
Recent Progress in the Characterization, Synthesis, Delivery Procedures, Treatment Strategies, and Precision of Antimicrobial Peptides

 ,
,  ,
,  , , ,
, , ,  and
and 
Abstract
1. Introduction
1.1. Antimicrobial Peptide Synthesis and Characterization
1.2. AMPs in the Era of Antibiotic Resistance
1.3. AMPs as Novel Antibacterial Agents with High Selectivity Indexes (SI)
2. AMPs with Anti-Inflammatory Properties
2.1. Role of AMPs in Medicine
2.2. Nanotechnology-Based Delivery Systems for AMPs
3. AMP-Doped and AMP-Releasing Biomaterials and Coating
3.1. AMPS and Mesenchymal Stem Cells
3.2. AMPs and Biofilms
3.3. Cytotoxicity of AMPs
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q. The antimicrobial peptides and their potential clinical applications. Am. J. Transl. Res. 2019, 11, 3919. [Google Scholar]
- Magana, M.; Pushpanathan, M.; Santos, A.L.; Leanse, L.; Fernandez, M.; Ioannidis, A.; Giulianotti, M.A.; Apidianakis, Y.; Bradfute, S.; Ferguson, A.L. The value of antimicrobial peptides in the age of resistance. Lancet Infect. Dis. 2020, 20, e216–e230. [Google Scholar] [CrossRef]
- Shen, X.; Zhang, Y.; Mao, Q.; Huang, Z.; Yan, T.; Lin, T.; Chen, W.; Wang, Y.; Cai, X.; Liang, Y. Peptide–Polymer Conjugates: A Promising Therapeutic Solution for Drug-Resistant Bacteria. Int. J. Polym. Sci. 2022, 2022, 7610951. [Google Scholar] [CrossRef]
- Qin, S.; Xiao, W.; Zhou, C.; Pu, Q.; Deng, X.; Lan, L.; Liang, H.; Song, X.; Wu, M. Pseudomonas aeruginosa: Pathogenesis, virulence factors, antibiotic resistance, interaction with host, technology advances and emerging therapeutics. Signal Transduct. Target. Ther. 2022, 7, 199. [Google Scholar] [CrossRef]
- Christaki, E.; Marcou, M.; Tofarides, A. Antimicrobial resistance in bacteria: Mechanisms, evolution, and persistence. J. Mol. Evol. 2020, 88, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.L.; Nosanchuk, J.D. Fungal diseases as neglected pathogens: A wake-up call to public health officials. PLoS Negl. Trop. Dis. 2020, 14, e0007964. [Google Scholar] [CrossRef] [PubMed]
- Lei, M.; Jayaraman, A.; Van Deventer, J.A.; Lee, K. Engineering selectively targeting antimicrobial peptides. Annu. Rev. Biomed. Eng. 2021, 23, 339–357. [Google Scholar] [CrossRef]
- Song, X.; Liu, P.; Liu, X.; Wang, Y.; Wei, H.; Zhang, J.; Yu, L.; Yan, X.; He, Z. Dealing with MDR bacteria and biofilm in the post-antibiotic era: Application of antimicrobial peptides-based nano-formulation. Mater. Sci. Eng. C 2021, 128, 112318. [Google Scholar] [CrossRef]
- Divyashree, M.; Mani, M.K.; Reddy, D.; Kumavath, R.; Ghosh, P.; Azevedo, V.; Barh, D. Clinical applications of antimicrobial peptides (AMPs): Where do we stand now? Protein Pept. Lett. 2020, 27, 120–134. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Hong, T.; Cui, P.; Wang, J.; Xia, J. Antimicrobial peptides towards clinical application: Delivery and formulation. Adv. Drug Deliv. Rev. 2021, 175, 113818. [Google Scholar] [CrossRef]
- Webber, M.J.; Pashuck, E.T. (Macro) molecular self-assembly for hydrogel drug delivery. Adv. Drug Deliv. Rev. 2021, 172, 275–295. [Google Scholar] [CrossRef] [PubMed]
- Barrón-González, M.; Montes-Aparicio, A.V.; Cuevas-Galindo, M.E.; Orozco-Suárez, S.; Barrientos, R.; Alatorre, A.; Querejeta, E.; Trujillo-Ferrara, J.G.; Farfán-García, E.D.; Soriano-Ursúa, M.A. Boron-containing compounds on neurons: Actions and potential applications for treating neurodegenerative diseases. J. Inorg. Biochem. 2023, 238, 112027. [Google Scholar] [CrossRef] [PubMed]
- Makvandi, P.; Wang, C.Y.; Zare, E.N.; Borzacchiello, A.; Niu, L.N.; Tay, F.R. Metal-based nanomaterials in biomedical applications: Antimicrobial activity and cytotoxicity aspects. Adv. Funct. Mater. 2020, 30, 1910021. [Google Scholar] [CrossRef]
- Kupnik, K.; Primožič, M.; Kokol, V.; Leitgeb, M. Nanocellulose in drug delivery and antimicrobially active materials. Polymers 2020, 12, 2825. [Google Scholar] [CrossRef] [PubMed]
- Ullah, I.; Ali, E. Bioavailability of Antibiotics and Their Toxicity. Antibiot. Antimicrob. Resist. Genes: Environ. Occur. Treat. Technol. 2020, 211–238, 1006. [Google Scholar]
- Elnagdy, S.; AlKhazindar, M. The potential of antimicrobial peptides as an antiviral therapy against COVID-19. ACS Pharmacol. Transl. Sci. 2020, 3, 780–782. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Chen, H.; Song, Y.; Qin, Z.; Xu, L.; He, N.; Tan, Y.; Dessie, W. Advancements, challenges and future perspectives on peptide-based drugs: Focus on antimicrobial peptides. Eur. J. Pharm. Sci. 2022, 181, 106363. [Google Scholar] [CrossRef]
- King, T.A.; Kandemir, J.M.; Walsh, S.J.; Spring, D.R. Photocatalytic methods for amino acid modification. Chem. Soc. Rev. 2021, 50, 39–57. [Google Scholar] [CrossRef]
- Chen, C.H.; Bepler, T.; Pepper, K.; Fu, D.; Lu, T.K. Synthetic molecular evolution of antimicrobial peptides. Curr. Opin. Biotechnol. 2022, 75, 102718. [Google Scholar] [CrossRef]
- Bakare, O.O.; Gokul, A.; Fadaka, A.O.; Wu, R.; Niekerk, L.-A.; Barker, A.M.; Keyster, M.; Klein, A. Plant antimicrobial peptides (PAMPs): Features, applications, production, expression, and challenges. Molecules 2022, 27, 3703. [Google Scholar] [CrossRef]
