone, E.; Perluigi, M.; Butterfield, D.A.; Feliers, D.; Lee, H.J.; Kasinath, B.S.; et al. Redox and Peroxidase Activities of the Hemoglobin Superfamily: Relevance to Health and Disease. Antioxidants Redox Signal. 2017, 26, 763–776. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, A. An intracellular transit iron pool. Ciba. Found Symp. 1976, 51, 91–106. [Google Scholar]
- Kontoghiorghes, G.J.; Efstathiou, A.; Kleanthous, M.; Michaelides, Y.; Kolnagou, A. Risk/Benefit Assessment, Advantages Over Other Drugs and Targeting Methods in the Use of Deferiprone as a Pharmaceutical Antioxidant in Iron Loading and Non Iron Loading Conditions. Hemoglobin 2009, 33, 386–397. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J.; Kontoghiorghe, C.N. Prospects for the introduction of targeted antioxidant drugs for the prevention and treatment of diseases related to free radical pathology. Expert Opin. Investig. Drugs 2019, 28, 593–603. [Google Scholar] [CrossRef]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Hou, W.; Song, X.; Yu, Y.; Huang, J.; Sun, X.; Kang, R.; Tang, D. Ferroptosis: Process and function. Cell Death Differ. 2016, 23, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.Y.; Dixon, S.J. Mechanisms of ferroptosis. Cell Mol. Life Sci. 2016, 73, 2195–2209. [Google Scholar] [CrossRef]
- Lu, B.; Chen, X.B.; Ying, M.D.; He, Q.J.; Cao, J.; Yang, B. The Role of Ferroptosis in Cancer Development and Treatment Response. Front. Pharmacol. 2018, 8, 992. [Google Scholar] [CrossRef]
- Li, G.; Liang, Y.; Yang, H.; Zhang, W.; Xie, T. The Research Landscape of Ferroptosis in Cancer: A Bibliometric Analysis. Front. Cell Dev. Biol. 2022, 10, 841724. [Google Scholar] [CrossRef]
- Wang, D.; Tang, L.; Zhang, Y.; Ge, G.; Jiang, X.; Mo, Y.; Wu, P.; Deng, X.; Li, L.; Zuo, S.; et al. Regulatory pathways and drugs associated with ferroptosis in tumors. Cell Death Dis. 2022, 13, 544. [Google Scholar] [CrossRef]
- Liu, L.; Li, L.; Li, M.; Luo, Z. Autophagy-Dependent Ferroptosis as a Therapeutic Target in Cancer. Chemmedchem 2021, 16, 2942–2950. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Huang, H.; Fan, S.; Zheng, B.; Wu, J.; Zhang, J.; Pi, J.; Xu, J.-F. Ferroptosis: A mixed blessing for infectious diseases. Front. Pharmacol. 2022, 13, 992734. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Xu, W.; Hou, Y.; Wang, S.; Zhang, H.; Ran, M.; Huang, Y.; Wang, Y.; Yang, G. The multifaceted role of ferroptosis in kidney diseases. Chem. Interactions 2022, 365, 110107. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Chen, X.-Z.; Wang, Y.-H.; Cheng, X.-L.; Zhao, Y.; Zhou, L.-Y.; Wang, K. Emerging roles of ferroptosis in cardiovascular diseases. Cell Death Discov. 2022, 8, 394. [Google Scholar] [CrossRef]
- Thapa, K.; Khan, H.; Kanojia, N.; Singh, T.G.; Kaur, A.; Kaur, G. Therapeutic Insights on Ferroptosis in Parkinson’s disease. Eur. J. Pharmacol. 2022, 930, 175133. [Google Scholar] [CrossRef]
- Jacobs, W.; Lammens, M.; Kerckhofs, A.; Voets, E.; Van San, E.; Van Coillie, S.; Peleman, C.; Mergeay, M.; Sirimsi, S.; Matheeussen, V.; et al. Fatal lymphocytic cardiac damage in coronavirus disease 2019 (COVID-19): Autopsy reveals a ferroptosis signature. ESC Heart Fail. 2020, 7, 3772–3781. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J. Deferiprone: A Forty-Year-Old Multi-Targeting Drug with Possible Activity against COVID-19 and Diseases of Similar Symptomatology. Int. J. Mol. Sci. 2022, 23, 6735. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, Y.; Guo, L.; Gao, W.; Tang, T.-L.; Yan, M. Interaction between macrophages and ferroptosis. Cell Death Dis. 2022, 13, 355. [Google Scholar] [CrossRef]
- Gao, M.; Monian, P.; Pan, Q.; Zhang, W.; Xiang, J.; Jiang, X. Ferroptosis is an autophagic cell death process. Cell Res. 2016, 26, 1021–1032. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Q.; Song, H.; Zhang, Y.; Chen, H.; Liu, P.; Sun, T.; Jiang, C. A Triple Therapeutic Strategy with Antiexosomal Iron Efflux for Enhanced Ferroptosis Therapy and Immunotherapy. Small 2022, 18, e2201704. [Google Scholar] [CrossRef]
- Hou, W.; Xie, Y.; Song, X.; Sun, X.; Lotze, M.T.; Zeh, H.J., 3rd; Kang, R.; Tang, D. Autophagy promotes ferroptosis by degradation of ferritin. Autophagy 2016, 12, 1425–1428. [Google Scholar] [CrossRef]
- Park, E.; Chung, S.W. ROS-mediated autophagy increases intracellular iron levels and ferroptosis by ferritin and transferrin receptor regulation. Cell Death Dis. 2019, 10, 822. [Google Scholar] [CrossRef] [PubMed]
- Kyriacou, K.; Michaelides, Y.; Senkus, R.; Simamonian, K.; Pavlides, N.; Antoniades, L.; Zambartas, C. Ultrastructural pathology of the heart in patients with beta-thalassaemia major. Ultrastruct. Pathol. 2000, 24, 75–81. [Google Scholar] [CrossRef]
- Iancu, T.C.; Neustein, H.B.; Landing, B.H. The Liver in Thalassaemia Major: Ultra-Structural Observations. Ciba Foundation Symposium 51-Iron Metabolism; John Wiley & Sons, Ltd.: Chichester, UK, 1977; pp. 293–316. [Google Scholar] [CrossRef]
- Kolnagou, A.; Michaelides, Y.; Kontos, C.; Kyriacou, K.; Kontoghiorghes, G.J. Myocyte Damage and Loss of Myofibers is the Potential Mechanism of Iron Overload Toxicity in Congestive Cardiac Failure in Thalassemia. Complete Reversal of the Cardiomyopathy and Normalization of Iron Load by Deferiprone. Hemoglobin 2008, 32, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J.; Kolnagou, A.; Kontoghiorghe, C.N. New targeted therapies and diagnostic methods for iron overload diseases. Front. Biosci. 2018, 10, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Aessopos, A.; Kati, M.; Farmakis, D. Heart disease in thalassemia intermedia: A review of the underlying pathophysiology. Haematologica 2007, 92, 658–665. [Google Scholar] [CrossRef]
- Sonakul, D.; Pacharee, P.; Wasi, P.; Fucharoen, S. Cardiac pathology in 47 patients with beta thalassaemia/haemoglobin E. Southeast Asian J. Trop. Med. Public Health 1984, 15, 554–563. [Google Scholar]
- Mancuso, A. Evidence-based medicine and Management of Hepatocellular Carcinoma in Thalassemia. BMC Gastroenterol. 2020, 20, 409. [Google Scholar] [CrossRef]
- Ricchi, P. Liver fibrosis in young patients with transfusion-dependent thalassaemia (TDT), understanding the role of steatosis. Br. J. Haematol. 2023, 200, 399–400. [Google Scholar] [CrossRef]
- Tolis, G.J.; Vlachopapadopoulou, E.; Karydis, I. Reproductive health in patients with beta-thalassemia. Curr. Opin. Pediatr. 1996, 8, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Angelopoulos, N.G.; Zervas, A.; Livadas, S.; Adamopoulos, I.; Giannopoulos, D.; Goula, A.; Tolis, G. Reduced insulin secretion in normoglycaemic patients with β-thalassaemia major. Diabet. Med. 2006, 23, 1327–1331. [Google Scholar] [CrossRef] [PubMed]
- Economides, C.P.; Soteriades, E.S.; Hadjigavriel, M.; Seimenis, I.; Karantanas, A. Iron deposits in the knee joints of a thalassemic patient. Acta Radiol. Short Rep. 2013, 2, 1–5. [Google Scholar] [CrossRef]
- Mavrogeni, S.I.; Gotsis, E.D.; Markussis, V.; Tsekos, N.; Politis, C.; Vretou, E.; Kermastinos, D. T2 relaxation time study of iron overload in b-thalassemia. MAGMA 1998, 6, 7–12. [Google Scholar] [CrossRef]
- Anderson, L.; Holden, S.; Davis, B.; Prescott, E.; Charrier, C.; Bunce, N.; Firmin, D.; Wonke, B.; Porter, J.; Walker, J.; et al. Cardiovascular T2-star (T2*) magnetic resonance for the early diagnosis of myocardial iron overload. Eur. Heart J. 2001, 22, 2171–2179. [Google Scholar] [CrossRef] [PubMed]
