
The Influence of Single Nucleotide Polymorphisms on Vitamin D Receptor Protein Levels and Function in Chronic Liver Disease

 ,
,  ,
,  , , , ,
, , , , 
 and
and 
Abstract
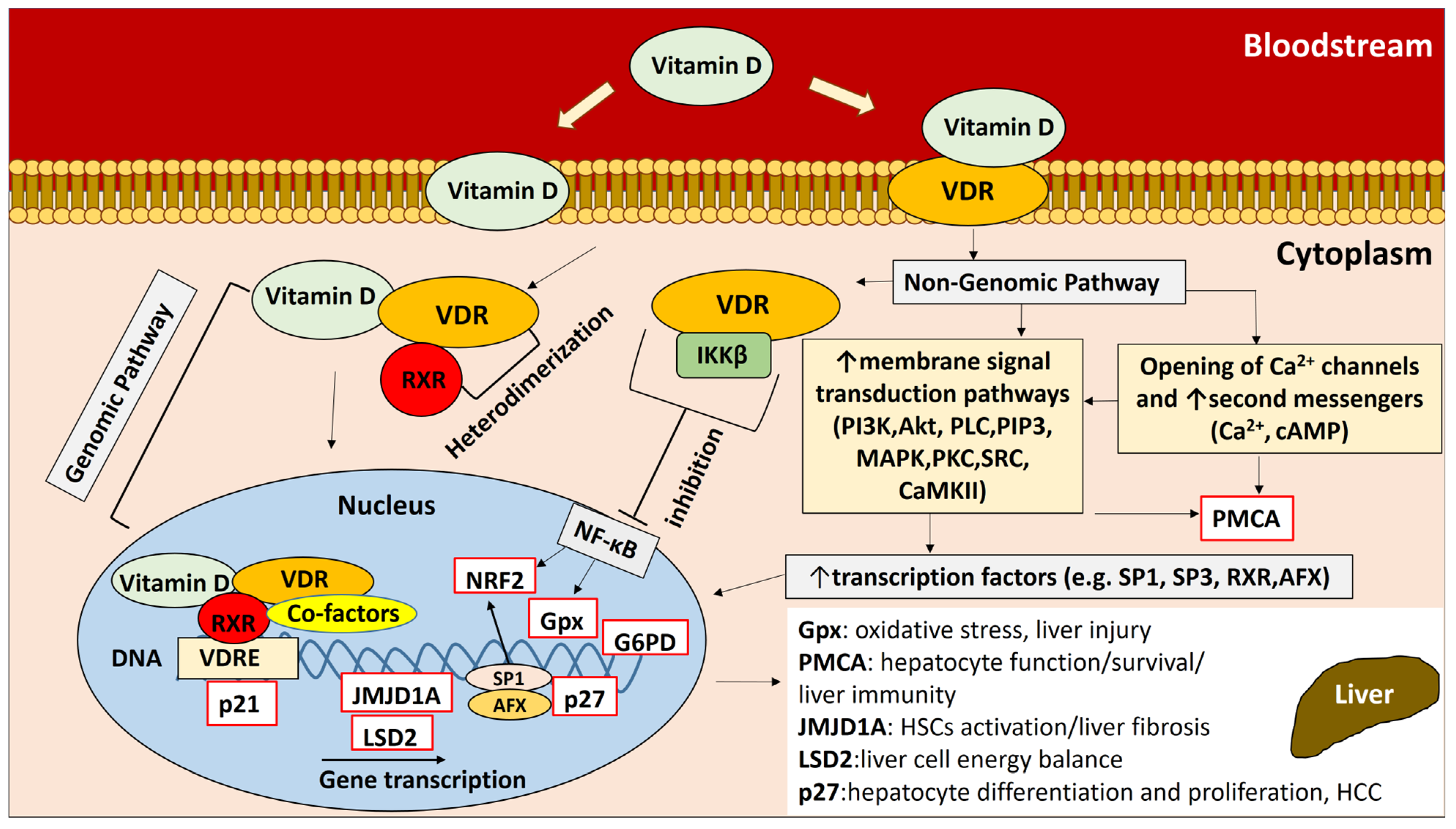
1. Introduction
2. Results
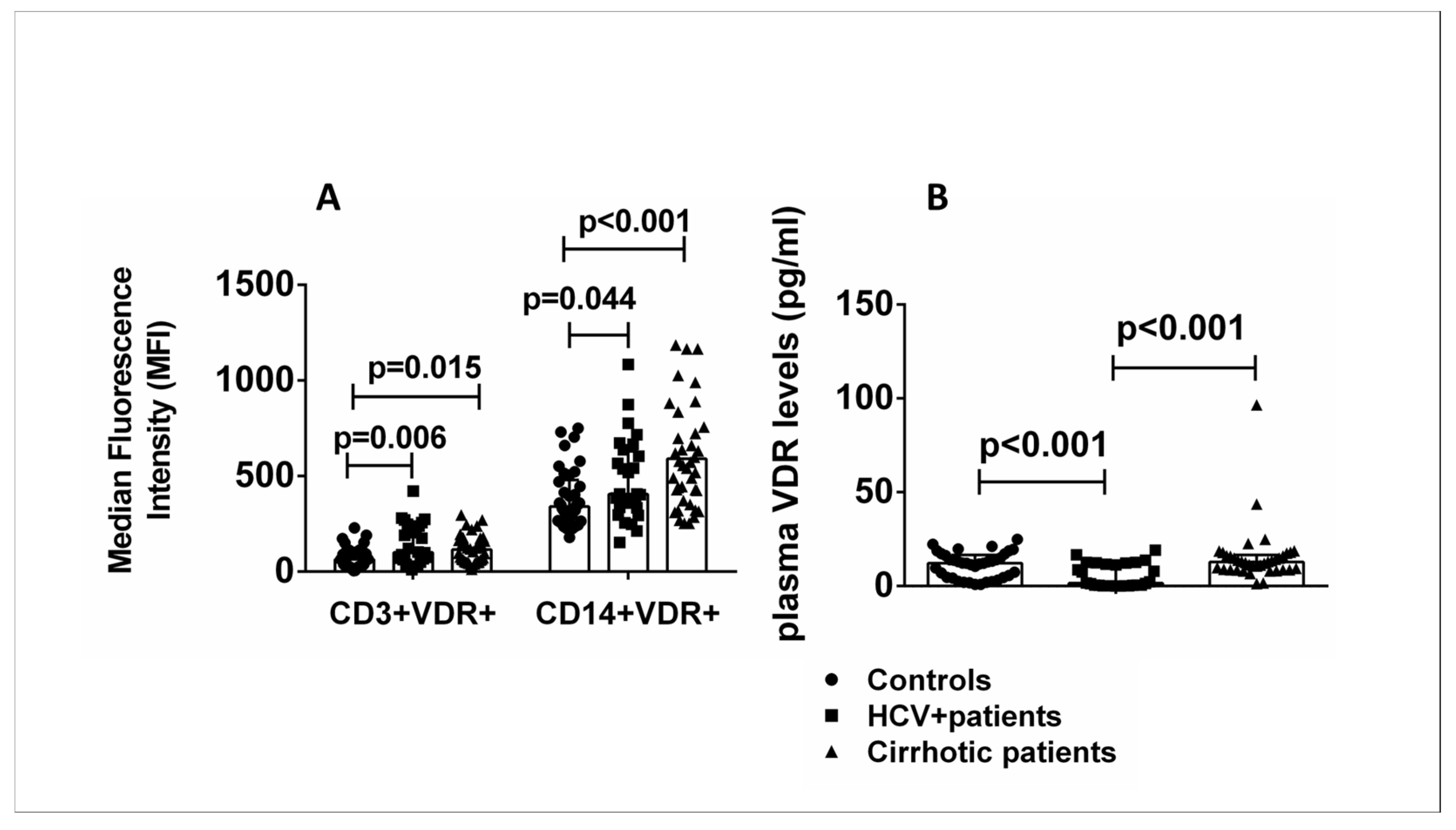
2.1. VDR Expression Levels in Specific Cell Subsets and Plasma from Healthy Controls Compared with HCV+ and Cirrhotic Patients
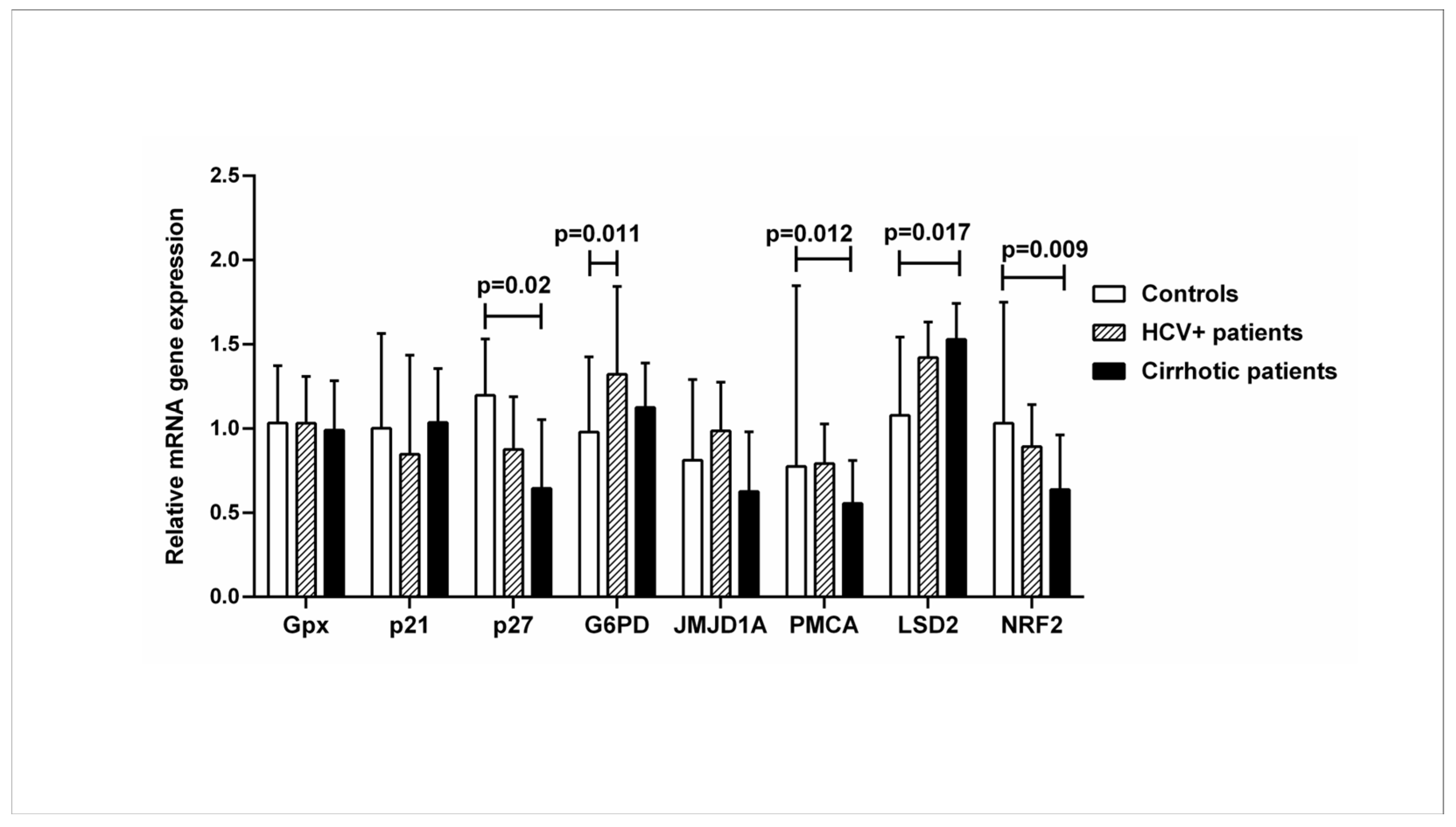
2.2. Expression of Genes Downstream of the Vitamin D-VDR Pathway in Healthy Controls Compared with HCV+ and Cirrhotic Patients
2.3. Relationship between VDR Expression Levels in Specific Cell Subsets and Liver Cirrhosis
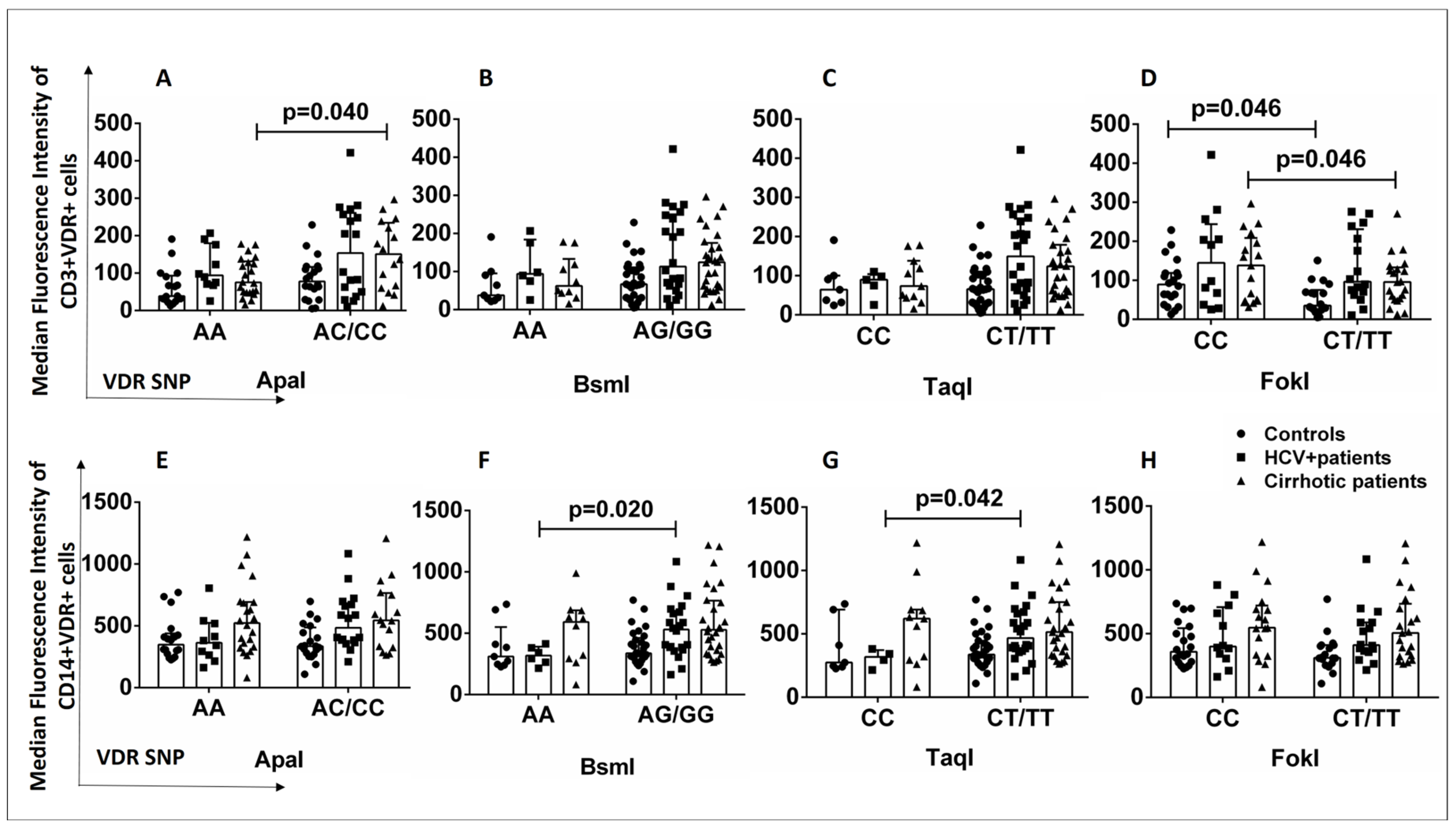
