
Nitric Oxide Function and Nitric Oxide Synthase Evolution in Aquatic Chordates

 , ,
, ,  ,
,
Abstract
1. Introduction
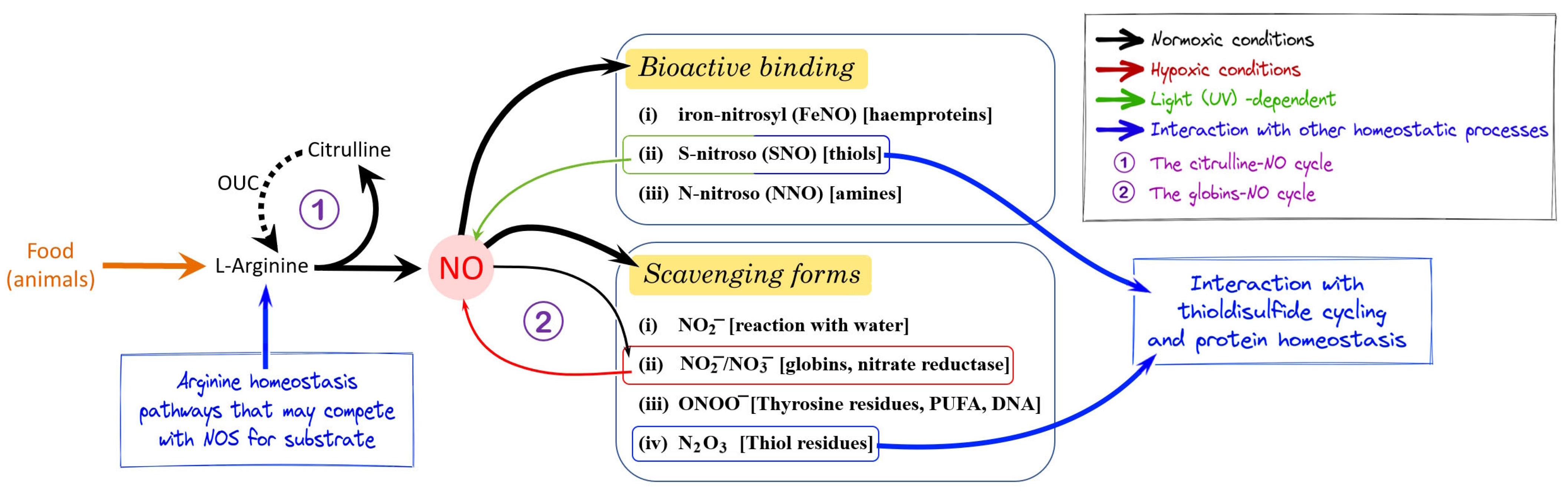
2. NO Homeostasis
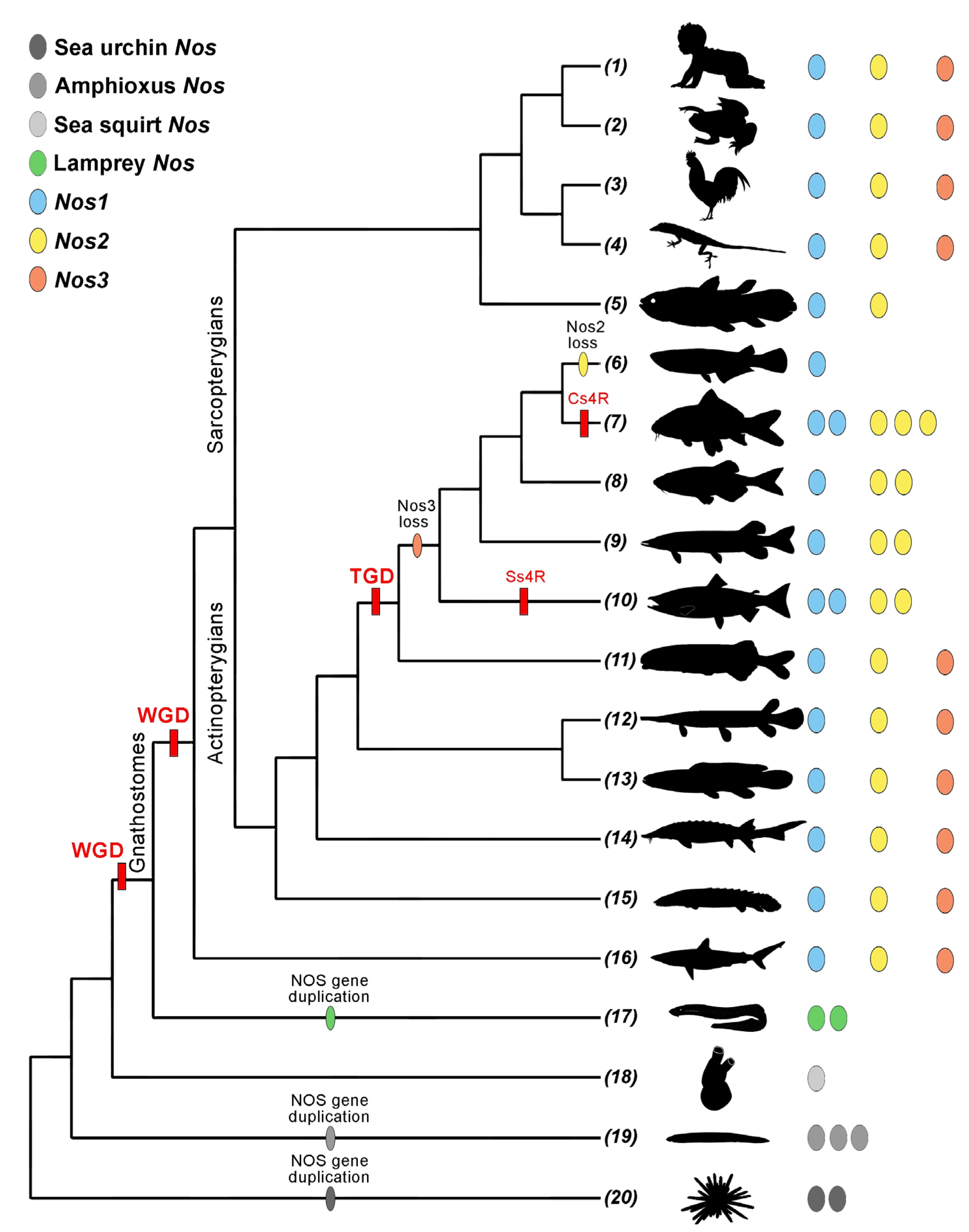
3. Nos Evolution in Deuterostomes
4. NO Function in Cephalochordates
5. NO Function in Tunicates
6. NO Function in Teleosts
6.1. Role of NO in Fish Reproduction and Development
6.2. Role of NO in Teleost Cardiorespiratory Function, Osmoregulation, and Feeding
6.3. Role of NO in the Immune System of Fish
6.4. Role of NO in the Teleost Nervous System, and Control of Locomotion and Behavior
7. NO Function in Amphibia
NO and Metamorphosis in Anura
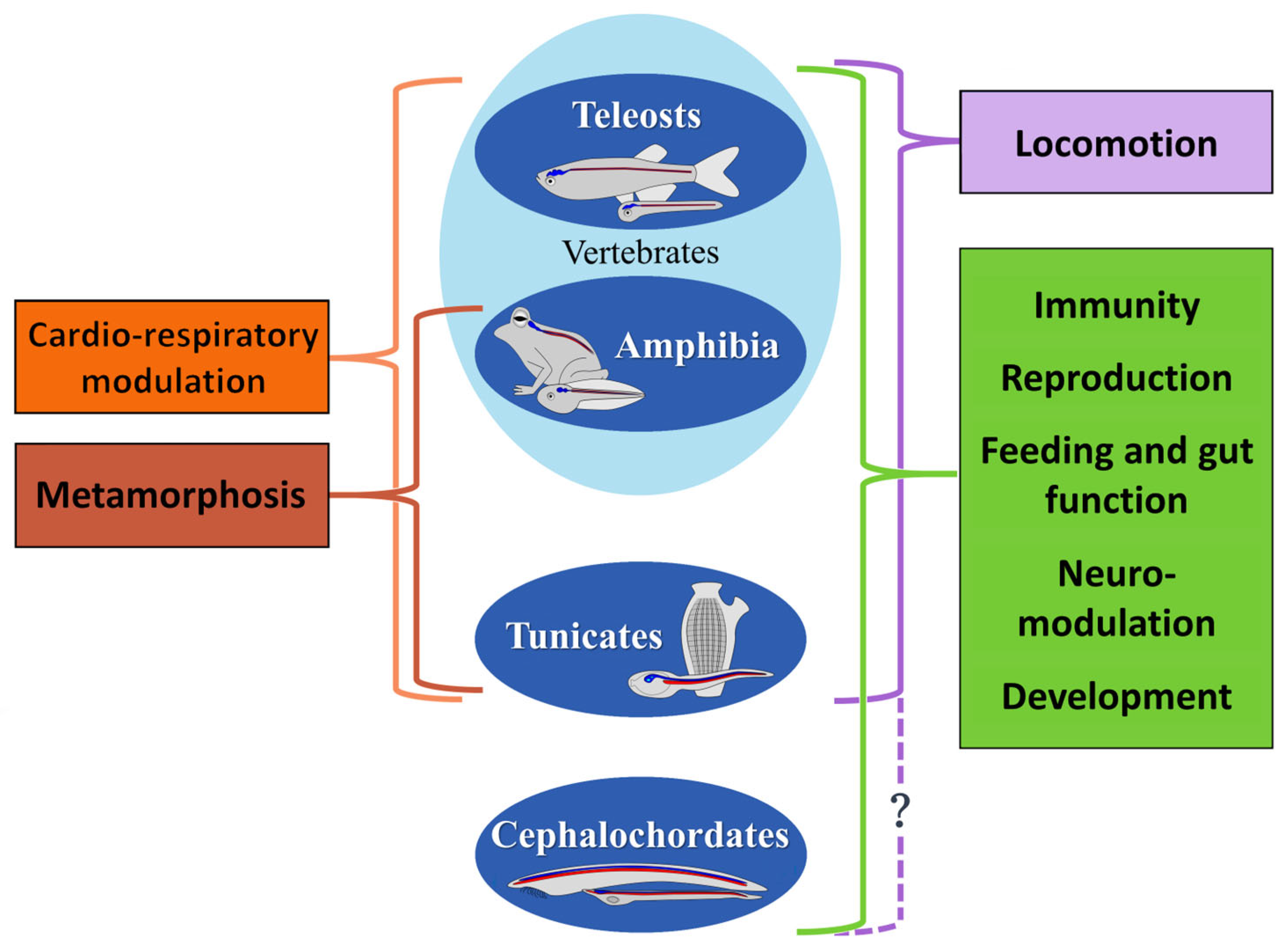
8. Conservation of NO Functions in Aquatic Chordates
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hansen, M.N.; Jensen, F.B. Nitric Oxide Metabolites in Goldfish under Normoxic and Hypoxic Conditions. J. Exp. Biol. 2010, 213, 3593–3602. [Google Scholar] [CrossRef] [PubMed]
- Moroz, L.L.; Romanova, D.Y.; Nikitin, M.A.; Sohn, D.; Kohn, A.B.; Neveu, E.; Varoqueaux, F.; Fasshauer, D. The Diversification and Lineage-Specific Expansion of Nitric Oxide Signaling in Placozoa: Insights in the Evolution of Gaseous Transmission. Sci. Rep. 2020, 10, 13020. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, M.; Giordano, D.; Verde, C.; Poole, R.K. The Evolution of Nitric Oxide Function: From Reactivity in the Prebiotic Earth to Examples of Biological Roles and Therapeutic Applications. Antioxidants 2022, 11, 1222. [Google Scholar] [CrossRef] [PubMed]
- Hirst, D.G.; Robson, T. Nitric Oxide Physiology and Pathology. Methods Mol. Biol. 2011, 704, 1–13. [Google Scholar] [CrossRef]
- Locascio, A.; Vassalli, Q.A.; Castellano, I.; Palumbo, A. Novel Insights on Nitric Oxide Synthase and NO Signaling in Ascidian Metamorphosis. Int. J. Mol. Sci. 2022, 23, 3505. [Google Scholar] [CrossRef]
- Annona, G.; Holland, N.D.; D’Aniello, S. Evolution of the Notochord. Evodevo 2015, 6, 30. [Google Scholar] [CrossRef]
- Gupta, R.S. Molecular Signatures That Are Distinctive Characteristics of the Vertebrates and Chordates and Supporting a Grouping of Vertebrates with the Tunicates. Mol. Phylogenet. Evol. 2016, 94, 383–391. [Google Scholar] [CrossRef]
- McNab, B.K. The Physiological Ecology of Vertebrates: A View from Energetics; Comstock Publishing Associates, Cornell University Press: Ithaca, NY, USA, 2002; 576p. [Google Scholar]
- Gattuso, A.; Garofalo, F.; Cerra, M.C.; Imbrogno, S. Hypoxia Tolerance in Teleosts: Implications of Cardiac Nitrosative Signals. Front. Physiol. 2018, 9, 366. [Google Scholar] [CrossRef]
- Thomas, D.D.; Heinecke, J.L.; Ridnour, L.A.; Cheng, R.Y.; Kesarwala, A.H.; Switzer, C.H.; McVicar, D.W.; Roberts, D.D.; Glynn, S.; Fukuto, J.M.; et al. Signaling and Stress: The Redox Landscape in NOS2 Biology. Free Radic. Biol. Med. 2015, 87, 204–225. [Google Scholar] [CrossRef]
- Potter, L.R. Guanylyl Cyclase Structure, Function and Regulation. Cell Signal. 2011, 23, 1921–1926. [Google Scholar] [CrossRef]
- Mason, M.G.; Nicholls, P.; Wilson, M.T.; Cooper, C.E. Nitric Oxide Inhibition of Respiration Involves Both Competitive (Heme) and Noncompetitive (Copper) Binding to Cytochrome c Oxidase. Proc. Natl. Acad. Sci. USA 2006, 103, 708–713. [Google Scholar] [CrossRef]
- Sarti, P.; Forte, E.; Mastronicola, D.; Giuffrè, A.; Arese, M. Cytochrome c Oxidase and Nitric Oxide in Action: Molecular Mechanisms and Pathophysiological Implications. Biochim. Biophys. Acta 2012, 1817, 610–619. [Google Scholar] [CrossRef]
- Poderoso, J.J.; Helfenberger, K.; Poderoso, C. The Effect of Nitric Oxide on Mitochondrial Respiration. Nitric Oxide 2019, 88, 61–72. [Google Scholar] [CrossRef]
- Rochon, E.R.; Corti, P. Globins and Nitric Oxide Homeostasis in Fish Embryonic Development. Mar. Genomics 2020, 49, 100721. [Google Scholar] [CrossRef]
- Pérez de la Lastra, J.M.; Juan, C.A.; Plou, F.J.; Pérez-Lebeña, E. The Nitration of Proteins, Lipids and DNA by Peroxynitrite Derivatives-Chemistry Involved and Biological Relevance. Stresses 2022, 2, 53–64. [Google Scholar] [CrossRef]
- Xiong, Y.; Uys, J.D.; Tew, K.D.; Townsend, D.M. S-Glutathionylation: From Molecular Mechanisms to Health Outcomes. Antioxid. Redox Signal. 2011, 15, 233. [Google Scholar] [CrossRef]
- Radzinski, M.; Oppenheim, T.; Metanis, N.; Reichmann, D. The Cys Sense: Thiol Redox Switches Mediate Life Cycles of Cellular Proteins. Biomolecules 2021, 11, 469. [Google Scholar] [CrossRef]
- Förstermann, U.; Sessa, W.C. Nitric Oxide Synthases: Regulation and Function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef]
- Allen, A.E.; Dupont, C.L.; Oborník, M.; Horák, A.; Nunes-Nesi, A.; McCrow, J.P.; Zheng, H.; Johnson, D.A.; Hu, H.; Fernie, A.R.; et al. Evolution and Metabolic Significance of the Urea Cycle in Photosynthetic Diatoms. Nature 2011, 473, 203–207. [Google Scholar] [CrossRef]
- Song, X.; Teng, L.; Zhao, X. Re-Examining the Origin and Functions of the Urea Cycle. Res. Sq. 2022. [Google Scholar] [CrossRef]
- Jansson, E.Å.; Huang, L.; Malkey, R.; Govoni, M.; Nihlén, C.; Olsson, A.; Stensdotter, M.; Petersson, J.; Holm, L.; Weitzberg, E.; et al. A Mammalian Functional Nitrate Reductase That Regulates Nitrite and Nitric Oxide Homeostasis. Nat. Chem. Biol. 2008, 4, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.N.; Lundberg, J.O.; Filice, M.; Fago, A.; Christensen, N.M.G.; Jensen, F.B. The Roles of Tissue Nitrate Reductase Activity and Myoglobin in Securing Nitric Oxide Availability in Deeply Hypoxic Crucian Carp. J. Exp. Biol. 2016, 219, 3875–3883. [Google Scholar] [CrossRef] [PubMed]