- Pennell, D.J. T2* Magnetic Resonance and Myocardial Iron in Thalassemia. Ann. N. Y. Acad. Sci. 2005, 1054, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Kolnagou, A.; Yazman, D.; Economides, C.; Eracleous, E.; Kontoghiorghes, G.J. Uses and Limitations of Serum Ferritin, Magnetic Resonance Imaging T2 and T2* in the Diagnosis of Iron Overload and in the Ferrikinetics of Normalization of the Iron Stores in Thalassemia Using the International Committee on Chelation Deferiprone/Deferoxamine Combination Protocol. Hemoglobin 2009, 33, 312–322. [Google Scholar] [CrossRef]
- Rahav, G.; Volach, V.; Shapiro, M.; Rund, D.; Rachmilewitz, E.A.; Goldfarb, A. Severe infections in thalassaemic patients: Prevalence and predisposing factors. Br. J. Haematol. 2006, 133, 667–674. [Google Scholar] [CrossRef]
- Weinberg, E.D. Iron availability and infection. Biochim. Biophys. Acta (BBA) Gen. Subj. 2009, 1790, 600–605. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J.; Kolnagou, A.; Skiada, A.; Petrikkos, G. The Role of Iron and Chelators on Infections in Iron Overload and Non Iron Loaded Conditions: Prospects for the Design of New Antimicrobial Therapies. Hemoglobin 2010, 34, 227–239. [Google Scholar] [CrossRef]
- Kolnagou, A.; Michaelides, Y.; Kontoghiorghe, C.N.; Kontoghiorghes, G.J. The importance of spleen, spleen iron, and splenectomy for determining total body iron load, ferrikinetics, and iron toxicity in thalassemia major patients. Toxicol. Mech. Methods 2013, 23, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Skandalis, K.; Vlachos, C.; Pliakou, X.; Gaitanis, G.; Kapsali, E.; Bassukas, I.D. Higher Serum Ferritin Levels Correlate with an Increased Risk of Cutaneous Morbidity in Adult Patients with β-Thalassemia: A Single-Center Retrospective Study. Acta Haematol. 2016, 135, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Anderson, L.J.; Westwood, M.A.; Prescott, E.; Walker, J.M.; Pennell, D.J.; Wonke, B. Development of Thalassaemic Iron Overload Cardiomyopathy despite Low Liver Iron Levels and Meticulous Compliance to Desferrioxamine. Acta Haematol. 2006, 115, 106–108. [Google Scholar] [CrossRef] [PubMed]
- Kolnagou, A.; Economides, C.; Eracleous, E.; Kontoghiorghes, G.J. Low Serum Ferritin Levels are Misleading for Detecting Cardiac Iron Overload and Increase the Risk of Cardiomyopathy in Thalassemia Patients. The Importance of Cardiac Iron Overload Monitoring Using Magnetic Resonance Imaging T2 and T2*. Hemoglobin 2006, 30, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Papakonstantinou, O.; Alexopoulou, E.; Economopoulos, N.; Benekos, O.; Kattamis, A.; Kostaridou, S.; Ladis, V.; Efstathopoulos, E.; Gouliamos, A.; Kelekis, N.L. Assessment of iron distribution between liver, spleen, pancreas, bone marrow, and myocardium by means of R2 relaxometry with MRI in patients with beta-thalassemia major. J. Magn. Reson. Imaging 2009, 29, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Kolnagou, A.; Natsiopoulos, K.; Kleanthous, M.; Ioannou, A.; Kontoghiorghes, G.J. Liver iron and serum ferritin levels are misleading for estimating cardiac, pancreatic, splenic and total body iron load in thalassemia patients: Factors influencing the heterogenic distribution of excess storage iron in organs as identified by MRI T2*. Toxicol. Mech. Methods 2013, 23, 48–56. [Google Scholar] [CrossRef]
- Wood, J.C. Diagnosis and management of transfusion iron overload: The role of imaging. Am. J. Hematol. 2007, 82 (Suppl. 12), 1132–1135. [Google Scholar] [CrossRef]
- Chacko, J.; Pennell, D.J.; Tanner, M.A.; Hamblin, T.J.; Wonke, B.; Levy, T.; Thomas, P.W.; Killick, S.B. Myocardial iron loading by magnetic resonance imaging T2* in good prognostic myelodysplastic syndrome patients on long-term blood transfusions. Br. J. Haematol. 2007, 138, 587–593. [Google Scholar] [CrossRef]
- Bs, C.J.B.; Coates, T.D.; Wood, J.C. Spleen R2 and R2* in iron-overloaded patients with sickle cell disease and thalassemia major. J. Magn. Reson. Imaging 2009, 29, 357–364. [Google Scholar] [CrossRef]
- Kontoghiorghe, C.N.; Kolnagou, A.; Kontoghiorghes, G.J. Dietary and pharmacological factors affecting iron absorption in mice and man (Comment for a Letter to the editor). Haematologica 2016, 101, e120–e121. [Google Scholar] [CrossRef]
- Kolnagou, A.; Kleanthous, M.; Kontoghiorghes, G.J. Efficacy, Compliance and Toxicity Factors Are Affecting the Rate of Normalization of Body Iron Stores in Thalassemia Patients Using The Deferiprone and Deferoxamine Combination Therapy. Hemoglobin 2011, 35, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Kolnagou, A.; Kontoghiorghe, C.N.; Kontoghiorghes, G.J. Transfusion-Related Acute Lung Injury (TRALI) in two Thalassaemia Patients Caused by the Same Multiparous Blood Donor. Mediterr. J. Hematol. Infect. Dis. 2017, 9, e2017060. [Google Scholar] [PubMed]
- Barton, J.C.; Edwards, C.Q. (Eds.) Hemochromatosis: Genetics, Pathophysiology, Diagnosis and Treatment; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Feder, J.N.; Gnirke, A.; Thomas, W.; Tsuchihashi, Z.; Ruddy, D.A.; Basava, A.; Dormishian, F.; Domingo, R., Jr.; Ellis, M.C.; Fullan, A.; et al. A novel MHC class I–like gene is mutated in patients with hereditary haemochromatosis. Nat. Genet. 1996, 13, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Pietrangelo, A. Hereditary Hemochromatosis—A New Look at an Old Disease. N. Engl. J. Med. 2004, 350, 2383–2397. [Google Scholar] [CrossRef]
- Allen, K.J.; Gurrin, L.C.; Constantine, C.C.; Osborne, N.J.; Delatycki, M.B.; Nicoll, A.J.; McLaren, C.E.; Bahlo, M.; Nisselle, A.E.; Vulpe, C.D.; et al. Iron-overload-related disease in HFE hereditary hemochromatosis. N. Engl. J. Med. 2008, 358, 221–230. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J. The Vital Role Played by Deferiprone in the Transition of Thalassaemia from a Fatal to a Chronic Disease and Challenges in Its Repurposing for Use in Non-Iron-Loaded Diseases. Pharmaceuticals 2023, 16, 1016. [Google Scholar] [CrossRef]
- Tenneti, P.; Chojecki, A.; Knovich, M.A. Iron overload in the HCT patient: A review. Bone Marrow Transplant. 2021, 56, 1794–1804. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J. How to manage iron toxicity in post-allogeneic hematopoietic stem cell transplantation? Expert. Rev. Hematol. 2020, 13, 299–302. [Google Scholar] [CrossRef]
- Meloni, A.; Pistoia, L.; Gamberini, M.R.; Ricchi, P.; Cecinati, V.; Sorrentino, F.; Cuccia, L.; Allò, M.; Righi, R.; Fina, P.; et al. The Link of Pancreatic Iron with Glucose Metabolism and Cardiac Iron in Thalassemia Intermedia: A Large, Multicenter Observational Study. J. Clin. Med. 2021, 10, 5561. [Google Scholar] [CrossRef]
- Verberckmoes, B.; Dekeyzer, S.; Decaestecker, K. The Dark Pituitary: Hemochromatosis as a Lesser-Known Cause of Pituitary Dysfunction. J. Belg. Soc. Radiol. 2022, 106, 63. [Google Scholar] [CrossRef]
- Byrne, D.; Walsh, J.P.; Murphy, R.T.; McMorrow, J.; Fagan, A.J.; Kecler Poetrzyk, A.; Durand, M.E.; Norris, S.; McKiernan, S.; King, G.; et al. Cardiac Magnetic Resonance at 3.0 T in Patients with C282Y Homozygous Hereditary Hemochromatosis: Superiority of Radial and Circumferential Strain Over Cardiac T2* Measurements at Baseline and at Post Venesection Follow-up. J. Thorac. Imaging. 2022, 37, 300–306. [Google Scholar] [CrossRef]
- Zhou, Y.; Luo, J. Relationship between Iron deposition and T lymphocytes in children with β-thalassemia with haematopoietic stem cell transplantation. Front. Pediatr. 2022, 10, 939157. [Google Scholar] [CrossRef]