- Song, W.; Kong, X.; Hua, Y.; Chen, Y.; Zhang, C.; Chen, Y. Identification of antibacterial peptides generated from enzymatic hydrolysis of cottonseed proteins. LWT 2020, 125, 109199. [Google Scholar] [CrossRef]
- Amiri, S.; Mokarram, R.R.; Khiabani, M.S.; Bari, M.R.; Khaledabad, M.A. Characterization of antimicrobial peptides produced by Lactobacillus acidophilus LA-5 and Bifidobacterium lactis BB-12 and their inhibitory effect against foodborne pathogens. LWT 2022, 153, 112449. [Google Scholar] [CrossRef]
- Prasad, A.K.; Tiwari, C.; Ray, S.; Holden, S.; Armstrong, D.A.; Rosengren, K.J.; Rodger, A.; Panwar, A.S.; Martin, L.L. Secondary structure transitions for a family of amyloidogenic, antimicrobial uperin 3 peptides in contact with sodium dodecyl sulfate. ChemPlusChem 2022, 87, e202100408. [Google Scholar] [CrossRef] [PubMed]
- Lesur, A.; Dittmar, G. The clinical potential of prm-PASEF mass spectrometry. Expert Rev. Proteom. 2021, 18, 75–82. [Google Scholar] [CrossRef]
- Shi, J.; Su, R.-q.; Zhang, W.-t.; Chen, J. Purification and the secondary structure of a novel angiotensin I-converting enzyme (ACE) inhibitory peptide from the alcalase hydrolysate of seahorse protein. J. Food Sci. Technol. 2020, 57, 3927–3934. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.L.; Hampton, K.J.; Rosales, A.M.; Muturi, E.J. Multiple mosquito AMPs are needed to potentiate their antifungal effect against entomopathogenic fungi. Front. Microbiol. 2022, 13, 1062383. [Google Scholar] [CrossRef] [PubMed]
- Etebu, E.; Arikekpar, I. Antibiotics: Classification and mechanisms of action with emphasis on molecular perspectives. Int. J. Appl. Microbiol. Biotechnol. Res. 2016, 4, 90–101. [Google Scholar]
- Weledji, E.P.; Weledji, E.K.; Assob, J.C.; Nsagha, D.S. Pros, cons and future of antibiotics. New Horiz. Transl. Med. 2017, 4, 9–14. [Google Scholar] [CrossRef]
- Reygaert, W.C. An overview of the antimicrobial resistance mechanisms of bacteria. AIMS Microbiol. 2018, 4, 482. [Google Scholar] [CrossRef]
- Flasche, S.; Atkins, K.E. Balancing Benefits and Risks of Antibiotic Use; Oxford University Press US: Oxford, UK, 2018; Volume 218, pp. 1351–1353. [Google Scholar]
- Martens, E.; Demain, A.L. The antibiotic resistance crisis, with a focus on the United States. J. Antibiot. 2017, 70, 520–526. [Google Scholar] [CrossRef]
- Nelson, D.W.; Moore, J.E.; Rao, J.R. Antimicrobial resistance (AMR): Significance to food quality and safety. Food Qual. Saf. 2019, 3, 15–22. [Google Scholar] [CrossRef]
- Hayes, J.F. Fighting Back against Antimicrobial Resistance with Comprehensive Policy and Education: A Narrative Review. Antibiotics 2022, 11, 644. [Google Scholar] [CrossRef]
- Ribeiro da Cunha, B.; Fonseca, L.P.; Calado, C.R. Antibiotic discovery: Where have we come from, where do we go? Antibiotics 2019, 8, 45. [Google Scholar] [CrossRef]
- Benfield, A.H.; Henriques, S.T. Mode-of-action of antimicrobial peptides: Membrane disruption vs. intracellular mechanisms. Front. Med. Technol. 2020, 2, 610997. [Google Scholar] [CrossRef]
- Rima, M.; Rima, M.; Fajloun, Z.; Sabatier, J.-M.; Bechinger, B.; Naas, T. Antimicrobial peptides: A potent alternative to antibiotics. Antibiotics 2021, 10, 1095. [Google Scholar] [CrossRef]
- Zhu, Y.; Hao, W.; Wang, X.; Ouyang, J.; Deng, X.; Yu, H.; Wang, Y. Antimicrobial peptides, conventional antibiotics, and their synergistic utility for the treatment of drug-resistant infections. Med. Res. Rev. 2022, 42, 1377–1422. [Google Scholar] [CrossRef]
- Decker, A.P.; Mechesso, A.F.; Wang, G. Expanding the Landscape of Amino Acid-Rich Antimicrobial Peptides: Definition, Deployment in Nature, Implications for Peptide Design and Therapeutic Potential. Int. J. Mol. Sci. 2022, 23, 12874. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Koh, J.-J.; Liu, S.; Lakshminarayanan, R.; Verma, C.S.; Beuerman, R.W. Membrane active antimicrobial peptides: Translating mechanistic insights to design. Front. Neurosci. 2017, 11, 73. [Google Scholar] [CrossRef] [PubMed]
- Bechinger, B.; Gorr, S.-U. Antimicrobial peptides: Mechanisms of action and resistance. J. Dent. Res. 2017, 96, 254–260. [Google Scholar] [CrossRef]
- Huang, X.; Li, G. Antimicrobial Peptides and Cell-Penetrating Peptides: Non-Antibiotic Membrane-Targeting Strategies against Bacterial Infections. Infect. Drug Resist. 2023, 16, 1203–1219. [Google Scholar] [CrossRef]
- Moretta, A.; Scieuzo, C.; Petrone, A.M.; Salvia, R.; Manniello, M.D.; Franco, A.; Lucchetti, D.; Vassallo, A.; Vogel, H.; Sgambato, A. Antimicrobial peptides: A new hope in biomedical and pharmaceutical fields. Front. Cell. Infect. Microbiol. 2021, 11, 668632. [Google Scholar] [CrossRef]
- Malanovic, N.; Lohner, K. Antimicrobial peptides targeting gram-positive bacteria. Pharmaceuticals 2016, 9, 59. [Google Scholar] [CrossRef] [PubMed]
- Bakare, O.O.; Gokul, A.; Wu, R.; Niekerk, L.-A.; Klein, A.; Keyster, M. Biomedical relevance of novel anticancer peptides in the sensitive treatment of cancer. Biomolecules 2021, 11, 1120. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, S.; Bharath Prasad, A.; Mehta, C.H.; Nayak, U.Y. Antimicrobial peptide polymers: No escape to ESKAPE pathogens—A review. World J. Microbiol. Biotechnol. 2020, 36, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial peptides: Classification, design, application and research progress in multiple fields. Front. Microbiol. 2020, 11, 2559. [Google Scholar] [CrossRef]
