
Antifungal Mechanism of Phenazine-1-Carboxylic Acid against Pestalotiopsis kenyana


Abstract
1. Introduction
2. Results and Discussion
2.1. Antifungal Activity
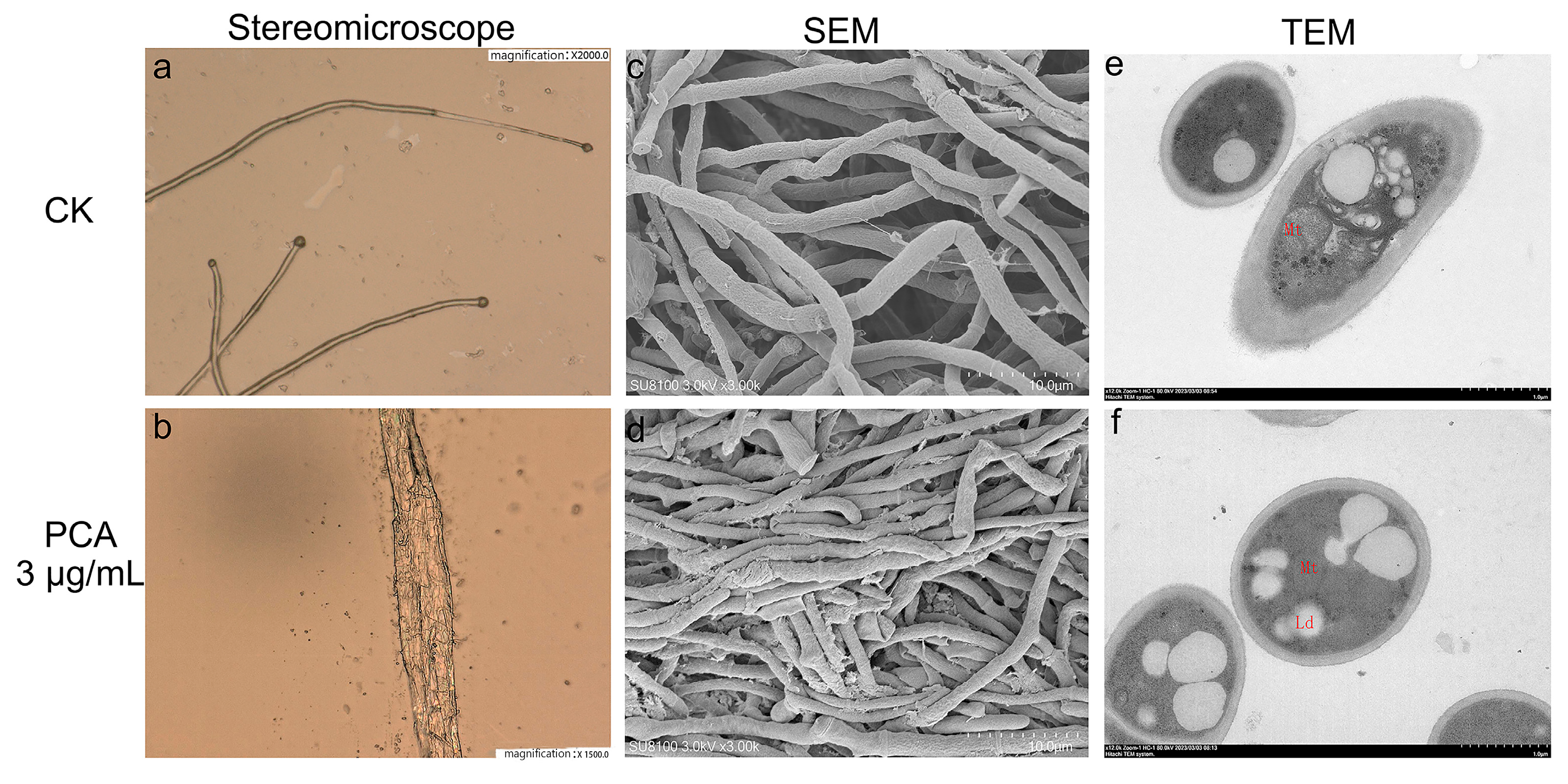
2.2. Effects on the Morphology of P. kenyana
2.3. Effect on ROS Production and Mitochondrial Membrane Potential
2.4. Effect on the Cell Membrane Permeability and Cellular Leakage
2.5. Effect on Nuclear Morphology of P. kenyana
2.6. Transcriptome Analysis
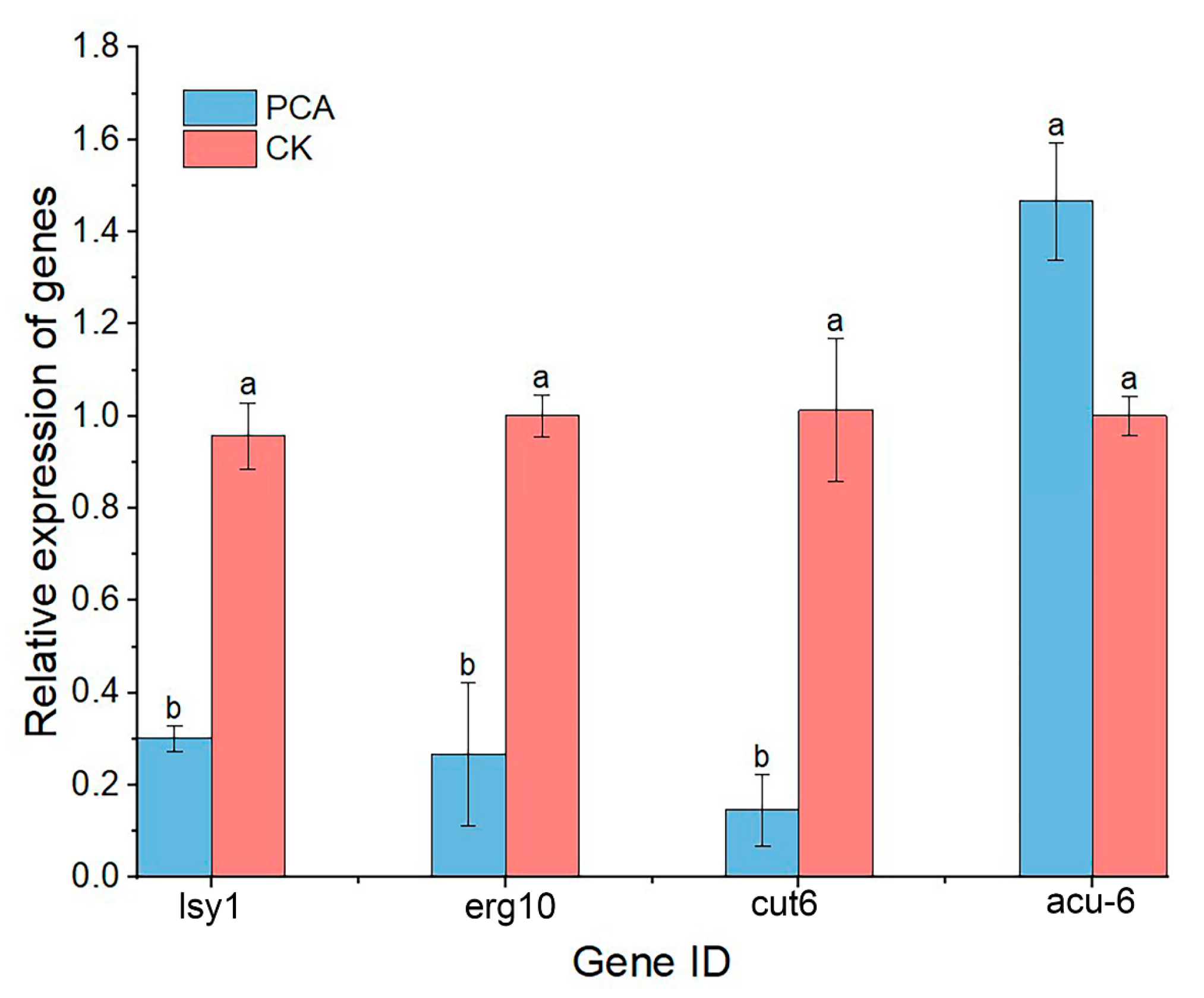
2.7. Gene Expression Analysis
3. Materials and Methods
3.1. Biomaterials
3.2. Antifungal Assay
3.3. Histocytological Assay
3.4. Reactive Oxygen Species (ROS) Accumulation and Mitochondrial Membrane Potential Assay
3.5. Observation of the Cell Membrane of P. kenyana
3.6. Nuclear Staining Assay
3.7. Transcriptome Analysis
3.8. qRT-PCR Analysis
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Tuyen, D.T.; Trung, N.T.; Thao, N.T.; Le Thanh, N.S.; Dai Nguyen, N.P.; Anh Tuyet, N.T.; Cuong, N.T.; Chan, S.S.; Khoo, K.S.; Show, P.L. Antifungal activity of secondary metabolites purified from Bacillus subtilis isolated in Vietnam and evaluated on in vitro and in vivo models. Int. Biodeterior. Biodegrad. 2023, 179, 105558. [Google Scholar] [CrossRef]
- Ren, H.; Wu, Y.; Ahmed, T.; Qi, X.; Li, B. Response of resistant and susceptible bayberry cultivars to infection of twig blight pathogen by histological observation and gibberellin related genes expression. Pathogens 2021, 10, 402. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.Y.; Lu, L.M.; Ni, H.Z.; Wang, Y.; Wang, Y.G.; Li, G.Q. First report of pestalotiopsis mangiferae and P. vismiae causing twig dieback of Myrica rubra in China. Plant Dis. 2012, 96, 588. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Voglmayr, H.; Xue, H.; Piao, C.G.; Li, Y. Morphology and phylogeny of Pestalotiopsis (Sporocadaceae, Amphisphaeriales) from Fagaceae leaves in China. Microbiol. Spectr. 2022, 10, e03272-22. [Google Scholar] [CrossRef]
