

New Insights and Potential Therapeutic Interventions in Metabolic Diseases

 ,
,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. Methods
3. The Rise of Metabolic Diseases
4. Genetic Basis
4.1. Genetic Basis of Phenylketonuria and Cystic Fibrosis
4.2. Common Mechanism in Genetic Mutations
4.3. Therapies against Molecular Mechanism
5. Psychological and Behavioral Basis
5.1. Experiential Avoidance and Metabolic Diseases
5.2. Reward Centre and Metabolic Diseases
6. Movement, Physical Activity, and Metabolic Diseases
6.1. Physical Activity
6.1.1. Types of Exercise and Metabolic Diseases
6.1.2. Intensity and Metabolic Diseases
7. Nutrition and Metabolic Diseases
7.1. High-Fat Diets and Metabolic Diseases
7.2. Low-Fat Diets
8. Single-Cell Transcriptomics in Metabolic Diseases
9. Gut Microbiota in Metabolic Diseases
9.1. Gut Microbiome and Obesity
9.2. Gut Microbiome and Dyslipidemia
9.3. Gut, Inflammation and Insulin Resistance
9.4. Future Lines of Intervention
10. Epigenetics and Metabolic Diseases
10.1. Epigenetic Modifications on Inflammation
10.2. Epigenetic Modifications and Gut Microbiome
10.3. Epigenetic Modifications and Expression of Genes
11. Advanced Imaging Techniques and Metabolic Diseases
12. Cell-Based Therapies in Metabolic Diseases
13. Practical Applications
- Patient Education and Support: Providing educational resources and support to patients is crucial in empowering them to manage their metabolic diseases effectively. This includes educating patients about their condition, treatment options, lifestyle modifications, and the importance of medication adherence. Patient support groups and counseling services can also play a significant role in providing emotional support and practical guidance [339,340,341].
- Telemedicine and Remote Monitoring: Embracing telemedicine and remote monitoring technologies can improve accessibility and continuity of care for individuals with metabolic diseases. Remote consultations, mobile applications, and wearable devices enable healthcare providers to monitor patients’ progress, provide real-time feedback, and adjust treatment plans accordingly, even from a distance [342,343,344].
- Behavioral Interventions: Implementing behavioral interventions, such as cognitive-behavioral therapy, motivational interviewing, and mindfulness-based techniques, can help individuals with metabolic diseases make sustainable lifestyle changes. These interventions focus on addressing psychological barriers, promoting self-efficacy, and facilitating behavior modification to support long-term adherence to healthy habits. Early Detection and Risk Assessment: The insights gained from the research on metabolic diseases can be applied in the development of screening tools and risk assessment strategies. By identifying individuals who are at a higher risk of developing metabolic diseases, healthcare professionals can intervene early with targeted interventions, such as lifestyle modifications, to prevent or delay the onset of these conditions [345].
- Personalized Treatment Approaches: The understanding of genetic, psychological, and behavioral factors in metabolic diseases can inform personalized treatment plans. Healthcare providers can consider an individual’s genetic predisposition, psychological factors such as stress and emotional eating, and behavioral patterns to tailor interventions that address the specific needs and challenges of each patient [335,345].
- Nutritional Interventions: The role of nutrition in the development and management of metabolic diseases is crucial. The findings from research can guide the development of evidence-based dietary guidelines and interventions. These interventions can promote healthier eating habits, such as reducing the consumption of processed foods high in fats, sugars, and salt, and increasing the intake of nutrient-dense foods like fruits, vegetables, whole grains, and lean proteins [29,346].
- Exercise Prescription: Prescribing personalized exercise programs, considering factors such as fitness level, health status, and personal preferences, can promote physical activity and improve metabolic health. Collaborating with exercise physiologists or certified fitness professionals can help in designing safe and effective exercise routines and providing ongoing support and guidance [347,348].
- Lifestyle Modification Programs: Physical activity is closely linked to metabolic health. Practical applications derived from research can inform the design of lifestyle modification programs that encourage regular exercise and physical activity. These programs can be tailored to different age groups, fitness levels, and individual preferences, and can include a combination of aerobic exercise, strength training, and flexibility exercises [349].
- Integrative Approaches: Incorporating emerging technologies and therapies can enhance the diagnosis and treatment of metabolic diseases. For example, single-cell transcriptomics can provide insights into cellular-level dysregulation, allowing for targeted therapeutic approaches. Gut microbiota analysis can inform interventions targeting the gut-brain axis and metabolic health. Additionally, advanced imaging techniques can aid in the early detection and monitoring of metabolic diseases [350,351].
- Patient Education and Empowerment: Practical applications of research can be used to develop educational materials and resources for patients with metabolic diseases. Empowering patients with knowledge about their condition, its underlying mechanisms, and the importance of lifestyle modifications can enhance their motivation and adherence to treatment plans [352].
- Long-Term Monitoring and Follow-Up: Regular monitoring of metabolic parameters, such as blood glucose levels, lipid profiles, and body composition, is crucial for evaluating treatment effectiveness and identifying potential complications. Implementing regular follow-up appointments and utilizing digital tools for remote monitoring can facilitate ongoing assessment and timely intervention [335].
14. Limitations and Challenges
- Due to the wide diversity of physical exercise, cognitive, and nutritional protocols, it has been difficult to select studies that favor each of the metabolic diseases in particular.
- In diseases as complex and multifactorial as metabolic diseases, establishing general action guidelines is difficult. The connection between psychological and behavioral factors, nutrition, and metabolic diseases demonstrates the need for an integrative approach to disease prevention and treatment.
- The genetic and epigenetic basis of metabolic diseases is still not fully understood, and interventional therapies are still being thoroughly researched.
- Single-cell transcriptomics is an emerging and promising discipline of molecular biology that has the potential to provide unmatched insight into metabolic diseases; however, there are still few studies on the subject, and more information is needed for a full understanding of the processes.
- To assess the long-term stability of microbiota donor engraftment in humans and its associated phenotypes, further studies with larger sample sizes and durations are needed, and the provision of references should be improved in the future.
15. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heindel, J.J.; Blumberg, B.; Cave, M.; Machtinger, R.; Mantovani, A.; Mendez, M.A.; Nadal, A.; Palanza, P.; Panzica, G.; Sargis, R.; et al. Metabolism Disrupting Chemicals and Metabolic Disorders. Reprod. Toxicol. Elmsford N. Y. 2017, 68, 3–33. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.M. Highlighting Diabetes Mellitus: The Epidemic Continues. Arterioscler. Thromb. Vasc. Biol. 2018, 38, e1–e8. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.L.; Neuschwander-Tetri, B.A.; Rinella, M.; Sanyal, A.J. Mechanisms of NAFLD Development and Therapeutic Strategies. Nat. Med. 2018, 24, 908–922. [Google Scholar] [CrossRef] [PubMed]
- Saklayen, M.G. The Global Epidemic of the Metabolic Syndrome. Curr. Hypertens. Rep. 2018, 20, 12. [Google Scholar] [CrossRef]
- Pryce, J.E.; Parker Gaddis, K.L.; Koeck, A.; Bastin, C.; Abdelsayed, M.; Gengler, N.; Miglior, F.; Heringstad, B.; Egger-Danner, C.; Stock, K.F.; et al. Invited Review: Opportunities for Genetic Improvement of Metabolic Diseases. J. Dairy Sci. 2016, 99, 6855–6873. [Google Scholar] [CrossRef]
- Cao, H. Adipocytokines in Obesity and Metabolic Disease. J. Endocrinol. 2014, 220, T47–T59. [Google Scholar] [CrossRef]
- Willer, C.J.; Schmidt, E.M.; Sengupta, S.; Peloso, G.M.; Gustafsson, S.; Kanoni, S.; Ganna, A.; Chen, J.; Buchkovich, M.L.; Mora, S.; et al. Discovery and Refinement of Loci Associated with Lipid Levels. Nat. Genet. 2013, 45, 1274–1283. [Google Scholar] [CrossRef]
- Grazioli, E.; Dimauro, I.; Mercatelli, N.; Wang, G.; Pitsiladis, Y.; Di Luigi, L.; Caporossi, D. Physical Activity in the Prevention of Human Diseases: Role of Epigenetic Modifications. BMC Genom. 2017, 18 (Suppl. S8), 802. [Google Scholar] [CrossRef]
- Jacobsen, S.C.; Gillberg, L.; Bork-Jensen, J.; Ribel-Madsen, R.; Lara, E.; Calvanese, V.; Ling, C.; Fernandez, A.F.; Fraga, M.F.; Poulsen, P.; et al. Young Men with Low Birthweight Exhibit Decreased Plasticity of Genome-Wide Muscle DNA Methylation by High-Fat Overfeeding. Diabetologia 2014, 57, 1154–1158. [Google Scholar] [CrossRef]
- Anwar, A.; Liu, Y.; Dong, R.; Bai, L.; Yu, X.; Li, Y. The Physiological and Molecular Mechanism of Brassinosteroid in Response to Stress: A Review. Biol. Res. 2018, 51, 46. [Google Scholar] [CrossRef]
- Gallardo-Alfaro, L.; Bibiloni, M.D.M.; Mascaró, C.M.; Montemayor, S.; Ruiz-Canela, M.; Salas-Salvadó, J.; Corella, D.; Fitó, M.; Romaguera, D.; Vioque, J.; et al. Leisure-Time Physical Activity, Sedentary Behaviour and Diet Quality Are Associated with Metabolic Syndrome Severity: The PREDIMED-Plus Study. Nutrients 2020, 12, 1013. [Google Scholar] [CrossRef]
- Owen, N.; Healy, G.N.; Matthews, C.E.; Dunstan, D.W. Too Much Sitting: The Population Health Science of Sedentary Behavior. Exerc. Sport Sci. Rev. 2010, 38, 105–113. [Google Scholar] [CrossRef]
- Siri-Tarino, P.W.; Sun, Q.; Hu, F.B.; Krauss, R.M. Saturated Fatty Acids and Risk of Coronary Heart Disease: Modulation by Replacement Nutrients. Curr. Atheroscler. Rep. 2010, 12, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Te Morenga, L.A.; Levers, M.T.; Williams, S.M.; Brown, R.C.; Mann, J. Comparison of High Protein and High Fiber Weight-Loss Diets in Women with Risk Factors for the Metabolic Syndrome: A Randomized Trial. Nutr. J. 2011, 10, 40. [Google Scholar] [CrossRef]
- Pot, G.K. Sleep and Dietary Habits in the Urban Environment: The Role of Chrono-Nutrition. Proc. Nutr. Soc. 2018, 77, 189–198. [Google Scholar] [CrossRef]
- Rothschild, J.; Hoddy, K.K.; Jambazian, P.; Varady, K.A. Time-Restricted Feeding and Risk of Metabolic Disease: A Review of Human and Animal Studies. Nutr. Rev. 2014, 72, 308–318. [Google Scholar] [CrossRef]
- Segerstolpe, Å.; Palasantza, A.; Eliasson, P.; Andersson, E.-M.; Andréasson, A.-C.; Sun, X.; Picelli, S.; Sabirsh, A.; Clausen, M.; Bjursell, M.K.; et al. Single-Cell Transcriptome Profiling of Human Pancreatic Islets in Health and Type 2 Diabetes. Cell Metab. 2016, 24, 593–607. [Google Scholar] [CrossRef] [PubMed]
- Vijay, J.; Gauthier, M.-F.; Biswell, R.L.; Louiselle, D.A.; Johnston, J.J.; Cheung, W.A.; Belden, B.; Pramatarova, A.; Biertho, L.; Gibson, M.; et al. Single-Cell Analysis of Human Adipose Tissue Identifies Depot and Disease Specific Cell Types. Nat. Metab. 2020, 2, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, A.; Feng, D.; Pessin, J.E.; Shinoda, K. The Impact of Single-Cell Genomics on Adipose Tissue Research. Int. J. Mol. Sci. 2020, 21, 4773. [Google Scholar] [CrossRef]
- Dusaulcy, R.; Handgraaf, S.; Visentin, F.; Howald, C.; Dermitzakis, E.T.; Philippe, J.; Gosmain, Y. High-Fat Diet Impacts More Changes in Beta-Cell Compared to Alpha-Cell Transcriptome. PLoS ONE 2019, 14, e0213299. [Google Scholar] [CrossRef]
- Lu, K.-Y.; Primus Dass, K.T.; Lin, S.-Z.; Harn, H.-J.; Liu, S.-P. The Application of Stem Cell Therapy and Brown Adipose Tissue Transplantation in Metabolic Disorders. Cytotherapy 2020, 22, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Espeland, M.A.; Hayden, K.M.; Lockhart, S.N.; Yassine, H.N.; Hoscheidt, S.; Yasar, S.; Luchsinger, J.A.; Neiberg, R.H.; Diaz Brinton, R.; Carmichael, O. Sex-Related Differences in Brain Volumes and Cerebral Blood Flow Among Overweight and Obese Adults with Type 2 Diabetes: Exploratory Analyses From the Action for Health in Diabetes Brain Magnetic Resonance Imaging Study. J. Gerontol. A Biol. Sci. Med. Sci. 2020, 75, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Latteri, S.; Sofia, M.; Puleo, S.; Di Vincenzo, A.; Cinti, S.; Castorina, S. Mechanisms Linking Bariatric Surgery to Adipose Tissue, Glucose Metabolism, Fatty Liver Disease and Gut Microbiota. Langenbeck’s Arch. Surg. 2023, 408, 101. [Google Scholar] [CrossRef] [PubMed]
- Rebelos, E.; Rinne, J.O.; Nuutila, P.; Ekblad, L.L. Brain Glucose Metabolism in Health, Obesity, and Cognitive Decline-Does Insulin Have Anything to Do with It? A Narrative Review. J. Clin. Med. 2021, 10, 1532. [Google Scholar] [CrossRef] [PubMed]
- Pahk, K.; Joung, C.; Kwon, H.W.; Kim, S. Chronic Physical Exercise Alleviates Stress-Associated Amygdala Metabolic Activity in Obese Women: A Prospective Serial (18)F-FDG PET/CT Study. Front. Endocrinol. 2022, 13, 1046838. [Google Scholar] [CrossRef]
- van den Bergh, J.P.; Szulc, P.; Cheung, A.M.; Bouxsein, M.; Engelke, K.; Chapurlat, R. The Clinical Application of High-Resolution Peripheral Computed Tomography (HR-PQCT) in Adults: State of the Art and Future Directions. Osteoporos. Int. 2021, 32, 1465–1485. [Google Scholar] [CrossRef]
- Clemente-Suárez, V.J.; Redondo-Flórez, L.; Beltrán-Velasco, A.I.; Martín-Rodríguez, A.; Martínez-Guardado, I.; Navarro-Jiménez, E.; Laborde-Cárdenas, C.C.; Tornero-Aguilera, J.F. The Role of Adipokines in Health and Disease. Biomedicines 2023, 11, 1290. [Google Scholar] [CrossRef]
- Clemente-Suárez, V.J.; Martín-Rodríguez, A.; Redondo-Flórez, L.; Ruisoto, P.; Navarro-Jiménez, E.; Ramos-Campo, D.J.; Tornero-Aguilera, J.F. Metabolic Health, Mitochondrial Fitness, Physical Activity, and Cancer. Cancers 2023, 15, 814. [Google Scholar] [CrossRef]
- Clemente-Suárez, V.J.; Mielgo-Ayuso, J.; Martín-Rodríguez, A.; Ramos-Campo, D.J.; Redondo-Flórez, L.; Tornero-Aguilera, J.F. The Burden of Carbohydrates in Health and Disease. Nutrients 2022, 14, 3809. [Google Scholar] [CrossRef]
- Boulangé, C.L.; Neves, A.L.; Chilloux, J.; Nicholson, J.K.; Dumas, M.-E. Impact of the Gut Microbiota on Inflammation, Obesity, and Metabolic Disease. Genome Med. 2016, 8, 42. [Google Scholar] [CrossRef]
- Chooi, Y.C.; Ding, C.; Magkos, F. The Epidemiology of Obesity. Metabolism 2019, 92, 6–10. [Google Scholar] [CrossRef]
- Stanhope, K.L. Sugar Consumption, Metabolic Disease and Obesity: The State of the Controversy. Crit. Rev. Clin. Lab. Sci. 2016, 53, 52–67. [Google Scholar] [CrossRef]
- Barroso, I.; McCarthy, M.I. The Genetic Basis of Metabolic Disease. Cell 2019, 177, 146–161. [Google Scholar] [CrossRef] [PubMed]
- Kopp, W. How Western Diet And Lifestyle Drive The Pandemic Of Obesity And Civilization Diseases. Diabetes Metab. Syndr. Obes. 2019, 12, 2221–2236. [Google Scholar] [CrossRef] [PubMed]
- Aranceta-Bartrina, J.; Gianzo-Citores, M.; Pérez-Rodrigo, C. Prevalence of Overweight, Obesity and Abdominal Obesity in the Spanish Population Aged 3 to 24 Years. The ENPE Study. Rev. Esp. Cardiol. 2020, 73, 290–299. [Google Scholar] [CrossRef]
- Jackson, S.E.; Llewellyn, C.H.; Smith, L. The Obesity Epidemic—Nature via Nurture: A Narrative Review of High-Income Countries. SAGE Open Med. 2020, 8, 2050312120918265. [Google Scholar] [CrossRef] [PubMed]
- Katzmarzyk, P.T.; Chaput, J.-P.; Fogelholm, M.; Hu, G.; Maher, C.; Maia, J.; Olds, T.; Sarmiento, O.L.; Standage, M.; Tremblay, M.S.; et al. International Study of Childhood Obesity, Lifestyle and the Environment (ISCOLE): Contributions to Understanding the Global Obesity Epidemic. Nutrients 2019, 11, 848. [Google Scholar] [CrossRef]
- Wang, Y.; Beydoun, M.A.; Min, J.; Xue, H.; Kaminsky, L.A.; Cheskin, L.J. Has the Prevalence of Overweight, Obesity and Central Obesity Levelled off in the United States? Trends, Patterns, Disparities, and Future Projections for the Obesity Epidemic. Int. J. Epidemiol. 2020, 49, 810–823. [Google Scholar] [CrossRef]
- Fernández, R. Impacto del Sobrepeso y la Obesidad en el Producto Interior Bruto a Nivel Mundial de 2020 a 2035. Available online: https://es.statista.com/estadisticas/1371046/impacto-del-sobrepeso-y-de-la-obesidad-en-el-producto-interior-bruto-a-nivel-mundial/ (accessed on 25 May 2023).
