
Cell Cycle Regulation by NF-YC in Drosophila Eye Imaginal Disc: Implications for Synchronization in the Non-Proliferative Region


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Depletion of NF-YC Leads to Ectopic Expression of CycA and CycB in the Non-Proliferative Region
2.2. NF-YC Is Not Essential for Expression of Dpp, Hh, and Hth in the Late 3rd Instar Eye Imaginal Disc
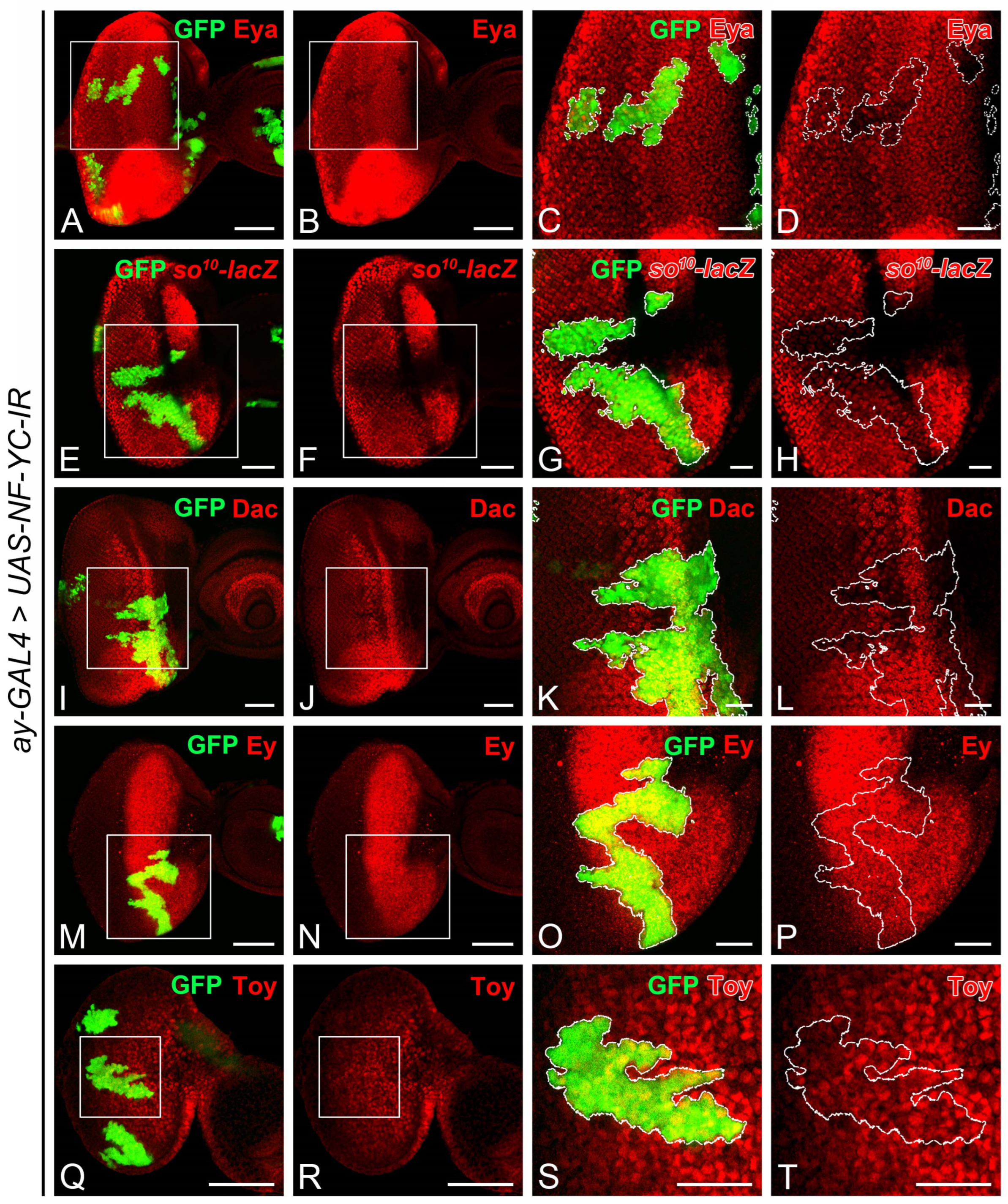
2.3. NF-YC Depletion Reduces Expression of Transcriptional Cofactor Eyes Absent
2.4. Eya Overexpression Rescues CycA and CycB Mislocalization Resulting from NF-YC Depletion
3. Discussion
4. Materials and Methods
4.1. Drosophila Stocks and Genetics
4.2. Drosophila Whole-Mount Immunostaining
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ready, D.F.; Hanson, T.E.; Benzer, S. Development of the Drosophila retina, a neurocrystalline lattice. Dev. Biol. 1976, 53, 217–240. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, A.; Ready, D.F. Neuronal differentiation in Drosophila ommatidium. Dev. Biol. 1987, 120, 366–376. [Google Scholar] [CrossRef]
- Campos-Ortega, J.A.; Hofbauer, A. Cell clones and pattern formation: On the lineage of photoreceptor cells in the compound eye of Drosophila. Wilehm Roux. Arch. Dev. Biol. 1977, 181, 227–245. [Google Scholar] [CrossRef] [PubMed]
- Escudero, L.M.; Freeman, M. Mechanism of G1 arrest in the Drosophila eye imaginal disc. BMC Dev. Biol. 2007, 7, 13. [Google Scholar] [CrossRef] [PubMed]
- Thomas, B.J.; Gunning, D.A.; Cho, J.; Zipursky, L. Cell cycle progression in the developing Drosophila eye: Roughex encodes a novel protein required for the establishment of G1. Cell 1994, 77, 1003–1014. [Google Scholar] [CrossRef]
- Thomas, B.J.; Zavitz, K.H.; Dong, X.; Lane, M.E.; Weigmann, K.; Finley, R.L., Jr.; Brent, R.; Lehner, C.F.; Zipursky, S.L. Roughex down-regulates G2 cyclins in G1. Genes Dev. 1997, 11, 1289–1298. [Google Scholar] [CrossRef]
- Firth, L.C.; Baker, N.E. Extracellular signals responsible for spatially regulated proliferation in the differentiating Drosophila eye. Dev. Cell 2005, 8, 541–551. [Google Scholar] [CrossRef]
- Du, W. Suppression of the rbf null mutants by a de2f1 allele that lacks transactivation domain. Development 2000, 127, 367–379. [Google Scholar] [CrossRef]
- Jarman, A.P.; Sun, Y.; Jan, L.Y.; Jan, Y.N. Role of the proneural gene, atonal, in formation of Drosophila chordotonal organs and photoreceptors. Development 1995, 121, 2019–2030. [Google Scholar] [CrossRef]
- Krek, W.; Nigg, E.A. Differential phosphorylation of vertebrate p34cdc2 kinase at the G1/S and G2/M transitions of the cell cycle: Identification of major phosphorylation sites. EMBO J. 1991, 10, 305–316. [Google Scholar] [CrossRef]
- Strausfeld, U.; Labbe, J.C.; Fesquet, D.; Cavadore, J.C.; Picard, A.; Sadhu, K.; Russell, P.; Doree, M. Dephosphorylation and activation of a p34cdc2/cyclin B complex in vitro by human CDC25 protein. Nature 1991, 351, 242–245. [Google Scholar] [CrossRef]