2.4. Relationship between VDR Polymorphisms and VDR Expression Levels in T Lymphocytes and Monocytes from Healthy Controls and HCV+ and Cirrhotic Patients
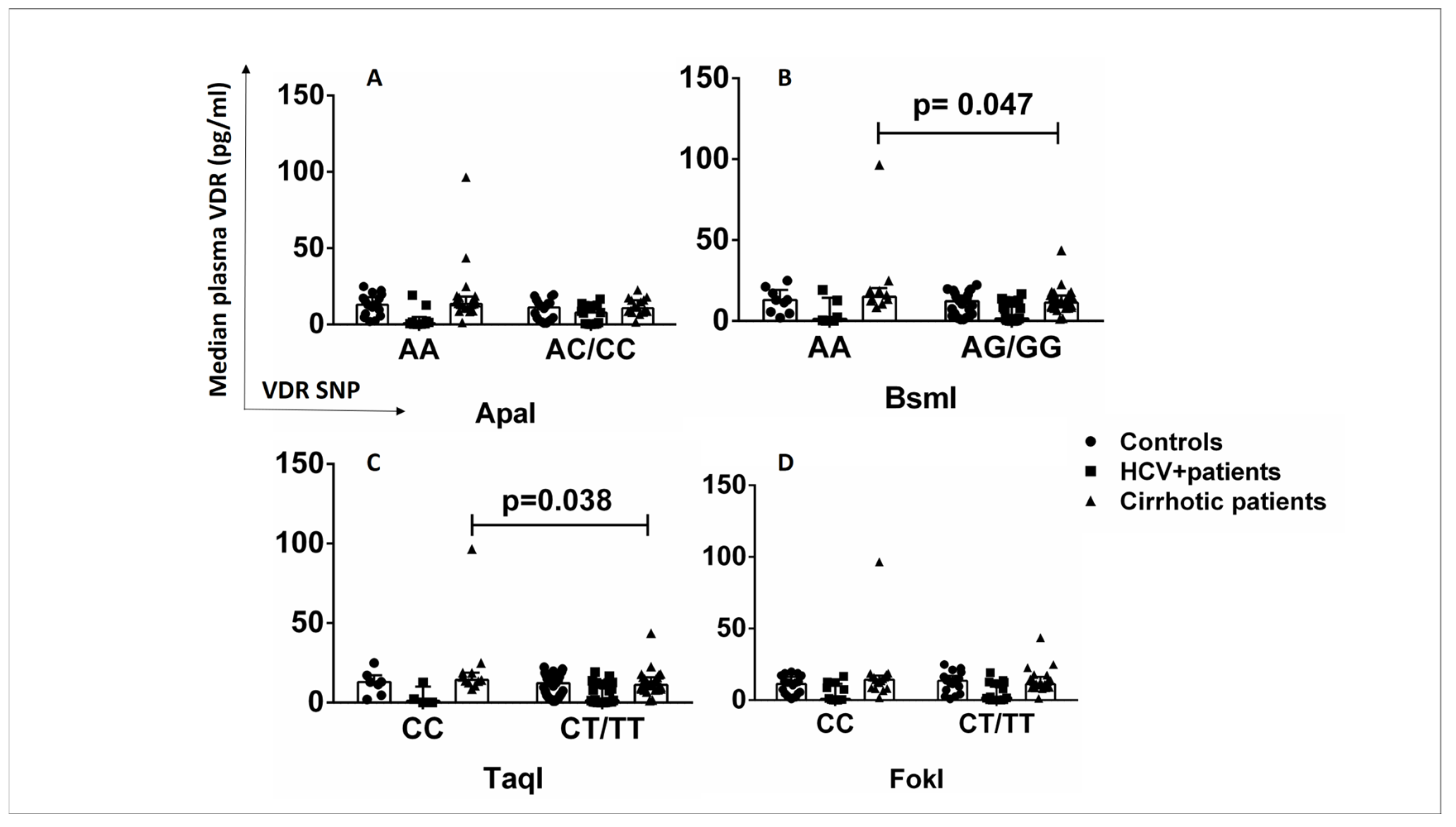
2.5. Relationship between VDR Polymorphisms and Plasma VDR Levels in Healthy Controls and HCV+ and Cirrhotic Patients
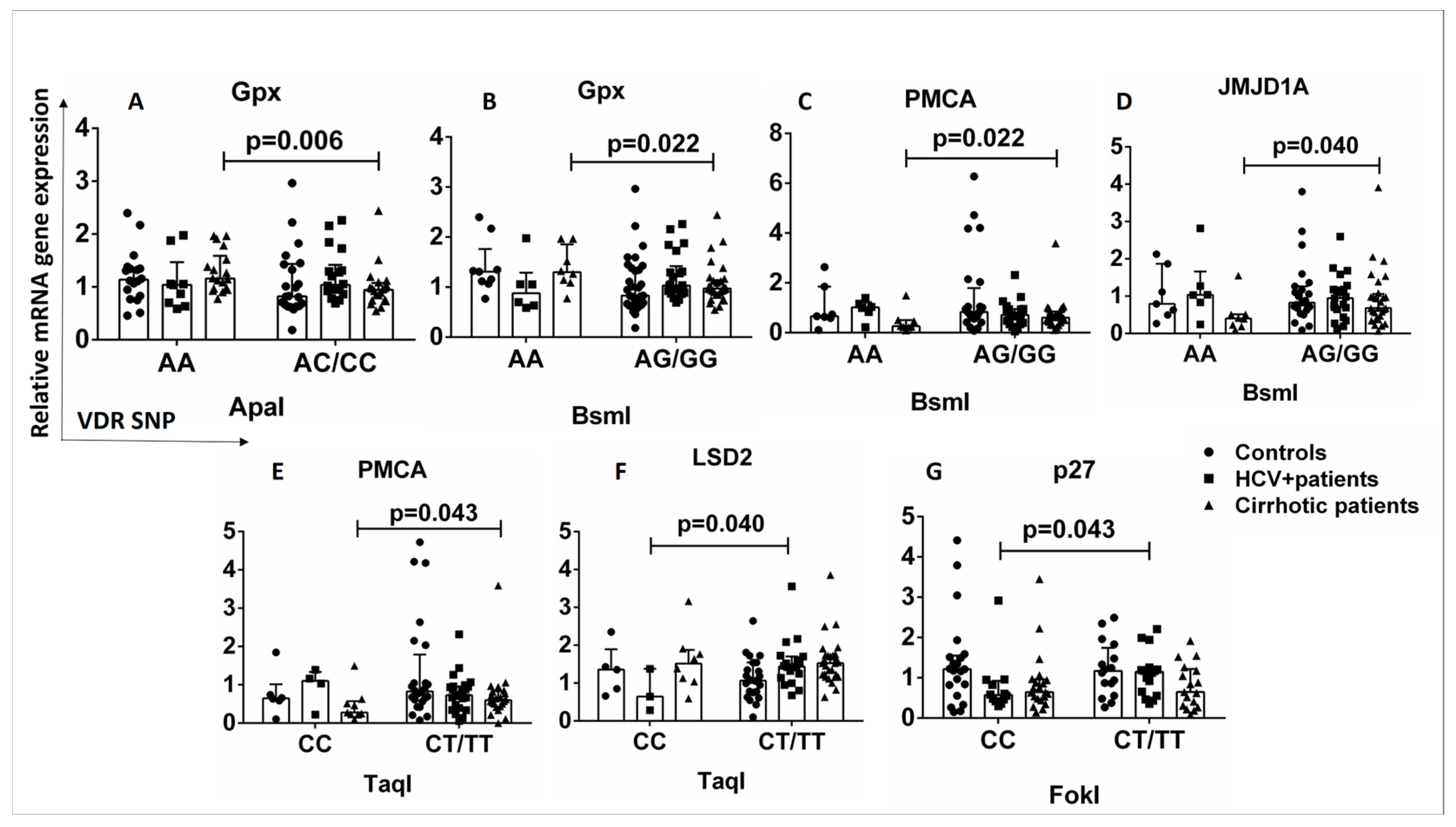
2.6. Relationship between VDR SNPs and Expression of Genes Downstream of the Vitamin D-VDR Pathway in Healthy Controls and HCV+ and Cirrhotic Patients
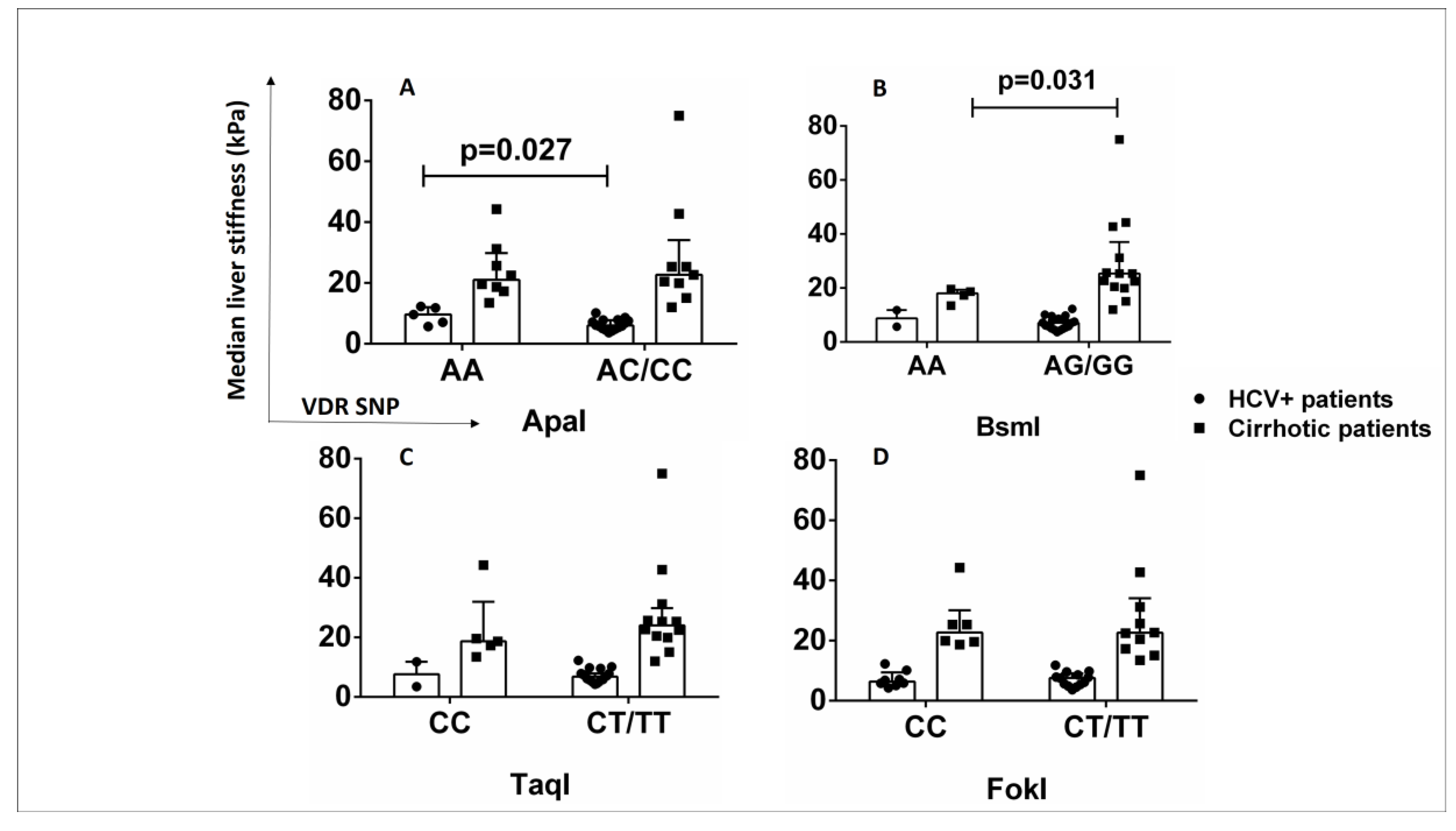
2.7. ApaI SNP Is Associated with Increased Liver Stiffness in HCV+ Patients
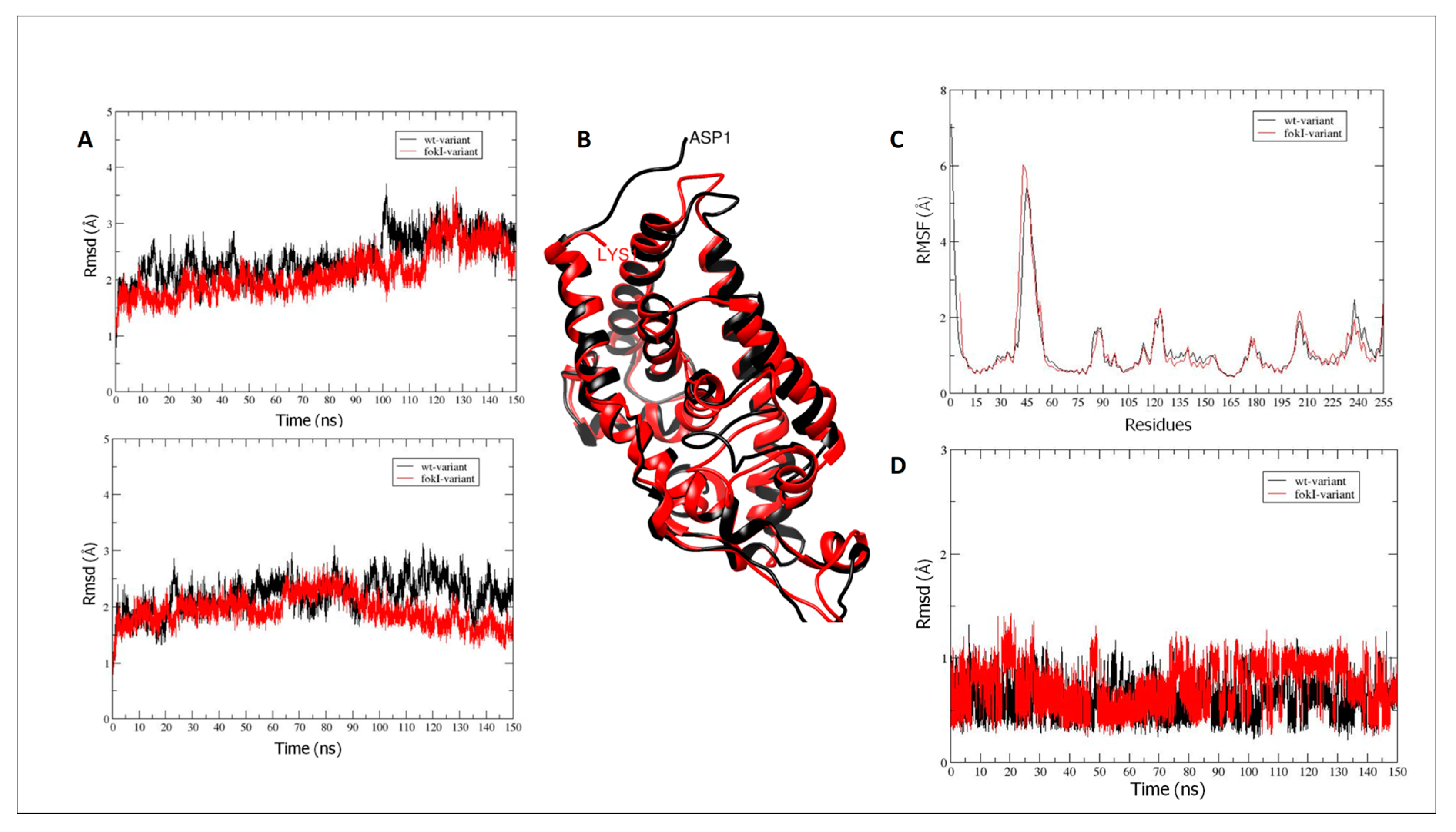
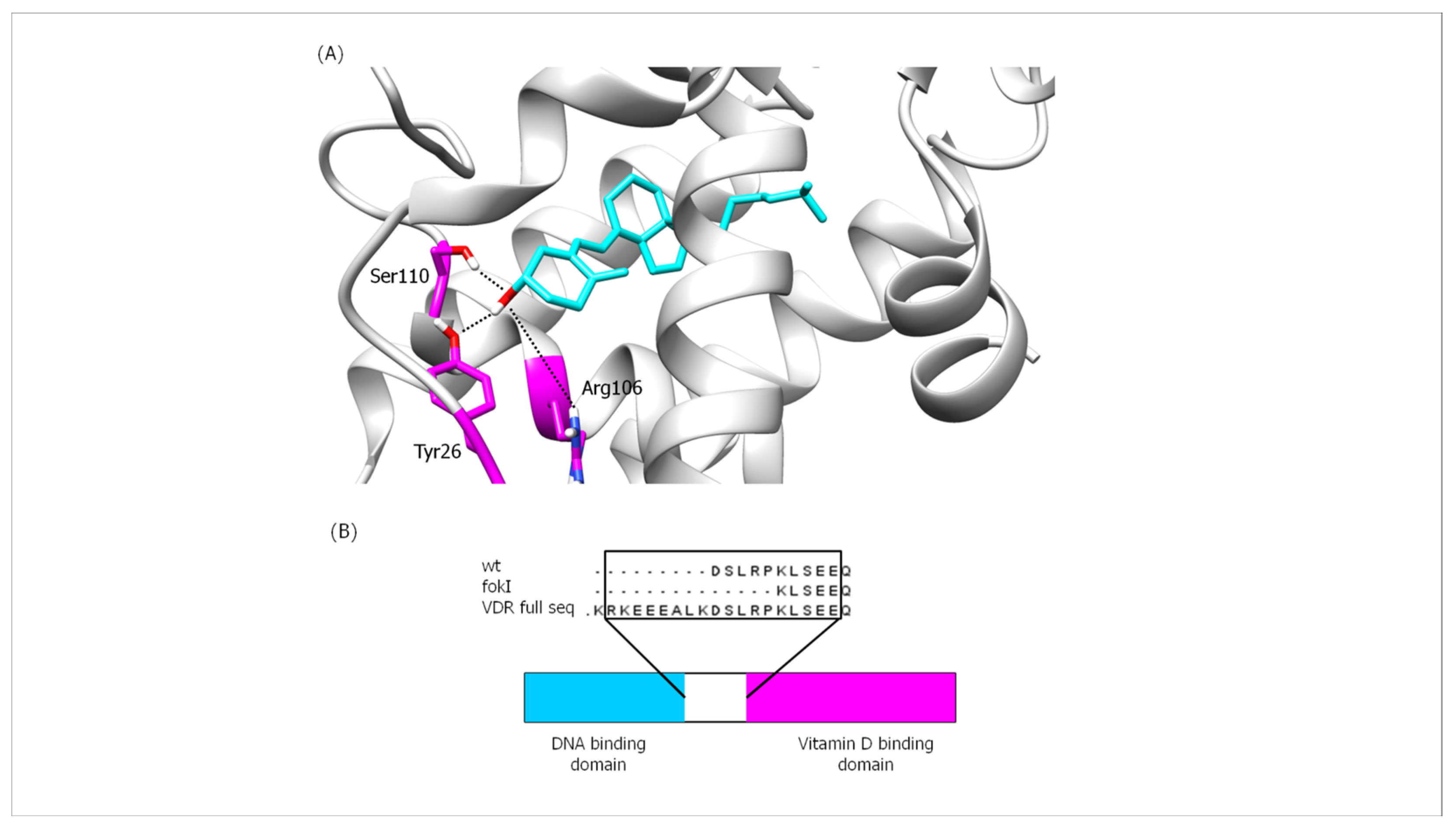