- Lovic, D.; Piperidou, A.; Zografou, I.; Grassos, H.; Pittaras, A.; Manolis, A. The Growing Epidemic of Diabetes Mellitus. Curr. Vasc. Pharmacol. 2020, 18, 104–109. [Google Scholar] [CrossRef]
- Standl, E.; Khunti, K.; Hansen, T.B.; Schnell, O. The Global Epidemics of Diabetes in the 21st Century: Current Situation and Perspectives. Eur. J. Prev. Cardiol. 2019, 26 (Suppl. S1), 7–14. [Google Scholar] [CrossRef]
- Reisinger, C.; Nkeh-Chungag, B.N.; Fredriksen, P.M.; Goswami, N. The Prevalence of Pediatric Metabolic Syndrome—A Critical Look on the Discrepancies between Definitions and Its Clinical Importance. Int. J. Obes. 2021, 45, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.; Weitzman, M.; Auinger, P.; Nguyen, M.; Dietz, W.H. Prevalence of a Metabolic Syndrome Phenotype in Adolescents: Findings from the Third National Health and Nutrition Examination Survey, 1988–1994. Arch. Pediatr. Adolesc. Med. 2003, 157, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Cruz, M.L.; Weigensberg, M.J.; Huang, T.T.-K.; Ball, G.; Shaibi, G.Q.; Goran, M.I. The Metabolic Syndrome in Overweight Hispanic Youth and the Role of Insulin Sensitivity. J. Clin. Endocrinol. Metab. 2004, 89, 108–113. [Google Scholar] [CrossRef]
- Lekka, D.E.; Brucknerova, J.; Salingova, A.; Sebova, C.; Ostrozlikova, M.; Ziburova, J.; Nemcovic, M.; Sestak, S.; Bellova, J.; Pakanova, Z.; et al. Congenital Disorders of Glycosylation—An Umbrella Term for Rapidly Expanding Group of Rare Genetic Metabolic Disorders—Importance of Physical Investigation. Bratisl. Lek. Listy 2021, 122, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Tomiyama, A.J. Stress and Obesity. Annu. Rev. Psychol. 2019, 70, 703–718. [Google Scholar] [CrossRef]
- Hewagalamulage, S.D.; Lee, T.K.; Clarke, I.J.; Henry, B.A. Stress, Cortisol, and Obesity: A Role for Cortisol Responsiveness in Identifying Individuals Prone to Obesity. Domest. Anim. Endocrinol. 2016, 56, S112–S120. [Google Scholar] [CrossRef]
- van der Valk, E.S.; Savas, M.; van Rossum, E.F.C. Stress and Obesity: Are There More Susceptible Individuals? Curr. Obes. Rep. 2018, 7, 193–203. [Google Scholar] [CrossRef]
- Roy, P.; Chakrabarty, S.; Chatterjee, D.; Bharati, P. Prevalence and Factors Associated with Overweight/Obesity in Adolescent School Girls: A Cross-Sectional Study in Kolkata, India. Anthropol. Rev. 2022, 85, 147–162. [Google Scholar] [CrossRef]
- Gómez-Pérez, D.; Cancino, M.; Moreno, P.I.; Ortiz, M.S. Weight Stigma, Chronic Stress, Unhealthy Diet, and Obesity in Chilean Adults. Int. J. Behav. Med. 2021, 28, 292–298. [Google Scholar] [CrossRef]
- James, W.P.T.; Johnson, R.J.; Speakman, J.R.; Wallace, D.C.; Frühbeck, G.; Iversen, P.O.; Stover, P.J. Nutrition and Its Role in Human Evolution. J. Intern. Med. 2019, 285, 533–549. [Google Scholar] [CrossRef]
- Ramón-Arbués, E.; Martínez Abadía, B.; Granada López, J.M.; Echániz Serrano, E.; Pellicer García, B.; Juárez Vela, R.; Guerrero Portillo, S.; Saéz Guinoa, M. Eating behavior and relationships with stress, anxiety, depression and insomnia in university students. Nutr. Hosp. 2019, 36, 1339–1345. [Google Scholar] [CrossRef] [PubMed]
- Kuo, L.E.; Czarnecka, M.; Kitlinska, J.B.; Tilan, J.U.; Kvetnanský, R.; Zukowska, Z. Chronic Stress, Combined with a High-Fat/High-Sugar Diet, Shifts Sympathetic Signaling toward Neuropeptide Y and Leads to Obesity and the Metabolic Syndrome. Ann. N. Y. Acad. Sci. 2008, 1148, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Jacques, A.; Chaaya, N.; Beecher, K.; Ali, S.A.; Belmer, A.; Bartlett, S. The Impact of Sugar Consumption on Stress Driven, Emotional and Addictive Behaviors. Neurosci. Biobehav. Rev. 2019, 103, 178–199. [Google Scholar] [CrossRef] [PubMed]
- Woods, N.P.; Tangpukdee, J.; Thepa, T.; Methakanchanasak, N. Consequences of Sleep Deprivation in Adult Diabetes Mellitus Type 2 Patients: An Integrative Review. Open Access Maced. J. Med. Sci. 2023, 11, 1–10. [Google Scholar] [CrossRef]
- Kawai, T.; Autieri, M.V.; Scalia, R. Adipose Tissue Inflammation and Metabolic Dysfunction in Obesity. Am. J. Physiol. Cell Physiol. 2021, 320, C375–C391. [Google Scholar] [CrossRef]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in Inflammation and Metabolic Disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef]
- Vockley, J.; Andersson, H.C.; Antshel, K.M.; Braverman, N.E.; Burton, B.K.; Frazier, D.M.; Mitchell, J.; Smith, W.E.; Thompson, B.H.; Berry, S.A. Phenylalanine Hydroxylase Deficiency: Diagnosis and Management Guideline. Genet. Med. Off. J. Am. Coll. Med. Genet. 2014, 16, 188–200. [Google Scholar] [CrossRef]
- Blau, N.; Bélanger-Quintana, A.; Demirkol, M.; Feillet, F.; Giovannini, M.; MacDonald, A.; Trefz, F.K.; van Spronsen, F. Management of Phenylketonuria in Europe: Survey Results from 19 Countries. Mol. Genet. Metab. 2010, 99, 109–115. [Google Scholar] [CrossRef]
- Han, L.; Roberts, M.; Luo, A.; Wei, S.; Slayden, O.D.; Macdonald, K.D. Functional Evaluation of the Cystic Fibrosis Transmembrane Conductance Regulator in the Endocervix†. Biol. Reprod. 2022, 107, 732–740. [Google Scholar] [CrossRef]
- Knowles, M.R.; Durie, P.R. What Is Cystic Fibrosis? N. Engl. J. Med. 2002, 347, 439–442. [Google Scholar] [CrossRef]
- Zimmet, P.; Alberti, K.G.; Shaw, J. Global and Societal Implications of the Diabetes Epidemic. Nature 2001, 414, 782–787. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D. Microbiota and Metabolites in Metabolic Diseases. Nat. Rev. Endocrinol. 2019, 15, 69–70. [Google Scholar] [CrossRef] [PubMed]
- Chandler, R.J.; Venditti, C.P. Gene Therapy for Metabolic Diseases. Transl. Sci. Rare Dis. 2016, 1, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Sacks, F.M.; Bray, G.A.; Carey, V.J.; Smith, S.R.; Ryan, D.H.; Anton, S.D.; McManus, K.; Champagne, C.M.; Bishop, L.M.; Laranjo, N.; et al. Comparison of Weight-Loss Diets with Different Compositions of Fat, Protein, and Carbohydrates. N. Engl. J. Med. 2009, 360, 859–873. [Google Scholar] [CrossRef] [PubMed]
- Koeberl, D.D.; Pinto, C.; Brown, T.; Chen, Y.T. Gene Therapy for Inherited Metabolic Disorders in Companion Animals. ILAR J. 2009, 50, 122–127. [Google Scholar] [CrossRef]
- Estruch, R.; Ros, E.; Salas-Salvadó, J.; Covas, M.-I.; Corella, D.; Arós, F.; Gómez-Gracia, E.; Ruiz-Gutiérrez, V.; Fiol, M.; Lapetra, J. Primary Prevention of Cardiovascular Disease with a Mediterranean Diet. N. Engl. J. Med. 2013, 368, 1279–1290. [Google Scholar] [CrossRef]
- Barrea, L.; Arnone, A.; Annunziata, G.; Muscogiuri, G.; Laudisio, D.; Salzano, C.; Pugliese, G.; Colao, A.; Savastano, S. Adherence to the Mediterranean Diet, Dietary Patterns and Body Composition in Women with Polycystic Ovary Syndrome (PCOS). Nutrients 2019, 11, 2278. [Google Scholar] [CrossRef]
- Naja, F.; Nasreddine, L.; Itani, L.; Adra, N.; Sibai, A.M.; Hwalla, N. Association between Dietary Patterns and the Risk of Metabolic Syndrome among Lebanese Adults. Eur. J. Nutr. 2013, 52, 97–105. [Google Scholar] [CrossRef]
- García-García, E.; De la Llata-Romero, M.; Kaufer-Horwitz, M.; Tusié-Luna, M.T.; Calzada-León, R.; Vázquez-Velázquez, V.; Barquera-Cervera, S.; Caballero-Romo, A.J.; Orozco, L.; Velásquez-Fernández, D.; et al. Obesity and metabolic syndrome as public health problem. A reflection. Arch. Cardiol. Mex. 2008, 78, 318–337. [Google Scholar]
- Palomino, A.M. The Role of Emotion in Eating Behavior Rol de La Emoción En La Conducta Alimentaria. Rev. Chil. Nutr. 2020, 47, 286–291. [Google Scholar] [CrossRef]
- Aguirre, P. Ricos Flacos y Gordos Pobres: La Alimentación En Crisis; Capital Intelectual: Buenos Aires, Argentina, 2004. [Google Scholar]
- García-Flores, C.L.; Martínez Moreno, A.G.; Beltrán Miranda, C.P.; Zepeda-Salvador, A.P.; Solano Santos, L.V. Satiation and satiety in the regulation of energy intake. Rev. Med. Chil. 2017, 145, 1172–1178. [Google Scholar] [CrossRef] [PubMed]
- López-Navarrete, G.E.; Perea-Martínez, A.; Loredo-Abdalá, A. Obesidad y Maltrato Infantil. Un Fenómeno Bidireccional. Acta Pediátr. Méx. 2008, 29, 342–346. [Google Scholar]
- Ruiz, L.D.; Zuelch, M.L.; Dimitratos, S.M.; Scherr, R.E. Adolescent Obesity: Diet Quality, Psychosocial Health, and Cardiometabolic Risk Factors. Nutrients 2019, 12, 43. [Google Scholar] [CrossRef] [PubMed]
- Luciano, C.; Hayes, S.C. Trastorno de Evitación Experiencial. Int. J. Clin. Health Psychol. 2001, 1, 109–157. [Google Scholar]
- Finger, I.d.R.; de Freitas, B.I.; Oliveira, M.d.S. Psychological Inflexibility in Overweight and Obese People from the Perspective of Acceptance and Commitment Therapy (ACT). Eat. Weight Disord. 2020, 25, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Hayes, S.C.; Wilson, K.G.; Gifford, E.V.; Follette, V.M.; Strosahl, K. Experimental Avoidance and Behavioral Disorders: A Functional Dimensional Approach to Diagnosis and Treatment. J. Consult. Clin. Psychol. 1996, 64, 1152–1168. [Google Scholar] [CrossRef] [PubMed]
- Russ, D.W.; Kent-Braun, J.A. Sex Differences in Human Skeletal Muscle Fatigue Are Eliminated under Ischemic Conditions. J. Appl. Physiol. 2003, 94, 2414–2422. [Google Scholar] [CrossRef]
- Ulrich-Lai, Y.M.; Fulton, S.; Wilson, M.; Petrovich, G.; Rinaman, L. Stress Exposure, Food Intake and Emotional State. Stress 2015, 18, 381–399. [Google Scholar] [CrossRef]
- Litwin, R.; Goldbacher, E.M.; Cardaciotto, L.; Gambrel, L.E. Negative Emotions and Emotional Eating: The Mediating Role of Experiential Avoidance. Eat. Weight Disord. 2017, 22, 97–104. [Google Scholar] [CrossRef]
- Leppanen, J.; Brown, D.; McLinden, H.; Williams, S.; Tchanturia, K. The Role of Emotion Regulation in Eating Disorders: A Network Meta-Analysis Approach. Front. Psychiatry 2022, 13, 793094. [Google Scholar] [CrossRef]
- Fahrenkamp, A.J.; Darling, K.E.; Ruzicka, E.B.; Sato, A.F. Food Cravings and Eating: The Role of Experiential Avoidance. Int. J. Environ. Res. Public Health 2019, 16, 1181. [Google Scholar] [CrossRef] [PubMed]
- Suez, J.; Cohen, Y.; Valdés-Mas, R.; Mor, U.; Dori-Bachash, M.; Federici, S.; Zmora, N.; Leshem, A.; Heinemann, M.; Linevsky, R.; et al. Personalized Microbiome-Driven Effects of Non-Nutritive Sweeteners on Human Glucose Tolerance. Cell 2022, 185, 3307–3328.e19. [Google Scholar] [CrossRef] [PubMed]
- González-Jiménez, E.; Schmidt Río-Valle, J. Regulation of dietary intake and energy balance: Factors and mechanisms involved. Nutr. Hosp. 2012, 27, 1850–1859. [Google Scholar] [CrossRef] [PubMed]
- Schulte, E.M.; Avena, N.M.; Gearhardt, A.N. Which Foods May Be Addictive? The Roles of Processing, Fat Content, and Glycemic Load. PLoS ONE 2015, 10, e0117959. [Google Scholar] [CrossRef]
- Muñoz-Rodríguez, J.R. Capítulo 1 El Cerebro Obeso. Cómo Reacciona Nuestro Cerebro Ante El Problema de La Obesidad. Aliment. Cereb. 2021, 23, 15. [Google Scholar]
- Neff, K.M.H.; Fay, A.; Saules, K.K. Foods and Nutritional Characteristics Associated with Addictive-Like Eating. Psychol. Rep. 2022, 125, 1937–1956. [Google Scholar] [CrossRef]
- Shefer, G.; Marcus, Y.; Stern, N. Is Obesity a Brain Disease? Neurosci. Biobehav. Rev. 2013, 37 Pt 2, 2489–2503. [Google Scholar] [CrossRef]
- Stice, E.; Spoor, S.; Bohon, C.; Veldhuizen, M.G.; Small, D.M. Relation of Reward from Food Intake and Anticipated Food Intake to Obesity: A Functional Magnetic Resonance Imaging Study. J. Abnorm. Psychol. 2008, 117, 924–935. [Google Scholar] [CrossRef]
- Mozaffarian, D.; Appel, L.J.; Van Horn, L. Components of a Cardioprotective Diet: New Insights. Circulation 2011, 123, 2870–2891. [Google Scholar] [CrossRef]
- Rochlani, Y.; Pothineni, N.V.; Kovelamudi, S.; Mehta, J.L. Metabolic Syndrome: Pathophysiology, Management, and Modulation by Natural Compounds. Ther. Adv. Cardiovasc. Dis. 2017, 11, 215–225. [Google Scholar] [CrossRef]