- Hlaiem, S.; Yangui, I.; Della Rocca, G.; Barberini, S.; Danti, R.; Ben Jamaa, M.L. First report of Pestalotiopsis biciliata causing dieback on Quercus coccifera and Pistacia lentiscus in Tunisia. Can. J. Plant Pathol. 2022, 44, 534–541. [Google Scholar] [CrossRef]
- Zhang, C.Q.; Liu, Y.H.; Wu, H.M.; Xu, B.C.; Sun, P.L.; Xu, Z.H. Baseline sensitivity of Pestalotiopsis microspora, which causes black spot disease on Chinese hickory (Carya cathayensis), to pyraclostrobin. Crop Prot. 2012, 42, 256–259. [Google Scholar] [CrossRef]
- Das, R.; Chutia, M.; Das, K.; Jha, D.K. Factors affecting sporulation of Pestalotiopsis disseminata causing grey blight disease of Persea bombycina Kost., the primary food plant of muga silkworm. Crop Prot. 2010, 29, 963–968. [Google Scholar] [CrossRef]
- Bhuiyan, M.A.B.; Islam, S.M.N.; Bukhari, M.A.I.; Kader, M.A.; Chowdhury, M.Z.H.; Alam, M.Z.; Abdullah, H.M.; Jenny, F. First report of Pestalotiopsis microspora causing leaf blight of banana in Bangladesh. Plant Dis. 2022, 106, 1518. [Google Scholar] [CrossRef]
- Zheng, X.-R.; Zhang, M.-J.; Chen, F.-M. Occurrence of Pestalotiopsis lushanensis causing leaf blight on Buddhist pine in China. Eur. J. Plant Pathol. 2022, 162, 655–665. [Google Scholar] [CrossRef]
- Yuan, S.Q.; Wang, Y.C.; Lei, L.; Hong, J.Y.; Yi, T.Y.; Hong, Y.Y. First report of Pestalotiopsis microspora causing leaf spot on Moyeam in China. Plant Dis. 2022, 106, 1996. [Google Scholar] [CrossRef]
- Liu, C.; Luo, F.; Zhu, T.; Han, S.; Li, S. Leaf spot disease caused by Pestalotiopsis kenyana on Zanthoxylum schinifolium in Sichuan Province, China. Plant Dis. 2021, 105, 3747. [Google Scholar] [CrossRef]
- Lu, L.M.; Chen, G.Q.; Hu, X.R.; Du, D.C.; Pu, Z.X.; Peng, A.T.; Cheng, B.P. Identification of Pestalotiopsis clavispora causing brown leaf spot on Chinese bayberry in China. Can. J. Plant Pathol. 2015, 37, 397–402. [Google Scholar] [CrossRef]
- Zhou, L.; Li, J.; Chen, F.; Chen, J.; Ye, J. First report of shoot blight on Cryptomeria japonica caused by Pestalotiopsis neglecta in China. Plant Dis. 2019, 103, 2140. [Google Scholar] [CrossRef]