- Pelegrino, M.T.; Paganotti, A.; Seabra, A.B.; Weller, R.B. Photochemistry of Nitric Oxide and S-Nitrosothiols in Human Skin. Histochem. Cell Biol. 2020, 153, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Casper, I.; Nowag, S.; Koch, K.; Hubrich, T.; Bollmann, F.; Henke, J.; Schmitz, K.; Kleinert, H.; Pautz, A. Post-Transcriptional Regulation of the Human Inducible Nitric Oxide Synthase (INOS) Expression by the Cytosolic Poly (A)-Binding Protein (PABP). Nitric Oxide 2013, 33, 6–17. [Google Scholar] [CrossRef]
- Pautz, A.; Li, H.; Kleinert, H. Regulation of NOS Expression in Vascular Diseases. Front. Biosci. Landmark 2021, 26, 85–101. [Google Scholar] [CrossRef]
- Lehnert, N.; Scheidt, W.R. Preface for the Inorganic Chemistry Forum: The Coordination Chemistry of Nitric Oxide and Its Significance for Metabolism, Signaling, and Toxicity in Biology. Inorg. Chem. 2010, 49, 6223–6225. [Google Scholar] [CrossRef]
- Hill, B.G.; Dranka, B.P.; Bailey, S.M.; Lancaster, J.R.; Darley-Usmar, V.M. What Part of NO Don’t You Understand? Some Answers to the Cardinal Questions in Nitric Oxide Biology. J. Biol. Chem. 2010, 285, 19699–19704. [Google Scholar] [CrossRef]
- Caldwell, R.B.; Toque, H.A.; Narayanan, S.P.; Caldwell, R.W. Arginase: An Old Enzyme with New Tricks. Trends Pharmacol. Sci. 2015, 36, 395–405. [Google Scholar] [CrossRef]
- Caldwell, R.W.; Rodriguez, P.C.; Toque, H.A.; Priya Narayanan, S.; Caldwell, R.B. Arginase: A Multifaceted Enzyme Important in Health and Disease. Physiol. Rev. 2018, 98, 641–665. [Google Scholar] [CrossRef]
- Momma, T.Y.; Ottaviani, J.I. There Is No Direct Competition between Arginase and Nitric Oxide Synthase for the Common Substrate L-Arginine. Nitric Oxide 2022, 129, 16–24. [Google Scholar] [CrossRef]
- Morris, S.M. Enzymes of Arginine Metabolism. J. Nutr. 2004, 134, 2743S–2747S. [Google Scholar] [CrossRef]
- Gambardella, J.; Khondkar, W.; Morelli, M.B.; Wang, X.; Santulli, G.; Trimarco, V. Arginine and Endothelial Function. Biomedicines 2020, 8, 277. [Google Scholar] [CrossRef]
- Martí i Líndez, A.-A.; Reith, W. Arginine-Dependent Immune Responses. Cell. Mol. Life Sci. 2021, 78, 5303–5324. [Google Scholar] [CrossRef]
- Premont, R.T.; Reynolds, J.D.; Zhang, R.; Stamler, J.S. Role of Nitric Oxide Carried by Hemoglobin in Cardiovascular Physiology. Circ. Res. 2020, 126, 129–158. [Google Scholar] [CrossRef]
- Bahadoran, Z.; Mirmiran, P.; Ghasemi, A. Role of Nitric Oxide in Insulin Secretion and Glucose Metabolism. Trends Endocrinol. Metab. 2020, 31, 118–130. [Google Scholar] [CrossRef]
- Bladowski, M.; Gawrys, J.; Gajecki, D.; Szahidewicz-Krupska, E.; Sawicz-Bladowska, A.; Doroszko, A. Role of the Platelets and Nitric Oxide Biotransformation in Ischemic Stroke: A Translative Review from Bench to Bedside. Oxid. Med. Cell. Longev. 2020, 2020, 2979260. [Google Scholar] [CrossRef]
- Tewari, D.; Sah, A.N.; Bawari, S.; Nabavi, S.F.; Dehpour, A.R.; Shirooie, S.; Braidy, N.; Fiebich, B.L.; Vacca, R.A.; Nabavi, S.M. Role of Nitric Oxide in Neurodegeneration: Function, Regulation, and Inhibition. Curr. Neuropharmacol. 2020, 19, 114–126. [Google Scholar] [CrossRef]
- Fukuto, J.M.; Perez-Ternero, C.; Zarenkiewicz, J.; Lin, J.; Hobbs, A.J.; Toscano, J.P. Hydropersulfides (RSSH) and Nitric Oxide (NO) Signaling: Possible Effects on S-Nitrosothiols (RS-NO). Antioxidants 2022, 11, 169. [Google Scholar] [CrossRef]
- Daiber, A.; Kröller-Schön, S.; Oelze, M.; Hahad, O.; Li, H.; Schulz, R.; Steven, S.; Münzel, T. Oxidative Stress and Inflammation Contribute to Traffic Noise-Induced Vascular and Cerebral Dysfunction via Uncoupling of Nitric Oxide Synthases. Redox Biol. 2020, 34, 101506. [Google Scholar] [CrossRef]
- Park, L.; Hochrainer, K.; Hattori, Y.; Ahn, S.J.; Anfray, A.; Wang, G.; Uekawa, K.; Seo, J.; Palfini, V.; Blanco, I.; et al. Tau Induces PSD95–Neuronal NOS Uncoupling and Neurovascular Dysfunction Independent of Neurodegeneration. Nat. Neurosci. 2020, 23, 1079–1089. [Google Scholar] [CrossRef]
- Andreakis, N.; D’Aniello, S.; Albalat, R.; Patti, F.P.; Garcia-Fernandez, J.; Procaccini, G.; Sordino, P.; Palumbo, A. Evolution of the Nitric Oxide Synthase Family in Metazoans. Mol. Biol. Evol. 2011, 28, 163–179. [Google Scholar] [CrossRef] [PubMed]
- Godtman, R.A.; Hallsberg, L.; Löf-Öhlin, Z.; Peeker, R.; Delbro, D. Constitutive Expression of Inducible Nitric Oxide Synthase in Healthy Rat Urothelium? Med. J. Swed. AB 2021, 55, 493–497. [Google Scholar] [CrossRef]
- Stuehr, D.J.; Haque, M.M. Nitric Oxide Synthase Enzymology in the 20 Years after the Nobel Prize. Br. J. Pharmacol. 2019, 176, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Annona, G.; Caccavale, F.; Pascual-Anaya, J.; Kuratani, S.; De Luca, P.; Palumbo, A.; D’Aniello, S. Nitric Oxide Regulates Mouth Development in Amphioxus. Sci. Rep. 2017, 7, 8432. [Google Scholar] [CrossRef]
- Comes, S.; Locascio, A.; Silvestre, F.; d’Ischia, M.; Russo, G.L.; Tosti, E.; Branno, M.; Palumbo, A. Regulatory Roles of Nitric Oxide during Larval Development and Metamorphosis in Ciona Intestinalis. Dev. Biol. 2007, 306, 772–784. [Google Scholar] [CrossRef]
- Yaguchi, J.; Yaguchi, S. Sea Urchin Larvae Utilize Light for Regulating the Pyloric Opening. BMC Biol. 2021, 19, 64. [Google Scholar] [CrossRef]
- Yaguchi, J.; Yaguchi, S. Evolution of Nitric Oxide Regulation of Gut Function. Proc. Natl. Acad. Sci. USA 2019, 116, 5607–5612. [Google Scholar] [CrossRef]