- Aguilar, M.; Bhuket, T.; Torres, S.; Liu, B.; Wong, R.J. Prevalence of the Metabolic Syndrome in the United States, 2003–2012. JAMA 2015, 313, 1973–1974. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Bergés, D.; Cabrera de León, A.; Sanz, H.; Elosua, R.; Guembe, M.J.; Alzamora, M.; Vega-Alonso, T.; Félix-Redondo, F.J.; Ortiz-Marrón, H.; Rigo, F.; et al. Metabolic Syndrome in Spain: Prevalence and Coronary Risk Associated with Harmonized Definition and WHO Proposal. DARIOS Study. Rev. Esp. Cardiol. 2012, 65, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z. The Metabolic Syndrome. Lancet 2005, 365, 1415–1428. [Google Scholar] [CrossRef] [PubMed]
- Ostman, C.; Smart, N.A.; Morcos, D.; Duller, A.; Ridley, W.; Jewiss, D. The Effect of Exercise Training on Clinical Outcomes in Patients with the Metabolic Syndrome: A Systematic Review and Meta-Analysis. Cardiovasc. Diabetol. 2017, 16, 110. [Google Scholar] [CrossRef] [PubMed]
- Moghetti, P.; Bacchi, E.; Brangani, C.; Donà, S.; Negri, C. Metabolic Effects of Exercise. Front. Horm. Res. 2016, 47, 44–57. [Google Scholar] [CrossRef]
- Evans, E.M.; Racette, S.B.; Peterson, L.R.; Villareal, D.T.; Greiwe, J.S.; Holloszy, J.O. Aerobic Power and Insulin Action Improve in Response to Endurance Exercise Training in Healthy 77–87 Yr Olds. J. Appl. Physiol. 2005, 98, 40–45. [Google Scholar] [CrossRef]
- Westcott, W.L. Resistance Training Is Medicine: Effects of Strength Training on Health. Curr. Sport. Med. Rep. 2012, 11, 209–216. [Google Scholar] [CrossRef]
- Hansen, D.; De Strijcker, D.; Calders, P. Impact of Endurance Exercise Training in the Fasted State on Muscle Biochemistry and Metabolism in Healthy Subjects: Can These Effects Be of Particular Clinical Benefit to Type 2 Diabetes Mellitus and Insulin-Resistant Patients? Sport. Med. 2017, 47, 415–428. [Google Scholar] [CrossRef]
- Sluik, D.; Buijsse, B.; Muckelbauer, R.; Kaaks, R.; Teucher, B.; Johnsen, N.F.; Tjønneland, A.; Overvad, K.; Ostergaard, J.N.; Amiano, P.; et al. Physical Activity and Mortality in Individuals with Diabetes Mellitus: A Prospective Study and Meta-Analysis. Arch. Intern. Med. 2012, 172, 1285–1295. [Google Scholar] [CrossRef]
- Borror, A.; Zieff, G.; Battaglini, C.; Stoner, L. The Effects of Postprandial Exercise on Glucose Control in Individuals with Type 2 Diabetes: A Systematic Review. Sport. Med. 2018, 48, 1479–1491. [Google Scholar] [CrossRef]
- Colberg, S.R.; Sigal, R.J.; Yardley, J.E.; Riddell, M.C.; Dunstan, D.W.; Dempsey, P.C.; Horton, E.S.; Castorino, K.; Tate, D.F. Physical Activity/Exercise and Diabetes: A Position Statement of the American Diabetes Association. Diabetes Care 2016, 39, 2065–2079. [Google Scholar] [CrossRef] [PubMed]
- Dunstan, D.W.; Daly, R.M.; Owen, N.; Jolley, D.; De Courten, M.; Shaw, J.; Zimmet, P. High-Intensity Resistance Training Improves Glycemic Control in Older Patients with Type 2 Diabetes. Diabetes Care 2002, 25, 1729–1736. [Google Scholar] [CrossRef] [PubMed]
- Reddy, R.; Wittenberg, A.; Castle, J.R.; El Youssef, J.; Winters-Stone, K.; Gillingham, M.; Jacobs, P.G. Effect of Aerobic and Resistance Exercise on Glycemic Control in Adults with Type 1 Diabetes. Can. J. Diabetes 2019, 43, 406–414.e1. [Google Scholar] [CrossRef]
- Sigal, R.J.; Kenny, G.P. Combined Aerobic and Resistance Exercise for Patients with Type 2 Diabetes. JAMA 2010, 304, 2298–2299. [Google Scholar] [CrossRef]
- Church, T.S.; Blair, S.N.; Cocreham, S.; Johannsen, N.; Johnson, W.; Kramer, K.; Mikus, C.R.; Myers, V.; Nauta, M.; Rodarte, R.Q.; et al. Effects of Aerobic and Resistance Training on Hemoglobin A1c Levels in Patients with Type 2 Diabetes: A Randomized Controlled Trial. JAMA 2010, 304, 2253–2262. [Google Scholar] [CrossRef] [PubMed]
- Hai-Guo, L.; Guo-Zheng, L.I. The Therapeutic Effect of Aerobic Exercise with Resistance Training in Elderly Men with Type 2 Diabetes Mellitus. Zhongguo Yingyong Shenglixue Zazhi = Chin. J. Appl. Physiol. 2016, 32, 529–532. [Google Scholar]
- Schwingshackl, L.; Missbach, B.; Dias, S.; König, J.; Hoffmann, G. Impact of Different Training Modalities on Glycaemic Control and Blood Lipids in Patients with Type 2 Diabetes: A Systematic Review and Network Meta-Analysis. Diabetologia 2014, 57, 1789–1797. [Google Scholar] [CrossRef]
- Snowling, N.J.; Hopkins, W.G. Effects of Different Modes of Exercise Training on Glucose Control and Risk Factors for Complications in Type 2 Diabetic Patients: A Meta-Analysis. Diabetes Care 2006, 29, 2518–2527. [Google Scholar] [CrossRef]
- Villareal, D.T.; Aguirre, L.; Gurney, A.B.; Waters, D.L.; Sinacore, D.R.; Colombo, E.; Armamento-Villareal, R.; Qualls, C. Aerobic or Resistance Exercise, or Both, in Dieting Obese Older Adults. N. Engl. J. Med. 2017, 376, 1943–1955. [Google Scholar] [CrossRef]
- Earnest, C.P.; Johannsen, N.M.; Swift, D.L.; Gillison, F.B.; Mikus, C.R.; Lucia, A.; Kramer, K.; Lavie, C.J.; Church, T.S. Aerobic and Strength Training in Concomitant Metabolic Syndrome and Type 2 Diabetes. Med. Sci. Sport. Exerc. 2014, 46, 1293–1301. [Google Scholar] [CrossRef]
- Ramalho, A.C.; de Lourdes Lima, M.; Nunes, F.; Cambuí, Z.; Barbosa, C.; Andrade, A.; Viana, A.; Martins, M.; Abrantes, V.; Aragão, C.; et al. The Effect of Resistance versus Aerobic Training on Metabolic Control in Patients with Type-1 Diabetes Mellitus. Diabetes Res. Clin. Pract. 2006, 72, 271–276. [Google Scholar] [CrossRef]
- Balducci, S.; Sacchetti, M.; Haxhi, J.; Orlando, G.; D’Errico, V.; Fallucca, S.; Menini, S.; Pugliese, G. Physical Exercise as Therapy for Type 2 Diabetes Mellitus. Diabetes Metab. Res. Rev. 2014, 30 (Suppl. S1), 13–23. [Google Scholar] [CrossRef]
- Jarvie, J.L.; Pandey, A.; Ayers, C.R.; McGavock, J.M.; Sénéchal, M.; Berry, J.D.; Patel, K.V.; McGuire, D.K. Aerobic Fitness and Adherence to Guideline-Recommended Minimum Physical Activity Among Ambulatory Patients with Type 2 Diabetes Mellitus. Diabetes Care 2019, 42, 1333–1339. [Google Scholar] [CrossRef]
- Pan, B.; Ge, L.; Xun, Y.-Q.; Chen, Y.-J.; Gao, C.-Y.; Han, X.; Zuo, L.-Q.; Shan, H.-Q.; Yang, K.-H.; Ding, G.-W.; et al. Exercise Training Modalities in Patients with Type 2 Diabetes Mellitus: A Systematic Review and Network Meta-Analysis. Int. J. Behav. Nutr. Phys. Act. 2018, 15, 72. [Google Scholar] [CrossRef] [PubMed]
- Kanaley, J.A.; Colberg, S.R.; Corcoran, M.H.; Malin, S.K.; Rodriguez, N.R.; Crespo, C.J.; Kirwan, J.P.; Zierath, J.R. Exercise/Physical Activity in Individuals with Type 2 Diabetes: A Consensus Statement from the American College of Sports Medicine. Med. Sci. Sport. Exerc. 2022, 54, 353–368. [Google Scholar] [CrossRef]
- Giannopoulou, I.; Fernhall, B.; Carhart, R.; Weinstock, R.S.; Baynard, T.; Figueroa, A.; Kanaley, J.A. Effects of Diet and/or Exercise on the Adipocytokine and Inflammatory Cytokine Levels of Postmenopausal Women with Type 2 Diabetes. Metabolism 2005, 54, 866–875. [Google Scholar] [CrossRef]
- Yassine, H.N.; Marchetti, C.M.; Krishnan, R.K.; Vrobel, T.R.; Gonzalez, F.; Kirwan, J.P. Effects of Exercise and Caloric Restriction on Insulin Resistance and Cardiometabolic Risk Factors in Older Obese Adults—A Randomized Clinical Trial. J. Gerontol. A Biol. Sci. Med. Sci. 2009, 64, 90–95. [Google Scholar] [CrossRef]
- Ross, R.; Janssen, I.; Dawson, J.; Kungl, A.-M.; Kuk, J.L.; Wong, S.L.; Nguyen-Duy, T.-B.; Lee, S.; Kilpatrick, K.; Hudson, R. Exercise-Induced Reduction in Obesity and Insulin Resistance in Women: A Randomized Controlled Trial. Obes. Res. 2004, 12, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Ross, R.; Dagnone, D.; Jones, P.J.; Smith, H.; Paddags, A.; Hudson, R.; Janssen, I. Reduction in Obesity and Related Comorbid Conditions after Diet-Induced Weight Loss or Exercise-Induced Weight Loss in Men. A Randomized, Controlled Trial. Ann. Intern. Med. 2000, 133, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Hwang, C.-L.; Lim, J.; Yoo, J.-K.; Kim, H.-K.; Hwang, M.-H.; Handberg, E.M.; Petersen, J.W.; Holmer, B.J.; Leey Casella, J.A.; Cusi, K.; et al. Effect of All-Extremity High-Intensity Interval Training vs. Moderate-Intensity Continuous Training on Aerobic Fitness in Middle-Aged and Older Adults with Type 2 Diabetes: A Randomized Controlled Trial. Exp. Gerontol. 2019, 116, 46–53. [Google Scholar] [CrossRef]
- Jelleyman, C.; Yates, T.; O’Donovan, G.; Gray, L.J.; King, J.A.; Khunti, K.; Davies, M.J. The Effects of High-Intensity Interval Training on Glucose Regulation and Insulin Resistance: A Meta-Analysis. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2015, 16, 942–961. [Google Scholar] [CrossRef] [PubMed]
- Gibala, M.J.; Little, J.P.; Macdonald, M.J.; Hawley, J.A. Physiological Adaptations to Low-Volume, High-Intensity Interval Training in Health and Disease. J. Physiol. 2012, 590, 1077–1084. [Google Scholar] [CrossRef] [PubMed]
- Fisher, G.; Brown, A.W.; Bohan Brown, M.M.; Alcorn, A.; Noles, C.; Winwood, L.; Resuehr, H.; George, B.; Jeansonne, M.M.; Allison, D.B. High Intensity Interval- vs Moderate Intensity- Training for Improving Cardiometabolic Health in Overweight or Obese Males: A Randomized Controlled Trial. PLoS ONE 2015, 10, e0138853. [Google Scholar] [CrossRef] [PubMed]
- Roberts, C.K.; Hevener, A.L.; Barnard, R.J. Metabolic Syndrome and Insulin Resistance: Underlying Causes and Modification by Exercise Training. Compr. Physiol. 2013, 3, 1–58. [Google Scholar] [CrossRef]
- Kirwan, J.P.; Sacks, J.; Nieuwoudt, S. The Essential Role of Exercise in the Management of Type 2 Diabetes. Cleve. Clin. J. Med. 2017, 84 (7 Suppl. S1), S15–S21. [Google Scholar] [CrossRef]
- Ordovas, J.M.; Ferguson, L.R.; Tai, E.S.; Mathers, J.C. Personalised Nutrition and Health. BMJ 2018, 361, bmj.k2173. [Google Scholar] [CrossRef]
- de Toro-Martín, J.; Arsenault, B.J.; Després, J.-P.; Vohl, M.-C. Precision Nutrition: A Review of Personalized Nutritional Approaches for the Prevention and Management of Metabolic Syndrome. Nutrients 2017, 9, 913. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.D.; Hu, F.B. Precision Nutrition for Prevention and Management of Type 2 Diabetes. Lancet Diabetes Endocrinol. 2018, 6, 416–426. [Google Scholar] [CrossRef]
- Ramos-Lopez, O.; Milagro, F.I.; Riezu-Boj, J.I.; Martinez, J.A. Epigenetic Signatures Underlying Inflammation: An Interplay of Nutrition, Physical Activity, Metabolic Diseases, and Environmental Factors for Personalized Nutrition. Inflamm. Res. 2021, 70, 29–49. [Google Scholar] [CrossRef]
- Ramallal, R.; Toledo, E.; Martínez, J.A.; Shivappa, N.; Hébert, J.R.; Martínez-González, M.A.; Ruiz-Canela, M. Inflammatory Potential of Diet, Weight Gain, and Incidence of Overweight/Obesity: The SUN Cohort. Obesity 2017, 25, 997–1005. [Google Scholar] [CrossRef]
- Christ, A.; Lauterbach, M.; Latz, E. Western Diet and the Immune System: An Inflammatory Connection. Immunity 2019, 51, 794–811. [Google Scholar] [CrossRef] [PubMed]
- Guariguata, L.; Whiting, D.R.; Hambleton, I.; Beagley, J.; Linnenkamp, U.; Shaw, J.E. Global Estimates of Diabetes Prevalence for 2013 and Projections for 2035. Diabetes Res. Clin. Pract. 2014, 103, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Rakhra, V.; Galappaththy, S.L.; Bulchandani, S.; Cabandugama, P.K. Obesity and the Western Diet: How We Got Here. Mo. Med. 2020, 117, 536–538. [Google Scholar] [PubMed]
- Aleksandrova, K.; Koelman, L.; Rodrigues, C.E. Dietary Patterns and Biomarkers of Oxidative Stress and Inflammation: A Systematic Review of Observational and Intervention Studies. Redox Biol. 2021, 42, 101869. [Google Scholar] [CrossRef]
- van den Brink, W.; van Bilsen, J.; Salic, K.; Hoevenaars, F.P.M.; Verschuren, L.; Kleemann, R.; Bouwman, J.; Ronnett, G.V.; van Ommen, B.; Wopereis, S. Current and Future Nutritional Strategies to Modulate Inflammatory Dynamics in Metabolic Disorders. Front. Nutr. 2019, 6, 129. [Google Scholar] [CrossRef] [PubMed]
- Balistreri, C.R.; Caruso, C.; Candore, G. The Role of Adipose Tissue and Adipokines in Obesity-Related Inflammatory Diseases. Mediat. Inflamm. 2010, 2010, 802078. [Google Scholar] [CrossRef]