- Aboobacker, F.N.; Dixit, G.; Lakshmi, K.M.; Korula, A.; Abraham, A.; George, B.; Mathews, V.; Srivastava, A. Outcome of iron reduction therapy in ex-thalassemics. PLoS ONE 2021, 16, e0238793. [Google Scholar] [CrossRef]
- Drayer, B.P.; Olanow, W.; Burger, P.; Johnson, G.A.; Herfkens, R.; Riederer, S. Parkinson plus syndrome: Diagnosis using high field MR imaging of brain iron. Radiology 1986, 159, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Norfray, J.F.; Chiaradonna, N.L.; Heiser, W.J.; Song, S.H.; Manyam, B.V.; Devleschoward, A.B.; Eastwood, L.M. Brain iron in patients with Parkinson disease: MR visualization using gradient modification. Am. J. Neuroradiol. 1988, 9, 237–240. [Google Scholar]
- Brooks, D.J.; Luthert, P.; Gadian, D.; Marsden, C.D. Does signal-attenuation on high-field T2-weighted MRI of the brain reflect regional cerebral iron deposition? Observations on the relationship between regional cerebral water proton T2 values and iron levels. J. Neurol. Neurosurg. Psychiatry 1989, 52, 108–111. [Google Scholar] [CrossRef] [PubMed]
- Bartzokis, G.; Sultzer, D.; Mintz, J.; Holt, L.E.; Marx, P.; Phelan, C.K.; Marder, S.R. In vivo evaluation of brain iron in Alzheimer’s disease and normal subjects using MRI. Biol. Psychiatry 1994, 35, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Giordano, N.; Fioravanti, A.; Sancasciani, S.; Marcolongo, R.; Borghi, C. Increased storage of iron and anaemia in rheumatoid arthritis: Usefulness of desferrioxamine. BMJ 1984, 289, 961–962. [Google Scholar] [CrossRef][Green Version]
- Zhou, J.Y.; Wong, J.H.; Berman, Z.T.; Lombardi, A.F.; Chang, E.Y.; von Drygalski, A. Bleeding with iron deposition and vascular remodelling in subchondral cysts: A newly discovered feature unique to haemophilic arthropathy. Haemophilia 2021, 27, e730–e738. [Google Scholar] [CrossRef]
- Badat, M.; Kaya, B.; Telfer, P. Combination-therapy with concurrent deferoxamine and deferiprone is effective in treating resistant cardiac iron-loading in aceruloplasminaemia. Br. J. Haematol. 2015, 171, 430–432. [Google Scholar] [CrossRef]
- Gold, R.; Lenox, R.H. Is there a rationale for iron supplementation in the treatment of akathisia? A review of the evidence. J. Clin. Psychiatry 1995, 56, 476–483. [Google Scholar]
- Imon, Y.; Yamaguchi, S.; Katayama, S.; Harada, A.; Yamamura, Y.; Nakamura, S. The Effect of Age and Disease on the MR Imaging T2 Low Signal Intensity Area in the Cerebral Cortex. Nippon. Ronen Igakkai Zasshi. Jpn. J. Geriatr. 1994, 31, 697–704. [Google Scholar] [CrossRef][Green Version]
- Lodi, R.; Hart, P.E.; Rajagopalan, B.; Taylor, D.J.; Crilley, J.G.; Bradley, J.L.; Blamire, A.M.; Manners, D.; Styles, P.; Schapira, A.H.; et al. Antioxidant treatment improves in vivo cardiac and skeletal muscle bioenergetics in patients with Friedreich’s ataxia. Ann. Neurol. 2001, 49, 590–596. [Google Scholar] [CrossRef]
- Koeppen, A.H.; Dickson, A.C. Iron in the Hallervorden-Spatz syndrome. Pediatr. Neurol. 2001, 25, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Salomão, R.P.A.; Pedroso, J.L.; Gama, M.T.D.; Dutra, L.A.; Maciel, R.H.; Godeiro-Junior, C.; Chien, H.F.; Teive, H.A.G.; Cardoso, F.; Barsottini, O.G.P. A diagnostic approach for neurodegeneration with brain iron accumulation: Clinical features, genetics and brain imaging. Arq. Neuro-Psiquiatr. 2016, 74, 587–596. [Google Scholar] [CrossRef]
- Umeno, A.; Biju, V.; Yoshida, Y. In vivo ROS production and use of oxidative stress-derived biomarkers to detect the onset of diseases such as Alzheimer’s disease, Parkinson’s disease, and diabetes. Free Radic. Res. 2017, 51, 413–427. [Google Scholar] [CrossRef]
- Brar, S.; Henderson, D.; Schenck, J.; Zimmerman, E.A. Iron Accumulation in the Substantia Nigra of Patients With Alzheimer Disease and Parkinsonism. Arch. Neurol. 2009, 66, 371–374. [Google Scholar] [CrossRef]
- Wallis, L.I.; Paley, M.N.; Graham, J.M.; Grünewald, R.A.; Wignall, E.L.; Joy, H.M.; Griffiths, P.D. MRI assessment of basal ganglia iron deposition in Parkinson’s disease. J. Magn. Reson. Imaging 2008, 28, 1061–1067. [Google Scholar] [CrossRef]
- Waldvogel, D.; van Gelderen, P.; Hallett, M. Increased iron in the dentate nucleus of patients with Friedrich’s ataxia. Ann Neurol. 1999, 46, 123–125. [Google Scholar] [CrossRef]
- Iankova, V.; Karin, I.; Klopstock, T.; Schneider, S.A. Emerging Disease-Modifying Therapies in Neurodegeneration With Brain Iron Accumulation (NBIA) Disorders. Front. Neurol. 2021, 12, 629414. [Google Scholar] [CrossRef]
- Weiss, G.; Ganz, T.; Goodnough, L.T. Anemia of inflammation. Blood 2019, 133, 40–50. [Google Scholar] [CrossRef]
- Ganz, T. Anemia of Inflammation. N. Engl. J. Med. 2019, 381, 1148–1157. [Google Scholar] [CrossRef]
- Marques, O.; Weiss, G.; Muckenthaler, M.U. The role of iron in chronic inflammatory diseases: From mechanisms to treatment options in anemia of inflammation. Blood 2022, 140, 2011–2023. [Google Scholar] [CrossRef]
- Cacoub, P.; Choukroun, G.; Cohen-Solal, A.; Luporsi, E.; Peyrin-Biroulet, L.; Peoc’H, K.; Andrieu, V.; Lasocki, S.; Puy, H.; Trochu, J. Iron deficiency screening is a key issue in chronic inflammatory diseases: A call to action. J. Intern. Med. 2022, 292, 542–556. [Google Scholar] [CrossRef]
- Buliga-Finis, O.N.; Ouatu, A.; Tanase, D.M.; Gosav, E.M.; Seritean Isac, P.N.; Richter, P.; Rezus, C. Managing Anemia: Point of Convergence for Heart Failure and Chronic Kidney Disease? Life 2023, 13, 1311. [Google Scholar] [CrossRef]
- Canny, S.P.; Orozco, S.L.; Thulin, N.K.; Hamerman, J.A. Immune Mechanisms in Inflammatory Anemia. Annu. Rev. Immunol. 2023, 41, 405–429. [Google Scholar] [CrossRef]
- Pépin, M.; Klimkowicz-Mrowiec, A.; Godefroy, O.; Delgado, P.; Carriazo, S.; Ferreira, A.C.; Golenia, A.; Malyszko, J.; Grodzicki, T.; Giannakou, K.; et al. Cognitive disorders in patients with chronic kidney disease: Approaches to prevention and treatment. Eur. J. Neurol. 2023, 30, 2899–2911. [Google Scholar] [CrossRef]
- Toplicanin, A.; Toncev, L.; Zaric, V.M.; Milutinovic, A.S. Autoimmune Hemolytic Anemia in Inflammatory Bowel Disease—Report of a Case and Review of the Literature. Life 2022, 12, 1784. [Google Scholar] [CrossRef]
- Rashidi, A.; Garimella, P.S.; Al-Asaad, A.; Kharadjian, T.; Torres, M.N.; Thakkar, J. Anemia Management in the Cancer Patient With CKD and End-Stage Kidney Disease. Adv. Chronic Kidney Dis. 2022, 29, 180–187.e1. [Google Scholar] [CrossRef]
- Heras-Benito, M. Renal anemia: Current treatments and emerging molecules. Rev. Clin. Esp. 2023, 223, 433–439. [Google Scholar] [CrossRef]
- Ku, E.; Del Vecchio, L.; Eckardt, K.-U.; Haase, V.H.; Johansen, K.L.; Nangaku, M.; Tangri, N.; Waikar, S.S.; Więcek, A.; Cheung, M.; et al. Novel anemia therapies in chronic kidney disease: Conclusions from a Kidney Disease: Improving Global Outcomes (KDIGO) Controversies Conference. Kidney Int. 2023; in press. [Google Scholar] [CrossRef]
- Afsar, B.; Afsar, R.E. Hypoxia-inducible factors and essential hypertension: Narrative review of experimental and clinical data. Pharmacol. Rep. 2023, 75, 861–875. [Google Scholar] [CrossRef]
- Kobayashi, M.; Suhara, T.; Baba, Y.; Kawasaki, N.K.; Higa, J.K.; Matsui, T. Pathological Roles of Iron in Cardiovascular Disease. Curr. Drug Targets 2018, 19, 1068–1076. [Google Scholar] [CrossRef]