- Qi, M.; Xie, C.X.; Chen, Q.W.; Yu, Z.D. Pestalotiopsis trachicarpicola, a novel pathogen causes twig blight of Pinus bungeana (Pinaceae: Pinoideae) in China. Antonie Van Leeuwenhoek 2021, 114, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Yin, Q.; Qiu, C.; Yang, Y.; Guo, D.; Huang, H.; Tang, Q.; Jiang, S.; Zhao, X.; Chen, Z. Transcriptome profiling of the leaf spot pathogen, Pestalotiopsis trachicarpicola, and its host, tea (Camellia sinensis), during infection. Plant Dis. 2022, 106, 2247–2252. [Google Scholar] [CrossRef]
- Liu, C.; Guo, H.; Liu, H.; Yu, J.; Li, S.; Zhu, T.; Owusu, A.M.; Li, S. Differential metabolomics reveals pathogenesis of Pestalotiopsis kenyana causing leaf spot disease of Zanthoxylum schinifolium. J. Fungi 2022, 8, 1208. [Google Scholar] [CrossRef]
- Jing, Y.; Lian-Nan, L.; Xiao-Bo, Z.; Yue, W.; Bing, B.; Guo-Cai, Z.; Chuan-Shan, Z. Sodium pheophorbide a has photoactivated fungicidal activity against Pestalotiopsis neglecta. Pestic. Biochem. Physiol. 2019, 158, 25–31. [Google Scholar] [CrossRef]
- Yang, J.; Ji, J.Y.; Zhang, B.W.; Chen, Y.Z.; Wang, S.R.; Zhang, G.C.; Zhang, J. Transcriptome and cell wall degrading enzyme-related gene analysis of Pestalotiopsis neglecta in response to sodium pheophorbide a. Pestic. Biochem. Physiol. 2020, 169, 104639. [Google Scholar] [CrossRef]
- Ahmed, T.; Ren, H.; Noman, M.; Shahid, M.; Liu, M.; Ali, M.A.; Zhang, J.; Tian, Y.; Qi, X.; Li, B. Green synthesis and characterization of zirconium oxide nanoparticles by using a native Enterobacter sp. and its antifungal activity against bayberry twig blight disease pathogen Pestalotiopsis versicolor. NanoImpact 2021, 21, 100281. [Google Scholar] [CrossRef]
- Ruomeng, B.; Meihao, O.; Siru, Z.; Shichen, G.; Yixian, Z.; Junhong, C.; Ruijie, M.; Yuan, L.; Gezhi, X.; Xingyu, C.; et al. Degradation strategies of pesticide residue: From chemicals to synthetic biology. Synth. Syst. Biotechnol. 2023, 8, 302–313. [Google Scholar] [CrossRef]
- Xu, Y.Q. Green microorganism based antimicrobial: Shenqinmycin(M18). Fine Spec. Chem. 2004, 12, 8–9,17. [Google Scholar] [CrossRef]