- Marshall, J.A.; Hamman, R.F.; Baxter, J. High-Fat, Low-Carbohydrate Diet and the Etiology of Non-Insulin-Dependent Diabetes Mellitus: The San Luis Valley Diabetes Study. Am. J. Epidemiol. 1991, 134, 590–603. [Google Scholar] [CrossRef]
- Nakamura, Y.K.; Omaye, S.T. Metabolic Diseases and Pro- and Prebiotics: Mechanistic Insights. Nutr. Metab. 2012, 9, 60. [Google Scholar] [CrossRef]
- Manzel, A.; Muller, D.N.; Hafler, D.A.; Erdman, S.E.; Linker, R.A.; Kleinewietfeld, M. Role of “Western Diet” in Inflammatory Autoimmune Diseases. Curr. Allergy Asthma Rep. 2014, 14, 404. [Google Scholar] [CrossRef]
- Canella, D.S.; Levy, R.B.; Martins, A.P.B.; Claro, R.M.; Moubarac, J.-C.; Baraldi, L.G.; Cannon, G.; Monteiro, C.A. Ultra-Processed Food Products and Obesity in Brazilian Households (2008–2009). PLoS ONE 2014, 9, e92752. [Google Scholar] [CrossRef]
- Aune, D.; Norat, T.; Romundstad, P.; Vatten, L.J. Whole Grain and Refined Grain Consumption and the Risk of Type 2 Diabetes: A Systematic Review and Dose-Response Meta-Analysis of Cohort Studies. Eur. J. Epidemiol. 2013, 28, 845–858. [Google Scholar] [CrossRef]
- Tørris, C.; Molin, M.; Småstuen, M.C. Lean Fish Consumption Is Associated with Beneficial Changes in the Metabolic Syndrome Components: A 13-Year Follow-Up Study from the Norwegian Tromsø Study. Nutrients 2017, 9, 247. [Google Scholar] [CrossRef] [PubMed]
- Di Daniele, N.; Noce, A.; Vidiri, M.F.; Moriconi, E.; Marrone, G.; Annicchiarico-Petruzzelli, M.; D’Urso, G.; Tesauro, M.; Rovella, V.; De Lorenzo, A. Impact of Mediterranean Diet on Metabolic Syndrome, Cancer and Longevity. Oncotarget 2017, 8, 8947–8979. [Google Scholar] [CrossRef] [PubMed]
- De Lorenzo, A.; Petroni, M.L.; De Luca, P.P.; Andreoli, A.; Morini, P.; Iacopino, L.; Innocente, I.; Perriello, G. Use of Quality Control Indices in Moderately Hypocaloric Mediterranean Diet for Treatment of Obesity. Diabetes Nutr. Metab. 2001, 14, 181–188. [Google Scholar] [PubMed]
- Pitsavos, C.; Panagiotakos, D.B.; Tzima, N.; Chrysohoou, C.; Economou, M.; Zampelas, A.; Stefanadis, C. Adherence to the Mediterranean Diet Is Associated with Total Antioxidant Capacity in Healthy Adults: The ATTICA Study. Am. J. Clin. Nutr. 2005, 82, 694–699. [Google Scholar] [CrossRef] [PubMed]
- Christ, A.; Latz, E. The Western Lifestyle Has Lasting Effects on Metaflammation. Nat. Rev. Immunol. 2019, 19, 267–268. [Google Scholar] [CrossRef]
- Kratz, M.; Baars, T.; Guyenet, S. The Relationship between High-Fat Dairy Consumption and Obesity, Cardiovascular, and Metabolic Disease. Eur. J. Nutr. 2013, 52, 1–24. [Google Scholar] [CrossRef]
- Giugliano, D.; Ceriello, A.; Esposito, K. The Effects of Diet on Inflammation: Emphasis on the Metabolic Syndrome. J. Am. Coll. Cardiol. 2006, 48, 677–685. [Google Scholar] [CrossRef]
- de Souza, R.J.; Mente, A.; Maroleanu, A.; Cozma, A.I.; Ha, V.; Kishibe, T.; Uleryk, E.; Budylowski, P.; Schünemann, H.; Beyene, J.; et al. Intake of Saturated and Trans Unsaturated Fatty Acids and Risk of All Cause Mortality, Cardiovascular Disease, and Type 2 Diabetes: Systematic Review and Meta-Analysis of Observational Studies. BMJ 2015, 351, h3978. [Google Scholar] [CrossRef]
- Sami, W.; Ansari, T.; Butt, N.S.; Hamid, M.R.A. Effect of Diet on Type 2 Diabetes Mellitus: A Review. Int. J. Health Sci. (Qassim) 2017, 11, 65–71. [Google Scholar] [PubMed]
- Feldeisen, S.E.; Tucker, K.L. Nutritional Strategies in the Prevention and Treatment of Metabolic Syndrome. Appl. Physiol. Nutr. Metab. 2007, 32, 46–60. [Google Scholar] [CrossRef] [PubMed]
- Marrone, G.; Guerriero, C.; Palazzetti, D.; Lido, P.; Marolla, A.; Di Daniele, F.; Noce, A. Vegan Diet Health Benefits in Metabolic Syndrome. Nutrients 2021, 13, 817. [Google Scholar] [CrossRef] [PubMed]
- Chiu, Y.-F.; Hsu, C.-C.; Chiu, T.H.T.; Lee, C.-Y.; Liu, T.-T.; Tsao, C.K.; Chuang, S.-C.; Hsiung, C.A. Cross-Sectional and Longitudinal Comparisons of Metabolic Profiles between Vegetarian and Non-Vegetarian Subjects: A Matched Cohort Study. Br. J. Nutr. 2015, 114, 1313–1320. [Google Scholar] [CrossRef]
- Chiang, J.-K.; Lin, Y.-L.; Chen, C.-L.; Ouyang, C.-M.; Wu, Y.-T.; Chi, Y.-C.; Huang, K.-C.; Yang, W.-S. Reduced Risk for Metabolic Syndrome and Insulin Resistance Associated with Ovo-Lacto-Vegetarian Behavior in Female Buddhists: A Case-Control Study. PLoS ONE 2013, 8, e71799. [Google Scholar] [CrossRef] [PubMed]
- Tørris, C.; Småstuen, M.C.; Molin, M. Nutrients in Fish and Possible Associations with Cardiovascular Disease Risk Factors in Metabolic Syndrome. Nutrients 2018, 10, 952. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.S.; Calle, M.; Fernandez, M.L. Healthy Plant-Based Diets Improve Dyslipidemias, Insulin Resistance, and Inflammation in Metabolic Syndrome. A Narrative Review. Adv. Nutr. 2023, 14, 44–54. [Google Scholar] [CrossRef]
- Kim, D.-H.; Kim, H.; Jeong, D.; Kang, I.-B.; Chon, J.-W.; Kim, H.-S.; Song, K.-Y.; Seo, K.-H. Kefir Alleviates Obesity and Hepatic Steatosis in High-Fat Diet-Fed Mice by Modulation of Gut Microbiota and Mycobiota: Targeted and Untargeted Community Analysis with Correlation of Biomarkers. J. Nutr. Biochem. 2017, 44, 35–43. [Google Scholar] [CrossRef]
- Unno, T.; Choi, J.-H.; Hur, H.-G.; Sadowsky, M.J.; Ahn, Y.-T.; Huh, C.-S.; Kim, G.-B.; Cha, C.-J. Changes in Human Gut Microbiota Influenced by Probiotic Fermented Milk Ingestion. J. Dairy Sci. 2015, 98, 3568–3576. [Google Scholar] [CrossRef]
- Ménard, S.; Candalh, C.; Bambou, J.C.; Terpend, K.; Cerf-Bensussan, N.; Heyman, M. Lactic Acid Bacteria Secrete Metabolites Retaining Anti-Inflammatory Properties after Intestinal Transport. Gut 2004, 53, 821–828. [Google Scholar] [CrossRef]
- Green, M.; Arora, K.; Prakash, S. Microbial Medicine: Prebiotic and Probiotic Functional Foods to Target Obesity and Metabolic Syndrome. Int. J. Mol. Sci. 2020, 21, 2890. [Google Scholar] [CrossRef]
- van der Wijst, M.G.P.; de Vries, D.H.; Groot, H.E.; Trynka, G.; Hon, C.-C.; Nawijn, M.C.; Idaghdour, Y.; van der Harst, P.; Ye, C.J.; Powell, J. Single-Cell EQTLGen Consortium: A Personalized Understanding of Disease. arXiv 2019, arXiv:arXiv1909.12550. [Google Scholar]
- Ding, S.; Chen, X.; Shen, K. Single-Cell RNA Sequencing in Breast Cancer: Understanding Tumor Heterogeneity and Paving Roads to Individualized Therapy. Cancer Commun. 2020, 40, 329–344. [Google Scholar] [CrossRef] [PubMed]
- Hrovatin, K.; Fischer, D.S.; Theis, F.J. Toward Modeling Metabolic State from Single-Cell Transcriptomics. Mol. Metab. 2022, 57, 101396. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, H.J.R.; Patikas, N.; Foskolou, S.; Field, S.F.; Park, J.-E.; Byrne, M.L.; Bassett, A.R.; Metzakopian, E. Single-Cell Transcriptomics of Parkinson’s Disease Human In Vitro Models Reveals Dopamine Neuron-Specific Stress Responses. Cell Rep. 2020, 33, 108263. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, Q.; Wu, D.; Chen, G. Exploring Additional Valuable Information from Single-Cell RNA-Seq Data. Front. Cell Dev. Biol. 2020, 8, 593007. [Google Scholar] [CrossRef]
- Wagner, A.; Regev, A.; Yosef, N. Revealing the Vectors of Cellular Identity with Single-Cell Genomics. Nat. Biotechnol. 2016, 34, 1145–1160. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-S.; Pinto, S.M.; Getnet, D.; Nirujogi, R.S.; Manda, S.S.; Chaerkady, R.; Madugundu, A.K.; Kelkar, D.S.; Isserlin, R.; Jain, S.; et al. A Draft Map of the Human Proteome. Nature 2014, 509, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.J.; Schug, J.; Won, K.-J.; Liu, C.; Naji, A.; Avrahami, D.; Golson, M.L.; Kaestner, K.H. Single-Cell Transcriptomics of the Human Endocrine Pancreas. Diabetes 2016, 65, 3028–3038. [Google Scholar] [CrossRef]
- Biddy, B.A.; Kong, W.; Kamimoto, K.; Guo, C.; Waye, S.E.; Sun, T.; Morris, S.A. Single-Cell Mapping of Lineage and Identity in Direct Reprogramming. Nature 2018, 564, 219–224. [Google Scholar] [CrossRef]
- Jaitin, D.A.; Adlung, L.; Thaiss, C.A.; Weiner, A.; Li, B.; Descamps, H.; Lundgren, P.; Bleriot, C.; Liu, Z.; Deczkowska, A.; et al. Lipid-Associated Macrophages Control Metabolic Homeostasis in a Trem2-Dependent Manner. Cell 2019, 178, 686–698.e14. [Google Scholar] [CrossRef]
- Su, Q.; Kim, S.Y.; Adewale, F.; Zhou, Y.; Aldler, C.; Ni, M.; Wei, Y.; Burczynski, M.E.; Atwal, G.S.; Sleeman, M.W.; et al. Single-Cell RNA Transcriptome Landscape of Hepatocytes and Non-Parenchymal Cells in Healthy and NAFLD Mouse Liver. iScience 2021, 24, 103233. [Google Scholar] [CrossRef]
- Vickers, M.H. Early Life Nutrition, Epigenetics and Programming of Later Life Disease. Nutrients 2014, 6, 2165–2178. [Google Scholar] [CrossRef]
- Wang, Y.J.; Kaestner, K.H. Single-Cell RNA-Seq of the Pancreatic Islets—A Promise Not yet Fulfilled? Cell Metab. 2019, 29, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Ruebel, M.L.; Cotter, M.; Sims, C.R.; Moutos, D.M.; Badger, T.M.; Cleves, M.A.; Shankar, K.; Andres, A. Obesity Modulates Inflammation and Lipid Metabolism Oocyte Gene Expression: A Single-Cell Transcriptome Perspective. J. Clin. Endocrinol. Metab. 2017, 102, 2029–2038. [Google Scholar] [CrossRef] [PubMed]
- Burska, A.N.; Roget, K.; Blits, M.; Soto Gomez, L.; van de Loo, F.; Hazelwood, L.D.; Verweij, C.L.; Rowe, A.; Goulielmos, G.N.; van Baarsen, L.G.M.; et al. Gene Expression Analysis in RA: Towards Personalized Medicine. Pharmacogenom. J. 2014, 14, 93–106. [Google Scholar] [CrossRef]
- Bao, K.; Cui, Z.; Wang, H.; Xiao, H.; Li, T.; Kong, X.; Liu, T. Pseudotime Ordering Single-Cell Transcriptomic of β Cells Pancreatic Islets in Health and Type 2 Diabetes. Phenomics 2021, 1, 199–210. [Google Scholar] [CrossRef]
- Shulse, C.N.; Cole, B.J.; Ciobanu, D.; Lin, J.; Yoshinaga, Y.; Gouran, M.; Turco, G.M.; Zhu, Y.; O’Malley, R.C.; Brady, S.M.; et al. High-Throughput Single-Cell Transcriptome Profiling of Plant Cell Types. Cell Rep. 2019, 27, 2241–2247.e4. [Google Scholar] [CrossRef]
- Shreiner, A.B.; Kao, J.Y.; Young, V.B. The Gut Microbiome in Health and in Disease. Curr. Opin. Gastroenterol. 2015, 31, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Cerdá, B.; Pérez, M.; Pérez-Santiago, J.D.; Tornero-Aguilera, J.F.; González-Soltero, R.; Larrosa, M. Gut Microbiota Modification: Another Piece in the Puzzle of the Benefits of Physical Exercise in Health? Front. Physiol. 2016, 7, 51. [Google Scholar] [CrossRef]
- Vijay, A.; Valdes, A.M. Role of the Gut Microbiome in Chronic Diseases: A Narrative Review. Eur. J. Clin. Nutr. 2022, 76, 489–501. [Google Scholar] [CrossRef]
- Hou, H.; Chen, D.; Zhang, K.; Zhang, W.; Liu, T.; Wang, S.; Dai, X.; Wang, B.; Zhong, W.; Cao, H. Gut Microbiota-Derived Short-Chain Fatty Acids and Colorectal Cancer: Ready for Clinical Translation? Cancer Lett. 2022, 526, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Sankararaman, S.; Noriega, K.; Velayuthan, S.; Sferra, T.; Martindale, R. Gut Microbiome and Its Impact on Obesity and Obesity-Related Disorders. Curr. Gastroenterol. Rep. 2023, 25, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Hales, C.M.; Carroll, M.D.; Fryar, C.D.; Ogden, C.L. Prevalence of Obesity among Adults and Youth: United States, 2015–2016; NCHS Data Brief; No. 288; U.S. Department OF Health and Human Services: Washington, DC, USA, 2017; pp. 1–8. [Google Scholar]
- Dabke, K.; Hendrick, G.; Devkota, S. The Gut Microbiome and Metabolic Syndrome. J. Clin. Investig. 2019, 129, 4050–4057. [Google Scholar] [CrossRef]
- Stanford, J.; Charlton, K.; Stefoska-Needham, A.; Zheng, H.; Bird, L.; Borst, A.; Fuller, A.; Lambert, K. Associations Among Plant-Based Diet Quality, Uremic Toxins, and Gut Microbiota Profile in Adults Undergoing Hemodialysis Therapy. J. Ren. Nutr. Off. J. Counc. Ren. Nutr. Natl. Kidney Found. 2021, 31, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Raoul, P.; Cintoni, M.; Palombaro, M.; Basso, L.; Rinninella, E.; Gasbarrini, A.; Mele, M.C. Food Additives, a Key Environmental Factor in the Development of IBD through Gut Dysbiosis. Microorganisms 2022, 10, 167. [Google Scholar] [CrossRef] [PubMed]