- Chan, L.W.; Hern, K.E.; Ngambenjawong, C.; Lee, K.; Kwon, E.J.; Hung, D.T.; Bhatia, S.N. Selective permeabilization of gram-negative bacterial membranes using multivalent peptide constructs for antibiotic sensitization. ACS Infect. Dis. 2021, 7, 721–732. [Google Scholar] [CrossRef]
- Mangmee, S.; Reamtong, O.; Kalambaheti, T.; Roytrakul, S.; Sonthayanon, P. Antimicrobial Peptide Modifications against Clinically Isolated Antibiotic-Resistant Salmonella. Molecules 2021, 26, 4654. [Google Scholar] [CrossRef]
- Bakare, O.O.; Fadaka, A.O.; Klein, A.; Pretorius, A. Dietary effects of antimicrobial peptides in therapeutics. All Life 2020, 13, 78–91. [Google Scholar] [CrossRef]
- Yang, N.; Aminov, R.; Franco, O.L.; de la Fuente-Nunez, C.; Wang, J. Community series in antimicrobial peptides: Molecular design, structure function relationship and biosynthesis optimization. Front. Microbiol. 2023, 14, 1125426. [Google Scholar] [CrossRef]
- Yan, Y.; Li, Y.; Zhang, Z.; Wang, X.; Niu, Y.; Zhang, S.; Xu, W.; Ren, C. Advances of peptides for antibacterial applications. Colloids Surf. B Biointerfaces 2021, 202, 111682. [Google Scholar] [CrossRef]
- Timón, M.L.; Andrés, A.I.; Otte, J.; Petrón, M.J. Antioxidant peptides (<3 kDa) identified on hard cow milk cheese with rennet from different origin. Food Res. Int. 2019, 120, 643–649. [Google Scholar]
- Carratalá, J.V.; Serna, N.; Villaverde, A.; Vázquez, E.; Ferrer-Miralles, N. Nanostructured antimicrobial peptides: The last push towards clinics. Biotechnol. Adv. 2020, 44, 107603. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, P.; Glossop, H.; Meikle, T.G.; Aburto-Medina, A.; Conn, C.E.; Sarojini, V.; Valery, C. Molecular engineering of antimicrobial peptides: Microbial targets, peptide motifs and translation opportunities. Biophys. Rev. 2021, 13, 35–69. [Google Scholar] [CrossRef]
- Gupta, A.; Mumtaz, S.; Li, C.-H.; Hussain, I.; Rotello, V.M. Combatting antibiotic-resistant bacteria using nanomaterials. Chem. Soc. Rev. 2019, 48, 415–427. [Google Scholar] [CrossRef]
- Biernacki, K.; Daśko, M.; Ciupak, O.; Kubiński, K.; Rachon, J.; Demkowicz, S. Novel 1, 2, 4-oxadiazole derivatives in drug discovery. Pharmaceuticals 2020, 13, 111. [Google Scholar] [CrossRef]
- Pranantyo, D.; Zhang, K.; Si, Z.; Hou, Z.; Chan-Park, M.B. Smart multifunctional polymer systems as alternatives or supplements of antibiotics to overcome bacterial resistance. Biomacromolecules 2022, 23, 1873–1891. [Google Scholar] [CrossRef]
- Rapacka-Zdończyk, A.; Woźniak, A.; Michalska, K.; Pierański, M.; Ogonowska, P.; Grinholc, M.; Nakonieczna, J. Factors determining the susceptibility of bacteria to antibacterial photodynamic inactivation. Front. Med. 2021, 8, 642609. [Google Scholar] [CrossRef]
- Ciura, K.; Ptaszyńska, N.; Kapica, H.; Pastewska, M.; Łęgowska, A.; Rolka, K.; Kamysz, W.; Sawicki, W.; Greber, K.E. Can immobilized artificial membrane chromatography support the characterization of antimicrobial peptide origin derivatives? Antibiotics 2021, 10, 1237. [Google Scholar] [CrossRef]
- Zhang, H.; Zhen, J.; Zhang, R.; Wanyan, Y.; Liu, K.; Yuan, X.; Tao, L.; Chen, Y. Cathelicidin hCAP18/LL-37 promotes cell proliferation and suppresses antitumor activity of 1, 25 (OH) 2D3 in hepatocellular carcinoma. Cell Death Discov. 2022, 8, 27. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, M.A.; Antunes, J.C.; Seabra, C.L.; Tohidi, S.D.; Reis, S.; Amorim, M.T.P.; Felgueiras, H.P. Tiger 17 and pexiganan as antimicrobial and hemostatic boosters of cellulose acetate-containing poly (vinyl alcohol) electrospun mats for potential wound care purposes. Int. J. Biol. Macromol. 2022, 209, 1526–1541. [Google Scholar] [CrossRef] [PubMed]
- Shen, P.; Ding, K.; Wang, L.; Tian, J.; Huang, X.; Zhang, M.; Dang, X. In vitro and in vivo antimicrobial activity of antimicrobial peptide Jelleine-I against foodborne pathogen Listeria monocytogenes. Int. J. Food Microbiol. 2023, 387, 110050. [Google Scholar] [CrossRef]
- Chee, P.Y.; Mang, M.; Lau, E.S.; Tan, L.T.-H.; He, Y.-W.; Lee, W.-L.; Pusparajah, P.; Chan, K.-G.; Lee, L.-H.; Goh, B.-H. Epinecidin-1, an antimicrobial peptide derived from grouper (Epinephelus coioides): Pharmacological activities and applications. Front. Microbiol. 2019, 10, 2631. [Google Scholar] [CrossRef] [PubMed]
- Westrich, J.A.; Vermeer, D.W.; Colbert, P.L.; Spanos, W.C.; Pyeon, D. The multifarious roles of the chemokine CXCL14 in cancer progression and immune responses. Mol. Carcinog. 2020, 59, 794–806. [Google Scholar] [CrossRef] [PubMed]
- Van Bramer, S.; Glatfelter, A.; Bai, S.; Dybowski, C.; Neue, G.; Perry, D. Solid-state 207Pb NMR studies of lead-group 16 and mixed transition-metal/lead-group 16 element-containing materials. Magn. Reson. Chem. 2006, 44, 357–365. [Google Scholar] [CrossRef]
- Huang, C.; Yang, X.; Huang, J.; Liu, X.; Yang, X.; Jin, H.; Huang, Q.; Li, L.; Zhou, R. Porcine beta-defensin 2 provides protection against bacterial infection by a direct bactericidal activity and alleviates inflammation via interference with the TLR4/NF-κB pathway. Front. Immunol. 2019, 10, 1673. [Google Scholar] [CrossRef]