- Caccavale, F.; Annona, G.; Subirana, L.; Escriva, H.; Bertrand, S.; D’Aniello, S. Crosstalk between Nitric Oxide and Retinoic Acid Pathways Is Essential for Amphioxus Pharynx Development. Elife 2021, 10, e58295. [Google Scholar] [CrossRef]
- Chen, D.; Lin, Y.; Zhang, H. Characterization and Expression of Two Amphioxus DDAH Genes Originating from an Amphioxus-Specific Gene Duplication. Gene 2008, 410, 75–81. [Google Scholar] [CrossRef]
- Lin, Y.S.; Chen, D.Y.; Zhang, W.; Cai, Z.P.; Chen, Z.K.; Zhang, N.; Mao, B.Y.; Zhang, H.W. Characterization of the Immune Defense Related Tissues, Cells, and Genes in Amphioxus. Sci. China Life Sci. 2011, 54, 999–1004. [Google Scholar] [CrossRef][Green Version]
- Caccavale, F.; Coppola, U.; Vassalli, Q.A.; La Vecchia, C.; Palumbo, A.; D’Aniello, E.; Locascio, A.; Ristoratore, F.; D’Aniello, S. Transphyletic Conservation of Nitric Oxide Synthase Regulation in Cephalochordates and Tunicates. Dev. Genes Evol. 2020, 230, 329–338. [Google Scholar] [CrossRef]
- Donald, J.A.; Forgan, L.G.; Cameron, M.S. The Evolution of Nitric Oxide Signalling in Vertebrate Blood Vessels. J. Comp. Physiol. B 2015, 185, 153–171. [Google Scholar] [CrossRef]
- Annona, G.; Sato, I.; Pascual-Anaya, J.; Osca, D.; Braasch, I.; Voss, R.; Stundl, J.; Soukup, V.; Ferrara, A.; Fontenot, Q.; et al. Evolution of the Nitric Oxide Synthase Family in Vertebrates and Novel Insights in Gill Development. Proc. R. Soc. B Biol. Sci. 2022, 289, 667. [Google Scholar] [CrossRef]
- Annona, G.; Ferran, J.L.; De Luca, P.; Conte, I.; Postlethwait, J.H.; D’Aniello, S. Expression Pattern of Nos1 in the Developing Nervous System of Ray-Finned Fish. Genes 2022, 13, 918. [Google Scholar] [CrossRef]
- Dehal, P.; Boore, J.L. Two Rounds of Whole Genome Duplication in the Ancestral Vertebrate. PLoS Biol. 2005, 3, e314. [Google Scholar] [CrossRef]
- Nakatani, Y.; Shingate, P.; Ravi, V.; Pillai, N.E.; Prasad, A.; McLysaght, A.; Venkatesh, B. Reconstruction of Proto-Vertebrate, Proto-Cyclostome and Proto-Gnathostome Genomes Provides New Insights into Early Vertebrate Evolution. Nat. Commun. 2021, 12, 4489. [Google Scholar] [CrossRef]
- Pasquier, J.; Braasch, I.; Batzel, P.; Cabau, C.; Montfort, J.; Nguyen, T.; Jouanno, E.; Berthelot, C.; Klopp, C.; Journot, L.; et al. Evolution of Gene Expression after Whole-Genome Duplication: New Insights from the Spotted Gar Genome. J. Exp. Zool. B Mol. Dev. Evol. 2017, 328, 709–721. [Google Scholar] [CrossRef]
- Godoy, L.; Gonzàlez-Duarte, R.; Albalat, R. S-Nitrosogluthathione Reductase Activity of Amphioxus ADH3: Insights into the Nitric Oxide Metabolism. Int. J. Biol. Sci. 2006, 2, 117. [Google Scholar] [CrossRef]
- Colasanti, M.; Venturini, G.; Merante, A.; Musci, G.; Lauro, G.M. Nitric Oxide Involvement in Hydra Vulgaris Very Primitive Olfactory-Like System. J. Neurosci. 1997, 17, 493–499. [Google Scholar] [CrossRef]
- Colasanti, M.; Venturini, G. Nitric Oxide in Invertebrates. Mol. Neurobiol. 1998, 17, 157–174. [Google Scholar] [CrossRef]
- Moroz, L.L.; Norekian, T.P.; Pirtle, T.J.; Robertson, K.J.; Satterlie, R.A. Distribution of NADPH-Diaphorase Reactivity and Effects of Nitric Oxide on Feeding and Locomotory Circuitry in the Pteropod Mollusc, Clione Limacina. J. Comp. Neurol. 2000, 427, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.; Sadamoto, H.; Ogawa, H.; Kitamura, Y.; Oka, K.; Tanishita, K.; Ito, E. Nitric Oxide Generation around Buccal Ganglia Accompanying Feeding Behavior in the Pond Snail, Lymnaea Stagnalis. Neurosci. Res. 2000, 38, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.I.; Tachibana, T.; Hasebe, Y.; Masuda, N.; Ueda, H. Peripheral or Central Administration of Nitric Oxide Synthase Inhibitor Affects Feeding Behavior in Chicks. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 148, 458–462. [Google Scholar] [CrossRef] [PubMed]
- Susswein, A.J.; Chiel, H.J. Nitric Oxide as a Regulator of Behavior: New Ideas from Aplysia Feeding. Prog. Neurobiol. 2012, 97, 304–317. [Google Scholar] [CrossRef]
- Hazut, N.; Rapps, K.; Kristt, D.A.; Susswein, A.J.; Weller, A. Nitric Oxide and L-Arginine Regulate Feeding in Satiated Rats. Appetite 2019, 132, 44–54. [Google Scholar] [CrossRef]
- Zhang, S.; Li, L.; Li, H.; Guo, H. Histochemical Localization of Constitutive Nitric Oxide Synthases in Amphioxus Branchiostoma Belcheri Tsingtauense. J. Mar. Biol. Assoc. UK 2002, 82, 1041–1042. [Google Scholar] [CrossRef]
- Yongjun, W.; Shicui, Z. Localization of Nitric Oxide Synthase in the Developing Gonads of Amphioxus Branchiostoma Belcheri Tsingtauense. Acta Oceanol. Sin. 2005, 24, 120–126. [Google Scholar]
- Ueda, N.; Degnan, S.M. Nitric Oxide Acts as a Positive Regulator to Induce Metamorphosis of the Ascidian Herdmania Momus. PLoS ONE 2013, 8, e72797. [Google Scholar] [CrossRef]
- Heyland, A.; Moroz, L.L. Signaling Mechanisms Underlying Metamorphic Transitions in Animals. Integr. Comp. Biol. 2006, 46, 743–759. [Google Scholar] [CrossRef]
- Paris, M.; Laudet, V. The History of a Developmental Stage: Metamorphosis in Chordates. Genesis 2008, 46, 657–672. [Google Scholar] [CrossRef]
- Castellano, I.; Ercolesi, E.; Palumbo, A. Nitric Oxide Affects ERK Signaling through Down-Regulation of MAP Kinase Phosphatase Levels during Larval Development of the Ascidian Ciona Intestinalis. PLoS ONE 2014, 9, e102907. [Google Scholar] [CrossRef]
- Bishop, C.D.; Bates, W.R.; Brandhorst, B.P. Regulation of Metamorphosis in Ascidians Involves NO/CGMP Signaling and HSP90. J. Exp. Zool. 2001, 289, 374–384. [Google Scholar] [CrossRef]
- Ercolesi, E.; Tedeschi, G.; Fiore, G.; Negri, A.; Maffioli, E.; D’Ischia, M.; Palumbo, A. Protein Nitration as Footprint of Oxidative Stress-Related Nitric Oxide Signaling Pathways in Developing Ciona Intestinalis. Nitric Oxide 2012, 27, 18–24. [Google Scholar] [CrossRef]