- Eslami, M.; Bahar, A.; Hemati, M.; Rasouli Nejad, Z.; Mehranfar, F.; Karami, S.; Kobyliak, N.M.; Yousefi, B. Dietary Pattern, Colonic Microbiota and Immunometabolism Interaction: New Frontiers for Diabetes Mellitus and Related Disorders. Diabet. Med. 2021, 38, e14415. [Google Scholar] [CrossRef]
- Ma, X.; Kong, Y.; Xu, H.; Bi, Q.; Liang, M.; Mai, K.; Zhang, Y. Short-Term Alternate Feeding between Terrestrially Sourced Oil-and Fish Oil-Based Diets Modulates the Intestinal Microecology of Juvenile Turbot. Biology 2023, 12, 650. [Google Scholar] [CrossRef]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia Muciniphila in Overweight and Obese Human Volunteers: A Proof-of-Concept Exploratory Study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef]
- Weitkunat, K.; Stuhlmann, C.; Postel, A.; Rumberger, S.; Fankhänel, M.; Woting, A.; Petzke, K.J.; Gohlke, S.; Schulz, T.J.; Blaut, M.; et al. Short-Chain Fatty Acids and Inulin, but Not Guar Gum, Prevent Diet-Induced Obesity and Insulin Resistance through Differential Mechanisms in Mice. Sci. Rep. 2017, 7, 6109. [Google Scholar] [CrossRef]
- Zinöcker, M.K.; Lindseth, I.A. The Western Diet-Microbiome-Host Interaction and Its Role in Metabolic Disease. Nutrients 2018, 10, 365. [Google Scholar] [CrossRef]
- Finger, A.-M.; Kramer, A. Mammalian Circadian Systems: Organization and Modern Life Challenges. Acta Physiol. 2021, 231, e13548. [Google Scholar] [CrossRef]
- Kaczmarek, J.L.; Thompson, S.V.; Holscher, H.D. Complex Interactions of Circadian Rhythms, Eating Behaviors, and the Gastrointestinal Microbiota and Their Potential Impact on Health. Nutr. Rev. 2017, 75, 673–682. [Google Scholar] [CrossRef]
- Leone, V.; Gibbons, S.M.; Martinez, K.; Hutchison, A.L.; Huang, E.Y.; Cham, C.M.; Pierre, J.F.; Heneghan, A.F.; Nadimpalli, A.; Hubert, N.; et al. Effects of Diurnal Variation of Gut Microbes and High-Fat Feeding on Host Circadian Clock Function and Metabolism. Cell Host Microbe 2015, 17, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Thaiss, C.A.; Levy, M.; Grosheva, I.; Zheng, D.; Soffer, E.; Blacher, E.; Braverman, S.; Tengeler, A.C.; Barak, O.; Elazar, M.; et al. Hyperglycemia Drives Intestinal Barrier Dysfunction and Risk for Enteric Infection. Science 2018, 359, 1376–1383. [Google Scholar] [CrossRef]
- Kopin, L.; Lowenstein, C. Dyslipidemia. Ann. Intern. Med. 2017, 167, ITC81–ITC96. [Google Scholar] [CrossRef]
- Xiao, S.; Zhang, Z.; Chen, M.; Zou, J.; Jiang, S.; Qian, D.; Duan, J. Xiexin Tang Ameliorates Dyslipidemia in High-Fat Diet-Induced Obese Rats via Elevating Gut Microbiota-Derived Short Chain Fatty Acids Production and Adjusting Energy Metabolism. J. Ethnopharmacol. 2019, 241, 112032. [Google Scholar] [CrossRef] [PubMed]
- Ambroselli, D.; Masciulli, F.; Romano, E.; Catanzaro, G.; Besharat, Z.M.; Massari, M.C.; Ferretti, E.; Migliaccio, S.; Izzo, L.; Ritieni, A.; et al. New Advances in Metabolic Syndrome, from Prevention to Treatment: The Role of Diet and Food. Nutrients 2023, 15, 640. [Google Scholar] [CrossRef]
- Onrust, L.; Ducatelle, R.; Van Driessche, K.; De Maesschalck, C.; Vermeulen, K.; Haesebrouck, F.; Eeckhaut, V.; Van Immerseel, F. Steering Endogenous Butyrate Production in the Intestinal Tract of Broilers as a Tool to Improve Gut Health. Front. Vet. Sci. 2015, 2, 75. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, C.; Jiang, Q.; Yin, Y. Butyrate in Energy Metabolism: There Is Still More to Learn. Trends Endocrinol. Metab. 2021, 32, 159–169. [Google Scholar] [CrossRef]
- Schoeler, M.; Caesar, R. Dietary Lipids, Gut Microbiota and Lipid Metabolism. Rev. Endocr. Metab. Disord. 2019, 20, 461–472. [Google Scholar] [CrossRef]
- Xu, H.; Wang, X.; Feng, W.; Liu, Q.; Zhou, S.; Liu, Q.; Cai, L. The Gut Microbiota and Its Interactions with Cardiovascular Disease. Microb. Biotechnol. 2020, 13, 637–656. [Google Scholar] [CrossRef] [PubMed]
- Cheru, L.; Saylor, C.F.; Lo, J. Gastrointestinal Barrier Breakdown and Adipose Tissue Inflammation. Curr. Obes. Rep. 2019, 8, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Cinti, S.; Mitchell, G.; Barbatelli, G.; Murano, I.; Ceresi, E.; Faloia, E.; Wang, S.; Fortier, M.; Greenberg, A.S.; Obin, M.S. Adipocyte Death Defines Macrophage Localization and Function in Adipose Tissue of Obese Mice and Humans. J. Lipid Res. 2005, 46, 2347–2355. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Yuan, M.; Frantz, D.F.; Melendez, P.A.; Hansen, L.; Lee, J.; Shoelson, S.E. Local and Systemic Insulin Resistance Resulting from Hepatic Activation of IKK-Beta and NF-KappaB. Nat. Med. 2005, 11, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Kamada, N.; Chen, G.Y.; Inohara, N.; Núñez, G. Control of Pathogens and Pathobionts by the Gut Microbiota. Nat. Immunol. 2013, 14, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Anders, H.-J.; Andersen, K.; Stecher, B. The Intestinal Microbiota, a Leaky Gut, and Abnormal Immunity in Kidney Disease. Kidney Int. 2013, 83, 1010–1016. [Google Scholar] [CrossRef] [PubMed]
- Motta, J.-P.; Wallace, J.L.; Buret, A.G.; Deraison, C.; Vergnolle, N. Gastrointestinal Biofilms in Health and Disease. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 314–334. [Google Scholar] [CrossRef]
- Geurts, L.; Lazarevic, V.; Derrien, M.; Everard, A.; Van Roye, M.; Knauf, C.; Valet, P.; Girard, M.; Muccioli, G.G.; François, P.; et al. Altered Gut Microbiota and Endocannabinoid System Tone in Obese and Diabetic Leptin-Resistant Mice: Impact on Apelin Regulation in Adipose Tissue. Front. Microbiol. 2011, 2, 149. [Google Scholar] [CrossRef]
- Kang, C.; Wang, B.; Kaliannan, K.; Wang, X.; Lang, H.; Hui, S.; Huang, L.; Zhang, Y.; Zhou, M.; Chen, M.; et al. Gut Microbiota Mediates the Protective Effects of Dietary Capsaicin against Chronic Low-Grade Inflammation and Associated Obesity Induced by High-Fat Diet. MBio 2017, 8, e00470-17. [Google Scholar] [CrossRef]
- Thaiss, C.A.; Levy, M.; Korem, T.; Dohnalová, L.; Shapiro, H.; Jaitin, D.A.; David, E.; Winter, D.R.; Gury-BenAri, M.; Tatirovsky, E.; et al. Microbiota Diurnal Rhythmicity Programs Host Transcriptome Oscillations. Cell 2016, 167, 1495–1510.e12. [Google Scholar] [CrossRef]
- Quraishi, M.N.; Widlak, M.; Bhala, N.; Moore, D.; Price, M.; Sharma, N.; Iqbal, T.H. Systematic Review with Meta-Analysis: The Efficacy of Faecal Microbiota Transplantation for the Treatment of Recurrent and Refractory Clostridium Difficile Infection. Aliment. Pharmacol. Ther. 2017, 46, 479–493. [Google Scholar] [CrossRef]
- de Groot, P.F.; Frissen, M.N.; de Clercq, N.C.; Nieuwdorp, M. Fecal Microbiota Transplantation in Metabolic Syndrome: History, Present and Future. Gut Microbes 2017, 8, 253–267. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The Gut Microbiota as an Environmental Factor That Regulates Fat Storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef] [PubMed]
- Proença, I.M.; Allegretti, J.R.; Bernardo, W.M.; de Moura, D.T.H.; Ponte Neto, A.M.; Matsubayashi, C.O.; Flor, M.M.; Kotinda, A.P.S.T.; de Moura, E.G.H. Fecal Microbiota Transplantation Improves Metabolic Syndrome Parameters: Systematic Review with Meta-Analysis Based on Randomized Clinical Trials. Nutr. Res. 2020, 83, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Turroni, S.; Magnani, M.; Kc, P.; Lesnik, P.; Vidal, H.; Heer, M. Gut Microbiome and Space Travelers’ Health: State of the Art and Possible Pro/Prebiotic Strategies for Long-Term Space Missions. Front. Physiol. 2020, 11, 553929. [Google Scholar] [CrossRef] [PubMed]
- Ozougwu, J.C.; Obimba, K.C.; Belonwu, C.D.; Unakalamba, C.B. The Pathogenesis and Pathophysiology of Type 1 and Type 2 Diabetes Mellitus. J. Physiol. Pathophysiol. 2013, 4, 46–57. [Google Scholar] [CrossRef]
- Al Aboud, N.M.; Tupper, C.; Jialal, I. Genetics, Epigenetic Mechanism; StatPearls: Treasure Island, FL, USA, 2023. [Google Scholar]
- Riva, P.; Ratti, A.; Venturin, M. The Long Non-Coding RNAs in Neurodegenerative Diseases: Novel Mechanisms of Pathogenesis. Curr. Alzheimer Res. 2016, 13, 1219–1231. [Google Scholar] [CrossRef]
- Abdul, Q.A.; Yu, B.P.; Chung, H.Y.; Jung, H.A.; Choi, J.S. Epigenetic Modifications of Gene Expression by Lifestyle and Environment. Arch. Pharm. Res. 2017, 40, 1219–1237. [Google Scholar] [CrossRef]
- Zheng, J.; Xiao, X.; Zhang, Q.; Yu, M. DNA Methylation: The Pivotal Interaction between Early-Life Nutrition and Glucose Metabolism in Later Life. Br. J. Nutr. 2014, 112, 1850–1857. [Google Scholar] [CrossRef] [PubMed]
- Biemann, R.; Blüher, M.; Isermann, B. Exposure to Endocrine-Disrupting Compounds Such as Phthalates and Bisphenol A Is Associated with an Increased Risk for Obesity. Best Pract. Res. Clin. Endocrinol. Metab. 2021, 35, 101546. [Google Scholar] [CrossRef] [PubMed]
- Barres, R.; Kirchner, H.; Rasmussen, M.; Yan, J.; Kantor, F.R.; Krook, A.; Näslund, E.; Zierath, J.R. Weight Loss after Gastric Bypass Surgery in Human Obesity Remodels Promoter Methylation. Cell Rep. 2013, 3, 1020–1027. [Google Scholar] [CrossRef]
- Rönn, T.; Volkov, P.; Davegårdh, C.; Dayeh, T.; Hall, E.; Olsson, A.H.; Nilsson, E.; Tornberg, A.; Dekker Nitert, M.; Eriksson, K.-F.; et al. A Six Months Exercise Intervention Influences the Genome-Wide DNA Methylation Pattern in Human Adipose Tissue. PLoS Genet. 2013, 9, e1003572. [Google Scholar] [CrossRef] [PubMed]
- Pinnick, K.E.; Karpe, F. DNA Methylation of Genes in Adipose Tissue. Proc. Nutr. Soc. 2011, 70, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and Chemokines: At the Crossroads of Cell Signalling and Inflammatory Disease. Biochim. Biophys. Acta 2014, 1843, 2563–2582. [Google Scholar] [CrossRef] [PubMed]
- Adcock, I.M.; Caramori, G.; Barnes, P.J. Chronic Obstructive Pulmonary Disease and Lung Cancer: New Molecular Insights. Respiration 2011, 81, 265–284. [Google Scholar] [CrossRef] [PubMed]
- Grabiec, A.M.; Korchynskyi, O.; Tak, P.P.; Reedquist, K.A. Histone Deacetylase Inhibitors Suppress Rheumatoid Arthritis Fibroblast-like Synoviocyte and Macrophage IL-6 Production by Accelerating MRNA Decay. Ann. Rheum. Dis. 2012, 71, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Castro, L.; Freeman, B.A. Reactive Oxygen Species in Human Health and Disease. Nutrition 2001, 17, 161+163–165. [Google Scholar] [CrossRef]
- Hosseini, H.; Teimouri, M.; Shabani, M.; Koushki, M.; Babaei Khorzoughi, R.; Namvarjah, F.; Izadi, P.; Meshkani, R. Resveratrol Alleviates Non-Alcoholic Fatty Liver Disease through Epigenetic Modification of the Nrf2 Signaling Pathway. Int. J. Biochem. Cell Biol. 2020, 119, 105667. [Google Scholar] [CrossRef]
- Yi, X.; Zhu, Q.-X.; Wu, X.-L.; Tan, T.-T.; Jiang, X.-J. Histone Methylation and Oxidative Stress in Cardiovascular Diseases. Oxid. Med. Cell. Longev. 2022, 2022, 6023710. [Google Scholar] [CrossRef]
- Kowluru, R.A.; Mishra, M. Oxidative Stress, Mitochondrial Damage and Diabetic Retinopathy. Biochim. Biophys. Acta 2015, 1852, 2474–2483. [Google Scholar] [CrossRef]
- Gallou-Kabani, C.; Vigé, A.; Gross, M.-S.; Boileau, C.; Rabes, J.-P.; Fruchart-Najib, J.; Jais, J.-P.; Junien, C. Resistance to High-Fat Diet in the Female Progeny of Obese Mice Fed a Control Diet during the Periconceptual, Gestation, and Lactation Periods. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E1095–E1100. [Google Scholar] [CrossRef] [PubMed]
- Shan, Q.; Zheng, G.; Zhu, A.; Cao, L.; Lu, J.; Wu, D.; Zhang, Z.; Fan, S.; Sun, C.; Hu, B.; et al. Epigenetic Modification of MiR-10a Regulates Renal Damage by Targeting CREB1 in Type 2 Diabetes Mellitus. Toxicol. Appl. Pharmacol. 2016, 306, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Surendran, S.; Vimaleswaran, K.S. A Nutrigenetic Approach to Examine the Relationship between Vitamin B12 Status and Cardio-metabolic Traits in Multiple Ethnic Groups–Findings from the GeNuIne Collaboration. Nutr. Bull. 2021, 46, 185–194. [Google Scholar] [CrossRef]