- Lopes, C.S.; Casares, F. Eye selector logic for a coordinated cell cycle exit. PLoS Genet. 2015, 11, e1004981. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, M.; Hafen, E. Hedgehog directly controls initiation and propagation of retinal differentiation in the Drosophila eye. Genes Dev. 1997, 11, 3254–3264. [Google Scholar] [CrossRef] [PubMed]
- Padgett, R.W.; Wozney, J.M.; Gelbart, W.M. Human BMP sequences can confer normal dorsal-ventral patterning in the Drosophila embryo. Proc. Natl. Acad. Sci. USA 1993, 90, 2905–2909. [Google Scholar] [CrossRef]
- Sampath, T.K.; Rashka, K.E.; Doctor, J.S.; Tucker, R.F.; Hoffmann, F.M. Drosophila transforming growth factor beta superfamily proteins induce endochondral bone formation in mammals. Proc. Natl. Acad. Sci. USA 1993, 90, 6004–6008. [Google Scholar] [CrossRef]
- Letsou, A.; Arora, K.; Wrana, J.L.; Simin, K.; Twombly, V.; Jamal, J.; Staehling-Hampton, K.; Hoffmann, F.M.; Gelbart, W.M.; Massague, J.; et al. Drosophila Dpp signaling is mediated by the punt gene product: A dual ligand-binding type II receptor of the TGF beta receptor family. Cell 1995, 80, 899–908. [Google Scholar] [CrossRef]
- Ruberte, E.; Marty, T.; Nellen, D.; Affolter, M.; Basler, K. An absolute requirement for both the type II and type I receptors, punt and thick veins, for dpp signaling in vivo. Cell 1995, 80, 889–897. [Google Scholar] [CrossRef] [PubMed]
- Burke, R.; Basler, K. Hedgehog-dependent patterning in the Drosophila eye can occur in the absence of Dpp signaling. Dev. Biol. 1996, 179, 360–368. [Google Scholar] [CrossRef]
- Ma, C.; Zhou, Y.; Beachy, P.A.; Moses, K. The segment polarity gene hedgehog is required for progression of the morphogenetic furrow in the developing Drosophila eye. Cell 1993, 75, 927–938. [Google Scholar] [CrossRef]
- Horsfield, J.; Penton, A.; Secombe, J.; Hoffman, F.M.; Richardson, H. decapentaplegic is required for arrest in G1 phase during Drosophila eye development. Development 1998, 125, 5069–5078. [Google Scholar] [CrossRef]
- Firth, L.C.; Bhattacharya, A.; Baker, N.E. Cell cycle arrest by a gradient of Dpp signaling during Drosophila eye development. BMC Dev. Biol. 2010, 10, 28. [Google Scholar] [CrossRef] [PubMed]
- Lopes, C.S.; Casares, F. hth maintains the pool of eye progenitors and its downregulation by Dpp and Hh couples retinal fate acquisition with cell cycle exit. Dev. Biol. 2010, 339, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, R. A survey of 178 NF-Y binding CCAAT boxes. Nucleic Acids Res. 1998, 26, 1135–1143. [Google Scholar] [CrossRef]
- Dorn, A.; Bollekens, J.; Staub, A.; Benoist, C.; Mathis, D. A multiplicity of CCAAT box-binding proteins. Cell 1987, 50, 863–872. [Google Scholar] [CrossRef]
- Li, G.; Zhao, H.; Wang, L.; Wang, Y.; Guo, X.; Xu, B. The animal nuclear factor Y: An enigmatic and important heterotrimeric transcription factor. Am. J. Cancer Res. 2018, 8, 1106–1125. [Google Scholar]
- Zwicker, J.; Lucibello, F.C.; Wolfraim, L.A.; Gross, C.; Truss, M.; Engeland, K.; Muller, R. Cell cycle regulation of the cyclin A, cdc25C and cdc2 genes is based on a common mechanism of transcriptional repression. EMBO J. 1995, 14, 4514–4522. [Google Scholar] [CrossRef] [PubMed]