- Zaitseva, O.V.; Romanov, V.N.; Markosova, T.G. Distribution of NADPH-Diaphorase Activity in Organs and Tissues of Adult Ascidians. Dokl. Biol. Sci. 2012, 444, 180–183. [Google Scholar] [CrossRef]
- de Barros, C.M.; De Carvalho, D.R.; Andrade, L.R.; Pavão, M.S.G.; Allodi, S. Nitric Oxide Production by Hemocytes of the Ascidian Styela Plicata. Cell Tissue Res. 2009, 338, 117–128. [Google Scholar] [CrossRef]
- de Barros, C.M.; de Abreu Mello, A.; Allodi, S. Norepinephrine Depresses the Nitric Oxide Production in the Ascidian Hemocytes. J. Invertebr. Pathol. 2012, 111, 182–185. [Google Scholar] [CrossRef][Green Version]
- Cioni, C.; Angiulli, E.; Toni, M. Nitric Oxide and the Neuroendocrine Control of the Osmotic Stress Response in Teleosts. Int. J. Mol. Sci. 2019, 20, 489. [Google Scholar] [CrossRef]
- Peter, M.C.S.; Gayathry, R.; Peter, V.S. Inducible Nitric Oxide Synthase/Nitric Oxide System as a Biomarker for Stress and Ease Response in Fish: Implication on Na+ Homeostasis During Hypoxia. Front. Physiol. 2022, 13, 795. [Google Scholar] [CrossRef]
- Filice, M.; Imbrogno, S.; Gattuso, A.; Cerra, M.C. Hypoxic and Thermal Stress: Many Ways Leading to the NOS/NO System in the Fish Heart. Antioxidants 2021, 10, 1401. [Google Scholar] [CrossRef]
- Li, J.; Zhou, W.; Wang, Y.; Niu, C. The Dual Role of CGMP in Oocyte Maturation of Zebrafish. Biochem. Biophys. Res. Commun. 2018, 499, 998–1003. [Google Scholar] [CrossRef]
- Stricker, S.A. Comparative Biology of Calcium Signaling during Fertilization and Egg Activation in Animals. Dev. Biol. 1999, 211, 157–176. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript Assembly and Quantification by RNA-Seq Reveals Unannotated Transcripts and Isoform Switching during Cell Differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Corti, P.; Xue, J.; Tejero, J.; Wajih, N.; Sun, M.; Stolz, D.B.; Tsang, M.; Kim-Shapiro, D.B.; Gladwin, M.T. Globin X Is a Six-Coordinate Globin That Reduces Nitrite to Nitric Oxide in Fish Red Blood Cells. Proc. Natl. Acad. Sci. USA 2016, 113, 8538–8543. [Google Scholar] [CrossRef]
- Lepiller, S.; Laurens, V.; Bouchot, A.; Herbomel, P.; Solary, E.; Chluba, J. Imaging of Nitric Oxide in a Living Vertebrate Using a Diaminofluorescein Probe. Free Radic. Biol. Med. 2007, 43, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Fritsche, R.; Schwerte, T.; Pelster, B. Nitric Oxide and Vascular Reactivity in Developing Zebrafish, Danio Rerio. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R2200–R2207. [Google Scholar] [CrossRef]
- Pelster, B.; Grillitsch, S.; Schwerte, T. NO as a Mediator during the Early Development of the Cardiovascular System in the Zebrafish. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2005, 142, 215–220. [Google Scholar] [CrossRef]
- North, T.E.; Goessling, W.; Peeters, M.; Li, P.; Ceol, C.; Lord, A.M.; Weber, G.J.; Harris, J.; Cutting, C.C.; Huang, P.; et al. Hematopoietic Stem Cell Development Is Dependent on Blood Flow. Cell 2009, 137, 736–748. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, P.; Wei, Y.; Gao, Y.; Patient, R.; Liu, F. A Blood Flow–Dependent Klf2a-NO Signaling Cascade Is Required for Stabilization of Hematopoietic Stem Cell Programming in Zebrafish Embryos. Blood 2011, 118, 4102–4110. [Google Scholar] [CrossRef]
- Veerkamp, J.; Rudolph, F.; Cseresnyes, Z.; Priller, F.; Otten, C.; Renz, M.; Schaefer, L.; Abdelilah-Seyfried, S. Unilateral Dampening of Bmp Activity by Nodal Generates Cardiac Left-Right Asymmetry. Dev. Cell 2013, 24, 660–667. [Google Scholar] [CrossRef]
- Holmqvist, B.; Ellingsen, B.; Forsell, J.; Zhdanova, I.; Alm, P. The Early Ontogeny of Neuronal Nitric Oxide Synthase Systems in the Zebrafish. J. Exp. Biol. 2004, 207, 923–935. [Google Scholar] [CrossRef]
- Bradley, S.; Tossell, K.; Lockley, R.; McDearmid, J.R. Nitric Oxide Synthase Regulates Morphogenesis of Zebrafish Spinal Cord Motoneurons. J. Neurosci. 2010, 30, 16818. [Google Scholar] [CrossRef]
- Kong, Y.; Grimaldi, M.; Curtin, E.; Dougherty, M.; Kaufman, C.; White, R.M.; Zon, L.I.; Liao, E.C. Neural Crest Development and Craniofacial Morphogenesis Is Coordinated by Nitric Oxide and Histone Acetylation. Chem. Biol. 2014, 21, 488–501. [Google Scholar] [CrossRef]
- Harumi, T.; Watanabe, T.; Yamamoto, T.; Tanabe, Y.; Suzuki, N. Expression of Membrane-Bound and Soluble Guanylyl Cyclase MRNAs in Embryonic and Adult Retina of the Medaka Fish Oryzias Latipes. Zool. Sci. 2003, 20, 133–140. [Google Scholar] [CrossRef][Green Version]
- Giordano, D.; Verde, C.; Corti, P. Nitric Oxide Production and Regulation in the Teleost Cardiovascular System. Antioxidants 2022, 11, 957. [Google Scholar] [CrossRef]
- Imbrogno, S.; Verri, T.; Filice, M.; Barca, A.; Schiavone, R.; Gattuso, A.; Cerra, M.C. Shaping the Cardiac Response to Hypoxia: NO and Its Partners in Teleost Fish. Curr. Res. Physiol. 2022, 5, 193–202. [Google Scholar] [CrossRef]
- Imbrogno, S.; Tota, B.; Gattuso, A. The Evolutionary Functions of Cardiac NOS/NO in Vertebrates Tracked by Fish and Amphibian Paradigms. Nitric Oxide 2011, 25, 1–10. [Google Scholar] [CrossRef]
- Newton, C.M.; Stoyek, M.R.; Croll, R.P.; Smith, F.M. Regional Innervation of the Heart in the Goldfish, Carassius Auratus: A Confocal Microscopy Study. J. Comp. Neurol. 2014, 522, 456–478. [Google Scholar] [CrossRef]
- Agnisola, C. Role of Nitric Oxide in the Control of Coronary Resistance in Teleosts. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2005, 142, 178–187. [Google Scholar] [CrossRef]