- Lancaster, E.E.; Vladimirov, V.I.; Riley, B.P.; Landry, J.W.; Roberson-Nay, R.; York, T.P. Large-Scale Integration of DNA Methylation and Gene Expression Array Platforms Identifies Both Cis and Trans Relationships. Epigenetics 2022, 17, 1753–1773. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Yin, J.; Zhang, J.; Ward, R.E.; Martin, R.J.; Lefevre, M.; Cefalu, W.T.; Ye, J. Butyrate Improves Insulin Sensitivity and Increases Energy Expenditure in Mice. Diabetes 2009, 58, 1509–1517. [Google Scholar] [CrossRef]
- D’Aquila, P.; Carelli, L.L.; De Rango, F.; Passarino, G.; Bellizzi, D. Gut Microbiota as Important Mediator Between Diet and DNA Methylation and Histone Modifications in the Host. Nutrients 2020, 12, 597. [Google Scholar] [CrossRef]
- Dick, K.J.; Nelson, C.P.; Tsaprouni, L.; Sandling, J.K.; Aïssi, D.; Wahl, S.; Meduri, E.; Morange, P.-E.; Gagnon, F.; Grallert, H.; et al. DNA Methylation and Body-Mass Index: A Genome-Wide Analysis. Lancet 2014, 383, 1990–1998. [Google Scholar] [CrossRef]
- Maiese, K.; Morhan, S.D.; Chong, Z.Z. Oxidative Stress Biology and Cell Injury during Type 1 and Type 2 Diabetes Mellitus. Curr. Neurovasc. Res. 2007, 4, 63–71. [Google Scholar] [CrossRef]
- Li, C.; Ni, Y.-Q.; Xu, H.; Xiang, Q.-Y.; Zhao, Y.; Zhan, J.-K.; He, J.-Y.; Li, S.; Liu, Y.-S. Roles and Mechanisms of Exosomal Non-Coding RNAs in Human Health and Diseases. Signal Transduct. Target. Ther. 2021, 6, 383. [Google Scholar] [CrossRef]
- Mthembu, S.X.H.; Mazibuko-Mbeje, S.E.; Ziqubu, K.; Nyawo, T.A.; Obonye, N.; Nyambuya, T.M.; Nkambule, B.B.; Silvestri, S.; Tiano, L.; Muller, C.J.F.; et al. Impact of Physical Exercise and Caloric Restriction in Patients with Type 2 Diabetes: Skeletal Muscle Insulin Resistance and Mitochondrial Dysfunction as Ideal Therapeutic Targets. Life Sci. 2022, 297, 120467. [Google Scholar] [CrossRef]
- Storniolo, C.E.; Casillas, R.; Bulló, M.; Castañer, O.; Ros, E.; Sáez, G.T.; Toledo, E.; Estruch, R.; Ruiz-Gutiérrez, V.; Fitó, M.; et al. A Mediterranean Diet Supplemented with Extra Virgin Olive Oil or Nuts Improves Endothelial Markers Involved in Blood Pressure Control in Hypertensive Women. Eur. J. Nutr. 2017, 56, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Heerboth, S.; Lapinska, K.; Snyder, N.; Leary, M.; Rollinson, S.; Sarkar, S. Use of Epigenetic Drugs in Disease: An Overview. Genet. Epigenet. 2014, 6, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Catana, C.; Guimaraes, A.R.; Rosen, B.R. PET and MR Imaging: The Odd Couple or a Match Made in Heaven? J. Nucl. Med. 2013, 54, 815–824. [Google Scholar] [CrossRef]
- Berg, E.; Cherry, S.R. Innovations in Instrumentation for Positron Emission Tomography. Semin. Nucl. Med. 2018, 48, 311–331. [Google Scholar] [CrossRef] [PubMed]
- Jones, T.; Townsend, D. History and Future Technical Innovation in Positron Emission Tomography. J. Med. Imaging 2017, 4, 11013. [Google Scholar] [CrossRef]
- Eriksson, O.; Laughlin, M.; Brom, M.; Nuutila, P.; Roden, M.; Hwa, A.; Bonadonna, R.; Gotthardt, M. In Vivo Imaging of Beta Cells with Radiotracers: State of the Art, Prospects and Recommendations for Development and Use. Diabetologia 2016, 59, 1340–1349. [Google Scholar] [CrossRef]
- Jodal, A.; Schibli, R.; Béhé, M. Targets and Probes for Non-Invasive Imaging of β-Cells. Eur. J. Nucl. Med. Mol. Imaging 2017, 44, 712–727. [Google Scholar] [CrossRef]
- Goel, S.; England, C.G.; Chen, F.; Cai, W. Positron Emission Tomography and Nanotechnology: A Dynamic Duo for Cancer Theranostics. Adv. Drug Deliv. Rev. 2017, 113, 157–176. [Google Scholar] [CrossRef]
- Culver, J.; Akers, W.; Achilefu, S. Multimodality Molecular Imaging with Combined Optical and SPECT/PET Modalities. J. Nucl. Med. 2008, 49, 169–172. [Google Scholar] [CrossRef]
- Ehlerding, E.B.; Cai, W. Harnessing the Power of Molecular Imaging for Precision Medicine. J. Nucl. Med. 2016, 57, 171–172. [Google Scholar] [CrossRef]
- Gotthardt, M.; Eizirik, D.L.; Cnop, M.; Brom, M. Beta Cell Imaging—A Key Tool in Optimized Diabetes Prevention and Treatment. Trends Endocrinol. Metab. 2014, 25, 375–377. [Google Scholar] [CrossRef] [PubMed]
- DiMeglio, L.A.; Evans-Molina, C.; Oram, R.A. Type 1 Diabetes. Lancet 2018, 391, 2449–2462. [Google Scholar] [CrossRef]
- Wei, W.; Ehlerding, E.B.; Lan, X.; Luo, Q.-Y.; Cai, W. Molecular Imaging of β-Cells: Diabetes and Beyond. Adv. Drug Deliv. Rev. 2019, 139, 16–31. [Google Scholar] [CrossRef] [PubMed]
- Matveyenko, A.V.; Butler, P.C. Relationship between Beta-Cell Mass and Diabetes Onset. Diabetes Obes. Metab. 2008, 10 (Suppl. S4), 23–31. [Google Scholar] [CrossRef] [PubMed]
- Butler, A.E.; Janson, J.; Bonner-Weir, S.; Ritzel, R.; Rizza, R.A.; Butler, P.C. Beta-Cell Deficit and Increased Beta-Cell Apoptosis in Humans with Type 2 Diabetes. Diabetes 2003, 52, 102–110. [Google Scholar] [CrossRef]
- Laybutt, D.R.; Kaneto, H.; Hasenkamp, W.; Grey, S.; Jonas, J.-C.; Sgroi, D.C.; Groff, A.; Ferran, C.; Bonner-Weir, S.; Sharma, A.; et al. Increased Expression of Antioxidant and Antiapoptotic Genes in Islets That May Contribute to Beta-Cell Survival during Chronic Hyperglycemia. Diabetes 2002, 51, 413–423. [Google Scholar] [CrossRef]
- Weir, G.C.; Bonner-Weir, S. Five Stages of Evolving Beta-Cell Dysfunction during Progression to Diabetes. Diabetes 2004, 53 (Suppl. S3), S16–S21. [Google Scholar] [CrossRef]
- Di Gialleonardo, V.; de Vries, E.F.J.; Di Girolamo, M.; Quintero, A.M.; Dierckx, R.A.J.O.; Signore, A. Imaging of β-Cell Mass and Insulitis in Insulin-Dependent (Type 1) Diabetes Mellitus. Endocr. Rev. 2012, 33, 892–919. [Google Scholar] [CrossRef]
- Andralojc, K.; Srinivas, M.; Brom, M.; Joosten, L.; de Vries, I.J.M.; Eizirik, D.L.; Boerman, O.C.; Meda, P.; Gotthardt, M. Obstacles on the Way to the Clinical Visualisation of Beta Cells: Looking for the Aeneas of Molecular Imaging to Navigate between Scylla and Charybdis. Diabetologia 2012, 55, 1247–1257. [Google Scholar] [CrossRef]
- Blomberg, B.A.; Codreanu, I.; Cheng, G.; Werner, T.J.; Alavi, A. Beta-Cell Imaging: Call for Evidence-Based and Scientific Approach. Mol. Imaging Biol. 2013, 15, 123–130. [Google Scholar] [CrossRef]
- Laurent, D.; Vinet, L.; Lamprianou, S.; Daval, M.; Filhoulaud, G.; Ktorza, A.; Wang, H.; Sewing, S.; Juretschke, H.-P.; Glombik, H.; et al. Pancreatic β-Cell Imaging in Humans: Fiction or Option? Diabetes Obes. Metab. 2016, 18, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Graves, S.A.; Hernandez, R.; Valdovinos, H.F.; Ellison, P.A.; Engle, J.W.; Barnhart, T.E.; Cai, W.; Nickles, R.J. Preparation and in Vivo Characterization of 51MnCl2 as PET Tracer of Ca2+ Channel-Mediated Transport. Sci. Rep. 2017, 7, 3033. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, R.; Graves, S.A.; Gregg, T.; VanDeusen, H.R.; Fenske, R.J.; Wienkes, H.N.; England, C.G.; Valdovinos, H.F.; Jeffery, J.J.; Barnhart, T.E.; et al. Radiomanganese PET Detects Changes in Functional β-Cell Mass in Mouse Models of Diabetes. Diabetes 2017, 66, 2163–2174. [Google Scholar] [CrossRef] [PubMed]
- Maffei, A.; Liu, Z.; Witkowski, P.; Moschella, F.; Del Pozzo, G.; Liu, E.; Herold, K.; Winchester, R.J.; Hardy, M.A.; Harris, P.E. Identification of Tissue-Restricted Transcripts in Human Islets. Endocrinology 2004, 145, 4513–4521. [Google Scholar] [CrossRef] [PubMed]
- Anlauf, M.; Eissele, R.; Schäfer, M.K.-H.; Eiden, L.E.; Arnold, R.; Pauser, U.; Klöppel, G.; Weihe, E. Expression of the Two Isoforms of the Vesicular Monoamine Transporter (VMAT1 and VMAT2) in the Endocrine Pancreas and Pancreatic Endocrine Tumors. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2003, 51, 1027–1040. [Google Scholar] [CrossRef]
- Weihe, E.; Eiden, L.E. Chemical Neuroanatomy of the Vesicular Amine Transporters. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2000, 14, 2435–2449. [Google Scholar] [CrossRef]
- Freeby, M.; Goland, R.; Ichise, M.; Maffei, A.; Leibel, R.; Harris, P. VMAT2 Quantitation by PET as a Biomarker for Beta-Cell Mass in Health and Disease. Diabetes Obes. Metab. 2008, 10 (Suppl. S4), 98–108. [Google Scholar] [CrossRef]
- Inabnet, W.B.; Milone, L.; Harris, P.; Durak, E.; Freeby, M.J.; Ahmed, L.; Sebastian, M.; Lifante, J.-C.; Bessler, M.; Korner, J. The Utility of [11C] Dihydrotetrabenazine Positron Emission Tomography Scanning in Assessing β-Cell Performance after Sleeve Gastrectomy and Duodenal-Jejunal Bypass. Surgery 2010, 147, 303–309. [Google Scholar] [CrossRef]
- Goland, R.; Freeby, M.; Parsey, R.; Saisho, Y.; Kumar, D.; Simpson, N.; Hirsch, J.; Prince, M.; Maffei, A.; Mann, J.J.; et al. 11C-Dihydrotetrabenazine PET of the Pancreas in Subjects with Long-Standing Type 1 Diabetes and in Healthy Controls. J. Nucl. Med. 2009, 50, 382–389. [Google Scholar] [CrossRef]
- Shen, W.; Wang, Z.; Punyanita, M.; Lei, J.; Sinav, A.; Kral, J.G.; Imielinska, C.; Ross, R.; Heymsfield, S.B. Adipose Tissue Quantification by Imaging Methods: A Proposed Classification. Obes. Res. 2003, 11, 5–16. [Google Scholar] [CrossRef]
- THOMAS, L.W. The Chemical Composition of Adipose Tissue of Man and Mice. Q. J. Exp. Physiol. Cogn. Med. Sci. 1962, 47, 179–188. [Google Scholar] [CrossRef]
- Wang, Y.; Lobstein, T. Worldwide Trends in Childhood Overweight and Obesity. Int. J. Pediatr. Obes. IJPO Off. J. Int. Assoc. Study Obes. 2006, 1, 11–25. [Google Scholar] [CrossRef]
- Flegal, K.M.; Carroll, M.D.; Ogden, C.L.; Curtin, L.R. Prevalence and Trends in Obesity among US Adults, 1999–2008. JAMA 2010, 303, 235–241. [Google Scholar] [CrossRef]
- Borga, M.; West, J.; Bell, J.D.; Harvey, N.C.; Romu, T.; Heymsfield, S.B.; Dahlqvist Leinhard, O. Advanced Body Composition Assessment: From Body Mass Index to Body Composition Profiling. J. Investig. Med. 2018, 66, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Demerath, E.W.; Reed, D.; Rogers, N.; Sun, S.S.; Lee, M.; Choh, A.C.; Couch, W.; Czerwinski, S.A.; Chumlea, W.C.; Siervogel, R.M.; et al. Visceral Adiposity and Its Anatomical Distribution as Predictors of the Metabolic Syndrome and Cardiometabolic Risk Factor Levels. Am. J. Clin. Nutr. 2008, 88, 1263–1271. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Fox, C.S.; Hickson, D.A.; May, W.D.; Hairston, K.G.; Carr, J.J.; Taylor, H.A. Impact of Abdominal Visceral and Subcutaneous Adipose Tissue on Cardiometabolic Risk Factors: The Jackson Heart Study. J. Clin. Endocrinol. Metab. 2010, 95, 5419–5426. [Google Scholar] [CrossRef] [PubMed]
- Neeland, I.J.; Ayers, C.R.; Rohatgi, A.K.; Turer, A.T.; Berry, J.D.; Das, S.R.; Vega, G.L.; Khera, A.; McGuire, D.K.; Grundy, S.M.; et al. Associations of Visceral and Abdominal Subcutaneous Adipose Tissue with Markers of Cardiac and Metabolic Risk in Obese Adults. Obesity 2013, 21, E439–E447. [Google Scholar] [CrossRef] [PubMed]
- Iwasa, M.; Mifuji-Moroka, R.; Hara, N.; Ishidome, M.; Iwata, K.; Sugimoto, R.; Tanaka, H.; Fujita, N.; Kobayashi, Y.; Takei, Y. Visceral Fat Volume Predicts New-Onset Type 2 Diabetes in Patients with Chronic Hepatitis C. Diabetes Res. Clin. Pract. 2011, 94, 468–470. [Google Scholar] [CrossRef]
- Kurioka, S.; Murakami, Y.; Nishiki, M.; Sohmiya, M.; Koshimura, K.; Kato, Y. Relationship between Visceral Fat Accumulation and Anti-Lipolytic Action of Insulin in Patients with Type 2 Diabetes Mellitus. Endocr. J. 2002, 49, 459–464. [Google Scholar] [CrossRef]
- van der Poorten, D.; Milner, K.-L.; Hui, J.; Hodge, A.; Trenell, M.I.; Kench, J.G.; London, R.; Peduto, T.; Chisholm, D.J.; George, J. Visceral Fat: A Key Mediator of Steatohepatitis in Metabolic Liver Disease. Hepatology 2008, 48, 449–457. [Google Scholar] [CrossRef]
- Britton, K.A.; Massaro, J.M.; Murabito, J.M.; Kreger, B.E.; Hoffmann, U.; Fox, C.S. Body Fat Distribution, Incident Cardiovascular Disease, Cancer, and All-Cause Mortality. J. Am. Coll. Cardiol. 2013, 62, 921–925. [Google Scholar] [CrossRef] [PubMed]
- Doyle, S.L.; Donohoe, C.L.; Lysaght, J.; Reynolds, J.V. Visceral Obesity, Metabolic Syndrome, Insulin Resistance and Cancer. Proc. Nutr. Soc. 2012, 71, 181–189. [Google Scholar] [CrossRef]