- Kim, J.; Jacob, B.; Jang, M.; Kwak, C.; Lee, Y.; Son, K.; Lee, S.; Jung, I.D.; Jeong, M.S.; Kwon, S.-H. Development of a novel short 12-meric papiliocin-derived peptide that is effective against Gram-negative sepsis. Sci. Rep. 2019, 9, 3817. [Google Scholar] [CrossRef]
- Zhang, J.; Sun, Y.; Kang, Y.; Shang, D. Antimicrobial peptide temporin-1CEa isolated from frog skin secretions inhibits the proinflammatory response in lipopolysaccharide-stimulated RAW264. 7 murine macrophages through the MyD88-dependent signaling pathway. Mol. Immunol. 2021, 132, 227–235. [Google Scholar] [CrossRef]
- Choi, H.; Hwang, J.S.; Lee, D.G. Coprisin exerts antibacterial effects by inducing apoptosis-like death in Escherichia coli. IUBMB Life 2016, 68, 72–78. [Google Scholar] [CrossRef]
- Nugteren, S.; den Uil, S.H.; Delis-van Diemen, P.M.; Simons-Oosterhuis, Y.; Lindenbergh-Kortleve, D.J.; van Haaften, D.H.; Stockmann, H.B.; Sanders, J.; Meijer, G.A.; Fijneman, R.J. High expression of secretory leukocyte protease inhibitor (SLPI) in stage III micro-satellite stable colorectal cancer is associated with reduced disease recurrence. Sci. Rep. 2022, 12, 12174. [Google Scholar] [CrossRef] [PubMed]
- Dissanayake, S.; He, J.; Yang, S.H.; Brimble, M.A.; Harris, P.W.; Cameron, A.J. Flow-Based Fmoc-SPPS Preparation and SAR Study of Cathelicidin-PY Reveals Selective Antimicrobial Activity. Molecules 2023, 28, 1993. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jiang, B.; Wang, K.; Dai, J.; Dong, C.; Wang, Y.; Zhang, P.; Li, M.; Xu, W.; Wei, L. A cathelicidin antimicrobial peptide from Hydrophis cyanocinctus inhibits Zika virus infection by downregulating expression of a viral entry factor. J. Biol. Chem. 2022, 298, 102471. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, R.; Wang, F.; Cai, X.; Wang, Y. Antimicrobial peptide GL13K immobilized onto SLA-treated titanium by silanization: Antibacterial effect against methicillin-resistant Staphylococcus aureus (MRSA). RSC Adv. 2022, 12, 6918–6929. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Jiménez, J.; Berraquero-García, C.; Pérez-Gálvez, R.; García-Moreno, P.J.; Espejo-Carpio, F.J.; Guadix, A.; Guadix, E.M. Peptides and protein hydrolysates exhibiting anti-inflammatory activity: Sources, structural features and modulation mechanisms. Food Funct. 2022, 13, 12510–12540. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Fu, L.; Wen, W.; Dong, N. The dual antimicrobial and immunomodulatory roles of host defense peptides and their applications in animal production. J. Anim. Sci. Biotechnol. 2022, 13, 141. [Google Scholar] [CrossRef] [PubMed]
- van Dijk, A.; Anten, J.; Bakker, A.; Evers, N.; Hoekstra, A.T.; Chang, J.-C.; Scheenstra, M.R.; Veldhuizen, E.J.; Netea, M.G.; Berkers, C.R. Innate immune training of human macrophages by cathelicidin analogs. Front. Immunol. 2022, 13, 777530. [Google Scholar] [CrossRef]
- Hwang, C.-H.; Lee, N.-K.; Paik, H.-D. The anti-cancer potential of heat-killed Lactobacillus brevis KU15176 upon AGS cell lines through intrinsic apoptosis pathway. Int. J. Mol. Sci. 2022, 23, 4073. [Google Scholar] [CrossRef]
- Mu, L.; Zhou, L.; Yang, J.; Zhuang, L.; Tang, J.; Liu, T.; Wu, J.; Yang, H. The first identified cathelicidin from tree frogs possesses anti-inflammatory and partial LPS neutralization activities. Amino Acids 2017, 49, 1571–1585. [Google Scholar] [CrossRef]
- Brady, D.; Grapputo, A.; Romoli, O.; Sandrelli, F. Insect cecropins, antimicrobial peptides with potential therapeutic applications. Int. J. Mol. Sci. 2019, 20, 5862. [Google Scholar] [CrossRef]
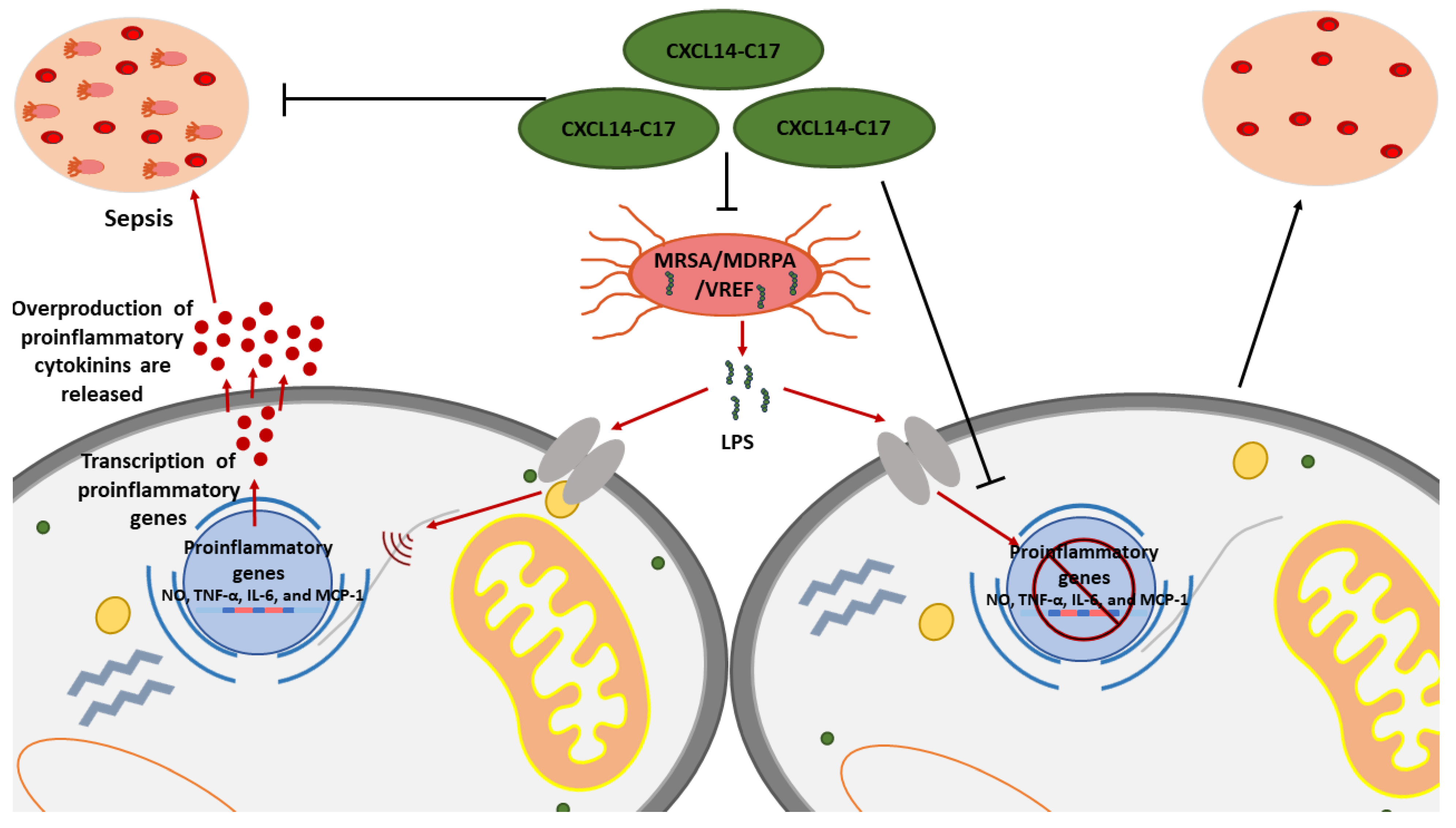
- Rajasekaran, G.; Kumar, S.D.; Nam, J.; Jeon, D.; Kim, Y.; Lee, C.W.; Park, I.-S.; Shin, S.Y. Antimicrobial and anti-inflammatory activities of chemokine CXCL14-derived antimicrobial peptide and its analogs. Biochim. Biophys. Acta (BBA) Biomembr. 2019, 1861, 256–267. [Google Scholar] [CrossRef]
- Kong, D.; Hua, X.; Zhou, R.; Cui, J.; Wang, T.; Kong, F.; You, H.; Liu, X.; Adu-Amankwaah, J.; Guo, G. Antimicrobial and Anti-Inflammatory Activities of MAF-1-Derived Antimicrobial Peptide Mt6 and Its D-Enantiomer D-Mt6 against Acinetobacter baumannii by Targeting Cell Membranes and Lipopolysaccharide Interaction. Microbiol. Spectr. 2022, 10, e01312–e01322. [Google Scholar] [CrossRef]
- Klubthawee, N.; Adisakwattana, P.; Hanpithakpong, W.; Somsri, S.; Aunpad, R. A novel, rationally designed, hybrid antimicrobial peptide, inspired by cathelicidin and aurein, exhibits membrane-active mechanisms against Pseudomonas aeruginosa. Sci. Rep. 2020, 10, 9117. [Google Scholar] [CrossRef] [PubMed]
- de Azevedo dos Santos, L.; Taveira, G.B.; da Silva, M.S.; da Silva Gebara, R.; da Silva Pereira, L.; Perales, J.; Teixeira-Ferreira, A.; de Oliveira Mello, É.; de Oliveira Carvalho, A.; Rodrigues, R. Antimicrobial peptides from Capsicum chinense fruits: Agronomic alternatives against phytopathogenic fungi. Biosci. Rep. 2020, 40, BSR20200950. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.A.; Abd El-Aziz, S.; Elbadry, H.M.; Samy, A.; Tamer, T.M. Prevalence, antimicrobial resistance profile, and characterization of multi-drug resistant bacteria from various infected wounds in North Egypt. Saudi J. Biol. Sci. 2022, 29, 2978–2988. [Google Scholar] [CrossRef]