- Agnisola, C.; Pellegrino, D. Role of Nitric Oxide in Vascular Regulation in Fish. Adv Exp Biol 2007, 1, 293–310. [Google Scholar] [CrossRef]
- Olson, K.R.; Donald, J.A. Nervous Control of Circulation—The Role of Gasotransmitters, NO, CO, and H2S. Acta Histochem. 2009, 111, 244–256. [Google Scholar] [CrossRef]
- Imanishi, Y.; Yang, L.; Sokal, I.; Filipek, S.; Palczewski, K.; Baehr, W. Diversity of Guanylate Cyclase-Activating Proteins (GCAPs) in Teleost Fish: Characterization of Three Novel GCAPs (GCAP4, GCAP5, GCAP7) from Zebrafish (Danio Rerio) and Prediction of Eight GCAPs (GCAP1-8) in Pufferfish (Fugu rubripes). J. Mol. Evol. 2004, 59, 204–217. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Saraiva, L.R.; Ahuja, G.; Ivandic, I.; Syed, A.S.; Marioni, J.C.; Korsching, S.I.; Logan, D.W. Molecular and Neuronal Homology between the Olfactory Systems of Zebrafish and Mouse. Sci. Rep. 2015, 5, 11487. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Yao, Y.; Harumi, T.; Suzuki, N. Localization of the Nitric Oxide/CGMP Signaling Pathway-Related Genes and Influences of Morpholino Knock-down of Soluble Guanylyl Cyclase on Medaka Fish Embryogenesis. Zool. Sci. 2003, 20, 181–191. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mikami, T.; Kusakabe, T.; Suzuki, N. Tandem Organization of Medaka Fish Soluble Guanylyl Cyclase A1 and Β1 Subunit Genes. J. Biol. Chem. 1999, 274, 18567–18573. [Google Scholar] [CrossRef]
- Imbrogno, S.; Capria, C.; Tota, B.; Jensen, F.B. Nitric Oxide Improves the Hemodynamic Performance of the Hypoxic Goldfish (Carassius auratus) Heart. Nitric Oxide 2014, 42, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Vishnolia, K.K.; Rakovic, A.; Hoene, C.; Tarhbalouti, K.; Aherrahrou, Z.; Erdmann, J. SGC Activity and Regulation of Blood Flow in a Zebrafish Model System. Front. Physiol. 2021, 12, 217. [Google Scholar] [CrossRef]
- Xiyuan, Z.; Fink, R.H.A.; Mosqueira, M. NO-SGC Pathway Modulates Ca2+ Release and Muscle Contraction in Zebrafish Skeletal Muscle. Front. Physiol. 2017, 8, 607. [Google Scholar] [CrossRef]
- Pellegrino, D.; Sprovieri, E.; Mazza, R.; Randall, D.J.; Tota, B. Nitric Oxide-CGMP-Mediated Vasoconstriction and Effects of Acetylcholine in the Branchial Circulation of the Eel. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2002, 132, 447–457. [Google Scholar] [CrossRef]
- Hyndman, K.A.; Choe, K.P.; Havird, J.C.; Rose, R.E.; Piermarini, P.M.; Evans, D.H. Neuronal Nitric Oxide Synthase in the Gill of the Killifish, Fundulus Heteroclitus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2006, 144, 510–519. [Google Scholar] [CrossRef]
- Mauceri, A.; Fasulo, S.; Ainis, L.; Licata, A.; Rita Lauriano, E.; Martfnez, A.; Mayer, B.; Zaccone, G. Neuronal Nitric Oxide Synthase (NNOS) Expression in the Epithelial Neuroendocrine Cell System and Nerve Fibers in the Gill of the Catfish, Heteropneustes Fossilis. Acta Histochem. 1999, 101, 437–448. [Google Scholar] [CrossRef]
- Gibbins, I.L.; Olsson, C.; Holmgren, S. Distribution of Neurons Reactive for NADPH-Diaphorase in the Branchial Nerves of a Teleost Fish, Gadus Morhua. Neurosci. Lett. 1995, 193, 113–116. [Google Scholar] [CrossRef]
- Zaccone, G.; Capillo, G.; Aragona, M.; Alesci, A.; Cupello, C.; Lauriano, E.R.; Guerrera, M.C.; Kuciel, M.; Zuwala, K.; Germana, A.; et al. Gill Structure and Neurochemical Markers in the African Bonytongue (Heterotis niloticus): A Preliminary Study. Acta Histochem. 2022, 124, 151954. [Google Scholar] [CrossRef]
- Garofalo, F.; Santovito, G.; Amelio, D. Morpho-Functional Effects of Heat Stress on the Gills of Antarctic T. bernacchii and C. hamatus. Mar. Pollut. Bull. 2019, 141, 194–204. [Google Scholar] [CrossRef]
- Tipsmark, C.K. Regulation of Na+/K+-ATPase Activity by Nitric Oxide in the Kidney and Gill of the Brown Trout (Salmo trutta). J. Exp. Biol. 2003, 206, 1503–1510. [Google Scholar] [CrossRef]
- Ebbesson, L.O.E. Nitric Oxide Synthase in the Gill of Atlantic Salmon: Colocalization with and Inhibition of Na+,K+-ATPase. J. Exp. Biol. 2005, 208, 1011–1017. [Google Scholar] [CrossRef]
- Jiménez, A.; Esteban, F.J.; Sánchez-López, A.M.; Pedrosa, J.A.; Del Moral, M.L.; Hernández, R.; Blanco, S.; Barroso, J.B.; Rodrigo, J.; Peinado, M.A. Immunohistochemical Localisation of Neuronal Nitric Oxide Synthase in the Rainbow Trout Kidney. J. Chem. Neuroanat. 2001, 21, 289–294. [Google Scholar] [CrossRef]
- Trischitta, F.; Pidalà, P.; Faggio, C. Nitric Oxide Modulates Ionic Transport in the Isolated Intestine of the Eel, Anguilla Anguilla. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 148, 368–373. [Google Scholar] [CrossRef]
- Peter, V.S. Nitric Oxide Rectifies Acid-Base Disturbance and Modifies Thyroid Hormone Activity during Net Confinement of Air-Breathing Fish (Anabas testudineus Bloch). Gen. Comp. Endocrinol. 2013, 181, 115–121. [Google Scholar] [CrossRef]
- Gerber, L.; Madsen, S.S.; Jensen, F.B. Cortisol Regulates Nitric Oxide Synthase in Freshwater and Seawater Acclimated Rainbow Trout, Oncorhynchus mykiss. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2017, 204, 1–8. [Google Scholar] [CrossRef]
- Gerber, L.; Jensen, F.B.; Madsen, S.S. Dynamic Changes in Nitric Oxide Synthase Expression Are Involved in Seawater Acclimation of Rainbow Trout Oncorhynchus mykiss. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 314, R552–R562. [Google Scholar] [CrossRef]