- Zwicker, J.; Gross, C.; Lucibello, F.C.; Truss, M.; Ehlert, F.; Engeland, K.; Muller, R. Cell cycle regulation of cdc25C transcription is mediated by the periodic repression of the glutamine-rich activators NF-Y and Sp1. Nucleic Acids Res. 1995, 23, 3822–3830. [Google Scholar] [CrossRef] [PubMed]
- Korner, K.; Jerome, V.; Schmidt, T.; Muller, R. Cell cycle regulation of the murine cdc25B promoter: Essential role for nuclear factor-Y and a proximal repressor element. J. Biol. Chem. 2001, 276, 9662–9669. [Google Scholar] [CrossRef] [PubMed]
- Bolognese, F.; Wasner, M.; Dohna, C.L.; Gurtner, A.; Ronchi, A.; Muller, H.; Manni, I.; Mossner, J.; Piaggio, G.; Mantovani, R.; et al. The cyclin B2 promoter depends on NF-Y, a trimer whose CCAAT-binding activity is cell-cycle regulated. Oncogene 1999, 18, 1845–1853. [Google Scholar] [CrossRef]
- Hu, Q.; Maity, S.N. Stable expression of a dominant negative mutant of CCAAT binding factor/NF-Y in mouse fibroblast cells resulting in retardation of cell growth and inhibition of transcription of various cellular genes. J. Biol. Chem. 2000, 275, 4435–4444. [Google Scholar] [CrossRef]
- Manni, I.; Mazzaro, G.; Gurtner, A.; Mantovani, R.; Haugwitz, U.; Krause, K.; Engeland, K.; Sacchi, A.; Soddu, S.; Piaggio, G. NF-Y mediates the transcriptional inhibition of the cyclin B1, cyclin B2, and cdc25C promoters upon induced G2 arrest. J. Biol. Chem. 2001, 276, 5570–5576. [Google Scholar] [CrossRef] [PubMed]
- Benatti, P.; Dolfini, D.; Vigano, A.; Ravo, M.; Weisz, A.; Imbriano, C. Specific inhibition of NF-Y subunits triggers different cell proliferation defects. Nucleic Acids Res. 2011, 39, 5356–5368. [Google Scholar] [CrossRef] [PubMed]
- Gurtner, A.; Manni, I.; Fuschi, P.; Mantovani, R.; Guadagni, F.; Sacchi, A.; Piaggio, G. Requirement for down-regulation of the CCAAT-binding activity of the NF-Y transcription factor during skeletal muscle differentiation. Mol. Biol. Cell 2003, 14, 2706–2715. [Google Scholar] [CrossRef] [PubMed]
- Farina, A.; Manni, I.; Fontemaggi, G.; Tiainen, M.; Cenciarelli, C.; Bellorini, M.; Mantovani, R.; Sacchi, A.; Piaggio, G. Down-regulation of cyclin B1 gene transcription in terminally differentiated skeletal muscle cells is associated with loss of functional CCAAT-binding NF-Y complex. Oncogene 1999, 18, 2818–2827. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yoshioka, Y.; Ly, L.L.; Yamaguchi, M. Transcription factor NF-Y is involved in differentiation of R7 photoreceptor cell in Drosophila. Biol. Open 2012, 1, 19–29. [Google Scholar] [CrossRef]
- Ly, L.L.; Suyari, O.; Yoshioka, Y.; Tue, N.T.; Yoshida, H.; Yamaguchi, M. dNF-YB plays dual roles in cell death and cell differentiation during Drosophila eye development. Gene 2013, 520, 106–118. [Google Scholar] [CrossRef]
- Morey, M.; Yee, S.K.; Herman, T.; Nern, A.; Blanco, E.; Zipursky, S.L. Coordinate control of synaptic-layer specificity and rhodopsins in photoreceptor neurons. Nature 2008, 456, 795–799. [Google Scholar] [CrossRef][Green Version]
- Yoshioka, Y.; Suyari, O.; Yamada, M.; Ohno, K.; Hayashi, Y.; Yamaguchi, M. Complex interference in the eye developmental pathway by Drosophila NF-YA. Genesis 2007, 45, 21–31. [Google Scholar] [CrossRef]