- Xu, S.; Pan, X.; Luo, J.; Wu, J.; Zhou, Z.; Liang, X.; He, Y.; Zhou, M. Effects of phenazine-1-carboxylic acid on the biology of the plant-pathogenic bacterium Xanthomonas oryzae pv. oryzae. Pestic. Biochem. Physiol. 2015, 117, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, A.; Bahmani, Z.; Castaldi, S.; Masi, M.; Isticato, R.; Abdollahzadeh, J.; Amini, J.; Evidente, A. Phenazine-1-carboxylic acid (PCA), produced for the first time as an antifungal metabolite by Truncatella Angustata, a causal agent of grapevine trunk diseases (GTDs) in Iran. J. Agric. Food Chem. 2021, 69, 12143–12147. [Google Scholar] [CrossRef]
- Simionato, A.S.; Navarro, M.O.P.; de Jesus, M.L.A.; Barazetti, A.R.; da Silva, C.S.; Simoes, G.C.; Balbi-Pena, M.I.; de Mello, J.C.P.; Panagio, L.A.; de Almeida, R.S.C.; et al. The effect of phenazine-1-carboxylic acid on mycelial growth of Botrytis cinerea produced by Pseudomonas aeruginosa LV strain. Front. Microbiol. 2017, 8, 1102. [Google Scholar] [CrossRef] [PubMed]
- Roquigny, R.; Novinscak, A.; Arseneault, T.; Joly, D.L.; Filion, M. Transcriptome alteration in Phytophthora infestans in response to phenazine-1-carboxylic acid production by Pseudomonas fluorescens strain LBUM223. BMC Genom. 2018, 19, 474. [Google Scholar] [CrossRef]
- Jiang, L.; Goh, D.X.; Koh, J.H.Z.; Chan, X.; Brennan, N.A.; Barathi, V.A.; Hoang, Q.V. Applications of genomics and transcriptomics in precision medicine for myopia control or prevention. Biomolecules 2023, 13, 494. [Google Scholar] [CrossRef]
- Turra, D.; Nordzieke, D.; Vitale, S.; El Ghalid, M.; Di Pietro, A. Hyphal chemotropism in fungal pathogenicity. Semin. Cell Dev. Biol. 2016, 57, 69–75. [Google Scholar] [CrossRef]
- Zhang, M.; Ji, P.; Li, Z.; Sun, Z.; Tran, N.T.; Li, S. Catalase regulates the homeostasis of hemolymph microbiota and autophagy of the hemocytes in mud crab (Scylla paramamosain). Aquac. Rep. 2022, 25, 101237. [Google Scholar] [CrossRef]