- Murawska, N.; Fabisiak, A.; Fichna, J. Anemia of Chronic Disease and Iron Deficiency Anemia in Inflammatory Bowel Diseases:Pathophysiology, Diagnosis, and Treatment. Inflamm. Bowel Dis. 2016, 22, 1198–1208. [Google Scholar] [CrossRef] [PubMed]
- Nataatmadja, M.S.; Francis, R. Recurrent severe hypophosphatemia following intravenous iron administration. Clin. Case Rep. 2020, 8, 243–246. [Google Scholar] [CrossRef]
- Kshirsagar, A.V.; Li, X. Long-Term Risks of Intravenous Iron in End-Stage Renal Disease Patients. Adv. Chronic Kidney Dis. 2019, 26, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Basak, T.; Kanwar, R.K. Iron imbalance in cancer: Intersection of deficiency and overload. Cancer Med. 2022, 11, 3837–3853. [Google Scholar] [CrossRef]
- Vassiliou, E.; Farias-Pereira, R. Impact of Lipid Metabolism on Macrophage Polarization: Implications for Inflammation and Tumor Immunity. Int. J. Mol. Sci. 2023, 24, 12032. [Google Scholar] [CrossRef]
- Zhou, Z.; Xu, B.; Hu, N.; Guo, Z.; Bao, W.; Shao, B.; Yang, W. Targeting the Macrophage-Ferroptosis Crosstalk: A Novel Insight into Tumor Immunotherapy. Front. Biosci. 2022, 27, 203. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Fang, T.; Wei, S.; Chai, S.; Yang, H.; Tao, M.; Cao, Y. Macrophages in melanoma: A double-edged sword and targeted therapy strategies (Review). Exp. Ther. Med. 2022, 24, 640. [Google Scholar] [CrossRef]
- Furgiuele, S.; Descamps, G.; Cascarano, L.; Boucq, A.; Dubois, C.; Journe, F.; Saussez, S. Dealing with Macrophage Plasticity to Address Therapeutic Challenges in Head and Neck Cancers. Int. J. Mol. Sci. 2022, 23, 6385. [Google Scholar] [CrossRef] [PubMed]
- Hetzel, M.; Ackermann, M.; Lachmann, N. Beyond “Big Eaters”: The Versatile Role of Alveolar Macrophages in Health and Disease. Int. J. Mol. Sci. 2021, 22, 3308. [Google Scholar] [CrossRef]
- Li, J.; Jia, Y.C.; Ding, Y.X.; Bai, J.; Cao, F.; Li, F. The crosstalk between ferroptosis and mitochondrial dynamic regulatory networks. Int. J. Biol. Sci. 2023, 19, 2756–2771. [Google Scholar] [CrossRef]
- Lee, S.; Hwang, N.; Seok, B.G.; Lee, S.; Lee, S.J.; Chung, S.W. Autophagy mediates an amplification loop during ferroptosis. Cell Death Dis. 2023, 14, 464. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Kang, R.; Tang, D.; Liu, J. Monitoring Lysosome Function in Ferroptosis. Methods Mol. Biol. 2023, 2712, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Denz, H.; Huber, P.; Landmann, R.; Orth, B.; Wachter, H.; Fuchs, D. Association between the activation of macrophages, changes of iron metabolism and the degree of anaemia in patients with malignant disorders. Eur. J. Haematol. 1992, 48, 244–248. [Google Scholar] [CrossRef]
- Cohen, L.A.; Gutierrez, L.; Weiss, A.; Leichtmann-Bardoogo, Y.; Zhang, D.L.; Crooks, D.R.; Sougrat, R.; Morgenstern, A.; Galy, B.; Hentze, M.W.; et al. Serum ferritin is derived primarily from macrophages through a non-classical secretory pathway. Blood 2010, 116, 1574–1584. [Google Scholar] [CrossRef]
- Wang, W.; Knovich, M.A.; Coffman, L.G.; Torti, F.M.; Torti, S.V. Serum ferritin: Past, present and future. Biochim. Biophys Acta 2010, 1800, 760–769. [Google Scholar] [CrossRef]
- Plays, M.; Müller, S.; Rodriguez, R. Chemistry and biology of ferritin. Metallomics 2021, 3, mfab021. [Google Scholar] [CrossRef]
- Yuan, X.M.; Li, W.; Baird, S.K.; Carlsson, M.; Melefors, Ö. Secretion of ferritin by iron-laden macrophages and influence of lipoproteins. Free Radic. Res. 2004, 38, 1133–1142. [Google Scholar] [CrossRef]
- Leftin, A.; Zhao, H.; Turkekul, M.; de Stanchina, E.; Manova, K.; Koutcher, J.A. Iron deposition is associated with differential macrophage infiltration and therapeutic response to iron chelation in prostate cancer. Sci. Rep. 2017, 7, 11632. [Google Scholar] [CrossRef]
- Yan, S.; Dong, X.; Ding, D.; Xue, J.; Wang, X.; Huang, Y.; Pan, Z.; Sun, H.; Ren, Q.; Dou, W.; et al. Iron deposition in ovarian endometriosis evaluated by magnetic resonance imaging R2* correlates with ovarian function. Reprod. Biomed. Online, 2023; in press. [Google Scholar] [CrossRef]
- Lattuada, D.; Uberti, F.; Colciaghi, B.; Morsanuto, V.; Maldi, E.; Squarzanti, D.F.; Molinari, C.; Boldorini, R.; Bulfoni, A.; Colombo, P.; et al. Fimbrial Cells Exposure to Catalytic Iron Mimics Carcinogenic Changes. Int. J. Gynecol. Cancer 2015, 25, 389–398. [Google Scholar] [CrossRef]
- Pontillo, G.; Petracca, M.; Monti, S.; Quarantelli, M.; Lanzillo, R.; Costabile, T.; Carotenuto, A.; Tortora, F.; Elefante, A.; Morra, V.B.; et al. Clinical correlates of R1 relaxometry and magnetic susceptibility changes in multiple sclerosis: A multi-parameter quantitative MRI study of brain iron and myelin. Eur Radiol. 2023, 33, 2185–2194. [Google Scholar] [CrossRef] [PubMed]
- Zachariou, V.; Bauer, C.E.; Pappas, C.; Gold, B.T. High cortical iron is associated with the disruption of white matter tracts supporting cognitive function in healthy older adults. Cereb. Cortex 2023, 33, 4815–4828. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhang, M.; Li, Y.; Li, Y.; Gong, H.; Li, J.; Zhang, Y.; Zhang, C.; Yan, F.; Sun, B.; et al. Alterations in brain iron deposition with progression of late-life depression measured by magnetic resonance imaging (MRI)-based quantitative susceptibility mapping. Quant. Imaging Med. Surg. 2022, 12, 3873–3888. [Google Scholar] [CrossRef]
- Pecorelli, A.; Franceschi, P.; Braccischi, L.; Izzo, F.; Renzulli, M.; Golfieri, R. MRI Appearance of Focal Lesions in Liver Iron Overload. Diagnostics 2022, 12, 891. [Google Scholar] [CrossRef]
- Blanusa, M.; Varnai, V.M.; Piasek, M.; Kostial, K. Chelators as Antidotes of Metal Toxicity: Therapeutic and Experimental Aspects. Curr. Med. Chem. 2005, 12, 2771–2794. [Google Scholar] [CrossRef]
- Perron, N.R.; Brumaghim, J.L. A Review of the Antioxidant Mechanisms of Polyphenol Compounds Related to Iron Binding. Cell Biochem. Biophys. 2009, 53, 75–100. [Google Scholar] [CrossRef]
- Kontoghiorghe, C.N.; Kolnagou, A.; Kontoghiorghes, G.J. Phytochelators Intended for Clinical Use in Iron Overload, Other Diseases of Iron Imbalance and Free Radical Pathology. Molecules 2015, 20, 20841–20872. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J. Advances on Chelation and Chelator Metal Complexes in Medicine. Int. J. Mol. Sci. 2020, 21, 2499. [Google Scholar] [CrossRef]
- Gulani, V.; Calamante, F.; Shellock, F.G.; Kanal, E.; Reeder, S.B. Gadolinium deposition in the brain: Summary of evidence and recommendations. Lancet Neurol. 2017, 16, 564–570. [Google Scholar] [CrossRef] [PubMed]
- García-Rodríguez, M.D.C.; Hernández-Cortés, L.M.; Mendoza-Núñez, V.M.; Arenas-Huertero, F. Effects of green tea polyphenols against metal-induced genotoxic damage: Underlying mechanistic pathways. J. Toxicol. Environ. Health Part B 2023, 1–16, in press. [Google Scholar] [CrossRef]
- Teranishi, K. Near-infrared chemiluminescence imaging of superoxide anion production in kidneys with iron3+-nitrilotriacetate-induced acute renal oxidative stress in rats. J. Photochem. Photobiol. B Biol. 2022, 228, 112391. [Google Scholar] [CrossRef]
- Yamamoto, R.; Williams, G.; Frankel, H.; Weisburger, J. 8-Hydroxyquinoline: Chronic toxicity and inhibitory effect on the carcinogenicity of N-2-fluorenylacetamide. Toxicol. Appl. Pharmacol. 1971, 19, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Konorev, D.; Yao, L.; Turesky, R.J. Multi-DNA Adduct and Abasic Site Quantitation In Vivo by Nano-Liquid Chromatography/High-Resolution Orbitrap Tandem Mass Spectrometry: Methodology for Biomonitoring Colorectal DNA Damage. Chem. Res. Toxicol. 2022, 35, 1519–1532. [Google Scholar] [CrossRef]