2.8. Bioinformatics Analysis of the Interaction Affinity between VDR Protein and Vitamin D in the Absence (VDR wt Isoform) and Presence of the FokI VDR Polymorphism (VDR FokI Isoform)
2.8.1. VDR Vitamin D-Binding Domain
2.8.2. Interactions between Hydrogen Bonds (Hb)
3. Discussion
4. Materials and Methods
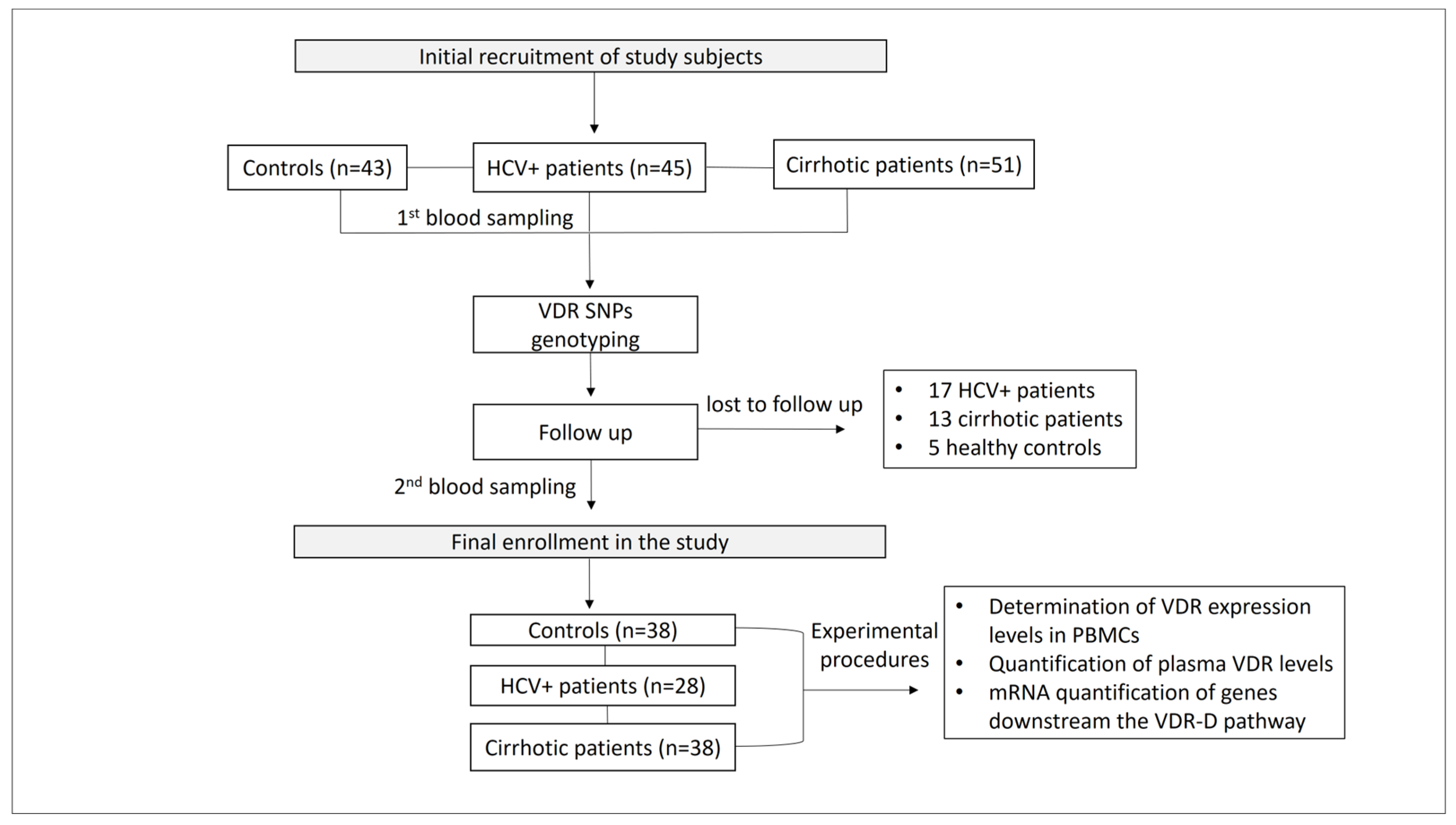
4.1. Patients
4.2. DNA Extraction
4.3. Genotyping of VDR Polymorphisms
4.4. Flow Cytometry Analysis of Peripheral Blood Mononuclear Cells
4.5. Enzyme-Linked Immunosorbent Assay (ELISA)
4.6. Quantitative Real-Time PCR (qPCR)
4.7. Structure Preparation and Molecular Dynamics (MD) Simulations
- Structure preparation: The crystallographic structure with PDB code 1ie91 was used to generate the parameters for the wild-type (wt) domain of the VDR and the FokI variant. USCF Chimera (v1.11.2) [74] was used to design the vitamin D molecule optimized for ab initio calculations using GAMESS software version 2019 R1 [75].MD simulations: MD simulations were performed using AMBER14 software [76]. The ff14SB [77] and GAFF [78] force fields were used to assign partial atomic charges for the amino acid residues and the vitamin D molecule, respectively. A detailed description of the methods used for the design and MD simulations [79,80,81,82,83,84,85,86,87,88,89,90,91,92,93] can be found in the Supplementary Material (Section S4).