- Liu, J.; Jia, B.; Li, Z.; Li, W. Reactive oxygen species-responsive polymer drug delivery systems. Front. Bioeng. Biotechnol. 2023, 11, 1115603. [Google Scholar] [CrossRef]
- Houston, R.; Sekine, Y.; Larsen, M.B.; Murakami, K.; Mullett, S.J.; Wendell, S.G.; Narendra, D.P.; Chen, B.B.; Sekine, S. Discovery of bactericides as an acute mitochondrial membrane damage inducer. Mol. Biol. Cell 2021, 32, ar32. [Google Scholar] [CrossRef]
- Ghosh, P.; Roy, A.; Hess, D.; Ghosh, A.; Das, S. Deciphering the mode of action of a mutant Allium sativum Leaf Agglutinin (mASAL), a potent antifungal protein on Rhizoctonia solani. BMC Microbiol. 2015, 15, 237. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Yan, D.; Li, M.; Li, D.; Zhang, S.; Fan, G.; Peng, L.; Pan, S. Ergosterol depletion under bifonazole treatment induces cell membrane damage and triggers a ROS-mediated mitochondrial apoptosis in Penicillium expansum. Fungal Biol. 2022, 126, 1–10. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, R.; Xue, H.; Bi, Y.; Li, L.; Zhang, Q.; Kouasseu, C.J.; Nan, M.; Prusky, D. Ozone controls potato dry rot development and diacetoxyscirpenol accumulation by targeting the cell membrane and affecting the growth of Fusarium sulphureus. Physiol. Mol. Plant Pathol. 2022, 118, 101785. [Google Scholar] [CrossRef]
- Ma, Y.; Gao, K.; Yu, H.; Liu, W.; Qin, Y.; Xing, R.; Liu, S.; Li, P. C-coordinated O-carboxymethyl chitosan Cu(II) complex exerts antifungal activity by disrupting the cell membrane integrity of Phytophthora capsici Leonian. Carbohydr. Polym. 2021, 261, 117821. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Wang, Z.; Zhong, T.; Guo, M.; Huang, L.; Yang, L.; Kan, J.; Zalan, Z.; Hegyi, F.; Takacs, K.; et al. Antifungal mechanisms of volatile organic compounds produced by Pseudomonas fluorescens ZX as biological fumigants against Botrytis cinerea. Microbiol. Res. 2023, 267, 127253. [Google Scholar] [CrossRef]
- Chen, B.; Co, C.; Ho, C.C. Cell shape dependent regulation of nuclear morphology. Biomaterials 2015, 67, 129–136. [Google Scholar] [CrossRef]