- Ekstedt, M.; Franzén, L.E.; Mathiesen, U.L.; Thorelius, L.; Holmqvist, M.; Bodemar, G.; Kechagias, S. Long-Term Follow-up of Patients with NAFLD and Elevated Liver Enzymes. Hepatology 2006, 44, 865–873. [Google Scholar] [CrossRef]
- Goodpaster, B.H.; Kelley, D.E.; Thaete, F.L.; He, J.; Ross, R. Skeletal Muscle Attenuation Determined by Computed Tomography Is Associated with Skeletal Muscle Lipid Content. J. Appl. Physiol. 2000, 89, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Marcus, R.L.; Addison, O.; Dibble, L.E.; Foreman, K.B.; Morrell, G.; Lastayo, P. Intramuscular Adipose Tissue, Sarcopenia, and Mobility Function in Older Individuals. J. Aging Res. 2012, 2012, 629637. [Google Scholar] [CrossRef]
- Betz, M.J.; Enerbäck, S. Human Brown Adipose Tissue: What We Have Learned So Far. Diabetes 2015, 64, 2352–2360. [Google Scholar] [CrossRef] [PubMed]
- Chechi, K.; Nedergaard, J.; Richard, D. Brown Adipose Tissue as an Anti-Obesity Tissue in Humans. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2014, 15, 92–106. [Google Scholar] [CrossRef] [PubMed]
- Nedergaard, J.; Bengtsson, T.; Cannon, B. Unexpected Evidence for Active Brown Adipose Tissue in Adult Humans. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E444–E452. [Google Scholar] [CrossRef] [PubMed]
- Borga, M.; Virtanen, K.A.; Romu, T.; Leinhard, O.D.; Persson, A.; Nuutila, P.; Enerbäck, S. Brown Adipose Tissue in Humans: Detection and Functional Analysis Using PET (Positron Emission Tomography), MRI (Magnetic Resonance Imaging), and DECT (Dual Energy Computed Tomography). Methods Enzymol. 2014, 537, 141–159. [Google Scholar] [CrossRef]
- Gifford, A.; Towse, T.F.; Walker, R.C.; Avison, M.J.; Welch, E.B. Characterizing Active and Inactive Brown Adipose Tissue in Adult Humans Using PET-CT and MR Imaging. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E95–E104. [Google Scholar] [CrossRef]
- Lunati, E.; Marzola, P.; Nicolato, E.; Fedrigo, M.; Villa, M.; Sbarbati, A. In Vivo Quantitative Lipidic Map of Brown Adipose Tissue by Chemical Shift Imaging at 4.7 Tesla. J. Lipid Res. 1999, 40, 1395–1400. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.H.; Smith, D.L.J.; Nayak, K.S.; Goran, M.I.; Nagy, T.R. Identification of Brown Adipose Tissue in Mice with Fat-Water IDEAL-MRI. J. Magn. Reson. Imaging 2010, 31, 1195–1202. [Google Scholar] [CrossRef] [PubMed]
- Romu, T.; Elander, L.; Leinhard, O.D.; Lidell, M.E.; Betz, M.J.; Persson, A.; Enerbäck, S.; Borga, M. Characterization of Brown Adipose Tissue by Water-Fat Separated Magnetic Resonance Imaging. J. Magn. Reson. Imaging 2015, 42, 1639–1645. [Google Scholar] [CrossRef] [PubMed]
- Pang, H.; Luo, S.; Xiao, Y.; Xia, Y.; Li, X.; Huang, G.; Xie, Z.; Zhou, Z. Emerging Roles of Exosomes in T1DM. Front. Immunol. 2020, 11, 593348. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.-X.; Zhang, D.-Y.; Khan, M.A.; Zheng, S.-Y.; Hu, X.-M.; Zhang, Q.; Yang, R.-H.; Xiong, K. Stem Cell Transplantation in the Treatment of Type 1 Diabetes Mellitus: From Insulin Replacement to Beta-Cell Replacement. Front. Endocrinol. 2022, 13, 859638. [Google Scholar] [CrossRef] [PubMed]
- Domouky, A.M.; Hegab, A.S.; Al-Shahat, A.; Raafat, N. Mesenchymal Stem Cells and Differentiated Insulin Producing Cells Are New Horizons for Pancreatic Regeneration in Type I Diabetes Mellitus. Int. J. Biochem. Cell Biol. 2017, 87, 77–85. [Google Scholar] [CrossRef]
- Boumaza, I.; Srinivasan, S.; Witt, W.T.; Feghali-Bostwick, C.; Dai, Y.; Garcia-Ocana, A.; Feili-Hariri, M. Autologous Bone Marrow-Derived Rat Mesenchymal Stem Cells Promote PDX-1 and Insulin Expression in the Islets, Alter T Cell Cytokine Pattern and Preserve Regulatory T Cells in the Periphery and Induce Sustained Normoglycemia. J. Autoimmun. 2009, 32, 33–42. [Google Scholar] [CrossRef]
- Szot, G.L.; Yadav, M.; Lang, J.; Kroon, E.; Kerr, J.; Kadoya, K.; Brandon, E.P.; Baetge, E.E.; Bour-Jordan, H.; Bluestone, J.A. Tolerance Induction and Reversal of Diabetes in Mice Transplanted with Human Embryonic Stem Cell-Derived Pancreatic Endoderm. Cell Stem Cell 2015, 16, 148–157. [Google Scholar] [CrossRef]
- Richardson, T.; Barner, S.; Candiello, J.; Kumta, P.N.; Banerjee, I. Capsule Stiffness Regulates the Efficiency of Pancreatic Differentiation of Human Embryonic Stem Cells. Acta Biomater. 2016, 35, 153–165. [Google Scholar] [CrossRef]
- Cai, J.; Wu, Z.; Xu, X.; Liao, L.; Chen, J.; Huang, L.; Wu, W.; Luo, F.; Wu, C.; Pugliese, A.; et al. Umbilical Cord Mesenchymal Stromal Cell with Autologous Bone Marrow Cell Transplantation in Established Type 1 Diabetes: A Pilot Randomized Controlled Open-Label Clinical Study to Assess Safety and Impact on Insulin Secretion. Diabetes Care 2016, 39, 149–157. [Google Scholar] [CrossRef]
- Wen, S.-J.; Zheng, X.-M.; Liu, L.-F.; Li, N.-N.; Mao, H.-A.; Huang, L.; Yuan, Q.-L. Effects of Primary Microglia and Astrocytes on Neural Stem Cells in in Vitro and in Vivo Models of Ischemic Stroke. Neural Regen. Res. 2021, 16, 1677–1685. [Google Scholar] [CrossRef] [PubMed]
- Sultan, N.; Amin, L.E.; Zaher, A.R.; Grawish, M.E.; Scheven, B.A. Dental Pulp Stem Cells Stimulate Neuronal Differentiation of PC12 Cells. Neural Regen. Res. 2021, 16, 1821–1828. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.-M.; Li, Z.-X.; Zhang, D.-Y.; Yang, Y.-C.; Fu, S.-A.; Zhang, Z.-Q.; Yang, R.-H.; Xiong, K. A Systematic Summary of Survival and Death Signalling during the Life of Hair Follicle Stem Cells. Stem Cell Res. Ther. 2021, 12, 453. [Google Scholar] [CrossRef] [PubMed]
- Francese, R.; Fiorina, P. Immunological and Regenerative Properties of Cord Blood Stem Cells. Clin. Immunol. 2010, 136, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.-M.; Zhang, Q.; Zhou, R.-X.; Wu, Y.-L.; Li, Z.-X.; Zhang, D.-Y.; Yang, Y.-C.; Yang, R.-H.; Hu, Y.-J.; Xiong, K. Programmed Cell Death in Stem Cell-Based Therapy: Mechanisms and Clinical Applications. World J. Stem Cells 2021, 13, 386–415. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Liu, F.; Wang, J.; Chen, X.; Xie, J.; Xiong, K. Epidermal Stem Cells in Wound Healing and Their Clinical Applications. Stem Cell Res. Ther. 2019, 10, 229. [Google Scholar] [CrossRef]
- Tahbaz, M.; Yoshihara, E. Immune Protection of Stem Cell-Derived Islet Cell Therapy for Treating Diabetes. Front. Endocrinol. 2021, 12, 716625. [Google Scholar] [CrossRef]
- Buron, F.; Reffet, S.; Badet, L.; Morelon, E.; Thaunat, O. Immunological Monitoring in Beta Cell Replacement: Towards a Pathophysiology-Guided Implementation of Biomarkers. Curr. Diab. Rep. 2021, 21, 19. [Google Scholar] [CrossRef]
- Shigemoto-Kuroda, T.; Oh, J.Y.; Kim, D.-K.; Jeong, H.J.; Park, S.Y.; Lee, H.J.; Park, J.W.; Kim, T.W.; An, S.Y.; Prockop, D.J.; et al. MSC-Derived Extracellular Vesicles Attenuate Immune Responses in Two Autoimmune Murine Models: Type 1 Diabetes and Uveoretinitis. Stem Cell Rep. 2017, 8, 1214–1225. [Google Scholar] [CrossRef]
- Karnieli, O.; Izhar-Prato, Y.; Bulvik, S.; Efrat, S. Generation of Insulin-Producing Cells from Human Bone Marrow Mesenchymal Stem Cells by Genetic Manipulation. Stem Cells 2007, 25, 2837–2844. [Google Scholar] [CrossRef]
- Chen, L.-B.; Jiang, X.-B.; Yang, L. Differentiation of Rat Marrow Mesenchymal Stem Cells into Pancreatic Islet Beta-Cells. World J. Gastroenterol. 2004, 10, 3016–3020. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Wu, D.-Q.; Hu, Y.-H.; Jin, G.-X.; Li, G.-D.; Sun, T.-W.; Li, F.-J. In Vitro Cultivation of Islet-like Cell Clusters from Human Umbilical Cord Blood-Derived Mesenchymal Stem Cells. Transl. Res. 2008, 151, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Chao, K.C.; Chao, K.F.; Fu, Y.S.; Liu, S.H. Islet-like Clusters Derived from Mesenchymal Stem Cells in Wharton’s Jelly of the Human Umbilical Cord for Transplantation to Control Type 1 Diabetes. PLoS ONE 2008, 3, e1451. [Google Scholar] [CrossRef]
- Lu, J.; Shen, S.-M.; Ling, Q.; Wang, B.; Li, L.-R.; Zhang, W.; Qu, D.-D.; Bi, Y.; Zhu, D.-L. One Repeated Transplantation of Allogeneic Umbilical Cord Mesenchymal Stromal Cells in Type 1 Diabetes: An Open Parallel Controlled Clinical Study. Stem Cell Res. Ther. 2021, 12, 340. [Google Scholar] [CrossRef]
- Wang, X.; Maxwell, K.G.; Wang, K.; Bowers, D.T.; Flanders, J.A.; Liu, W.; Wang, L.-H.; Liu, Q.; Liu, C.; Naji, A.; et al. A Nanofibrous Encapsulation Device for Safe Delivery of Insulin-Producing Cells to Treat Type 1 Diabetes. Sci. Transl. Med. 2021, 13, eabb4601. [Google Scholar] [CrossRef]
- Alagpulinsa, D.A.; Cao, J.J.L.; Driscoll, R.K.; Sîrbulescu, R.F.; Penson, M.F.E.; Sremac, M.; Engquist, E.N.; Brauns, T.A.; Markmann, J.F.; Melton, D.A.; et al. Alginate-Microencapsulation of Human Stem Cell-Derived β Cells with CXCL12 Prolongs Their Survival and Function in Immunocompetent Mice without Systemic Immunosuppression. Am. J. Transplant. Off. J. Am. Soc. Transplant. Am. Soc. Transpl. Surg. 2019, 19, 1930–1940. [Google Scholar] [CrossRef]
- Xie, M.; Hao, H.J.; Cheng, Y.; Xie, Z.Y.; Yin, Y.Q.; Zhang, Q.; Gao, J.Q.; Liu, H.Y.; Mu, Y.M.; Han, W.D. Adipose-Derived Mesenchymal Stem Cells Ameliorate Hyperglycemia through Regulating Hepatic Glucose Metabolism in Type 2 Diabetic Rats. Biochem. Biophys. Res. Commun. 2017, 483, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage Cells from Human Adipose Tissue: Implications for Cell-Based Therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef]
- Mikłosz, A.; Nikitiuk, B.E.; Chabowski, A. Using Adipose-Derived Mesenchymal Stem Cells to Fight the Metabolic Complications of Obesity: Where Do We Stand? Obes. Rev. Off. J. Int. Assoc. Study Obes. 2022, 23, e13413. [Google Scholar] [CrossRef]
- Bhansali, A.; Upreti, V.; Khandelwal, N.; Marwaha, N.; Gupta, V.; Sachdeva, N.; Sharma, R.R.; Saluja, K.; Dutta, P.; Walia, R.; et al. Efficacy of Autologous Bone Marrow-Derived Stem Cell Transplantation in Patients with Type 2 Diabetes Mellitus. Stem Cells Dev. 2009, 18, 1407–1416. [Google Scholar] [CrossRef]
- Valencia, J.; Blanco, B.; Yáñez, R.; Vázquez, M.; Herrero Sánchez, C.; Fernández-García, M.; Rodríguez Serrano, C.; Pescador, D.; Blanco, J.F.; Hernando-Rodríguez, M.; et al. Comparative Analysis of the Immunomodulatory Capacities of Human Bone Marrow- and Adipose Tissue-Derived Mesenchymal Stromal Cells from the Same Donor. Cytotherapy 2016, 18, 1297–1311. [Google Scholar] [CrossRef] [PubMed]
- Kocan, B.; Maziarz, A.; Tabarkiewicz, J.; Ochiya, T.; Banaś-Ząbczyk, A. Trophic Activity and Phenotype of Adipose Tissue-Derived Mesenchymal Stem Cells as a Background of Their Regenerative Potential. Stem Cells Int. 2017, 2017, 1653254. [Google Scholar] [CrossRef] [PubMed]
- Banas, A.; Teratani, T.; Yamamoto, Y.; Tokuhara, M.; Takeshita, F.; Osaki, M.; Kawamata, M.; Kato, T.; Okochi, H.; Ochiya, T. IFATS Collection: In Vivo Therapeutic Potential of Human Adipose Tissue Mesenchymal Stem Cells after Transplantation into Mice with Liver Injury. Stem Cells 2008, 26, 2705–2712. [Google Scholar] [CrossRef] [PubMed]
- El-Badawy, A.; Amer, M.; Abdelbaset, R.; Sherif, S.N.; Abo-Elela, M.; Ghallab, Y.H.; Abdelhamid, H.; Ismail, Y.; El-Badri, N. Adipose Stem Cells Display Higher Regenerative Capacities and More Adaptable Electro-Kinetic Properties Compared to Bone Marrow-Derived Mesenchymal Stromal Cells. Sci. Rep. 2016, 6, 37801. [Google Scholar] [CrossRef]
- Karaoz, E.; Okcu, A.; Ünal, Z.S.; Subasi, C.; Saglam, O.; Duruksu, G. Adipose Tissue-Derived Mesenchymal Stromal Cells Efficiently Differentiate into Insulin-Producing Cells in Pancreatic Islet Microenvironment Both in Vitro and in Vivo. Cytotherapy 2013, 15, 557–570. [Google Scholar] [CrossRef]
- Wang, M.; Song, L.; Strange, C.; Dong, X.; Wang, H. Therapeutic Effects of Adipose Stem Cells from Diabetic Mice for the Treatment of Type 2 Diabetes. Mol. Ther. 2018, 26, 1921–1930. [Google Scholar] [CrossRef]