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Moon, A.M.; Singal, A.G.; Tapper, E.B. Contemporary Epidemiology of Chronic Liver Disease and Cirrhosis. Clin. Gastroenterol. Hepatol. 2020, 18, 2650–2666. [Google Scholar] [CrossRef]
- Di Marco, L.; La Mantia, C.; Di Marco, V. Hepatitis C: Standard of Treatment and What to Do for Global Elimination. Viruses 2022, 14, 505. [Google Scholar] [CrossRef]
- Khatun, M.; Ray, R.B. Mechanisms Underlying Hepatitis C Virus-Associated Hepatic Fibrosis. Cells 2019, 8, 1249. [Google Scholar] [CrossRef]
- Liu, Y.B.; Chen, M.K. Epidemiology of liver cirrhosis and associated complications: Current knowledge and future directions. World J. Gastroenterol. 2022, 28, 5910–5930. [Google Scholar] [CrossRef]
- Triantos, C.; Aggeletopoulou, I.; Thomopoulos, K.; Mouzaki, A. Vitamin D-Liver Disease Association: Biological Basis and Mechanisms of Action. Hepatology 2021, 74, 1065–1073. [Google Scholar] [CrossRef]
- Christakos, S.; Dhawan, P.; Verstuyf, A.; Verlinden, L.; Carmeliet, G. Vitamin D: Metabolism, Molecular Mechanism of Action, and Pleiotropic Effects. Physiol. Rev. 2016, 96, 365–408. [Google Scholar] [CrossRef] [PubMed]
- Hanel, A.; Malmberg, H.R.; Carlberg, C. Genome-wide effects of chromatin on vitamin D signaling. J. Mol. Endocrinol. 2020, 64, R45–R56. [Google Scholar] [CrossRef]
- Bikle, D. Nonclassic actions of vitamin D. J. Clin. Endocrinol. Metab. 2009, 94, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Masuyama, H.; Brownfield, C.M.; St-Arnaud, R.; MacDonald, P.N. Evidence for ligand-dependent intramolecular folding of the AF-2 domain in vitamin D receptor-activated transcription and coactivator interaction. Mol. Endocrinol. 1997, 11, 1507–1517. [Google Scholar] [CrossRef]
- Saramäki, A.; Banwell, C.M.; Campbell, M.J.; Carlberg, C. Regulation of the human p21(waf1/cip1) gene promoter via multiple binding sites for p53 and the vitamin D3 receptor. Nucleic Acids Res. 2006, 34, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Pereira, F.; Barbáchano, A.; Singh, P.K.; Campbell, M.J.; Muñoz, A.; Larriba, M.J. Vitamin D has wide regulatory effects on histone demethylase genes. Cell Cycle 2012, 11, 1081–1089. [Google Scholar] [CrossRef] [PubMed]
- Szymczak-Pajor, I.; Śliwińska, A. Analysis of Association between Vitamin D Deficiency and Insulin Resistance. Nutrients 2019, 11, 794. [Google Scholar] [CrossRef]
- Fleet, J.C. Rapid, membrane-initiated actions of 1,25 dihydroxyvitamin D: What are they and what do they mean? J. Nutr. 2004, 134, 3215–3218. [Google Scholar] [CrossRef]
- Dwivedi, P.P.; Gao, X.H.; Tan, J.C.T.; Evdokiou, A.; Ferrante, A.; Morris, H.A.; May, B.K.; Hii, C.S.T. A role for the phosphatidylinositol 3-kinase--protein kinase C zeta-Sp1 pathway in the 1,25-dihydroxyvitamin D3 induction of the 25-hydroxyvitamin D3 24-hydroxylase gene in human kidney cells. Cell. Signal. 2010, 22, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Norman, A.W. Minireview: Vitamin D receptor: New assignments for an already busy receptor. Endocrinology 2006, 147, 5542–5548. [Google Scholar] [CrossRef] [PubMed]
- Hii, C.S.; Ferrante, A. The Non-Genomic Actions of Vitamin D. Nutrients 2016, 8, 135. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.J.; Liu, Z.G. Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, J.; Ge, X.; Du, J.; Deb, D.K.; Li, Y.C. Vitamin D receptor inhibits nuclear factor κB activation by interacting with IκB kinase β protein. J. Biol. Chem. 2013, 288, 19450–19458. [Google Scholar] [CrossRef]
- Wagle, A.; Jivraj, S.; Garlock, G.L.; Stapleton, S.R. Insulin regulation of glucose-6-phosphate dehydrogenase gene expression is rapamycin-sensitive and requires phosphatidylinositol 3-kinase. J. Biol. Chem. 1998, 273, 14968–14974. [Google Scholar] [CrossRef]
- Doroudi, M.; Schwartz, Z.; Boyan, B.D. Membrane-mediated actions of 1,25-dihydroxy vitamin D3: A review of the roles of phospholipase A2 activating protein and Ca2+/calmodulin-dependent protein kinase II. J. Steroid Biochem. Mol. Biol. 2015, 147, 81–84. [Google Scholar] [CrossRef]
- Kuo, T.H.; Liu, B.F.; Diglio, C.; Tsang, W. Regulation of the plasma membrane calcium pump gene expression by two signal transduction pathways. Arch. Biochem. Biophys. 1993, 305, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.C.; Chen, J.Y.; Hung, W.C. Vitamin D3 receptor/Sp1 complex is required for the induction of p27Kip1 expression by vitamin D3. Oncogene 2004, 23, 4856–4861. [Google Scholar] [CrossRef] [PubMed]
- Stahl, M.; Dijkers, P.F.; Kops, G.J.P.L.; Lens, S.M.A.; Coffer, P.J.; Burgering, B.M.T.; Medema, R.H. The forkhead transcription factor FoxO regulates transcription of p27Kip1 and Bim in response to IL-2. J. Immunol. 2002, 168, 5024–5031. [Google Scholar] [CrossRef]
- Medema, R.H.; Kops, G.J.; Bos, J.L.; Burgering, B.M. AFX-like Forkhead transcription factors mediate cell-cycle regulation by Ras and PKB through p27kip1. Nature 2000, 404, 782–787. [Google Scholar] [CrossRef] [PubMed]
- Siswanto, F.M.; Oguro, A.; Imaoka, S. Sp1 is a substrate of Keap1 and regulates the activity of CRL4AWDR23 ubiquitin ligase toward Nrf2. J. Biol. Chem. 2021, 296, 100704. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Guo, L.; Yang, Y.; Wang, Y.; Xia, S.; Gong, H.; Zhang, B.K.; Yan, M. Dissecting the Crosstalk Between Nrf2 and NF-κB Response Pathways in Drug-Induced Toxicity. Front. Cell Dev. Biol. 2022, 9, 809952. [Google Scholar] [CrossRef]
- Gascon-Barré, M.; Demers, C.; Mirshahi, A.; Néron, S.; Zalzal, S.; Nanci, A. The normal liver harbors the vitamin D nuclear receptor in nonparenchymal and biliary epithelial cells. Hepatology 2003, 37, 1034–1042. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.; Zhou, Y.; Wang, W.; Scott, J.; Kim, K.; Sun, Z.; Guo, L.; Lu, Y.; Gonzales, N.M.; Wu, H.; et al. Vitamin D Receptor Activation in Liver Macrophages Ameliorates Hepatic Inflammation, Steatosis, and Insulin Resistance in Mice. Hepatology 2020, 71, 1559–1574. [Google Scholar] [CrossRef] [PubMed]
- Elangovan, H.; Chahal, S.; Gunton, J.E. Vitamin D in liver disease: Current evidence and potential directions. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 4, 907–916. [Google Scholar] [CrossRef]
- Konstantakis, C.; Tselekouni, P.; Kalafateli, M.; Triantos, C. Vitamin D deficiency in patients with liver cirrhosis. Ann. Gastroenterol. 2016, 29, 297–306. [Google Scholar] [CrossRef]
- Aggeletopoulou, I.; Thomopoulos, K.; Mouzaki, A.; Triantos, C. Vitamin D-VDR Novel Anti-Inflammatory Molecules-New Insights into Their Effects on Liver Diseases. Int. J. Mol. Sci. 2022, 23, 8465. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Tu, X.; Zhu, Y.; Zhou, L.; Pfeiffer, T.; Feltens, R.; Stoecker, W.; Zhong, R. Genetic association of vitamin D receptor polymorphisms with autoimmune hepatitis and primary biliary cirrhosis in the Chinese. J. Gastroenterol. Hepatol. 2005, 20, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Kempińska-Podhorecka, A.; Wunsch, E.; Jarowicz, T.; Raszeja-Wyszomirska, J.; Loniewska, B.; Kaczmarczyk, M.; Milkiewicz, M.; Milkiewicz, P. Vitamin d receptor polymorphisms predispose to primary biliary cirrhosis and severity of the disease in polish population. Gastroenterol. Res. Pract. 2012, 2012, 408723. [Google Scholar] [CrossRef] [PubMed]