- Bhattacharjya, S.; Straus, S.K. Design, engineering and discovery of novel α-helical and β-boomerang antimicrobial peptides against drug resistant bacteria. Int. J. Mol. Sci. 2020, 21, 5773. [Google Scholar] [CrossRef]
- Mahlapuu, M.; Björn, C.; Ekblom, J. Antimicrobial peptides as therapeutic agents: Opportunities and challenges. Crit. Rev. Biotechnol. 2020, 40, 978–992. [Google Scholar] [CrossRef] [PubMed]
- Ghorani-Azam, A. Antimicrobial Peptides: Importance in Biomedicine, and Future Directions. EC Pharmacol. Toxicol. 2022, 10, 80–85. [Google Scholar]
- Lopes, B.S.; Hanafiah, A.; Nachimuthu, R.; Muthupandian, S.; Md Nesran, Z.N.; Patil, S. The role of antimicrobial peptides as antimicrobial and antibiofilm agents in tackling the silent pandemic of antimicrobial resistance. Molecules 2022, 27, 2995. [Google Scholar] [CrossRef]
- Boparai, J.K.; Sharma, P.K. Mini review on antimicrobial peptides, sources, mechanism and recent applications. Protein Pept. Lett. 2020, 27, 4–16. [Google Scholar] [CrossRef]
- Guryanova, S.V.; Ovchinnikova, T.V. Immunomodulatory and allergenic properties of antimicrobial peptides. Int. J. Mol. Sci. 2022, 23, 2499. [Google Scholar] [CrossRef]
- Nage, N.B.; Wadekar, V.N.; Baig, M. Antimicrobial peptides: Current perspectives and future promises. Microbes Infect. Dis. 2023, 187321. [Google Scholar] [CrossRef]
- Wang, G.; Vaisman, I.I.; van Hoek, M.L. Machine learning prediction of antimicrobial peptides. In Computational Peptide Science: Methods and Protocols; Springer: Berlin/Heidelberg, Germany, 2022; pp. 1–37. [Google Scholar]
- Kurpe, S.R.; Grishin, S.Y.; Surin, A.K.; Panfilov, A.V.; Slizen, M.V.; Chowdhury, S.D.; Galzitskaya, O.V. Antimicrobial and amyloidogenic activity of peptides. Can antimicrobial peptides be used against SARS-CoV-2? Int. J. Mol. Sci. 2020, 21, 9552. [Google Scholar] [CrossRef] [PubMed]
- Pirtskhalava, M.; Amstrong, A.A.; Grigolava, M.; Chubinidze, M.; Alimbarashvili, E.; Vishnepolsky, B.; Gabrielian, A.; Rosenthal, A.; Hurt, D.E.; Tartakovsky, M. DBAASP v3: Database of antimicrobial/cytotoxic activity and structure of peptides as a resource for development of new therapeutics. Nucleic Acids Res. 2021, 49, D288–D297. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, J.; Ikram, S.; Ahmad, F.; Rehman, I.U.; Mushtaq, M. SARS-CoV-2 RNA dependent RNA polymerase (RdRp)—A drug repurposing study. Heliyon 2020, 6, e04502. [Google Scholar] [CrossRef]
- Shinde, M.U.; Patwekar, M.; Patwekar, F.; Bajaber, M.A.; Medikeri, A.; Mohammad, F.S.; Mukim, M.; Soni, S.; Mallick, J.; Jawaid, T. Nanomaterials: A potential hope for life sciences from bench to bedside. J. Nanomater. 2022, 2022, 5968131. [Google Scholar] [CrossRef]
- Del Rio, D.D.F.; Sovacool, B.K.; Foley, A.M.; Griffiths, S.; Bazilian, M.; Kim, J.; Rooney, D. Decarbonizing the glass industry: A critical and systematic review of developments, sociotechnical systems and policy options. Renew. Sustain. Energy Rev. 2022, 155, 111885. [Google Scholar] [CrossRef]
- Kebede, L.; Masoomi Dezfooli, S.; Seyfoddin, A. Medicinal cannabis pharmacokinetics and potential methods of delivery. Pharm. Dev. Technol. 2022, 27, 202–214. [Google Scholar] [CrossRef]
- Patel, M.; Joshi, G.; Sawant, K.K. Nanotechnology in oral drug delivery: Salient aspects, state of art, and applications. Funct. Bionanomater. Biomol. Nanopart. 2020, 165–184, 466. [Google Scholar]
- Pant, A.; Mackraj, I.; Govender, T. Advances in sepsis diagnosis and management: A paradigm shift towards nanotechnology. J. Biomed. Sci. 2021, 28, 6. [Google Scholar] [CrossRef]
- Roque-Borda, C.A.; Da Silva, P.B.; Rodrigues, M.C.; Di Filippo, L.D.; Duarte, J.L.; Chorilli, M.; Vicente, E.F.; Garrido, S.S.; Pavan, F.R. Pharmaceutical Nanotechnology: Antimicrobial Peptides as potential new drugs against WHO list of critical, high, and medium priority bacteria. Eur. J. Med. Chem. 2022, 241, 114640. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, Y.; Shi, Y.; Song, H.; Yu, C. Antibiotic-free antibacterial strategies enabled by nanomaterials: Progress and perspectives. Adv. Mater. 2020, 32, 1904106. [Google Scholar] [CrossRef] [PubMed]
- Thapa, R.K.; Diep, D.B.; Tønnesen, H.H. Nanomedicine-based antimicrobial peptide delivery for bacterial infections: Recent advances and future prospects. J. Pharm. Investig. 2021, 51, 377–398. [Google Scholar] [CrossRef]
- Gupta, R.; Badhe, Y.; Mitragotri, S.; Rai, B. Permeation of nanoparticles across the intestinal lipid membrane: Dependence on shape and surface chemistry studied through molecular simulations. Nanoscale 2020, 12, 6318–6333. [Google Scholar] [CrossRef] [PubMed]
- Pinilla, C.M.B.; Lopes, N.A.; Brandelli, A. Lipid-based nanostructures for the delivery of natural antimicrobials. Molecules 2021, 26, 3587. [Google Scholar] [CrossRef]
- Martin-Serrano, Á.; Gómez, R.; Ortega, P.; de la Mata, F.J. Nanosystems as vehicles for the delivery of antimicrobial peptides (AMPs). Pharmaceutics 2019, 11, 448. [Google Scholar] [CrossRef]
- Kirtane, A.R.; Verma, M.; Karandikar, P.; Furin, J.; Langer, R.; Traverso, G. Nanotechnology approaches for global infectious diseases. Nat. Nanotechnol. 2021, 16, 369–384. [Google Scholar] [CrossRef] [PubMed]