- Zhu, B.Z.; Xu, D.; Qin, L.; Huang, C.H.; Xie, L.N.; Mao, L.; Shao, J.; Kalyanaraman, B. An unexpected new pathway for nitroxide radical production via more reactve nitrogen-centered amidyl radical intermediate during detoxification of the carcinogenic halogenated quinones by N-alkyl hydroxamic acids. Free Radic. Biol. Med. 2020, 146, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J. New Iron Metabolic Pathways and Chelation Targeting Strategies Affecting the Treatment of All Types and Stages of Cancer. Int. J. Mol. Sci. 2022, 23, 13990. [Google Scholar] [CrossRef]
- Williams, G.M.; Yamamoto, R.S. Absence of Stainable Iron From Preneoplastic and Neoplastic Lesions in Rat Liver With 8-Hydroxyquinoline-Induced Siderosis. Gynecol. Oncol. 1972, 49, 685–692. [Google Scholar] [CrossRef]
- Li, G.; Yang, J.; Zhao, G.; Shen, Z.; Yang, K.; Tian, L.; Zhou, Q.; Chen, Y.; Huang, Y. Dysregulation of ferroptosis may involve in the development of non-small-cell lung cancer in Xuanwei area. J. Cell Mol. Med. 2021, 25, 2872–2884, Erratum in: J. Cell Mol. Med. 2022, 26, 240. [Google Scholar] [CrossRef]
- Pease, C.; Rücker, T.; Birk, T. Review of the Evidence from Epidemiology, Toxicology, and Lung Bioavailability on the Carcinogenicity of Inhaled Iron Oxide Particulates. Chem. Res. Toxicol. 2016, 29, 237–254. [Google Scholar] [CrossRef]
- Lovera-Leroux, M.; Crobeddu, B.; Kassis, N.; Petit, P.X.; Janel, N.; Baeza-Squiban, A.; Andreau, K. The iron component of particulate matter is antiapoptotic: A clue to the development of lung cancer after exposure to atmospheric pollutants? Biochimie 2015, 118, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.; Zhou, L.; Gao, W.; Li, Y. Organic additives enhance Fenton treatment of nitrobenzene at near-neutral pH. Environ. Sci. Pollut. Res. 2015, 22, 7082–7092. [Google Scholar] [CrossRef]
- Bouvard, V.; Loomis, D.; Guyton, K.Z.; Grosse, Y.; El Ghissassi, F.; Benbrahim-Tallaa, L.; Guha, N.; Mattock, H.; Straif, K. Carcinogenicity of consumption of red and processed meat. Lancet Oncol. 2015, 16, 1599–1600. [Google Scholar] [CrossRef]
- Seiwert, N.; Wecklein, S.; Demuth, P.; Hasselwander, S.; Kemper, T.A.; Schwerdtle, T.; Brunner, T.; Fahrer, J. Heme oxygenase 1 protects human colonocytes against ROS formation, oxidative DNA damage and cytotoxicity induced by heme iron, but not inorganic iron. Cell Death Dis. 2020, 11, 787. [Google Scholar] [CrossRef]
- Rizzolo-Brime, L.; Orta-Ramirez, A.; Martin, Y.P.; Jakszyn, P. Nutritional Assessment of Plant-Based Meat Alternatives: A Comparison of Nutritional Information of Plant-Based Meat Alternatives in Spanish Supermarkets. Nutrients 2023, 15, 1325. [Google Scholar] [CrossRef]
- Niedermaier, T.; Gredner, T.; Hoffmeister, M.; Mons, U.; Brenner, H. Impact of Reducing Intake of Red and Processed Meat on Colorectal Cancer Incidence in Germany 2020 to 2050—A Simulation Study. Nutrients 2023, 15, 1020. [Google Scholar] [CrossRef] [PubMed]
- Martin, O.C.B.; Olier, M.; Ellero-Simatos, S.; Naud, N.; Dupuy, J.; Huc, L.; Taché, S.; Graillot, V.; Levêque, M.; Bézirard, V.; et al. Haem iron reshapes colonic luminal environment: Impact on mucosal homeostasis and microbiome through aldehyde formation. Microbiome 2019, 7, 72. [Google Scholar] [CrossRef]
- Sasso, A.; Latella, G. Role of Heme Iron in the Association Between Red Meat Consumption and Colorectal Cancer. Nutr. Cancer 2018, 70, 1173–1183. [Google Scholar] [CrossRef] [PubMed]
- Chiang, V.S.-C.; Quek, S.-Y. The relationship of red meat with cancer: Effects of thermal processing and related physiological mechanisms. Crit. Rev. Food Sci. Nutr. 2017, 57, 1153–1173. [Google Scholar] [CrossRef]
- Bastide, N.M.; Pierre, F.H.; Corpet, D.E. Heme Iron from Meat and Risk of Colorectal Cancer: A Meta-analysis and a Review of the Mechanisms Involved. Cancer Prev. Res. 2011, 4, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Cross, A.J.; Freedman, N.D.; Ren, J.; Ward, M.H.; Hollenbeck, A.R.; Schatzkin, A.; Sinha, R.; Abnet, C.C. Meat Consumption and Risk of Esophageal and Gastric Cancer in a Large Prospective Study. Am. J. Gastroenterol. 2011, 106, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, Y. Asbestos-induced mesothelial injury and carcinogenesis: Involvement of iron and reactive oxygen species. Pathol. Int. 2022, 72, 83–95. [Google Scholar] [CrossRef]
- Ito, F.; Yanatori, I.; Maeda, Y.; Nimura, K.; Ito, S.; Hirayama, T.; Nagasawa, H.; Kohyama, N.; Okazaki, Y.; Akatsuka, S.; et al. Asbestos conceives Fe(II)-dependent mutagenic stromal milieu through ceaseless macrophage ferroptosis and β-catenin induction in mesothelium. Redox Biol. 2020, 36, 101616. [Google Scholar] [CrossRef] [PubMed]
- Chew, S.H.; Toyokuni, S. Malignant mesothelioma as an oxidative stress-induced cancer: An update. Free. Radic. Biol. Med. 2015, 86, 166–178. [Google Scholar] [CrossRef] [PubMed]
- E Duncan, K.; Cook, P.M.; Gavett, S.H.; A Dailey, L.; Mahoney, R.K.; Ghio, A.J.; Roggli, V.L.; Devlin, R.B. In vitro determinants of asbestos fiber toxicity: Effect on the relative toxicity of Libby amphibole in primary human airway epithelial cells. Part. Fibre Toxicol. 2014, 11, 2. [Google Scholar] [CrossRef] [PubMed]
- Urso, L.; Cavallari, I.; Sharova, E.; Ciccarese, F.; Pasello, G.; Ciminale, V. Metabolic rewiring and redox alterations in malignant pleural mesothelioma. Br. J. Cancer 2020, 122, 52–61. [Google Scholar] [CrossRef]
- Ou, Z.; Wen, Q.; Deng, Y.; Yu, Y.; Chen, Z.; Sun, L. Cigarette smoking is associated with high level of ferroptosis in seminal plasma and affects semen quality. Reprod. Biol. Endocrinol. 2020, 18, 55. [Google Scholar] [CrossRef]
- Zhang, W.Z.; Butler, J.J.; Cloonan, S.M. Smoking-induced iron dysregulation in the lung. Free. Radic. Biol. Med. 2019, 133, 238–247. [Google Scholar] [CrossRef]
- Takahashi, M.; Mizumura, K.; Gon, Y.; Shimizu, T.; Kozu, Y.; Shikano, S.; Iida, Y.; Hikichi, M.; Okamoto, S.; Tsuya, K.; et al. Iron-Dependent Mitochondrial Dysfunction Contributes to the Pathogenesis of Pulmonary Fibrosis. Front. Pharmacol. 2022, 12, 643980. [Google Scholar] [CrossRef]
- Fibach, E.; Rachmilewitz, E.A. Iron overload in hematological disorders. Presse Med. 2017, 46 Pt 2, e296–e305. [Google Scholar] [CrossRef] [PubMed]
- Leitch, H.A.; Fibach, E.; Rachmilewitz, E. Toxicity of iron overload and iron overload reduction in the setting of hematopoietic stem cell transplantation for hematologic malignancies. Crit. Rev. Oncol. 2017, 113, 156–170. [Google Scholar] [CrossRef] [PubMed]
- Harrison, P.; Marwah, S.S.; Hughes, R.T.; Bareford, D. Non-transferrin bound iron and neutropenia after cytotoxic chemotherapy. J. Clin. Pathol. 1994, 47, 350–352. [Google Scholar] [CrossRef]
- Motoori, S.; Hosokawa, Y.; Miyazawa, T.; Iwamoto, N.; Kase, Y.; Kitagawa, N.; Saisho, H.; Furuse, J. Two cases of colorectal cancer presenting with periodic, transient elevation of serum iron after chemotherapy including irinotecan. Gan Kagaku Ryoho Cancer Chemother 2010, 37, 2185–2188. [Google Scholar]
- Łęcka, M.; Słomka, A.; Albrecht, K.; Żekanowska, E.; Romiszewski, M.; Styczyński, J. Unbalance in Iron Metabolism in Childhood Leukemia Converges with Treatment Intensity: Biochemical and Clinical Analysis. Cancers 2021, 13, 3029. [Google Scholar] [CrossRef] [PubMed]