- Porteus, C.S.; Pollack, J.; Tzaneva, V.; Kwong, R.W.M.; Kumai, Y.; Abdallah, S.J.; Zaccone, G.; Lauriano, E.R.; Milsom, W.K.; Perry, S.F. A Role for Nitric Oxide in the Control of Breathing in Zebrafish (Danio rerio). J. Exp. Biol. 2015, 218, 3746–3753. [Google Scholar] [CrossRef] [PubMed]
- Tota, B.; Amelio, D.; Cerra, M.C.; Garofalo, F. The Morphological and Functional Significance of the NOS/NO System in the Respiratory, Osmoregulatory, and Contractile Organs of the African Lungfish. Acta Histochem 2018, 120, 654–666. [Google Scholar] [CrossRef] [PubMed]
- Olsson, C.; Holmgren, S. The Control of Gut Motility. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2001, 128, 479–501. [Google Scholar] [CrossRef]
- Olsson, C.; Holmgren, S. Autonomic Control of Gut Motility: A Comparative View. Auton. Neurosci. 2011, 165, 80–101. [Google Scholar] [CrossRef]
- Ceccotti, C.; Giaroni, C.; Bistoletti, M.; Viola, M.; Crema, F.; Terova, G. Neurochemical Characterization of Myenteric Neurons in the Juvenile Gilthead Sea Bream (Sparus aurata) Intestine. PLoS ONE 2018, 13, e0201760. [Google Scholar] [CrossRef]
- Olsson, C.; Holmgren, S. Pacap and Nitric Oxide Inhibit Contractions in the Proximal Intestine of the Atlantic Cod, Gadus Morhua. J. Exp. Biol. 2000, 203, 575–583. [Google Scholar] [CrossRef]
- Holmberg, A.; Olsson, C.; Holmgren, S. The Effects of Endogenous and Exogenous Nitric Oxide on Gut Motility in Zebrafish Danio Rerio Embryos and Larvae. J. Exp. Biol. 2006, 209, 2472–2479. [Google Scholar] [CrossRef]
- Olsson, C.; Aldman, G.; Larsson, A.; Holmgren, S. Cholecystokinin Affects Gastric Emptying and Stomach Motility in the Rainbow Trout Oncorhynchus Mykiss. J. Exp. Biol. 1999, 202, 161–170. [Google Scholar] [CrossRef]
- Uyttebroek, L.; Shepherd, I.T.; Vanden Berghe, P.; Hubens, G.; Timmermans, J.P.; Van Nassauw, L. The Zebrafish Mutant Lessen: An Experimental Model for Congenital Enteric Neuropathies. Neurogastroenterol. Motil. 2016, 28, 345–357. [Google Scholar] [CrossRef]
- Bosi, G.; Maynard, B.J.; Pironi, F.; Dezfuli, B.S. Parasites and the Neuroendocrine Control of Fish Intestinal Function: An Ancient Struggle between Pathogens and Host. Parasitology 2022, 149, 1842–1861. [Google Scholar] [CrossRef]
- Zhu, W.; Su, J. Immune Functions of Phagocytic Blood Cells in Teleost. Rev. Aquac. 2022, 14, 630–646. [Google Scholar] [CrossRef]
- Mosser, D.M.; Edwards, J.P. Exploring the Full Spectrum of Macrophage Activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef]
- Denicola, A.; Rubbo, H.; Rodriguez, D.; Radi, R. Peroxynitrite-Mediated Cytotoxicity to Trypanosoma Cruzi. Arch. Biochem. Biophys. 1993, 304, 279–286. [Google Scholar] [CrossRef]
- Henard, C.A.; Vázquez-Torres, A. Nitric Oxide and Salmonella Pathogenesis. Front. Microbiol. 2011, 2, 84. [Google Scholar] [CrossRef]
- Wiegertjes, G.F.; Wentzel, A.S.; Spaink, H.P.; Elks, P.M.; Fink, I.R. Polarization of Immune Responses in Fish: The ‘Macrophages First’ Point of View. Mol. Immunol. 2016, 69, 146–156. [Google Scholar] [CrossRef]
- Neumann, N.F.; Fagan, D.; Belosevic, M. Macrophage Activating Factor(s) Secreted by Mitogen Stimulated Goldfish Kidney Leukocytes Synergize with Bacterial Lipopolysaccharide to Induce Nitric Oxide Production in Teleost Macrophages. Dev. Comp. Immunol. 1995, 19, 473–482. [Google Scholar] [CrossRef]
- Buentello, J.A.; Gatlin, D.M. Nitric Oxide Production in Activated Macrophages from Channel Catfish (Ictalurus Punctatus): Influence of Dietary Arginine and Culture Media. Aquaculture 1999, 179, 513–521. [Google Scholar] [CrossRef]
- Losada, A.P.; Bermúdez, R.; Faílde, L.D.; Quiroga, M.I. Quantitative and Qualitative Evaluation of INOS Expression in Turbot (Psetta maxima) Infected with Enteromyxum scophthalmi. Fish. Shellfish Immunol. 2012, 32, 243–248. [Google Scholar] [CrossRef]
- Saeij, J.P.J.; Stet, R.J.M.; Groeneveld, A.; Verburg-Van Kemenade, L.B.M.; Van Muiswinkel, W.B.; Wiegertjes, G.F. Molecular and Functional Characterization of a Fish Inducible-Type Nitric Oxide Synthase. Immunogenetics 2000, 51, 339–346. [Google Scholar] [CrossRef]
- Vojtech, L.N.; Sanders, G.E.; Conway, C.; Ostland, V.; Hansen, J.D. Host Immune Response and Acute Disease in a Zebrafish Model of Francisella Pathogenesis. Infect. Immun. 2009, 77, 914–925. [Google Scholar] [CrossRef]
- Campos-Perez, J.J.; Ward, M.; Grabowski, P.S.; Ellis, A.E.; Secombes, C.J. The Gills Are an Important Site of INOS Expression in Rainbow Trout Oncorhynchus Mykiss after Challenge with the Gram-Positive Pathogen Renibacterium salmoninarum. Immunology 2000, 99, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Forlenza, M.; Fink, I.R.; Raes, G.; Wiegertjes, G.F. Heterogeneity of Macrophage Activation in Fish. Dev. Comp. Immunol. 2011, 35, 1246–1255. [Google Scholar] [CrossRef] [PubMed]
- Wentzel, A.S.; Janssen, J.J.E.; de Boer, V.C.J.; van Veen, W.G.; Forlenza, M.; Wiegertjes, G.F. Fish Macrophages Show Distinct Metabolic Signatures Upon Polarization. Front. Immunol. 2020, 11, 152. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, V.; Mancuso, C.; Calvani, M.; Rizzarelli, E.; Butterfield, D.A.; Giuffrida Stella, A.M. Nitric Oxide in the Central Nervous System: Neuroprotection versus Neurotoxicity. Nat. Rev. Neurosci. 2007, 8, 766–775. [Google Scholar] [CrossRef]
- Øyan, A.M.; Nilsen, F.; Goksøyr, A.; Holmqvist, B. Partial cloning of constitutive and inducible nitric oxide synthases and detailed neuronal expression of NOS mRNA in the cerebellum and optic tectum of adult Atlantic salmon (Salmo salar). Mol. Brain Res. 2000, 78, 38–49. [Google Scholar] [CrossRef]