- Kazemzadeh-Narbat, M.; Cheng, H.; Chabok, R.; Alvarez, M.M.; De La Fuente-Nunez, C.; Phillips, K.S.; Khademhosseini, A. Strategies for antimicrobial peptide coatings on medical devices: A review and regulatory science perspective. Crit. Rev. Biotechnol. 2021, 41, 94–120. [Google Scholar] [CrossRef]
- Zhang, L.; Xue, Y.; Gopalakrishnan, S.; Li, K.; Han, Y.; Rotello, V.M. Antimicrobial peptide-loaded pectolite nanorods for enhancing wound-healing and biocidal activity of titanium. ACS Appl. Mater. Interfaces 2021, 13, 28764–28773. [Google Scholar] [CrossRef]
- Teixeira, M.A.; Antunes, J.C.; Seabra, C.L.; Fertuzinhos, A.; Tohidi, S.D.; Reis, S.; Amorim, M.T.P.; Ferreira, D.P.; Felgueiras, H.P. Antibacterial and hemostatic capacities of cellulose nanocrystalline-reinforced poly (vinyl alcohol) electrospun mats doped with Tiger 17 and pexiganan peptides for prospective wound healing applications. Biomater. Adv. 2022, 137, 212830. [Google Scholar] [CrossRef]
- Wang, C.-C.; Wei, S.-C.; Luo, S.-C. Recent advances and biomedical applications of peptide-integrated conducting polymers. ACS Appl. Bio Mater. 2022, 5, 1916–1933. [Google Scholar] [CrossRef]
- Bhattacharjee, B.; Ghosh, S.; Patra, D.; Haldar, J. Advancements in release-active antimicrobial biomaterials: A journey from release to relief. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2022, 14, e1745. [Google Scholar] [CrossRef]
- Vasanthan, J.; Gurusamy, N.; Rajasingh, S.; Sigamani, V.; Kirankumar, S.; Thomas, E.L.; Rajasingh, J. Role of human mesenchymal stem cells in regenerative therapy. Cells 2020, 10, 54. [Google Scholar] [CrossRef]
- Xuan, X.; Tian, C.; Zhao, M.; Sun, Y.; Huang, C. Mesenchymal stem cells in cancer progression and anticancer therapeutic resistance. Cancer Cell Int. 2021, 21, 595. [Google Scholar] [CrossRef] [PubMed]
- Moeinabadi-Bidgoli, K.; Rezaee, M.; Rismanchi, H.; Mohammadi, M.M.; Babajani, A. Mesenchymal stem cell-derived antimicrobial peptides as potential anti-neoplastic agents: New insight into anticancer mechanisms of stem cells and exosomes. Front. Cell Dev. Biol. 2022, 10, 900418. [Google Scholar] [CrossRef] [PubMed]
- Harman, R.M.; Yang, S.; He, M.K.; Van de Walle, G.R. Antimicrobial peptides secreted by equine mesenchymal stromal cells inhibit the growth of bacteria commonly found in skin wounds. Stem Cell Res. Ther. 2017, 8, 157. [Google Scholar] [CrossRef]
- Yagi, H.; Chen, A.F.; Hirsch, D.; Rothenberg, A.C.; Tan, J.; Alexander, P.G.; Tuan, R.S. Antimicrobial activity of mesenchymal stem cells against Staphylococcus aureus. Stem Cell Res. Ther. 2020, 11, 293. [Google Scholar] [CrossRef]
- Alfei, S.; Schito, A.M. From nanobiotechnology, positively charged biomimetic dendrimers as novel antibacterial agents: A review. Nanomaterials 2020, 10, 2022. [Google Scholar] [CrossRef]
- Zhang, Q.-Y.; Yan, Z.-B.; Meng, Y.-M.; Hong, X.-Y.; Shao, G.; Ma, J.-J.; Cheng, X.-R.; Liu, J.; Kang, J.; Fu, C.-Y. Antimicrobial peptides: Mechanism of action, activity and clinical potential. Mil. Med. Res. 2021, 8, 48. [Google Scholar] [CrossRef]
- Seyfi, R.; Kahaki, F.A.; Ebrahimi, T.; Montazersaheb, S.; Eyvazi, S.; Babaeipour, V.; Tarhriz, V. Antimicrobial peptides (AMPs): Roles, functions and mechanism of action. Int. J. Pept. Res. Ther. 2020, 26, 1451–1463. [Google Scholar] [CrossRef]
- Gorgun, C.; Ceresa, D.; Lesage, R.; Villa, F.; Reverberi, D.; Balbi, C.; Santamaria, S.; Cortese, K.; Malatesta, P.; Geris, L. Dissecting the effects of preconditioning with inflammatory cytokines and hypoxia on the angiogenic potential of mesenchymal stromal cell (MSC)-derived soluble proteins and extracellular vesicles (EVs). Biomaterials 2021, 269, 120633. [Google Scholar] [CrossRef]
- Zouggar, A. G9a/EHMT2 Methyltransferase Activity Controls Stem-Like Identity and Tumor-Initiating Function in Human Colorectal Cancer; Université d’Ottawa/University of Ottawa: Ottawa, ON, Canada, 2021. [Google Scholar]
- Hamazaki, T.; El Rouby, N.; Fredette, N.C.; Santostefano, K.E.; Terada, N. Concise review: Induced pluripotent stem cell research in the era of precision medicine. Stem Cells 2017, 35, 545–550. [Google Scholar] [CrossRef]
- Najmi, Z.; Kumar, A.; Scalia, A.C.; Cochis, A.; Obradovic, B.; Grassi, F.A.; Leigheb, M.; Lamghari, M.; Loinaz, I.; Gracia, R. Evaluation of nisin and LL-37 antimicrobial peptides as tool to preserve articular cartilage healing in a septic environment. Front. Bioeng. Biotechnol. 2020, 8, 561. [Google Scholar] [CrossRef]
- Specht, H.; Emmott, E.; Petelski, A.A.; Huffman, R.G.; Perlman, D.H.; Serra, M.; Kharchenko, P.; Koller, A.; Slavov, N. Single-cell proteomic and transcriptomic analysis of macrophage heterogeneity using SCoPE2. Genome Biol. 2021, 22, 50. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Wang, Y.; Liu, L.; He, J. Biofilms: The microbial “protective clothing” in extreme environments. Int. J. Mol. Sci. 2019, 20, 3423. [Google Scholar] [CrossRef] [PubMed]
- Mahto, K.U.; Priyadarshanee, M.; Samantaray, D.P.; Das, S. Bacterial biofilm and extracellular polymeric substances in the treatment of environmental pollutants: Beyond the protective role in survivability. J. Clean. Prod. 2022, 379, 134759. [Google Scholar] [CrossRef]
- Yasir, M.; Willcox, M.D.P.; Dutta, D. Action of antimicrobial peptides against bacterial biofilms. Materials 2018, 11, 2468. [Google Scholar] [CrossRef] [PubMed]