- Hoan, N.X.; Khuyen, N.; Giang, D.P.; Binh, M.T.; Toan, N.L.; Anh, D.T.; Trung, N.T.; Bang, M.H.; Meyer, C.G.; Velavan, T.P.; et al. Vitamin D receptor ApaI polymorphism associated with progression of liver disease in Vietnamese patients chronically infected with hepatitis B virus. BMC Med. Genet. 2019, 20, 201. [Google Scholar] [CrossRef] [PubMed]
- Barooah, P.; Saikia, S.; Bharadwaj, R.; Sarmah, P.; Bhattacharyya, M.; Goswami, B.; Medhi, S. Role of VDR, GC, and CYP2R1 Polymorphisms in the Development of Hepatocellular Carcinoma in Hepatitis C Virus-Infected Patients. Genet. Test. Mol. Biomark. 2019, 23, 325–331. [Google Scholar] [CrossRef]
- Baur, K.; Mertens, J.C.; Schmitt, J.; Iwata, R.; Stieger, B.; Eloranta, J.J.; Frei, P.; Stickel, F.; Dill, M.T.; Seifert, B.; et al. Combined effect of 25-OH vitamin D plasma levels and genetic vitamin D receptor (NR 1I1) variants on fibrosis progression rate in HCV patients. Liver Int. 2012, 32, 635–643. [Google Scholar] [CrossRef]
- Arai, T.; Atsukawa, M.; Tsubota, A.; Koeda, M.; Yoshida, Y.; Okubo, T.; Nakagawa, A.; Itokawa, N.; Kondo, C.; Nakatsuka, K.; et al. Association of vitamin D levels and vitamin D-related gene polymorphisms with liver fibrosis in patients with biopsy-proven nonalcoholic fat-ty liver disease. Dig. Liver Dis. 2019, 51, 1036–1042. [Google Scholar] [CrossRef]
- Triantos, C.; Aggeletopoulou, I.; Kalafateli, M.; Spantidea, P.I.; Vourli, G.; Diamantopou-lou, G.; Tapratzi, D.; Michalaki, M.; Manolakopoulos, S.; Gogos, C.; et al. Prognostic significance of vitamin D receptor (VDR) gene polymorphisms in liver cirrhosis. Sci. Rep. 2018, 8, 14065. [Google Scholar] [CrossRef]
- Wan, L.Y.; Zhang, Y.Q.; Chen, M.D.; Du, Y.Q.; Liu, C.B.; Wu, J.F. Relationship between Structure and Conformational Change of the Vitamin D Receptor Ligand Binding Domain in 1α,25-Dihydroxyvitamin D3 Signaling. Molecules 2015, 20, 20473–20486. [Google Scholar] [CrossRef]
- Ferrer-Suay, S.; Alonso-Iglesias, E.; Tortajada-Girbés, M.; Carrasco-Luna, J.; Codoñer-Franch, P. Vitamin D receptor gene ApaI and FokI polymorphisms and its association with inflammation and oxidative stress in vitamin D sufficient Caucasian Spanish children. Transl. Pediatr. 2021, 10, 103–111. [Google Scholar] [CrossRef]
- Wang, X.; Cheng, W.; Ma, Y.; Zhu, J. Vitamin D receptor gene FokI but not TaqI, ApaI, BsmI polymorphism is associated with Hashimoto’s thyroiditis: A meta-analysis. Sci. Rep. 2017, 7, 41540. [Google Scholar] [CrossRef] [PubMed]
- Yamagata, M.; Nakajima, S.; Tokita, A.; Sakai, N.; Yanagihara, I.; Yabuta, K.; Ozono, K. Analysis of the stable levels of messenger RNA derived from different polymorphic alleles in the vitamin D receptor gene. J. Bone Miner. Metab. 1999, 17, 164–170. [Google Scholar] [CrossRef] [PubMed]
- van Etten, E.; Verlinden, L.; Giulietti, A.; Ramos-Lopez, E.; Branisteanu, D.D.; Ferreira, G.B.; Overbergh, L.; Verstuyf, A.; Bouillon, R.; Roep, B.O.; et al. The vitamin D receptor gene FokI polymorphism: Functional impact on the immune system. Eur. J. Immunol. 2007, 37, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Dong, B.; Kim, K.H.; Choi, S.; Sun, Z.; Wu, N.; Wu, Y.; Scott, J.; Moore, D.D. Vitamin D Receptor Activation in Liver Macrophages Protects Against Hepatic Endoplasmic Reticulum Stress in Mice. Hepatology 2020, 71, 1453–1466. [Google Scholar] [CrossRef]
- Bell, S.J. The effect of vitamin D3 on hepcidin and IL-8 expression in monocytes. J. Hematol. 2013, 2, 1–7. [Google Scholar] [CrossRef]
- Khashim Alswailmi, F.; Shah, S.I.A.; Nawaz, H.; Al-Mazaideh, G.M. Molecular Mechanisms of Vitamin D-Mediated Immunomodulation. Galen. Med. J. 2021, 10, e2097. [Google Scholar] [CrossRef]
- Ding, N.; Yu, R.T.; Subramaniam, N.; Sherman, M.H.; Wilson, C.; Rao, R.; Leblanc, M.; Coulter, S.; He, M.; Scott, C.; et al. A vitamin D receptor/SMAD genomic circuit gates hepatic fibrotic response. Cell 2013, 153, 601–613. [Google Scholar] [CrossRef]
- Scalioni, L.P.; Santos, B.R.D.; Spritzer, P.M.; Villela-Nogueira, C.A.; Lewis-Ximenez, L.L.; Pollo-Flores, P.; Cathala Esberard, E.B.; Brandao-Mello, C.E.; Lampe, E.; Villar, L.M. Impact of vitamin D receptor and binding protein gene polymorphisms in clinical and laboratory data of HCV patients: Cross sectional study. Medicine 2018, 97, e9881. [Google Scholar] [CrossRef]
- Gisbert-Ferrándiz, L.; Cosin-Roger, J.; Hernández, C.; Macias-Ceja, D.C.; Ortiz-Masiá, D.; Salvador, P.; Wildenberg, M.E.; Esplugues, J.V.; Alos, R.; Navarro, F.; et al. The vitamin D receptor Taq I polymorphism is associated with reduced VDR and in-creased PDIA3 protein levels in human intestinal fibroblasts. J. Steroid Biochem. Mol. Biol. 2020, 202, 105720. [Google Scholar] [CrossRef]
- Barchetta, I.; Carotti, S.; Labbadia, G.; Gentilucci, U.V.; Muda, A.O.; Angelico, F.; Silecchia, G.; Leonetti, F.; Fraioli, A.; Picatdi, A.; et al. Liver vitamin D receptor, CYP2R1, and CYP27A1 expression: Relationship with liver histology and vitamin D3 levels in patients with nonalcoholic steatohepatitis or hepatitis C virus. Hepatology 2012, 56, 2180–2187. [Google Scholar] [CrossRef]
- Petta, S.; Grimaudo, S.; Tripodo, C.; Cabibi, D.; Calvaruso, M.; Di Cristina, A.; Guarnotta, C.; Macaluso, F.S.; Minissale, M.G.; Marchesini, G.; et al. The hepatic expression of vita-min D receptor is inversely associated with the severity of liver damage in genotype 1 chronic hepatitis C patients. J. Clin. Endocrinol. Metab. 2015, 100, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Behroozian, R.; Bayazidchi, M.; Rasooli, J. Systemic Inflammatory Response Syndrome and MELD Score in Hospital Outcome of Patients with Liver Cirrhosis. Middle East. J. Dig. Dis. 2012, 4, 168–172. [Google Scholar] [PubMed]
- Li, A.N.; Li, S.; Zhang, Y.J.; Xu, X.R.; Chen, Y.M.; Li, H.B. Resources and biological activities of natural polyphenols. Nutrients 2014, 6, 6020–6047. [Google Scholar] [CrossRef]
- Singal, A.K.; Jampana, S.C.; Weinman, S.A. Antioxidants as therapeutic agents for liver disease. Liver Int. 2011, 31, 1432–1448. [Google Scholar] [CrossRef]
- Medina, J.; Moreno-Otero, R. Pathophysiological basis for antioxidant therapy in chronic liver disease. Drugs 2005, 65, 2445–2461. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, S.; Takahashi, S.; Sasaki, T.; Kumagai, T.; Nagata, K. Progression of alcoholic and non-alcoholic steatohepatitis: Common metabolic aspects of innate immune system and oxidative stress. Drug. Metab. Pharmacokinet. 2011, 26, 30–46. [Google Scholar] [CrossRef]