- Wang, W.; Wang, M.; Ruan, Y.; Tan, J.; Wang, H.; Yang, T.; Li, J.; Zhou, Q. Ginkgolic acids impair mitochondrial function by decreasing mitochondrial biogenesis and promoting FUNDC1-dependent mitophagy. J. Agric. Food Chem. 2019, 67, 10097–10106. [Google Scholar] [CrossRef]
- Luo, Z.; Liu, S.; Du, G.; Xu, S.; Zhou, J.; Chen, J. Enhanced pyruvate production in Candida glabrata by carrier engineering. Biotechnol. Bioeng. 2018, 115, 473–482. [Google Scholar] [CrossRef]
- Zhang, S.; Wakai, S.; Sasakura, N.; Tsutsumi, H.; Hata, Y.; Ogino, C.; Kondo, A. Pyruvate metabolism redirection for biological production of commodity chemicals in aerobic fungus Aspergillus oryzae. Metab. Eng. 2020, 61, 225–237. [Google Scholar] [CrossRef]
- Wu, T.-L.; Zhang, B.-Q.; Luo, X.-F.; Li, A.-P.; Zhang, S.-Y.; An, J.-X.; Zhang, Z.-J.; Liu, Y.-Q. Antifungal efficacy of sixty essential oils and mechanism of oregano essential oil against Rhizoctonia solani. Ind. Crops Prod. 2023, 191, 115975. [Google Scholar] [CrossRef]
- Chen, W.; Xia, S.; Xiao, C. Complex coacervation microcapsules by tannic acid crosslinking prolong the antifungal activity of cinnamaldehyde against Aspergillus brasiliensis. Food Biosci. 2022, 47, 101686. [Google Scholar] [CrossRef]
- Zhang, Z.J.; Jiang, Z.Y.; Zhu, Q.; Zhong, G.H. Discovery of beta-Carboline Oxadiazole Derivatives as Fungicidal Agents against Rice Sheath Blight. J. Agric. Food Chem. 2018, 66, 9598–9607. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.F.; Dai, L.X.; Zhang, Z.J.; Yang, C.J.; Du, S.S.; Wu, T.L.; He, Y.H.; Zhu, J.K.; Liu, Y.Q.; Yan, Y.F.; et al. Integrated proteomics and transcriptomics analyses reveals the possible antifungal mechanism of an indoloquinoline alkaloid neocryptolepine against Rhizoctonia solani. J. Agric. Food Chem. 2021, 69, 6455–6464. [Google Scholar] [CrossRef]
- Yan, Y.F.; Yang, C.J.; Shang, X.F.; Zhao, Z.M.; Liu, Y.Q.; Zhou, R.; Liu, H.; Wu, T.L.; Zhao, W.B.; Wang, Y.L.; et al. Bioassay-guided isolation of two antifungal compounds from Magnolia officinalis, and the mechanism of action of honokiol. Pestic. Biochem. Physiol. 2020, 170, 104705. [Google Scholar] [CrossRef]