- Martín-Rodríguez, A.J. Respiration-induced biofilm formation as a driver for bacterial niche colonization. Trends Microbiol. 2022, 31, 1016. [Google Scholar] [CrossRef] [PubMed]
- Efremov, Y.M.; Zurina, I.M.; Presniakova, V.S.; Kosheleva, N.V.; Butnaru, D.V.; Svistunov, A.A.; Rochev, Y.A.; Timashev, P.S. Mechanical properties of cell sheets and spheroids: The link between single cells and complex tissues. Biophys. Rev. 2021, 13, 541–561. [Google Scholar] [CrossRef]
- Pandey, R.P.; Mukherjee, R.; Chang, C.-M. Emerging concern with imminent therapeutic strategies for treating resistance in biofilm. Antibiotics 2022, 11, 476. [Google Scholar] [CrossRef]
- Klümper, U.; Recker, M.; Zhang, L.; Yin, X.; Zhang, T.; Buckling, A.; Gaze, W.H. Selection for antimicrobial resistance is reduced when embedded in a natural microbial community. ISME J. 2019, 13, 2927–2937. [Google Scholar] [CrossRef]
- Kour, D.; Kaur, T.; Fahliyani, S.A.; Rastegari, A.A.; Yadav, N.; Yadav, A.N. Microbial biofilms in the human: Diversity and potential significances in health and disease. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2020; pp. 89–124. [Google Scholar]
- Pang, L.; Lin, H.; Yang, F.; Deng, D. Mechanisms of biofilm development and antibiofilm strategies. Front. Microbiol. 2023, 14, 1098. [Google Scholar] [CrossRef]
- Devanga Ragupathi, N.K.; Veeraraghavan, B.; Karunakaran, E.; Monk, P.N. Biofilm-mediated nosocomial infections and its association with antimicrobial resistance: Detection, prevention, and management. Front. Med. 2022, 9, 987011. [Google Scholar] [CrossRef]
- Ghosh, S.; Mukherjee, S.; Patra, D.; Haldar, J. Polymeric biomaterials for prevention and therapeutic intervention of microbial infections. Biomacromolecules 2022, 23, 592–608. [Google Scholar] [CrossRef] [PubMed]
- Parrino, B.; Schillaci, D.; Carnevale, I.; Giovannetti, E.; Diana, P.; Cirrincione, G.; Cascioferro, S. Synthetic small molecules as anti-biofilm agents in the struggle against antibiotic resistance. Eur. J. Med. Chem. 2019, 161, 154–178. [Google Scholar] [CrossRef]
- Batoni, G.; Maisetta, G.; Esin, S. Therapeutic potential of antimicrobial peptides in polymicrobial biofilm-associated infections. Int. J. Mol. Sci. 2021, 22, 482. [Google Scholar] [CrossRef]
- Ghosh, C.; Sarkar, P.; Issa, R.; Haldar, J. Alternatives to conventional antibiotics in the era of antimicrobial resistance. Trends Microbiol. 2019, 27, 323–338. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Caputo, G.A.; Kuroda, K. Amphiphilic polymer therapeutics: An alternative platform in the fight against antibiotic resistant bacteria. Biomater. Sci. 2021, 9, 2758–2767. [Google Scholar] [CrossRef] [PubMed]
- Haidari, H.; Melguizo-Rodríguez, L.; Cowin, A.J.; Kopecki, Z. Therapeutic potential of antimicrobial peptides for treatment of wound infection. Am. J. Physiol. Cell Physiol. 2023, 324, C29–C38. [Google Scholar] [CrossRef]
- Sahoo, A.; Swain, S.S.; Behera, A.; Sahoo, G.; Mahapatra, P.K.; Panda, S.K. Antimicrobial peptides derived from insects offer a novel therapeutic option to combat biofilm: A review. Front. Microbiol. 2021, 12, 661195. [Google Scholar] [CrossRef]
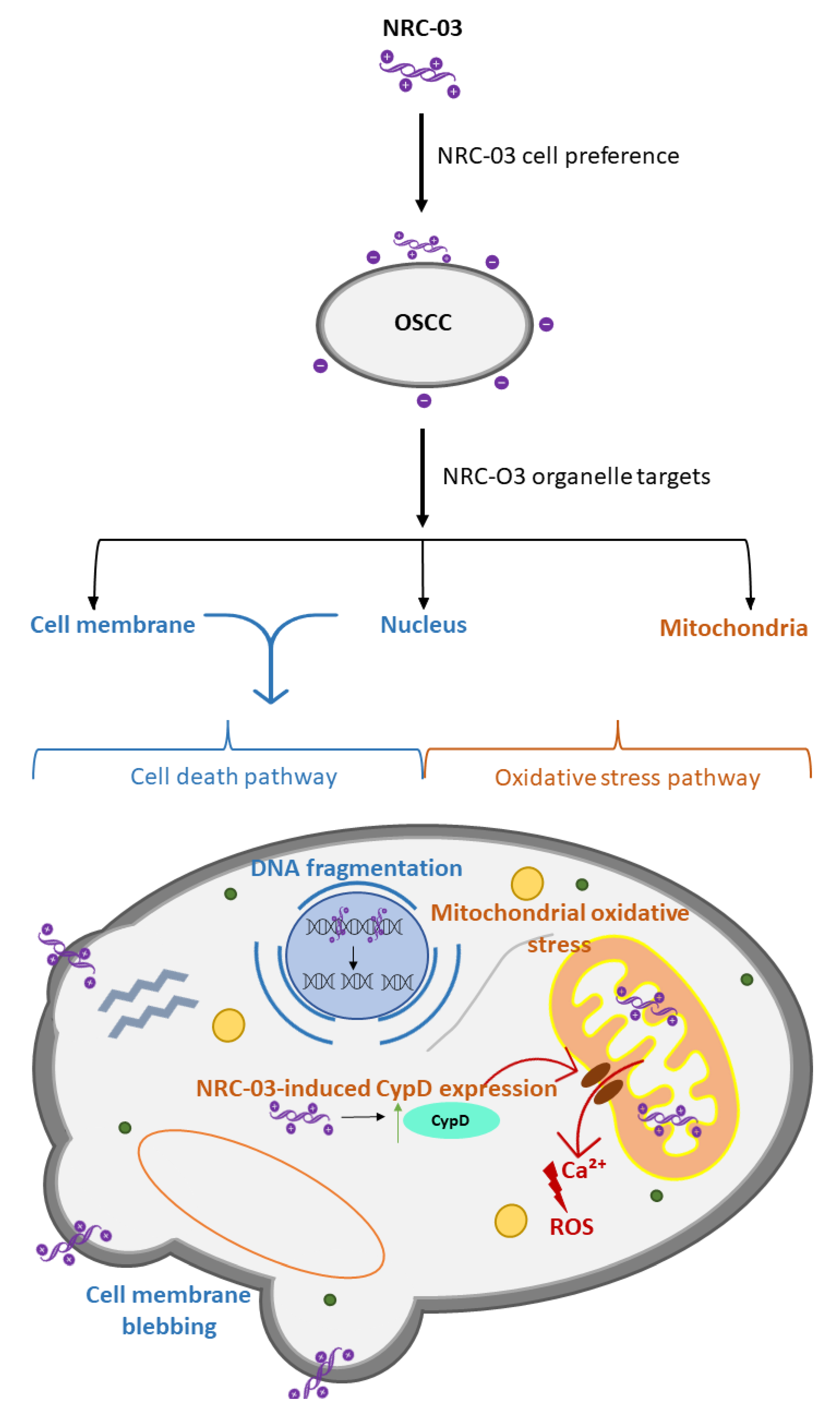
- Hou, D.; Hu, F.; Mao, Y.; Yan, L.; Zhang, Y.; Zheng, Z.; Wu, A.; Forouzanfar, T.; Pathak, J.L.; Wu, G. Cationic antimicrobial peptide NRC-03 induces oral squamous cell carcinoma cell apoptosis via CypD-mPTP axis-mediated mitochondrial oxidative stress. Redox Biol. 2022, 54, 102355. [Google Scholar] [CrossRef]
- Jia, F.; Zhang, Y.; Wang, J.; Peng, J.; Zhao, P.; Zhang, L.; Yao, H.; Ni, J.; Wang, K. The effect of halogenation on the antimicrobial activity, antibiofilm activity, cytotoxicity and proteolytic stability of the antimicrobial peptide Jelleine-I. Peptides 2019, 112, 56–66. [Google Scholar] [CrossRef]
- Gong, H.; Zhang, J.; Hu, X.; Li, Z.; Fa, K.; Liu, H.; Waigh, T.A.; McBain, A.; Lu, J.R. Hydrophobic control of the bioactivity and cytotoxicity of de novo-designed antimicrobial peptides. ACS Appl. Mater. Interfaces 2019, 11, 34609–34620. [Google Scholar] [CrossRef] [PubMed]
- Lai, Z.; Yuan, X.; Chen, H.; Zhu, Y.; Dong, N.; Shan, A. Strategies employed in the design of antimicrobial peptides with enhanced proteolytic stability. Biotechnol. Adv. 2022, 59, 107962. [Google Scholar] [CrossRef]
- Di, Y.; Lin, Q.; Chen, C.; Montelaro, R.; Doi, Y.; Deslouches, B. Enhanced therapeutic index of an antimicrobial peptide in mice by increasing safety and activity against multidrug-resistant bacteria. Sci. Adv. 2020, 6, eaay6817. [Google Scholar] [CrossRef] [PubMed]
- Beaudoin, T.; Stone, T.A.; Glibowicka, M.; Adams, C.; Yau, Y.; Ahmadi, S.; Bear, C.E.; Grasemann, H.; Waters, V.; Deber, C.M. Activity of a novel antimicrobial peptide against Pseudomonas aeruginosa biofilms. Sci. Rep. 2018, 8, 14728. [Google Scholar] [CrossRef]
- Deslouches, B.; Di, Y.P. Antimicrobial peptides with selective antitumor mechanisms: Prospect for anticancer applications. Oncotarget 2017, 8, 46635. [Google Scholar] [CrossRef] [PubMed]
- Sandín García, D. Design and Optimization of New Antimicrobial Peptides against Gram-Negative Bacteria. 2023. Available online: https://www.tdx.cat/handle/10803/688589#page=1 (accessed on 1 June 2023).