- Cichoż-Lach, H.; Michalak, A. Oxidative stress as a crucial factor in liver diseases. World J. Gastroenterol. 2014, 20, 8082–8091. [Google Scholar] [CrossRef]
- Delgado-Coello, B.; Trejo, R.; Mas-Oliva, J. Is there a specific role for the plasma membrane Ca2+-ATPase in the hepatocyte? Mol. Cell. Biochem. 2006, 285, 1–15. [Google Scholar] [CrossRef]
- Merino-Wong, M.; Niemeyer, B.A.; Alansary, D. Plasma Membrane Calcium ATPase Regulates Stoichiometry of CD4+ T-Cell Compartments. Front. Immunol. 2021, 12, 687242. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Wang, S.; Zhao, Y.; Lin, C.; Zhong, F.; Jin, L.; He, F.; Wang, H. Histone H3K9 demethylase JMJD1A modulates hepatic stellate cells activation and liver fibrosis by epigenetically regulating peroxisome proliferator-activated receptor γ. FASEB J. 2015, 29, 1830–1841. [Google Scholar] [CrossRef]
- Karytinos, A.; Forneris, F.; Profumo, A.; Ciossani, G.; Battaglioli, E.; Binda, C.; Mattevi, A. A novel mammalian flavin-dependent histone demethylase. J. Biol. Chem. 2009, 284, 17775–17782. [Google Scholar] [CrossRef]
- Nagaoka, K.; Hino, S.; Sakamoto, A.; Anan, K.; Takase, R.; Umehara, T.; Yokoyama, S.; Sasaki, Y.; Nakao, M. Lysine-specific demethylase 2 suppresses lipid influx and metabolism in hepatic cells. Mol. Cell. Biol. 2015, 35, 1068–1080. [Google Scholar] [CrossRef] [PubMed]
- Kharkar, V.; Ast, A.; Gupta, S.; Sharma, S. LSD2/KDM1B/AOF1 and human cancer path-ways: A literature review. Cancer Stud. Ther. J. 2016, 1, 1–5. [Google Scholar]
- Ilyin, G.P.; Glaise, D.; Gilot, D.; Baffet, G.; Guguen-Guillouzo, C. Regulation and role of p21 and p27 cyclin-dependent kinase inhibitors during hepatocyte differentiation and growth. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 285, G115–G127. [Google Scholar] [CrossRef] [PubMed]
- Costa-Guda, J.; Marinoni, I.; Molatore, S.; Pellegata, N.S.; Arnold, A. Somatic mutation and germline sequence abnormalities in CDKN1B, encoding p27Kip1, in sporadic parathyroid adenomas. J. Clin. Endocrinol. Metab. 2011, 96, E701–E706. [Google Scholar] [CrossRef] [PubMed]
- Marinoni, I.; Pellegata, N.S. p27kip1: A new multiple endocrine neoplasia gene? Neuroendocrinology 2011, 93, 19–28. [Google Scholar] [CrossRef]
- Matsuda, Y.; Wakai, T.; Hirose, Y.; Osawa, M.; Fujimaki, S.; Kubota, M. p27 Is a critical prognostic biomarker in non-alcoholic steatohepatitis-related hepatocellular carcinoma. Int. J. Mol. Sci. 2013, 14, 23499–23515. [Google Scholar] [CrossRef] [PubMed]
- Chorley, B.N.; Wang, X.; Campbell, M.R.; Pittman, G.S.; Noureddine, M.A.; Bell, D.A. Discovery and verification of functional single nucleotide polymorphisms in regulatory genomic regions: Current and developing technologies. Mutat. Res. 2008, 659, 147–157. [Google Scholar] [CrossRef]
- Sadee, W.; Wang, D.; Papp, A.C.; Pinsonneault, J.K.; Smith, R.M.; Moyer, R.A.; Johnson, A.D. Pharmacogenomics of the RNA world: Structural RNA polymorphisms in drug therapy. Clin. Pharmacol. Ther. 2011, 89, 355–365. [Google Scholar] [CrossRef]
- Drysdale, C.M.; McGraw, D.W.; Stack, C.B.; Stephens, J.C.; Judson, R.S.; Nandabalan, K.; Arnold, K.; Ruano, G.; Liggett, S.B. Complex promoter and coding region beta 2-adrenergic receptor haplotypes alter receptor expression and predict in vivo responsiveness. Proc. Natl. Acad. Sci. USA 2000, 97, 10483–10488. [Google Scholar] [CrossRef]
- Jalan, R.; Gines, P.; Olson, J.C.; Mookerjee, R.P.; Moreau, R.; Garcia-Tsao, G.; Arroyo, V.; Kamath, P.S. Acute-on chronic liver failure. J. Hepatol. 2012, 57, 1336–1348. [Google Scholar] [CrossRef] [PubMed]
- Luxenburger, H.; Neumann-Haefelin, C.; Thimme, R.; Boettler, T. HCV-Specific T Cell Responses During and After Chronic HCV Infection. Viruses 2018, 10, 645. [Google Scholar] [CrossRef]
- Kongsbak, M.; Levring, T.B.; Geisler, C.; von Essen, M.R. The vitamin d receptor and T cell function. Front. Immunol. 2013, 4, 148. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera--a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Barca, G.M.J.; Bertoni, C.; Carrington, L.; Datta, D.; De Silva, N.; Deustua, J.E.; Fedorov, D.G.; Gour, J.R.; Gunina, A.O.; Guidez, E.; et al. Recent developments in the general atomic and molecular electronic structure system. J. Chem. Phys. 2020, 152, 154102. [Google Scholar] [CrossRef]
- Case, D.A.; Babin, V.; Berryman, J.T.; Betz, R.M.; Cai, Q.; Cerutti, D.S.; Cheatham, T.E.; Darden, T.A.; Duke, R.E.; Gohlke, H.; et al. Amber 14. University of California, San Francisco. 2014. Available online: https://ambermd.org (accessed on 23 March 2022).
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. ff14SB: Improving the Accuracy of Protein Side Chain and Backbone Parameters from ff99SB. J. Chem. Theory Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wolf, R.M.; Caldwell, J.W.; Kollman, P.A.; Case, D.A. Development and testing of a general amber force field. J. Comput. Chem. 2004, 25, 1157–1174. [Google Scholar] [CrossRef] [PubMed]
- Tocchini-Valentini, G.; Rochel, N.; Wurtz, J.M.; Mitschler, A.; Moras, D. Crystal structures of the vitamin D receptor complexed to superagonist 20-epi ligands. Proc. Natl. Acad. Sci. USA 2001, 98, 5491–5496. [Google Scholar] [CrossRef]
- Schmidt, M.W.; Baldridge, K.K.; Boatz, J.A.; Elbert, S.T.; Gordon, M.S.; Jensen, J.H.; Koseki, S.; Matsunaga, N.; Nguyen, K.A.; Su, S.; et al. General atomic and molecular electronic structure system. J. Comput. Chem. 1993, 14, 1347–1363. [Google Scholar] [CrossRef]
- Gordon, M.S.; Schmidt, M.W. Advances in electronic structure theory: GAMESS a decade later. In Theory and Applications of Computational Chemistry; Elsevier: Amsterdam, The Netherlands, 2005; Charter 41; pp. 1167–1189. [Google Scholar] [CrossRef]
- Becke, A.D. Density-functional thermochemistry. III. The role of exact exchange. J. Chem. Phys. 1993, 98, 2155–2160. [Google Scholar] [CrossRef]
- Pople, J.A.; Gill, P.M.; Johnson, B.G. Kohn-Sham density-functional theory within a finite basis set. Chem. Phys. Lett. 1992, 199, 557–560. [Google Scholar] [CrossRef]
- Hertwig, R.H.; Koch, W. On the parameterization of the local correlation functional. What is Becke-3-LYP? Chem. Phys. Lett. 1997, 268, 345–351. [Google Scholar] [CrossRef]
- Bayly, C.I.; Cieplak, P.; Cornell, W.; Kollman, P.A. A well-behaved electrostatic potential based method using charge restraints for deriving atomic charges: The RESP model. J. Phys. Chem. 1993, 97, 10269–10280. [Google Scholar] [CrossRef]
- Dupradeau, F.Y.; Pigache, A.; Zaffran, T.; Savineau, C.; Lelong, R.; Grivel, N.; Lelong, D.; Rosanski, W.; Cieplak, P. The R.E.D. tools: Advances in RESP and ESP charge derivation and force field library building. Phys. Chem. Chem. Phys. 2010, 12, 7821–7839. [Google Scholar] [CrossRef]