- Zhou, L.; Bai, R.; Tian, J.; Liu, X.; Lu, D.; Zhu, P.; Liu, Y.; Zeng, L.; Luo, W.; Zhang, Y.; et al. Bioinformatic comparisons and tissue expression of the neuronal nitric oxide synthase (nNOS) gene from the red drum (Sciaenops ocellatus). Fish Shellfish. Immunol. 2009, 27, 577–584. [Google Scholar] [CrossRef]
- Brüning, G.; Hattwig, K.; Mayer, B. Nitric Oxide Synthase in the Peripheral Nervous System of the Goldfish, Carassius Auratus. Cell Tissue Res. 1996, 284, 87–98. [Google Scholar] [CrossRef]
- Zaccone, G.; Lauriano, E.R.; Kuciel, M.; Capillo, G.; Pergolizzi, S.; Alesci, A.; Ishimatsu, A.; Ip, Y.K.; Icardo, J.M. Identification and Distribution of Neuronal Nitric Oxide Synthase and Neurochemical Markers in the Neuroepithelial Cells of the Gill and the Skin in the Giant Mudskipper, Periophthalmodon schlosseri. Zoology 2017, 125, 41–52. [Google Scholar] [CrossRef]
- Penglee, R.; Gao, L.; Huang, Y.; Liu, L.; Nimitkul, S.; Bao, B. The Role of Nitric Oxide and Neuronal Nitric Oxide Synthase in Zebrafish (Danio rerio) Shoaling. Aquac. Fish 2021, 6, 565–573. [Google Scholar] [CrossRef]
- Carreño Gutiérrez, H.; O’Leary, A.; Freudenberg, F.; Fedele, G.; Wilkinson, R.; Markham, E.; van Eeden, F.; Reif, A.; Norton, W.H.J. Nitric Oxide Interacts with Monoamine Oxidase to Modulate Aggression and Anxiety-like Behaviour. Eur. Neuropsychopharmacol. 2020, 30, 30–43. [Google Scholar] [CrossRef]
- Filby, A.L.; Paull, G.C.; Hickmore, T.F.A.; Tyler, C.R. Unravelling the Neurophysiological Basis of Aggression in a Fish Model. BMC Genom. 2010, 11, 498. [Google Scholar] [CrossRef]
- Akinrinade, I.; Kareklas, K.; Teles, M.C.; Reis, T.K.; Gliksberg, M.; Petri, G.; Levkowitz, G.; Oliveira, R.F. Evolutionarily Conserved Role of Oxytocin in Social Fear Contagion in Zebrafish. Science 2023, 379, 1232–1237. [Google Scholar] [CrossRef]
- Rettori, V.; Canteros, G.; Renoso, R.; Gimeno, M.; McCann, S.M. Oxytocin Stimulates the Release of Luteinizing Hormone-Releasing Hormone from Medial Basal Hypothalamic Explants by Releasing Nitric Oxide. Proc. Natl. Acad. Sci. USA 1997, 94, 2741–2744. [Google Scholar] [CrossRef]
- Bordieri, L.; Bonaccorsi Di Patti, M.C.; Miele, R.; Cioni, C. Partial Cloning of Neuronal Nitric Oxide Synthase (NNOS) CDNA and Regional Distribution of NNOS MRNA in the Central Nervous System of the Nile Tilapia Oreochromis Niloticus. Mol. Brain Res. 2005, 142, 123–133. [Google Scholar] [CrossRef]
- Mobley, R.B.; Ray, E.J.; Maruska, K.P. Expression and Localization of Neuronal Nitric Oxide Synthase in the Brain and Sensory Tissues of the African Cichlid Fish Astatotilapia burtoni. J. Comp. Neurol. 2022, 530, 2901–2917. [Google Scholar] [CrossRef]
- Vielma, A.H.; Retamal, M.A.; Schmachtenberg, O. Nitric Oxide Signaling in the Retina: What Have We Learned in Two Decades? Brain Res. 2012, 1430, 112–125. [Google Scholar] [CrossRef]
- Singru, P.S.; Sakharkar, A.J.; Subhedar, N. Neuronal Nitric Oxide Synthase in the Olfactory System of an Adult Teleost Fish Oreochromis mossambicus. Brain Res. 2003, 977, 157–168. [Google Scholar] [CrossRef]
- Liepe, B.A.; Stone, C.; Koistinaho, J.; Copenhagen, D.R. Nitric Oxide Synthase in Muller Cells and Neurons of Salamander and Fish Retina. J. Neurosci. 1994, 14, 7641–7654. [Google Scholar] [CrossRef]
- Miles, G.B.; Sillar, K.T. Neuromodulation of Vertebrate Locomotor Control Networks. Physiology 2011, 26, 393–411. [Google Scholar] [CrossRef]
- Pirtle, T.J.; Satterlie, R.A. Cyclic Guanosine Monophosphate Modulates Locomotor Acceleration Induced by Nitric Oxide but Not Serotonin in Clione limacina Central Pattern Generator Swim Interneurons. Integr. Org. Biol. 2021, 3, obaa045. [Google Scholar] [CrossRef]
- Hachoumi, L.; Sillar, K.T. Developmental Stage-Dependent Switching in the Neuromodulation of Vertebrate Locomotor Central Pattern Generator Networks. Dev. Neurobiol. 2020, 80, 42–57. [Google Scholar] [CrossRef] [PubMed]
- Jay, M.; Bradley, S.; McDearmid, J.R. Effects of Nitric Oxide on Neuromuscular Properties of Developing Zebrafish Embryos. PLoS ONE 2014, 9, e86930. [Google Scholar] [CrossRef]
- Maake, C.; Kloas, W.; Szendefi, M.; Reinecke, M. Neurohormonal Peptides, Serotonin, and Nitric Oxide Synthase in the Enteric Nervous System and Endocrine Cells of the Gastrointestinal Tract of Neotenic and Thyroid Hormone-Treated Axolotls (Ambystoma mexicanum). Cell Tissue Res. 1999, 297, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Blom, J.J.; Blute, T.A.; Eldred, W.D. Functional Localization of the Nitric Oxide/CGMP Pathway in the Salamander Retina. Vis. Neurosci. 2009, 26, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Trajanovska, S.; Donald, J.A. Endothelial Nitric Oxide Synthase in the Amphibian, Xenopus Tropicalis. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2011, 158, 274–281. [Google Scholar] [CrossRef]
- Brunelli, E.; Perrotta, I.; Talarico, E.; Tripepi, S. Localization of Two Nitric Oxide Synthase Isoforms, ENOS and INOS, in the Skin of Triturus Italicus (Amphibia, Urodela) during Development. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2005, 142, 249–255. [Google Scholar] [CrossRef]
- Jeseta, M.; Marin, M.; Tichovska, H.; Melicharova, P.; Cailliau-Maggio, K.; Martoriati, A.; Lescuyer-Rousseau, A.; Beaujois, R.; Petr, J.; Sedmikova, M.; et al. Nitric Oxide-Donor SNAP Induces Xenopus Eggs Activation. PLoS ONE 2012, 7, e41509. [Google Scholar] [CrossRef]
- Jacox, L.; Sindelka, R.; Chen, J.; Rothman, A.; Dickinson, A.; Sive, H. The Extreme Anterior Domain Is an Essential Craniofacial Organizer Acting through Kinin-Kallikrein Signaling. Cell Rep. 2014, 8, 596–609. [Google Scholar] [CrossRef]