- Ito, K.; Awano, W.; Suzuki, K.; Hiromi, Y.; Yamamoto, D. The Drosophila mushroom body is a quadruple structure of clonal units each of which contains a virtually identical set of neurones and glial cells. Development 1997, 124, 761–771. [Google Scholar] [CrossRef]
- Fu, W.; Baker, N.E. Deciphering synergistic and redundant roles of Hedgehog, Decapentaplegic and Delta that drive the wave of differentiation in Drosophila eye development. Development 2003, 130, 5229–5239. [Google Scholar] [CrossRef]
- Masucci, J.D.; Miltenberger, R.J.; Hoffmann, F.M. Pattern-specific expression of the Drosophila decapentaplegic gene in imaginal disks is regulated by 3′ cis-regulatory elements. Genes Dev. 1990, 4, 2011–2023. [Google Scholar] [CrossRef] [PubMed]
- Bessa, J.; Gebelein, B.; Pichaud, F.; Casares, F.; Mann, R.S. Combinatorial control of Drosophila eye development by eyeless, homothorax, and teashirt. Genes Dev. 2002, 16, 2415–2427. [Google Scholar] [CrossRef] [PubMed]
- Karandikar, U.C.; Jin, M.; Jusiak, B.; Kwak, S.; Chen, R.; Mardon, G. Drosophila eyes absent is required for normal cone and pigment cell development. PLoS ONE 2014, 9, e102143. [Google Scholar] [CrossRef] [PubMed]
- Bui, Q.T.; Zimmerman, J.E.; Liu, H.; Bonini, N.M. Molecular analysis of Drosophila eyes absent mutants reveals features of the conserved Eya domain. Genetics 2000, 155, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Amoui, M.; Zhang, Z.; Mardon, G. Dachshund and eyes absent proteins form a complex and function synergistically to induce ectopic eye development in Drosophila. Cell 1997, 91, 893–903. [Google Scholar] [CrossRef]
- Pignoni, F.; Hu, B.; Zavitz, K.H.; Xiao, J.; Garrity, P.A.; Zipursky, S.L. The eye-specification proteins So and Eya form a complex and regulate multiple steps in Drosophila eye development. Cell 1997, 91, 881–891. [Google Scholar] [CrossRef]
- Halder, G.; Callaerts, P.; Flister, S.; Walldorf, U.; Kloter, U.; Gehring, W.J. Eyeless initiates the expression of both sine oculis and eyes absent during Drosophila compound eye development. Development 1998, 125, 2181–2191. [Google Scholar] [CrossRef]
- Ostrin, E.J.; Li, Y.; Hoffman, K.; Liu, J.; Wang, K.; Zhang, L.; Mardon, G.; Chen, R. Genome-wide identification of direct targets of the Drosophila retinal determination protein Eyeless. Genome Res. 2006, 16, 466–476. [Google Scholar] [CrossRef]
- Casimiro, M.C.; Crosariol, M.; Loro, E.; Li, Z.; Pestell, R.G. Cyclins and cell cycle control in cancer and disease. Genes Cancer 2012, 3, 649–657. [Google Scholar] [CrossRef]
- Edgar, B.A.; O’Farrell, P.H. Genetic control of cell division patterns in the Drosophila embryo. Cell 1989, 57, 177–187. [Google Scholar] [CrossRef]
- Mozer, B.A.; Easwarachandran, K. Pattern formation in the absence of cell proliferation: Tissue-specific regulation of cell cycle progression by string (stg) during Drosophila eye development. Dev. Biol. 1999, 213, 54–69. [Google Scholar] [CrossRef] [PubMed]