- Vanquelef, E.; Simon, S.; Marquant, G.; Garcia, E.; Klimerak, G.; Delepine, J.C.; Cieplak, P.; Dupradeau, F.Y. R.E.D. Server: A web service for deriving RESP and ESP charges and building force field libraries for new molecules and molecular fragments. Nucleic Acids Res. 2011, 39, W511–W517. [Google Scholar] [CrossRef]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Izaguirre, J.A.; Catarello, D.P.; Wozniak, J.M.; Skeel, R.D. Langevin stabilization of molecular dynamics. J. Chem. Phys. 2001, 114, 2090–2098. [Google Scholar] [CrossRef]
- Ryckaert, J.P.; Ciccotti, G.; Berendsen, H.J. Numerical integration of the cartesian equations of motion of a system with constraints: Molecular dynamics of n-alkanes. J. Comp. Phys. 1977, 23, 327–341. [Google Scholar] [CrossRef]
- Darden, T.; York, D.; Pedersen, L. Particle mesh Ewald: An N⋅log (N) method for Ewald sums in large systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef]
- Roe, D.R.; Cheatham, T.E. PTRAJ and CPPTRAJ: Software for Processing and Analysis of Molecular Dynamics Trajectory Data. J. Chem. Theory Comput. 2013, 9, 3084–3095. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Tanner, S.W.; Thompson, N.; Cheatham, T.E. Clustering Molecular Dynamics Trajectories: 1. Characterizing the Performance of Different Clustering Algorithms. J. Chem. Theory Comput. 2007, 3, 2312–2334. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Healthy Controls | Cirrhotic Patients | HCV+ Patients | ||||
|---|---|---|---|---|---|---|
| n= 38 | n = 38 | n = 28 | ||||
| n | % | n | % | n | % | |
| Sex (M/F) | 22/16 | 57.9/42.1 | 26/12 | 68.4/31.6 | 16/12 | 57.1/42.9 |
| Genotype (1/3/4) | - | - | - | - | 8/11/7 | 30.7/42.3/26.9 |
| Child Pugh stage (A/B/C) | - | - | 29/8/1 | 76.3/21.1/2.6 | - | - |
| Decompensation development (yes/no) | - | - | 9/29 | 23.7/76.3 | - | - |
| HCC development (yes/no) | - | - | 6/32 | 15.8/84.2 | - | - |
| Median | IQR | Median | IQR | Median | IQR | |
| Age (years) | 53.5 | 48–64.25 | 56.5 | 51–66 | 43.5 | 38–56.75 |
| Hb (g/dL) | - | - | 13.95 | 12.35–14.62 | 14.7 | 12.85–15.30 |
| Plt (K/μL) | - | - | 158,000 | 110,000–238,250 | 203,000 | 169,000–250,000 |
| INR | - | - | 1.09 | 0.96–1.23 | 0.94 | 0.90–1.04 |
| Creatinine (mg/dL) | - | - | 0.8 | 0.7–0.9 | 0.8 | 0.79–1 |
| SGOT (U/L) | - | - | 36 | 25–80.75 | 33.50 | 24.75–58.75 |
| SGPT (U/L) | - | - | 40 | 25–92.50 | 41 | 21.75–96.25 |
| G-GT (U/L) | - | - | 66 | 40.25–130.52 | 13.10 | 12.35–13.7 |
| ALP (U/L) | - | - | 107 | 82–141 | 65 | 52–83.50 |
| Albumin (g/dL) | - | - | 4.1 | 3.8–4.3 | 4.50 | 4.20–4.60 |
| Total Bilirubin (mg/dL) | - | - | 0.95 | 0.68–1.46 | 0.47 | 0.39–0.64 |
| Liver Stiffness (kPa) | - | - | 22.5 | 18–28.45 | 6.95 | 5.32–8.85 |
| HCV RNA (IU/L) | - | - | - | - | 122,500 | 28,500–528,000 |
| Child–Pugh score | - | - | 5.50 | 5–6.25 | - | - |
| MELD score | - | - | 7 | 6–9 | - | - |
| ApaI (rs7975232) | BsmI (rs1544410) | |||||||
| SNP location | Nucleotide change | SNP alleles | SNP location | Nucleotide change | SNP alleles | |||
| Intron 8 | A > C | A > a | Intron 8 | A > G | B > b | |||
| n (%) | AA (AA) | AC (Aa) | CC (aa) | AA (BB) | AG (Bb) | GG (bb) | ||
| Controls | 18 (47.4) | 15 (39.5) | 5 (13.2) | Reference group | 9 (23.7) | 18 (47.4) | 11 (38.9) | Reference group |
| Cirrhotics | 18 (47.4) | 11 (28.9) | 9 (23.7) | p = 0.448 | 9 (23.7) | 14 (36.8) | 15 (39.5) | p = 0.617 |
| HCV+ | 9 (32.1) | 15 (53.6) | 4 (14.3) | p = 0.487 | 7 (25) | 10 (35.7) | 11 (39.3) | p = 0.628 |
| TaqI (rs731236) | FokI (rs2228570) | |||||||
| SNP location | Nucleotide change | SNP alleles | SNP location | Nucleotide change | SNP alleles | |||
| Exon 9 | C > T | T > t | Eχon 2 | C > T | F > f | |||
| n (%) | CC (TT) | CT (Tt) | TT (tt) | CC (FF) | CT (Ff) | TT (ff) | ||
| Controls | 7 (18.4) | 18 (47.4) | 13 (34.2) | Reference group | 21 (55.3) | 15 (39.5) | 2 (5.3) | Reference group |
| Cirrhotics | 9 (23.7) | 16 (42.1) | 13 (34.2) | p = 0.875 | 21 (55.3) | 13 (34.2) | 4 (10.5) | p = 0.736 |
| HCV+ | 7 (25) | 10 (35.7) | 11 (39.3) | p = 0.653 | 13 (46.4) | 14 (50) | 1 (3.6) | p = 0.702 |
| CD3+VDR+ (MFI) | CD14+VDR+ (MFI) | Plasma VDR (pg/mL) | ||||
|---|---|---|---|---|---|---|
| Spearman’s Rho | p Value | Spearman’s Rho | p Value | Spearman’s Rho | p Value | |
| Child–Pugh (CP) score | −0.048 | 0.776 | −0.100 | 0.549 | 0.188 | 0.259 |
| MELD score | −0.197 | 0.236 | −0.347 | 0.033 | −0.010 | 0.955 |
| Gene | PCR primer (5′→3′) | PCR Product (bp) | PCR Cycling Parameters |
|---|---|---|---|
| Gpx | TATCGAGAATGTGGCGTCCC TCTTGGCGTTCTCCTGATGC | 143 | 95 °C 3 min, 95 °C 3 s, 60 °C 30 s |
| p21 | AGTCAGTTCCTTGTGGAGCC CATTAGCGCATCACAGTCGC | 184 | 95 °C 3 min, 95 °C 3 s, 63 °C 30 s |
| p27 | ACGTGCGAGTGTCTAACGG CGCCTCTTCCATGTCTCTGC | 158 | 95 °C 3 min, 95 °C 3 s, 60 °C 30 s |
| G6PD | GAAACGGTCGTACACTTCGG CCGACTGATGGAAGGCATCG | 153 | 95 °C 3 min, 95 °C 3 s, 60 °C 30 s |
| JMJD1A | CAGTTGCCTAAATGCCGA TGAATTGTAACCTCCTGAAGTG | 112 | 95 °C 3 min, 95 °C 3 s, 61 °C 30 s |
| PMCA | CAGTTATGTGGGGACGAAATG GCGGTGAGTCTTGAGTAATGC | 123 | 95 °C 3 min, 95 °C 3 s, 61 °C 30 s |
| LSD2 | GCGTGCTGATGTCTGTGATT TTGTGGGATCTGGGACCTC | 132 | 95 °C 3 min, 95 °C 3 s, 60 °C 30 s |
| NRF2 | GCGCAGACATTCCCGTTTG GACTGGGCTCTCGATGTGAC | 96 | 95 °C 3 min, 95 °C 3 s, 61 °C 30 s |
| TRPV5 | TGGCACTGTTCACCACCTTT CAATGATGGCGAAGGCGAAG | 114 | 95 °C 3 min, 95 °C 3 s, 62 °C 30 s |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tourkochristou, E.; Tsounis, E.P.; Tzoupis, H.; Aggeletopoulou, I.; Tsintoni, A.; Lourida, T.; Diamantopoulou, G.; Zisimopoulos, K.; Kafentzi, T.; de Lastic, A.-L.; et al. The Influence of Single Nucleotide Polymorphisms on Vitamin D Receptor Protein Levels and Function in Chronic Liver Disease. Int. J. Mol. Sci. 2023, 24, 11404. https://doi.org/10.3390/ijms241411404
Tourkochristou E, Tsounis EP, Tzoupis H, Aggeletopoulou I, Tsintoni A, Lourida T, Diamantopoulou G, Zisimopoulos K, Kafentzi T, de Lastic A-L, et al. The Influence of Single Nucleotide Polymorphisms on Vitamin D Receptor Protein Levels and Function in Chronic Liver Disease. International Journal of Molecular Sciences. 2023; 24(14):11404. https://doi.org/10.3390/ijms241411404
Chicago/Turabian StyleTourkochristou, Evanthia, Efthymios P. Tsounis, Haralampos Tzoupis, Ioanna Aggeletopoulou, Aggeliki Tsintoni, Theoni Lourida, Georgia Diamantopoulou, Konstantinos Zisimopoulos, Theodora Kafentzi, Anne-Lise de Lastic, and et al. 2023. "The Influence of Single Nucleotide Polymorphisms on Vitamin D Receptor Protein Levels and Function in Chronic Liver Disease" International Journal of Molecular Sciences 24, no. 14: 11404. https://doi.org/10.3390/ijms241411404
APA StyleTourkochristou, E., Tsounis, E. P., Tzoupis, H., Aggeletopoulou, I., Tsintoni, A., Lourida, T., Diamantopoulou, G., Zisimopoulos, K., Kafentzi, T., de Lastic, A.-L., Rodi, M., Tselios, T., Thomopoulos, K., Mouzaki, A., & Triantos, C. (2023). The Influence of Single Nucleotide Polymorphisms on Vitamin D Receptor Protein Levels and Function in Chronic Liver Disease. International Journal of Molecular Sciences, 24(14), 11404. https://doi.org/10.3390/ijms241411404