- Belotti, A.; Duca, L.; Borin, L.; Realini, S.; Renso, R.; Parma, M.; Pioltelli, P.; Pogliani, E.; Cappellini, M.D. Non transferrin bound iron (NTBI) in acute leukemias throughout conventional intensive chemotherapy: Kinetics of its appearance and potential predictive role in infectious complications. Leuk. Res. 2015, 39, 88–91. [Google Scholar] [CrossRef]
- Sahlstedt, L.; von Bonsdorff, L.; Ebeling, F.; Parkkinen, J.; Juvonen, E.; Ruutu, T. Non-transferrin-bound iron in haematological patients during chemotherapy and conditioning for autologous stem cell transplantation. Eur. J. Haematol. 2009, 83, 455–459. [Google Scholar] [CrossRef]
- von Bonsdorff, L.; Lindeberg, E.; Sahlstedt, L.; Lehto, J.; Parkkinen, J. Bleomycin-detectable Iron Assay for Non-Transferrin-bound Iron in Hematologic Malignancies. Clin. Chem. 2002, 48, 307–314. [Google Scholar] [CrossRef]
- Dürken, M.; Herrnring, C.; Finckh, B.; Nagel, S.; Nielsen, P.; Fischer, R.; Berger, H.M.; Moison, R.M.; Pichlmeier, U.; Kohlschütter, B.; et al. Impaired plasma antioxidative defense and increased nontransferrin-bound iron during high-dose chemotherapy and radiochemotherapy preceding bone marrow transplantation. Free Radic. Biol. Med. 2000, 28, 887–894. [Google Scholar] [CrossRef]
- Sahlstedt, L.; Ebeling, F.; Von Bonsdorff, L.; Parkkinen, J.; Ruutu, T. Non-transferrin-bound iron during allogeneic stem cell transplantation. Br. J. Haematol. 2001, 113, 836–838. [Google Scholar] [CrossRef]
- Miranda, J.L.; Mesquita, R.B.; Leite, A.; Silva, A.M.; Rangel, M.; Rangel, A.O. Non-transferrin-bound iron determination in blood serum using microsequential injection solid phase spectrometry—Proof of concept. Talanta 2023, 257, 124345. [Google Scholar] [CrossRef]
- Jóźwik, M.; Jóźwik, M.; Jóźwik, M.; Szczypka, M.; Gajewska, J.; Laskowska-Klita, T. Antioxidant defence of red blood cells and plasma in stored human blood. Clin. Chim. Acta 1997, 267, 129–142. [Google Scholar] [CrossRef]
- Ferrara, F.; Coppi, F.; Riva, R.; Ventura, P.; Ricci, A.; Mattioli, A.V.; Talarico, M.; Garuti, C.; Bevini, M.; Rochira, V.; et al. Labile plasma iron and echocardiographic parameters are associated with cardiac events in β-thalassemic patients. Eur. J. Clin. Investig. 2023, 53, e13954. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J. Iron Mobilization From Transferrin And Non-Transferrin-Bound-Iron by Deferiprone. Implications in the Treatment of Thalassemia, Anemia of Chronic Disease, Cancer and Other Conditions. Hemoglobin 2006, 30, 183–200. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J.; Weinberg, E.D. Iron: Mammalian defense systems, mechanisms of disease, and chelation therapy approaches. Blood Rev. 1995, 9, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.V.; Rajapurkar, M.M.; Baliga, R. The Role of Catalytic Iron in Acute Kidney Injury. Clin. J. Am. Soc. Nephrol. 2011, 6, 2329–2331. [Google Scholar] [CrossRef]
- Lele, S.S.; Mukhopadhyay, B.N.; Mardikar, M.M.; Patel, T.A.; Vasavada, A.K.; Banker, D.N.; Kapasi, K.D.; Chauhan, V.C.; Chawla, K.C.; Raju, S.R.; et al. Impact of catalytic iron on mortality in patients with acute coronary syndrome exposed to iodinated radiocontrast—The Iscom Study. Am. Heart J. 2013, 165, 744–751. [Google Scholar] [CrossRef]
- Rajapurkar, M.M.; Shah, S.V.; Lele, S.S.; Hegde, U.N.; Lensing, S.Y.; Gohel, K.; Mukhopadhyay, B.; Gang, S.; Eigenbrodt, M.L. Association of Catalytic Iron With Cardiovascular Disease. Am. J. Cardiol. 2012, 109, 438–442. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, M.; Nie, Y.; Yang, Y.; Tan, Y.; Du, Z.; Gao, S.; Li, W. Impact of iron overload on poor graft function after allo-HSCT in a patient with transfusion-dependent low-risk MDS: A case report and literature review. Medicine 2022, 101, e32012. [Google Scholar] [CrossRef]
- Brissot, P.; Bernard, D.G.; Brissot, E.; Loréal, O.; Troadec, M.-B. Rare anemias due to genetic iron metabolism defects. Mutat. Res. Mol. Mech. Mutagen. 2018, 777, 52–63. [Google Scholar] [CrossRef]
- Baravelli, C.M.; Aarsand, A.K.; Sandberg, S.; Tollånes, M.C. Porphyria cutanea tarda and patterns of long-term sick leave and disability pension: A 24-year nationwide matched-cohort study. Orphanet. J. Rare Dis. 2022, 17, 72. [Google Scholar] [CrossRef]
- Adams, P.C.; Horgan-Bell, C.; Walsh, S.; Sadikovic, B. Porphyria cutanea tarda associated with elevated serum ferritin, iron overload, and a bone morphogenetic protein 6 genetic variant. Can. Liver J. 2020, 3, 232–234. [Google Scholar] [CrossRef] [PubMed]
- Grace, R.F.; van Beers, E.J.; Corrons, J.-L.V.; Glader, B.; Glenthøj, A.; Kanno, H.; Kuo, K.H.M.; Lander, C.; Layton, D.M.; Pospíŝilová, D.; et al. The Pyruvate Kinase Deficiency Global Longitudinal (Peak) Registry: Rationale and study design. BMJ Open 2023, 13, e063605. [Google Scholar] [CrossRef] [PubMed]
- Julián-Serrano, S.; Yuan, F.; Barrett, M.J.; Pfeiffer, R.M.; Stolzenberg-Solomon, R.Z. Hemochromatosis, Iron Overload–Related Diseases, and Pancreatic Cancer Risk in the Surveillance, Epidemiology, and End Results (SEER)-Medicare. Cancer Epidemiol. Biomark. Prev. 2021, 30, 2136–2139. [Google Scholar] [CrossRef] [PubMed]
- Asleh, M.; Levitas, A.; Daniel, S.; Abu-Quider, A.; Ben-Harosh, M.; Kapelushnik, J. Hepatic and cardiac iron load as determined by MRI T2* in patients with congenital dyserythropoietic anemia type I. Ann. Hematol. 2020, 99, 2507–2512. [Google Scholar] [CrossRef]
- Beutler, E.; Felitti, V.; Gelbart, T.; Ho, N. Genetics of iron storage and hemochromatosis. Drug Metab. Dispos. 2001, 29 Pt 2, 495–499. [Google Scholar]
- Duan, G.; Li, J.; Duan, Y.; Zheng, C.; Guo, Q.; Li, F.; Zheng, J.; Yu, J.; Zhang, P.; Wan, M.; et al. Mitochondrial Iron Metabolism: The Crucial Actors in Diseases. Molecules 2022, 28, 29. [Google Scholar] [CrossRef] [PubMed]
- Pagano, G.; Talamanca, A.A.; Castello, G.; Cordero, M.D.; d’Ischia, M.; Gadaleta, M.N.; Pallardó, F.V.; Petrović, S.; Tiano, L.; Zatterale, A. Oxidative stress and mitochondrial dysfunction across broad-ranging pathologies: Toward mitochondria-targeted clinical strategies. Oxid. Med. Cell. Longev. 2014, 2014, 541230. [Google Scholar] [CrossRef]
- Uda, K.; Yashiro, M. Idiopathic Pulmonary Hemosiderosis. N. Engl. J. Med. 2023, 388, 2179. [Google Scholar] [CrossRef]
- Guo, Q.; Qian, C.; Qian, Z.-M. Iron metabolism and atherosclerosis. Trends Endocrinol. Metab. 2023, 34, 404–413. [Google Scholar] [CrossRef]
- Dempke, W.C.M.; Zielinski, R.; Winkler, C.; Silberman, S.; Reuther, S.; Priebe, W. Anthracycline-induced cardiotoxicity—Are we about to clear this hurdle? Eur. J. Cancer 2023, 185, 94–104. [Google Scholar] [CrossRef]
- Jones, I.C.; Dass, C.R. Doxorubicin-induced cardiotoxicity: Causative factors and possible interventions. J. Pharm. Pharmacol. 2022, 74, 1677–1688. [Google Scholar] [CrossRef]
- Sheibani, M.; Azizi, Y.; Shayan, M.; Nezamoleslami, S.; Eslami, F.; Farjoo, M.H.; Dehpour, A.R. Doxorubicin-Induced Cardiotoxicity: An Overview on Pre-clinical Therapeutic Approaches. Cardiovasc. Toxicol. 2022, 22, 292–310. [Google Scholar] [CrossRef]
- Menon, A.V.; Kim, J. Iron Promotes Cardiac Doxorubicin Retention and Toxicity Through Downregulation of the Mitochondrial Exporter ABCB8. Front. Pharmacol. 2022, 13, 817951. [Google Scholar] [CrossRef] [PubMed]
- Mody, H.; Vaidya, T.R.; Ait-Oudhia, S. In vitro to clinical translational pharmacokinetic/pharmacodynamic modeling of doxorubicin (DOX) and dexrazoxane (DEX) interactions: Safety assessment and optimization. Sci. Rep. 2023, 13, 3100. [Google Scholar] [CrossRef]