- Avedisov, S.N.; Krasnoselskaya, I.; Mortin, M.; Thomas, B.J. Roughex mediates G(1) arrest through a physical association with cyclin A. Mol. Cell Biol. 2000, 20, 8220–8229. [Google Scholar] [CrossRef] [PubMed]
- Penton, A.; Selleck, S.B.; Hoffmann, F.M. Regulation of cell cycle synchronization by decapentaplegic during Drosophila eye development. Science 1997, 275, 203–206. [Google Scholar] [CrossRef]
- Curtiss, J.; Mlodzik, M. Morphogenetic furrow initiation and progression during eye development in Drosophila: The roles of decapentaplegic, hedgehog and eyes absent. Development 2000, 127, 1325–1336. [Google Scholar] [CrossRef]
- Bui, Q.T.; Zimmerman, J.E.; Liu, H.; Gray-Board, G.L.; Bonini, N.M. Functional analysis of an eye enhancer of the Drosophila eyes absent gene: Differential regulation by eye specification genes. Dev. Biol. 2000, 221, 355–364. [Google Scholar] [CrossRef][Green Version]
- Blackman, R.K.; Sanicola, M.; Raftery, L.A.; Gillevet, T.; Gelbart, W.M. An extensive 3′ cis-regulatory region directs the imaginal disk expression of decapentaplegic, a member of the TGF-beta family in Drosophila. Development 1991, 111, 657–666. [Google Scholar] [CrossRef]
- Niimi, T.; Seimiya, M.; Kloter, U.; Flister, S.; Gehring, W.J. Direct regulatory interaction of the eyeless protein with an eye-specific enhancer in the sine oculis gene during eye induction in Drosophila. Development 1999, 126, 2253–2260. [Google Scholar] [CrossRef]
- Lin, M.D.; Lee, H.T.; Wang, S.C.; Li, H.R.; Hsien, H.L.; Cheng, K.W.; Chang, Y.D.; Huang, M.L.; Yu, J.K.; Chen, Y.H. Expression of phosphatase of regenerating liver family genes during embryogenesis: An evolutionary developmental analysis among Drosophila, amphioxus, and zebrafish. BMC Dev. Biol. 2013, 13, 18. [Google Scholar] [CrossRef]
- Kurant, E.; Pai, C.Y.; Sharf, R.; Halachmi, N.; Sun, Y.H.; Salzberg, A. Dorsotonals/homothorax, the Drosophila homologue of meis1, interacts with extradenticle in patterning of the embryonic PNS. Development 1998, 125, 1037–1048. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avellino, A.; Peng, C.-H.; Lin, M.-D. Cell Cycle Regulation by NF-YC in Drosophila Eye Imaginal Disc: Implications for Synchronization in the Non-Proliferative Region. Int. J. Mol. Sci. 2023, 24, 12203. https://doi.org/10.3390/ijms241512203
Avellino A, Peng C-H, Lin M-D. Cell Cycle Regulation by NF-YC in Drosophila Eye Imaginal Disc: Implications for Synchronization in the Non-Proliferative Region. International Journal of Molecular Sciences. 2023; 24(15):12203. https://doi.org/10.3390/ijms241512203
Chicago/Turabian StyleAvellino, Anthony, Chen-Huan Peng, and Ming-Der Lin. 2023. "Cell Cycle Regulation by NF-YC in Drosophila Eye Imaginal Disc: Implications for Synchronization in the Non-Proliferative Region" International Journal of Molecular Sciences 24, no. 15: 12203. https://doi.org/10.3390/ijms241512203
APA StyleAvellino, A., Peng, C.-H., & Lin, M.-D. (2023). Cell Cycle Regulation by NF-YC in Drosophila Eye Imaginal Disc: Implications for Synchronization in the Non-Proliferative Region. International Journal of Molecular Sciences, 24(15), 12203. https://doi.org/10.3390/ijms241512203