{kind=link}
{kind=link}
| AMP Database Codes | Specific Names | Sequences | Specific Functions | References |
|---|---|---|---|---|
| AP00351 | Citropin 1.1 | GLFDVIKKVASVIGGL | Peptidoglycan breakdown for its access to the cytoplasmic membrane | [59] |
| AP00557 | Cathelicidin 1 | RVKRVWPLVIRTVIAGYNLYRAIKKK | Bind and disrupt negatively charged membranes resulting in cell death | [60] |
| AP00952 | Pexiganan acetate | GIGKFLKKAKKFGKAFVKILKK | Disturbance/disruption of the cell membrane permeability | [61] |
| AP01210 | Jelleine-I | PFKLSLHL | Improvement of intestinal mucosa tight junction | [62] |
| AP01328 | Epinecidin-1 | GFIFHIIKGLFHAGKMIHGLV | Immunomodulatory and wound-healing functions | [63] |
| AP01372 | CXCL14 | SKCKCSRKGPKIRYSDVKKLEMKPKYPHCEEKMVIITTKSVSRYRGQEHCLHPKLQSTKRFIKWYNAWNEKRRVYEE | Inhibition of colorectal cancer migration, invasion, and epithelial-to-mesenchymal transition (EMT) by suppressing NF-kappaB signaling | [64] |
| AP01550 | Dybowskin-2CDYa | SAVGRHGRRFGLRKHRKH | Promotion of cell proliferation | [65] |
| AP01590 | porcine beta-defensin 2 | DHYICAKKGGTCNFSPCPLFNRIEGTCYSGKAKCCIR | Alleviation of inflammation via interference with the TLR4/NF-κB Pathway | [66] |
| AP01751 | Papiliocin | RWKIFKKIEKVGRNVRDGIIKAGPAVAVVGQAATVVK | Possession of broad-spectrum antibacterial activities | [67] |
| AP01794 | Temporin-1CEa | FVDLKKIANIINSIF | Possession of broad-spectrum antibacterial activities | [68] |
| AP01976 | Coprisin | VTCDVLSFEAKGIAVNHSACALHCIALRKKGGSCQNGVCVCRN | Prevention of Clostridium difficile-mediated acute inflammation and mucosal damage through selective antimicrobial activity | [69] |
| AP02095 | secretory leukocyte protease inhibitor | SGKSFKAGVCPPKKSAQCLRYKKPECQSDWQCPGKKRCCPDTCGIKCLDPVDTPNPTRRKPGKCPVTYGQCLMLNPPNFCEMDGQCKRDLKCCMGMCGKSCVSPVKA | Protection of tissues by inhibiting the proteases, such as cathepsin G, elastase, and trypsin from neutrophils; chymotrypsin and trypsin from pancreatic acinar cells; and chymase and tryptase from mast cells | [70] |
| AP02202 | Cathelicidin-PY | RKCNFLCKLKEKLRTVITSHIDKVLRPQG | Inhibition of the activation of Toll-like receptor 4 (TLR4) inflammatory response pathways induced by lipopolysaccharide (LPS) | [71] |
| AP02569 | Hc-CATH | KFFKRLLKSVRRAVKKFRKKPRLIGLSTLL | Hc-cath exerts an antimicrobial effect against P. aeruginosa and S. aureus and promotes Galleria survival | [72] |
| AP02570 | GL13K | GKIIKLKASLKLL | GL13K exhibits bactericidal activity against planktonic bacteria and low toxicity against mammalian cells | [73] |
| AP02621 | SibaCec | GKLTKDKLKRGAKKALNVASKVAPIVAAGASIAR | SibaCec can function as an immune regulator, inhibiting the host defense system against infection | [74] |
| AP02626 | Cl-CATH2 | LIQRGRFGRFLGRIRRFRPRINFDIRARGSIRLG | Cl-CATH2 exerts broad-spectrum but moderate antimicrobial activities | [75] |
| AP02629 | dCATH | KRFWQLVPLAIKIYRAWKRR | Activation of autophagy, which enhances antimicrobial effects against diverse pathogens | [76] |
| AP02703 | Periplanetasin-5 | MKTFLRLYRSLINKVLH | Control of inflammation by inhibiting phosphorylation of MAPKs, and reduction of degradation of IκB. | [77] |
| AP02901 | Cathelicidin-PP | ASENGKCNLLCLVKKKLRAVGNVIKTVVGKIA | Reduction of bacterial load in clinically relevant infection models | [78] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakare, O.O.; Gokul, A.; Niekerk, L.-A.; Aina, O.; Abiona, A.; Barker, A.M.; Basson, G.; Nkomo, M.; Otomo, L.; Keyster, M.; et al. Recent Progress in the Characterization, Synthesis, Delivery Procedures, Treatment Strategies, and Precision of Antimicrobial Peptides. Int. J. Mol. Sci. 2023, 24, 11864. https://doi.org/10.3390/ijms241411864
Bakare OO, Gokul A, Niekerk L-A, Aina O, Abiona A, Barker AM, Basson G, Nkomo M, Otomo L, Keyster M, et al. Recent Progress in the Characterization, Synthesis, Delivery Procedures, Treatment Strategies, and Precision of Antimicrobial Peptides. International Journal of Molecular Sciences. 2023; 24(14):11864. https://doi.org/10.3390/ijms241411864
Chicago/Turabian StyleBakare, Olalekan Olanrewaju, Arun Gokul, Lee-Ann Niekerk, Omolola Aina, Ademola Abiona, Adele Mariska Barker, Gerhard Basson, Mbukeni Nkomo, Laetitia Otomo, Marshall Keyster, and et al. 2023. "Recent Progress in the Characterization, Synthesis, Delivery Procedures, Treatment Strategies, and Precision of Antimicrobial Peptides" International Journal of Molecular Sciences 24, no. 14: 11864. https://doi.org/10.3390/ijms241411864
APA StyleBakare, O. O., Gokul, A., Niekerk, L.-A., Aina, O., Abiona, A., Barker, A. M., Basson, G., Nkomo, M., Otomo, L., Keyster, M., & Klein, A. (2023). Recent Progress in the Characterization, Synthesis, Delivery Procedures, Treatment Strategies, and Precision of Antimicrobial Peptides. International Journal of Molecular Sciences, 24(14), 11864. https://doi.org/10.3390/ijms241411864