- Rahimi, P.; Barootkoob, B.; ElHashash, A.; Nair, A. Efficacy of Dexrazoxane in Cardiac Protection in Pediatric Patients Treated With Anthracyclines. Cureus 2023, 15, e37308. [Google Scholar] [CrossRef] [PubMed]
- de Baat, E.C.; van Dalen, E.C.; Mulder, R.L.; Hudson, M.M.; Ehrhardt, M.J.; Engels, F.K.; Feijen, E.A.M.; Grotenhuis, H.B.; Leerink, J.M.; Kapusta, L.; et al. Primary cardioprotection with dexrazoxane in patients with childhood cancer who are expected to receive anthracyclines: Recommendations from the International Late Effects of Childhood Cancer Guideline Harmonization Group. Lancet Child Adolesc. Health 2022, 6, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, T.C.; Suntharalingam, K.; Lippard, S.J. The next generation of platinum drugs: Targeted Pt (II) agents, nanoparticle delivery, and Pt(IV) Prodrugs. Chem. Rev. 2016, 116, 3436–3486. [Google Scholar] [CrossRef]
- Peng, H.; Jin, H.; Zhuo, H.; Huang, H. Enhanced antitumor efficacy of cisplatin for treating ovarian cancer in vitro and in vivo via transferrin binding. Oncotarget 2017, 8, 45597–45611. [Google Scholar] [CrossRef]
- Aggarwal, A.; Dinda, A.K.; Mukhopadhyay, C.K. Effect of Cisplatin on Renal Iron Homeostasis Components: Implication in Nephropathy. ACS Omega 2022, 7, 27804–27817. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, J.; Ma, X.; Wang, D.; Ma, H.; Chang, Y.; Nie, G.; Jia, L.; Duan, X.; Liang, X.-J. Specific Hemosiderin Deposition in Spleen Induced by a Low Dose of Cisplatin: Altered Iron Metabolism and Its Implication as an Acute Hemosiderin Formation Model. Curr. Drug Metab. 2010, 11, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Makhdoumi, P.; Abnous, K.; Mehri, S.; Etemad, L.; Imenshahidi, M.; Karimi, G. Oral deferiprone administration ameliorates cisplatin-induced nephrotoxicity in rats. J. Pharm. Pharmacol. 2018, 70, 1357–1368. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J.; Jackson, M.J.; Lunec, J. In Vitro Screening of Iron Chelators Using Models of Free Radical Damage. Free. Radic. Res. Commun. 1986, 2, 115–124. [Google Scholar] [CrossRef]
- Crisponi, G.; Nurchi, V.M.; Lachowicz, J.I.; Crespo-Alonso, M.; Zoroddu, M.A.; Peana, M. Kill or cure: Misuse of chelation therapy for human diseases. Coord. Chem. Rev. 2015, 284, 278–285. [Google Scholar] [CrossRef]
- Choi, J.J.; McCarthy, M.W. Cefiderocol: A novel siderophore cephalosporin. Expert Opin. Investig. Drugs 2018, 27, 193–197. [Google Scholar] [CrossRef]
- Robins-Browne, R.; Prpic, J. Desferrioxamine and systemic yersiniosis. Lancet 1983, 322, 1372. [Google Scholar] [CrossRef]
- Boelaert, J.R.; Fenves, A.Z.; Coburn, J.W. Deferoxamine Therapy and Mucormycosis in Dialysis Patients: Report of an International Registry. Am. J. Kidney Dis. 1991, 18, 660–667. [Google Scholar] [CrossRef]
- Brock, J.H.; Licéaga, J.; Kontoghiorghes, G.J. The effect of synthetic iron chelators on bacterial growth in human serum. FEMS Microbiol. Immunol. 1988, 1, 55–60. [Google Scholar] [CrossRef]
- Djaldetti, M.; Fishman, P.; Notti, I.; Bessler, H. The effect of tetracycline administration on iron absorption in mice. Biomedicine 1981, 35, 150–152. [Google Scholar]
- Madaan, K.; Kaushik, D.; Verma, T. Hydroxyurea: A key player in cancer chemotherapy. Expert Rev. Anticancer. Ther. 2012, 2, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Konstantinou, E.; Pashalidis, I.; Kolnagou, A.; Kontoghiorghes, G.J. Interactions Of Hydroxycarbamide (Hydroxyurea) With Iron And Copper: Implications On Toxicity and Therapeutic Strategies. Hemoglobin 2011, 35, 237–246. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J.; Kolnagou, A.; Kontoghiorghe, C.N.; Mourouzidis, L.; Timoshnikov, V.A.; Polyakov, N.E. Trying to Solve the Puzzle of the Interaction of Ascorbic Acid and Iron: Redox, Chelation and Therapeutic Implications. Medicines 2020, 7, 45. [Google Scholar] [CrossRef] [PubMed]
- Ohno, S.; Ohno, Y.; Suzuki, N.; Soma, G.-I.; Inoue, M. High-dose vitamin C (ascorbic acid) therapy in the treatment of patients with advanced cancer. Anticancer. Res. 2009, 29, 809–815. [Google Scholar] [PubMed]
- Nielsen, T.K.; Højgaard, M.; Andersen, J.T.; Poulsen, H.E.; Lykkesfeldt, J.; Mikines, K.J. Elimination of Ascorbic Acid After High-Dose Infusion in Prostate Cancer Patients: A Pharmacokinetic Evaluation. Basic Clin. Pharmacol. Toxicol. 2015, 116, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Levine, M.; Conry-Cantilena, C.; Wang, Y.; Welch, R.W.; Washko, P.W.; Dhariwal, K.R.; Park, J.B.; Lazarev, A.; Graumlich, J.F.; King, J.; et al. Vitamin C pharmacokinetics in healthy volunteers: Evidence for a recommended dietary allowance. Proc. Natl. Acad. Sci. USA 1996, 93, 3704–3709. [Google Scholar] [CrossRef]
- Tarng, D.; Huang, T.; Wei, Y. Erythropoietin and iron: The role of ascorbic acid. Nephrol. Dial. Transplant. 2001, 16, 35–39. [Google Scholar] [CrossRef][Green Version]
- Sultana, T.; DeVita, M.V.; Michelis, M.F. Oral vitamin C supplementation reduces erythropoietin requirement in hemodialysis patients with functional iron deficiency. Int. Urol. Nephrol. 2016, 48, 1519–1524. [Google Scholar] [CrossRef]
- Sourabh, S.; Bhatia, P.; Jain, R. Favourable improvement in haematological parameters in response to oral iron and vitamin C combination in children with Iron Refractory Iron Deficiency Anemia (IRIDA) phenotype. Blood Cells Mol. Dis. 2019, 75, 26–29. [Google Scholar] [CrossRef]
- Scheers, N.; Sandberg, A.-S. Iron Transport through Ferroportin Is Induced by Intracellular Ascorbate and Involves IRP2 and HIF2α. Nutrients 2014, 6, 249–260. [Google Scholar] [CrossRef]
- Timoshnikov, V.A.; Selyutina, O.Y.; Polyakov, N.E.; Didichenko, V.; Kontoghiorghes, G.J. Mechanistic Insights of Chelator Complexes with Essential Transition Metals: Antioxidant/Pro-Oxidant Activity and Applications in Medicine. Int. J. Mol. Sci. 2022, 23, 1247. [Google Scholar] [CrossRef]
- Timoshnikov, V.A.; Kobzeva, T.V.; Polyakov, N.E.; Kontoghiorghes, G.J. Redox Interactions of Vitamin C and Iron: Inhibition of the Pro-Oxidant Activity by Deferiprone. Int. J. Mol. Sci. 2020, 21, 3967. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J. Deferiprone and Iron-Maltol: Forty Years since Their Discovery and Insights into Their Drug Design, Development, Clinical Use and Future Prospects. Int. J. Mol. Sci. 2023, 24, 4970. [Google Scholar] [CrossRef] [PubMed]
- Ueda, K.; Kim, H.J.; Zhao, J.; Song, Y.; Dunaief, J.L.; Sparrow, J.R. Iron promotes oxidative cell death caused by bisretinoids of retina. Proc. Natl. Acad. Sci. USA 2018, 115, 4963–4968. [Google Scholar] [CrossRef] [PubMed]
- Selyutina, O.Y.; Timoshnikov, V.A.; Polyakov, N.E.; Kontoghiorghes, G.J. Metal Complexes of Omadine (N-Hydroxypyridine-2-thione): Differences of Antioxidant and Pro-Oxidant Behavior in Light and Dark Conditions with Possible Toxicity Implications. Molecules 2023, 28, 4210. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J. Chemical, Pharmacological, Toxicological and Therapeutic Advances of Deferiprone (L1) and Other Iron and Aluminium Chelators. Arch. Toxicol. Suppl. 1996, 18, 202–214. [Google Scholar] [CrossRef] [PubMed]