- Guo, H.; Qiao, B.; Ji, X.; Wang, X.; Zhu, E. Antifungal activity and possible mechanisms of submicron chitosan dispersions against Alteraria alternata. Postharvest Biol. Technol. 2020, 161, 110883. [Google Scholar] [CrossRef]
- Zhao, Z.M.; Shang, X.F.; Lawoe, R.K.; Liu, Y.Q.; Zhou, R.; Sun, Y.; Yan, Y.F.; Li, J.C.; Yang, G.Z.; Yang, C.J. Anti-phytopathogenic activity and the possible mechanisms of action of isoquinoline alkaloid sanguinarine. Pestic. Biochem. Physiol. 2019, 159, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene_ID | log2FC | Primer Sequences (5′–3′) | Primer’s Efficiency |
|---|---|---|---|
| acu-6 | 2.03 | F-TGTATACGAAGTCGGTTCAGCC R-TAACCGTCAACGACGTAAATTC | 96 |
| erg10 | −1.73 | F-TCCAACACCCCGCACTACCTGC R-GCTGACCTCGACGGGGACGATC | 95 |
| lys1 | −1.31 | F-GTGAACAGTTCGCCAATGCCTA R-AGAGTGCTCGCGGAGGTGCTG | 97 |
| cut6 | −1.46 | F-AGCAGCCTCCCCAGGGTGAAT R-TCGGTACACTTGTTGAAGAAGTGG | 97 |
| actin | F-CTGGCACCGTCGTCGATGAAG R-AAGGTCCGCTCTCGTCGTACTC | 99 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xun, W.; Gong, B.; Liu, X.; Yang, X.; Zhou, X.; Jin, L. Antifungal Mechanism of Phenazine-1-Carboxylic Acid against Pestalotiopsis kenyana. Int. J. Mol. Sci. 2023, 24, 11274. https://doi.org/10.3390/ijms241411274
Xun W, Gong B, Liu X, Yang X, Zhou X, Jin L. Antifungal Mechanism of Phenazine-1-Carboxylic Acid against Pestalotiopsis kenyana. International Journal of Molecular Sciences. 2023; 24(14):11274. https://doi.org/10.3390/ijms241411274
Chicago/Turabian StyleXun, Weizhi, Bing Gong, Xingxin Liu, Xiuju Yang, Xia Zhou, and Linhong Jin. 2023. "Antifungal Mechanism of Phenazine-1-Carboxylic Acid against Pestalotiopsis kenyana" International Journal of Molecular Sciences 24, no. 14: 11274. https://doi.org/10.3390/ijms241411274
APA StyleXun, W., Gong, B., Liu, X., Yang, X., Zhou, X., & Jin, L. (2023). Antifungal Mechanism of Phenazine-1-Carboxylic Acid against Pestalotiopsis kenyana. International Journal of Molecular Sciences, 24(14), 11274. https://doi.org/10.3390/ijms241411274