- Vitt, L.J.; Caldwell, J.P. Herpetology: An Introductory Biology of Amphibians and Reptiles, 4th ed.; Academic Press: Cambridge, MA, USA, 2013; 757p. [Google Scholar] [CrossRef]
- Sillar, K.T.; Combes, D.; Ramanathan, S.; Molinari, M.; Simmers, J. Neuromodulation and Developmental Plasticity in the Locomotor System of Anuran Amphibians during Metamorphosis. Brain Res. Rev. 2008, 57, 94–102. [Google Scholar] [CrossRef]
- Cristino, L.; Florenzano, F.; Bentivoglio, M.; Guglielmotti, V. Nitric Oxide Synthase Expression and Cell Changes in Dorsal Root Ganglia and Spinal Dorsal Horn of Developing and Adult Rana Esculenta Indicate a Role of Nitric Oxide in Limb Metamorphosis. J. Comp. Neurol. 2004, 472, 423–436. [Google Scholar] [CrossRef]
- Johnson, J.; Manzo, W.; Gardner, E.; Menon, J. Reactive Oxygen Species and Anti-Oxidant Defenses in Tail of Tadpoles, Xenopus Laevis. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2013, 158, 101–108. [Google Scholar] [CrossRef]
- Hedrick, M.S.; Chen, A.K.; Jessop, K.L. Nitric Oxide Changes Its Role as a Modulator of Respiratory Motor Activity during Development in the Bullfrog (Rana catesbeiana). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2005, 142, 231–240. [Google Scholar] [CrossRef]
- Ueda, N.; Richards, G.S.; Degnan, B.M.; Kranz, A.; Adamska, M.; Croll, R.P.; Degnan, S.M. An Ancient Role for Nitric Oxide in Regulating the Animal Pelagobenthic Life Cycle: Evidence from a Marine Sponge. Sci. Rep. 2016, 6, 37546. [Google Scholar] [CrossRef]
- Moroz, L.L.; Meech, R.W.; Sweedler, J.V.; Mackie, G.O. Nitric Oxide Regulates Swimming in the Jellyfish Aglantha Digitale. J. Comp. Neurol. 2004, 471, 26–36. [Google Scholar] [CrossRef]
- Newcomb, J.M.; Watson, W.H. Modulation of Swimming in the Gastropod Melibe leonina by Nitric Oxide. J. Exp. Biol. 2002, 205, 397–403. [Google Scholar] [CrossRef]
- Mahapatra, C.; Achary, A.S.; Patra, D. Protein S-Nitrosylation: Nitric Oxide Signalling during Anuran Tail Regression. Acta Histochem. 2022, 124, 151899. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Type | Species | Formula | Role |
|---|---|---|---|
| oxidized | nitrate | NO3− | intermediate of NO transformation and source of nitrite under hypoxia |
| oxidized | nitrite | NO2− | signaling agent; intermediate of NO transformation |
| oxidized | peroxynitrite | ONOO− | intermediate of NO transformation; by-product under oxidative stress |
| oxidized | nitrogen dioxide | NO2 | intermediate of NO transformation; by-product under oxidative stress |
| oxidized | dinitrogen dioxide | N2O3 | mobile NO transporter |
| reduced | nitroxyl/nitroxide ion | HNO/NO− | signaling agent |
| reduced | nitroso-thiols | RS-NO | signaling agent; mobile NO transporter |
| reduced | hyponitrite | N2O22− | intermediate of NO transformation |
| Extended Nomenclature | Different Acronyms Present in Literature | Duplicates Nomenclature |
|---|---|---|
| neuronal Nos | nNos, nNOS, bNos, NOSI, NOS1, Nos1 * | Nos1a–Nos1b |
| Nos1α–Nos1β | ||
| inducible Nos | iNos, iNOS, NOSII, NOS2, Nos2 * | Nos2a–Nos2b |
| Nos2ba–Nos2bb | ||
| Nos2.1–Nos2.2 | ||
| Nos2α–Nos2β | ||
| endothelial Nos | eNos, eNOS, NOSIII, NOS3, Nos3 * |
| DEUTEROSTOMES | Nos GENES | REFs | |||||
|---|---|---|---|---|---|---|---|
| AMBULACRARIA | Echinoderms | Echinoids | Strongylocentrotus purpuratus Hemicentrotus pulcherrimus | 2 | [42] [47] [48] | ||
| CHORDATES | Cephalochordates | Asymmetron lucayanum | 3 | [42] [45] [49] [50] [51] | |||
| Branchiostoma floridae Branchiostoma lanceolatum Branchiostoma belcheri | 3 | ||||||
| Urochordates | Ascidians | Ciona robusta # Ciona savignyi | 1 | [42] [46] [52] | |||
| Cyclostomes | Lethenteron camtschaticum Petromyzon marinus | 2 | [42] [53] [54] [55] | ||||
| Eptatretus burgeri | |||||||
| Chondrichthyes | Scyliorhinus torazame | 3 | |||||
| Chiloscyllium punctatum | 2 * | ||||||
| Rhincodon typus | 2 * | ||||||
| Callorhinchus milii | 2 * | ||||||
| Actinopterygians | Polypteriformes | Polypterus senegalus | 3 | ||||
| Acipenseriformes | Acipenser ruthenus | 3 | |||||
| Holosteans | Amia calva Lepisosteus oculatus | 3 | |||||
| Teleosts | Paramormyrops kingsleyae | 3 | |||||
| Salmo salar | 4 | ||||||
| Esox lucius | 3 | ||||||
| Danio rerio | 3 | ||||||
| Cyprinus carpio | 5 | ||||||
| Oryzias latipes | 1 | ||||||
| Sarcopterygians | Coelacathiformes | Latimeria chalumnae | 2 * | ||||
| Tetrapods | Homo sapiens Anolis carolinensis Gallus gallus Xenopus tropicalis | 3 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Locascio, A.; Annona, G.; Caccavale, F.; D’Aniello, S.; Agnisola, C.; Palumbo, A. Nitric Oxide Function and Nitric Oxide Synthase Evolution in Aquatic Chordates. Int. J. Mol. Sci. 2023, 24, 11182. https://doi.org/10.3390/ijms241311182
Locascio A, Annona G, Caccavale F, D’Aniello S, Agnisola C, Palumbo A. Nitric Oxide Function and Nitric Oxide Synthase Evolution in Aquatic Chordates. International Journal of Molecular Sciences. 2023; 24(13):11182. https://doi.org/10.3390/ijms241311182
Chicago/Turabian StyleLocascio, Annamaria, Giovanni Annona, Filomena Caccavale, Salvatore D’Aniello, Claudio Agnisola, and Anna Palumbo. 2023. "Nitric Oxide Function and Nitric Oxide Synthase Evolution in Aquatic Chordates" International Journal of Molecular Sciences 24, no. 13: 11182. https://doi.org/10.3390/ijms241311182
APA StyleLocascio, A., Annona, G., Caccavale, F., D’Aniello, S., Agnisola, C., & Palumbo, A. (2023). Nitric Oxide Function and Nitric Oxide Synthase Evolution in Aquatic Chordates. International Journal of Molecular Sciences, 24(13), 11182. https://doi.org/10.3390/ijms241311182