- Campbell, N.; Hasinoff, B. Iron supplements: A common cause of drug interactions. Br. J. Clin. Pharmacol. 1991, 31, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Carota, G.; Distefano, A.; Spampinato, M.; Giallongo, C.; Broggi, G.; Longhitano, L.; Palumbo, G.A.; Parenti, R.; Caltabiano, R.; Giallongo, S.; et al. Neuroprotective Role of α-Lipoic Acid in Iron-Overload-Mediated Toxicity and Inflammation in In Vitro and In Vivo Models. Antioxidants 2022, 11, 1596. [Google Scholar] [CrossRef] [PubMed]
- Ott, D.B.; Hartwig, A.; Stillman, M.J. Competition between Al3+ and Fe3+ binding to human transferrin and toxicological implications: Structural investigations using ultra-high resolution ESI MS and CD spectroscopy. Metallomics 2019, 11, 968–981. [Google Scholar] [CrossRef] [PubMed]
- Crapper McLachlan, D.R.; Dalton, A.J.; Kruck, T.P.; Bell, M.Y.; Smith, W.L.; Kalow, W.; Andrews, D.F. Intramuscular desferrioxamine in patients with Alzheimer’s disease. Lancet 1991, 337, 1304–1308. [Google Scholar] [CrossRef]
- Nemeth, E.; Ganz, T. Hepcidin and Iron in Health and Disease. Annu. Rev. Med. 2023, 74, 261–277. [Google Scholar] [CrossRef]
- Ruchala, P.; Nemeth, E. The pathophysiology and pharmacology of hepcidin. Trends Pharmacol. Sci. 2014, 35, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Arezes, J.; Nemeth, E. Hepcidin and iron disorders: New biology and clinical approaches. Int. J. Lab. Hematol. 2015, 37 (Suppl. 1), 92–98. [Google Scholar] [CrossRef] [PubMed]
- Hawula, Z.J.; Wallace, D.F.; Subramaniam, V.N.; Rishi, G. Therapeutic Advances in Regulating the Hepcidin/Ferroportin Axis. Pharmaceuticals 2019, 12, 170. [Google Scholar] [CrossRef]
- Rishi, G.; Subramaniam, V.N. Biology of the iron efflux transporter, ferroportin. Adv. Protein. Chem. Struct. Biol. 2021, 123, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, S.; Belo, L.; Reis, F.; Santos-Silva, A. Iron therapy in chronic kidney disease: Recent changes, benefits and risks. Blood Rev. 2016, 30, 65–72. [Google Scholar] [CrossRef]
- Chekmarev, J.; Azad, M.G.; Richardson, D.R. The Oncogenic Signaling Disruptor, NDRG1: Molecular and Cellular Mechanisms of Activity. Cells 2021, 10, 2382. [Google Scholar] [CrossRef]
- Zhang, P.; Tchou-Wong, K.M.; Costa, M. Egr-1 mediates hypoxia-inducible transcription of the NDRG1 gene through an overlapping Egr-1/Sp1 binding site in the promoter. Cancer Res. 2007, 67, 9125–9133. [Google Scholar] [CrossRef]
- Orfanou, I.-M.; Argyros, O.; Papapetropoulos, A.; Tseleni-Balafouta, S.; Vougas, K.; Tamvakopoulos, C. Discovery and Pharmacological Evaluation of STEAP4 as a Novel Target for HER2 Overexpressing Breast Cancer. Front. Oncol. 2021, 11, 608201. [Google Scholar] [CrossRef]
- Wang, C.Y.; Canali, S.; Bayer, A.; Dev, S.; Agarwal, A.; Babitt, J.L. Iron, erythropoietin, and inflammation regulate hepcidin in Bmp2-deficient mice, but serum iron fails to induce hepcidin in Bmp6-deficient mice. Am. J. Hematol. 2019, 94, 240–248. [Google Scholar] [CrossRef]
- Camaschella, C.; Pagani, A.; Silvestri, L.; Nai, A. The mutual crosstalk between iron and erythropoiesis. Int. J. Hematol. 2022, 116, 182–191. [Google Scholar] [CrossRef]
- Kolnagou, A.; Kleanthous, M.; Kontoghiorghes, G.J. Reduction of body iron stores to normal range levels in thalassaemia by using a deferiprone/deferoxamine combination and their maintenance thereafter by deferiprone monotherapy. Eur. J. Haematol. 2010, 85, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Farmaki, K.; Tzoumari, I.; Pappa, C.; Chouliaras, G.; Berdoukas, V. Normalisation of total body iron load with very intensive combined chelation reverses cardiac and endocrine complications of thalassaemia major. Br. J. Haematol. 2010, 148, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Kolnagou, A.; Kontoghiorghe, C.N.; Kontoghiorghes, G.J. Prevention of Iron Overload and Long Term Maintenance of Normal Iron Stores in Thalassaemia Major Patients using Deferiprone or Deferiprone Deferoxamine Combination. Drug Res. 2017, 67, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J.; Kleanthous, M.; Kontoghiorghe, C.N. The History of Deferiprone (L1) and the Paradigm of the Complete Treatment of Iron Overload in Thalassaemia. Mediterr. J. Hematol. Infect. Dis. 2020, 12, e2020011. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A. Transfusional and other iron-loaded categories of patients |
|---|
| (a) Thalassemia |
| β-thalassemia major |
| β-thalassaemia intermedia: Heterozygous mild β+/severe β+. Homozygous β (o) or β+ with other genetic factors leading to increased γ chain production. Homozygous normal Hb A2 β thalassemia (type I). Heterozygous β (o) or β+/Normal Hb A2 β thalassemia (type I). |
| δβ thalasemia and High Persistent Fetal Hemoglobin (HPFH): Homozygous (G) γ, δβ thalasemia. Heterozygous β (o) or β+ thalasemia/δβ thalasemia. Heterozygous Greek HPFH/β thalassemia. |
| β or δβ thalasemia with structural hemoglobin variants: Hemoglobin S,C,E in association with β (o), β+ or δβ thalasemia. Hb E thalassemia. Hb E/β thalassemia |
| Severe heterozygous β thalasemia |
| Interactions of α and β thalasemia: α- and β-thalassemia |
| Interactions of α thalasemia and Hemoglobin S |
| (b) Sickle cell anemia |
| (c) Hematologic categories: Myelodysplastic syndrome, aplastic anemia, Fanconi anemia, Blackfan-Diamond anemia, pyruvate kinase deficiency, hematopoietic stem cell transplantation, hereditary sideroblastic anemia. |
| (d) Oncologic and other categories: Cancer, renal dialysis |
| B. Idiopathic hemochromatosis and other diseases related to increased iron absorption |
| Iron overload in liver disease. Iron poisoning. Dietary or iatrogenic iron intake. |
| C. Hemolytic and other anemias: Hemolytic diseases. Pernicious anemia. Prophyria cutanea tarda. Hereditary spherocytosis. Congenital dyserythropoietic anemia. Congenital atransferrinemia. Hereditary hypochromic anemia. |
| D. Anemia of chronic disease: Inflammatory diseases, chronic kidney disease, cancer etc. |
| E. Neurodegenerative diseases: Friedreich’s ataxia, Alzheimer’s disease, Parkinson’s disease, etc. |
| F. Drug and natural product iron toxicity: Drug toxicity, e.g., doxorubicin. Natural product toxicity, e.g., ascorbic acid |
| G. Dietary toxins: Processed meat carcinogens. Red meat carcinogens. Nitroso heme complexes. Other heme and iron complexes. |
| H. Environmental toxins: Toxic iron particles from gas fumes. Asbestos. |
| I. Free radical toxicity: Diseases of free radical pathology. Ischemia reperfusion injury. Storage of organs for transplantation. |
| K. Ferroptotic cell death related diseases: Almost all common and rare diseases. |
| L. Iron toxicity associated with abnormalities related to regulatory proteins of iron metabolism |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kontoghiorghes, G.J. Iron Load Toxicity in Medicine: From Molecular and Cellular Aspects to Clinical Implications. Int. J. Mol. Sci. 2023, 24, 12928. https://doi.org/10.3390/ijms241612928
Kontoghiorghes GJ. Iron Load Toxicity in Medicine: From Molecular and Cellular Aspects to Clinical Implications. International Journal of Molecular Sciences. 2023; 24(16):12928. https://doi.org/10.3390/ijms241612928
Chicago/Turabian StyleKontoghiorghes, George J. 2023. "Iron Load Toxicity in Medicine: From Molecular and Cellular Aspects to Clinical Implications" International Journal of Molecular Sciences 24, no. 16: 12928. https://doi.org/10.3390/ijms241612928
APA StyleKontoghiorghes, G. J. (2023). Iron Load Toxicity in Medicine: From Molecular and Cellular Aspects to Clinical Implications. International Journal of Molecular Sciences, 24(16), 12928. https://doi.org/10.3390/ijms241612928