- Hu, J.; Fu, Z.; Chen, Y.; Tang, N.; Wang, L.; Wang, F.; Sun, R.; Yan, S. Effects of Autologous Adipose-Derived Stem Cell Infusion on Type 2 Diabetic Rats. Endocr. J. 2015, 62, 339–352. [Google Scholar] [CrossRef]
- Sanap, A.; Bhonde, R.; Joshi, K. Conditioned Medium of Adipose Derived Mesenchymal Stem Cells Reverse Insulin Resistance through Downregulation of Stress Induced Serine Kinases. Eur. J. Pharmacol. 2020, 881, 173215. [Google Scholar] [CrossRef]
- Chen, T.; Wu, Y.; Gu, W.; Xu, Q. Response of Vascular Mesenchymal Stem/Progenitor Cells to Hyperlipidemia. Cell. Mol. Life Sci. 2018, 75, 4079–4091. [Google Scholar] [CrossRef]
- Takafuji, Y.; Hori, M.; Mizuno, T.; Harada-Shiba, M. Humoral Factors Secreted from Adipose Tissue-Derived Mesenchymal Stem Cells Ameliorate Atherosclerosis in Ldlr−/− Mice. Cardiovasc. Res. 2019, 115, 1041–1051. [Google Scholar] [CrossRef]
- Clemente-Suárez, V.J. Multidisciplinary intervention in the treatment of mixed anxiety and depression disorder. Physiol. Behav. 2020, 219, 112858. [Google Scholar] [CrossRef] [PubMed]
- Tornero-Aguilera, J.F.; Pelarigo, J.G.; Clemente-Suárez, V.J. Psychophysiological intervention to improve preparedness in military special operations forces. Aerosp. Med. Hum. Perform. 2019, 90, 953–958. [Google Scholar] [CrossRef]
- Clemente-Suárez, V.J.; Ramírez-Goerke, M.I.; Redondo-Flórez, L.; Beltrán-Velasco, A.I.; Martín-Rodríguez, A.; Ramos-Campo, D.J.; Navarro-Jiménez, E.; Yáñez-Sepúlveda, R.; Tornero-Aguilera, J.F. The Impact of Anorexia Nervosa and the Basis for Non-Pharmacological Interventions. Nutrients 2023, 15, 2594. [Google Scholar] [CrossRef] [PubMed]
- Bustamante-Sanchez, A.; Villegas-Mora, B.E.; Martínez-Guardado, I.; Tornero-Aguilera, J.F.; Ardigò, L.P.; Nobari, H.; Clemente-Suárez, V.J. Physical Activity and Nutritional Pattern Related to Maturation and Development. Sustainability 2022, 14, 16958. [Google Scholar] [CrossRef]
- Martínez-González, M.B.; Arenas-Rivera, C.P.; Cardozo-Rusinque, A.A.; Morales-Cuadro, A.R.; Acuña-Rodríguez, M.; Turizo-Palencia, Y.; Clemente-Suárez, V.J. Psychological and Gender Differences in a Simulated Cheating Coercion Situation at School. Soc. Sci. 2021, 10, 265. [Google Scholar] [CrossRef]
- Martínez-González, M.B.; Pérez-Pedraza, D.C.; Alfaro-Álvarez, J.; Reyes-Cervantes, C.; González-Malabet, M.; Clemente-Suárez, V.J. Women facing psychological abuse: How do they respond to maternal identity humiliation and body shaming? Int. J. Environ. Res. Public Health 2021, 18, 6627. [Google Scholar] [CrossRef] [PubMed]
- Ruisoto, P.; Ramírez, M.R.; García, P.A.; Paladines-Costa, B.; Vaca, S.L.; Clemente-Suárez, V.J. Social support mediates the effect of burnout on health in health care professionals. Front. Psychol. 2021, 11, 623587. [Google Scholar] [CrossRef]
- Soto-Baño, M.A.; Clemente-Suárez, V.J. Psicología de emergencias en España: Análisis actual, normativa y proposición reguladora. Pap. Psicól. 2021, 42, 46–55. [Google Scholar]
- Clemente-Suárez, V.J.; Martínez-González, M.B.; Benitez-Agudelo, J.C.; Navarro-Jiménez, E.; Beltran-Velasco, A.I.; Ruisoto, P.; Diaz Arroyo, E.; Laborde-Cárdenas, C.C.; Tornero-Aguilera, J.F. The impact of the COVID-19 pandemic on mental disorders. A critical review. Int. J. Environ. Res. Public Health 2021, 18, 10041. [Google Scholar] [CrossRef] [PubMed]
- Soto-Baño, M.A.; Clemente-Suárez, V.J. Psicología de emergencias en España: Delimitación conceptual, ámbitos de actuación y propuesta de un sistema asistencial. Pap. Psicól. 2021, 42, 56–66. [Google Scholar]
- Clemente-Suárez, V.J.; Dalamitros, A.A.; Beltran-Velasco, A.I.; Mielgo-Ayuso, J.; Tornero-Aguilera, J.F. Social and psychophysiological consequences of the COVID-19 pandemic: An extensive literature review. Front. Psychol. 2020, 11, 3077. [Google Scholar] [CrossRef] [PubMed]
- Clemente-Suárez, V.J.; Redondo-Flórez, L.; Rubio-Zarapuz, A.; Martínez-Guardado, I.; Navarro-Jiménez, E.; Tornero-Aguilera, J.F. Nutritional and exercise interventions in cancer-related cachexia: An extensive narrative review. Int. J. Environ. Res. Public Health 2022, 19, 4604. [Google Scholar] [CrossRef]
- Clemente-Suárez, V.J.; Bustamante-Sanchez, Á.; Tornero-Aguilera, J.F.; Ruisoto, P.; Mielgo-Ayuso, J. Inflammation in COVID-19 and the Effects of Non-Pharmacological Interventions during the Pandemic: A Review. Int. J. Mol. Sci. 2022, 23, 15584. [Google Scholar] [CrossRef] [PubMed]
- Clemente-Suárez, V.J.; Beltrán-Velasco, A.I.; Ramos-Campo, D.J.; Mielgo-Ayuso, J.; Nikolaidis, P.A.; Belando, N.; Tornero-Aguilera, J.F. Physical activity and COVID-19. The basis for an efficient intervention in times of COVID-19 pandemic. Physiol. Behav. 2022, 244, 113667. [Google Scholar] [CrossRef]
- Clemente-Suárez, V.J.; Navarro-Jiménez, E.; Moreno-Luna, L.; Saavedra-Serrano, M.C.; Jimenez, M.; Simón, J.A.; Tornero-Aguilera, J.F. The impact of the COVID-19 pandemic on social, health, and economy. Sustainability 2021, 13, 6314. [Google Scholar] [CrossRef]
- Clemente-Suárez, V.J.; Navarro-Jiménez, E.; Ruisoto, P.; Dalamitros, A.A.; Beltran-Velasco, A.I.; Hormeño-Holgado, A.; Laborde-Cárdenas, C.C.; Tornero-Aguilera, J.F. Performance of fuzzy multi-criteria decision analysis of emergency system in COVID-19 pandemic. An extensive narrative review. Int. J. Environ. Res. Public Health 2021, 18, 5208. [Google Scholar] [CrossRef] [PubMed]
- Clemente-Suárez, V.J.; Martín-Rodríguez, A.; Yáñez-Sepúlveda, R.; Tornero-Aguilera, J.F. Mitochondrial Transfer as a Novel Therapeutic Approach in Disease Diagnosis and Treatment. Int. J. Mol. Sci. 2023, 24, 8848. [Google Scholar] [CrossRef]
- Clemente-Suárez, V.J.; Navarro-Jiménez, E.; Jimenez, M.; Hormeño-Holgado, A.; Martinez-Gonzalez, M.B.; Benitez-Agudelo, J.C.; Perez-Palencia, N.; Laborde-Cárdenas, C.C.; Tornero-Aguilera, J.F. Impact of COVID-19 pandemic in public mental health: An extensive narrative review. Sustainability 2021, 13, 3221. [Google Scholar] [CrossRef]





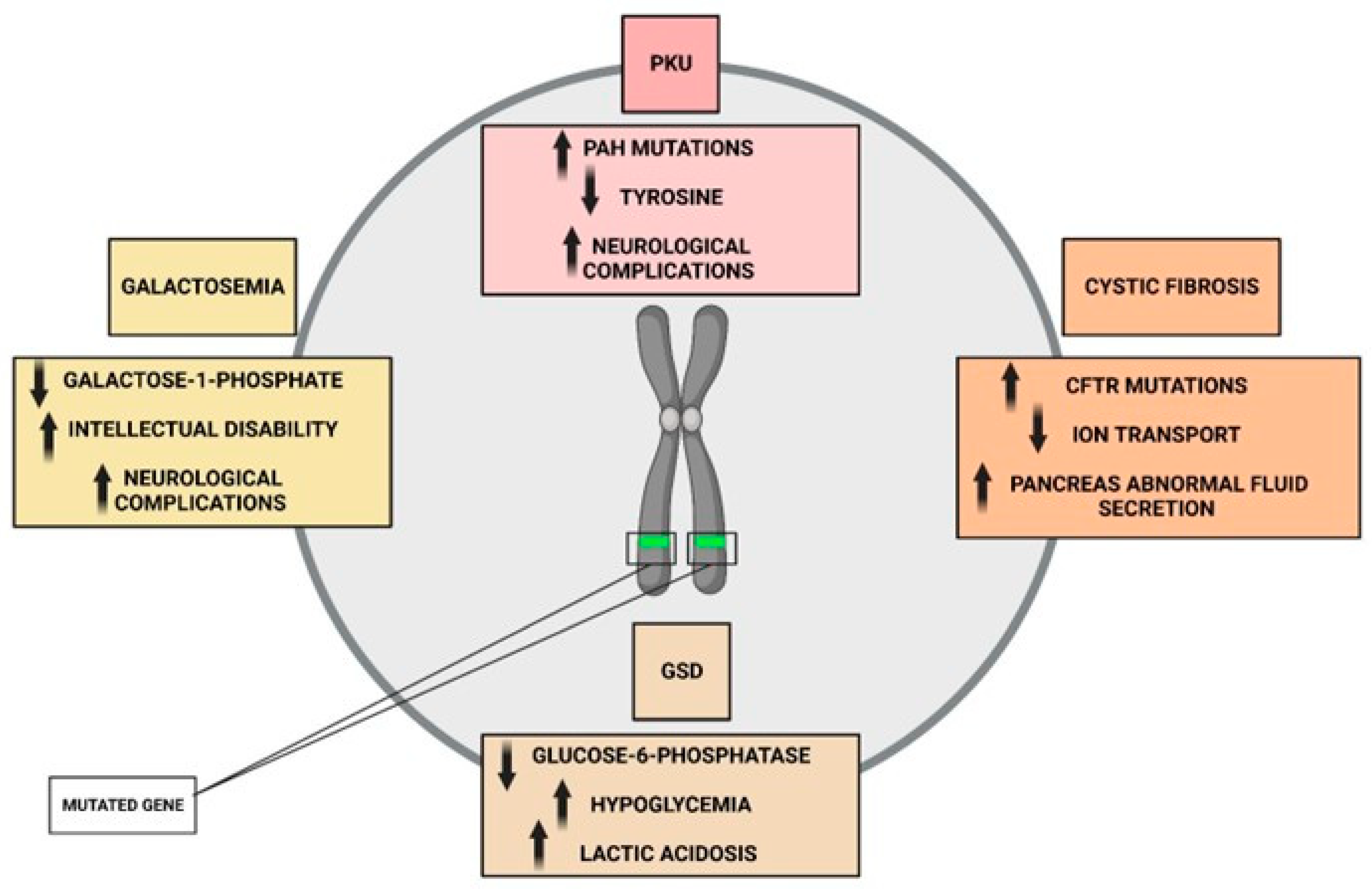{kind=link}
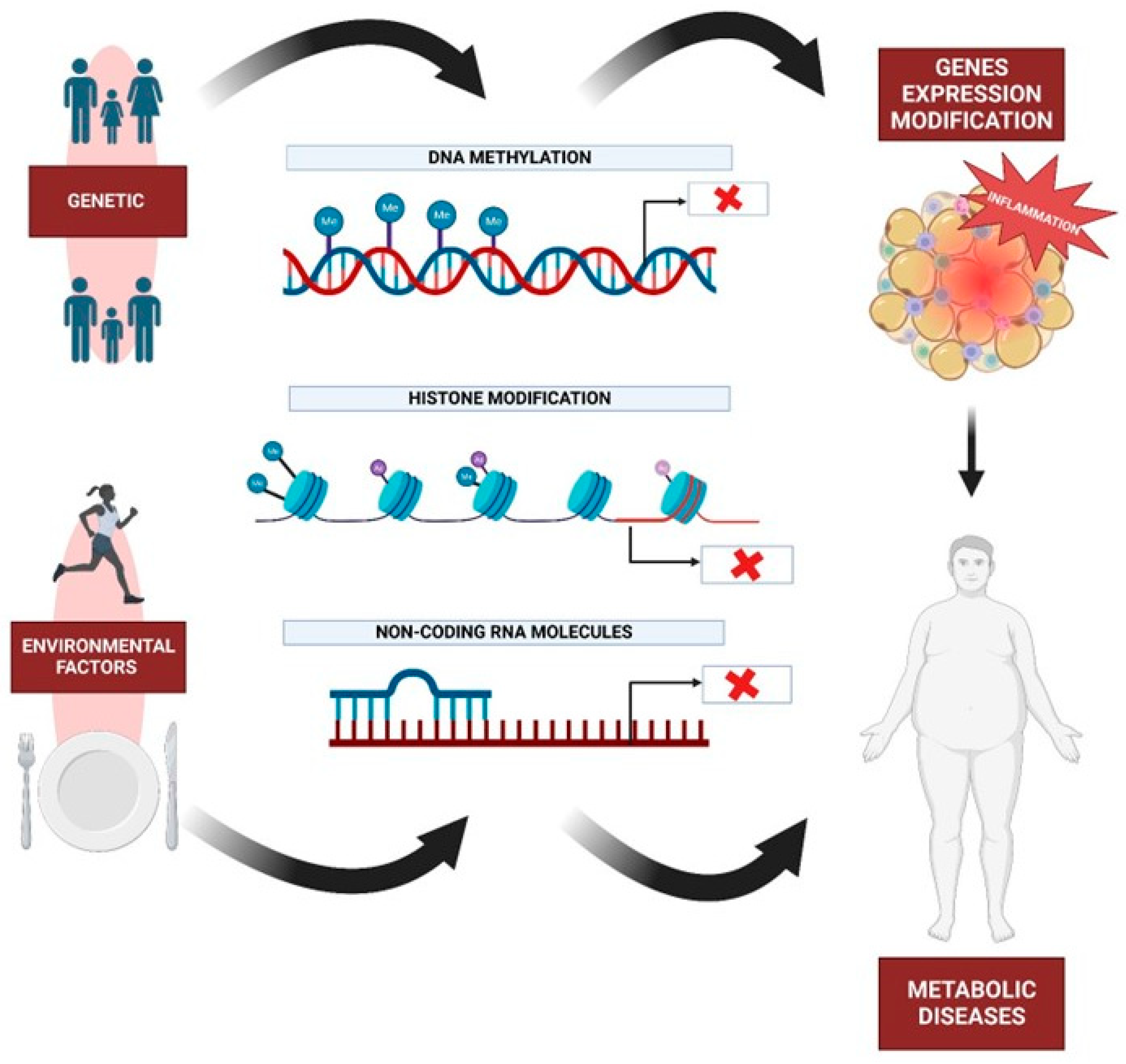{kind=link}
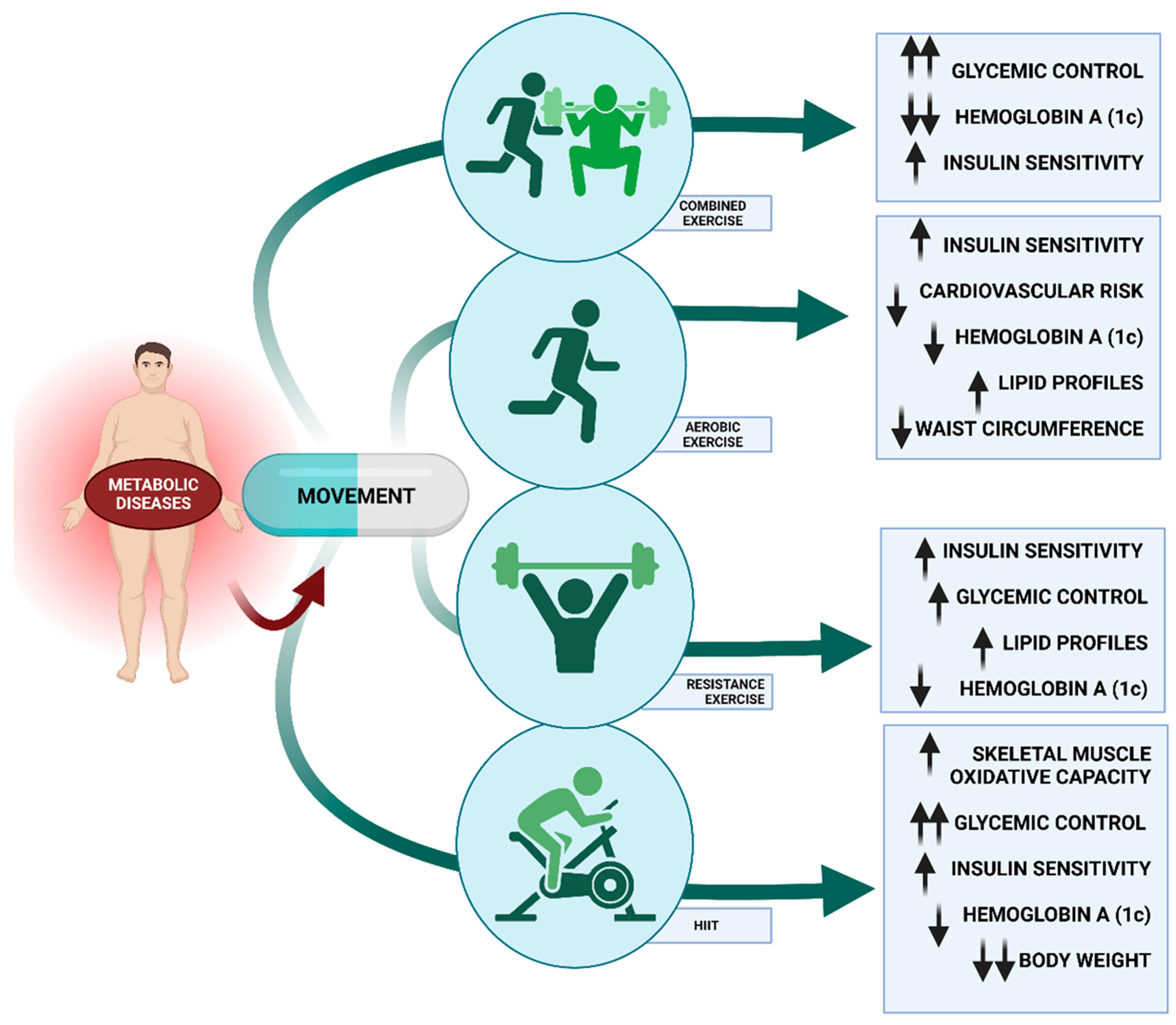{kind=link}
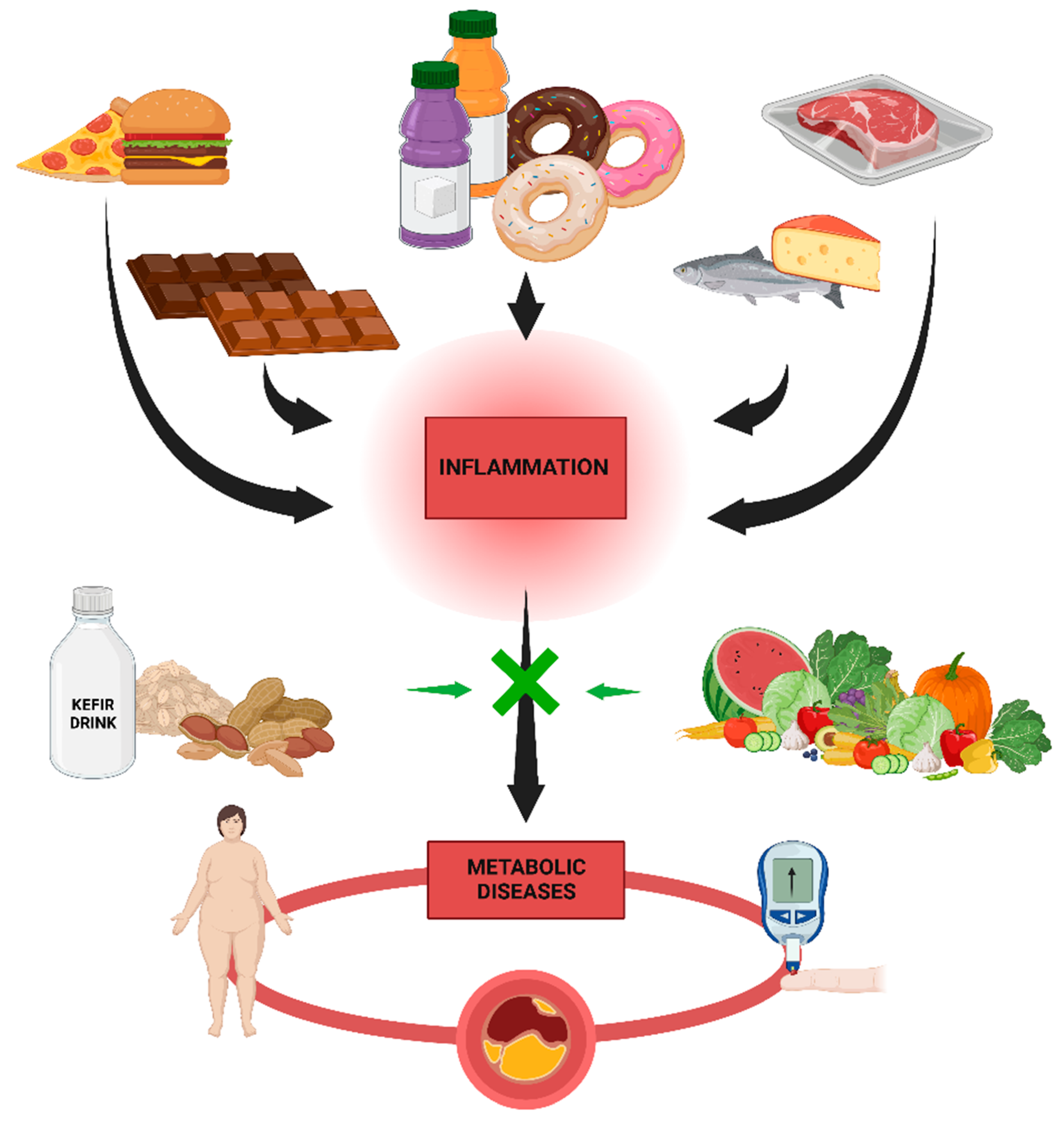{kind=link}
| Authors and Year | Study Title | Aim of Study | Main Outcomes | Section |
|---|---|---|---|---|
| Han et al. [60] | Gene Therapy for Metabolic Diseases | Asses viral-mediated gene addition approaches | Most of the current success has been achieved using a viral-mediated gene addition approach | Genetic basis |
| Litwin et al. [81] | Negative Emotions and Emotional Eating: The Mediating Role of Experiential Avoidance | Examine whether experiential avoidance would mediate the association between negative emotions and emotional eating | Experiential avoidance mediated the relationship between negative emotions and emotional eating | Psychological and behavioral basis |
| Hansen et al. [100] | Impact of Endurance Exercise Training in the Fasted State on Muscle Biochemistry and Metabolism in Healthy Subjects: Can These Effects Be of Particular Clinical Benefit to Type 2 Diabetes Mellitus and Insulin-Resistant Patients? | Describe the impact of endurance exercise (training) in the fasted versus fed state on parameters of muscle biochemistry and metabolism linked to glycemic control or insulin sensitivity in healthy subjects | Promising results of exercise (training) in the fasted state have been found in healthy subjects on parameters of muscle biochemistry and metabolism linked to insulin sensitivity and glycemic control | Physical Activity |
| Ramallal et al. [132] | Inflammatory Potential of Diet, Weight Gain, and Incidence of Overweight/Obesity: The SUN Cohort | Assess the association of the inflammatory potential of a diet using the dietary inflammatory index (DII) with average yearly weight changes and incident overweight/obesity. | Consistently, increases in average yearly weight gains were significantly associated with proinflammatory diets | Nutrition |
| Hovratin et al. [165] | Toward Modeling Metabolic State from Single-Cell Transcriptomics | Summarize the current state of single-cell metabolic measurement and modeling approaches, motivating the use of computational techniques | Single-cell metabolic modeling is a rising field that provides a new perspective for understanding cellular functions | Single-cell transcriptomics |
| Bao et al. [178] | Pseudotime Ordering Single-Cell Transcriptomic of β Cells Pancreatic Islets in Health and Type 2 Diabetes | Evaluate that human pancreatic β cells are heterogeneous and demonstrated the transcript alterations of β cell subpopulation in diabetes. | identified three major states including (1) Normal branch, (2) Obesity-like branch and (3) T2D-like branch based on biomarker genes and genes that give rise to bifurcation in the trajectory. β cell function-maintain-related genes, insulin expression-related genes, and T2D-related genes enriched in three branches, respectively | Single-cell transcriptomics |
| Sankararaman et al. [184] | Gut Microbiome and Its Impact on Obesity and Obesity-Related Disorders | Review the current literature on gut microbiome and its impact on obesity and obesity-related disorders | An altered gut microbial composition in obesity and obesity-related disorders is associated with enhanced energy extraction from the non-digestible dietary carbohydrates, increased gut permeability, and increased production of proinflammatory metabolites, such as lipopolysaccharides, resulting in systemic inflammation and insulin resistance | Gut microbiome |
| Rönn et al. [226] | A Six Months Exercise Intervention Influences the Genome-Wide DNA Methylation Pattern in Human Adipose Tissue | Describe the genome-wide pattern of DNA methylation in human adipose tissue from 23 healthy men, with a previous low level of physical activity, before and after a six months exercise intervention | A simultaneous change in mRNA expression was seen for 6 of those genes. To understand if genes that exhibit differential DNA methylation and mRNA expression in human adipose tissue in vivo affect adipocyte metabolism | Epigenetics |
| Berg et al. [248] | Innovations in Instrumentation for Positron Emission Tomography | Review the current state of the art in PET instrumentation, detectors and systems, describe the major limitations in PET as currently practiced, and offer our own personal insights into some of the recent and emerging technological innovations that we believe will impact the field | Combining these emerging technological and methodological advances promises to lead to a generation of PET scanners tailored for specific applications that really can claim to approach the fundamental limits set forth by the physics of radioactive decay and the statistics of the available signal. | Imaging techniques |
| Szot et al. [302] | Tolerance Induction and Reversal of Diabetes in Mice Transplanted with Human Embryonic Stem Cell-Derived Pancreatic Endoderm | Demonstrate that immunotherapies that target T cell costimulatory pathways block the rejection of xenogeneic human embryonic-stem-cell-derived pancreatic endoderm (hESC-PE) in mice | Costimulation blockade prevents rejection of xenogeneic hESC-derived islets; Short-term treatment induces long-term tolerance to xenogeneic hESC-derived islets; Tolerance induced by costimulation blockade is transferable independently of Tregs; Costimulation blockade prevents rejection of allogeneic; hESC islets by human PBMCs | Cell-based therapies |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clemente-Suárez, V.J.; Martín-Rodríguez, A.; Redondo-Flórez, L.; López-Mora, C.; Yáñez-Sepúlveda, R.; Tornero-Aguilera, J.F. New Insights and Potential Therapeutic Interventions in Metabolic Diseases. Int. J. Mol. Sci. 2023, 24, 10672. https://doi.org/10.3390/ijms241310672
Clemente-Suárez VJ, Martín-Rodríguez A, Redondo-Flórez L, López-Mora C, Yáñez-Sepúlveda R, Tornero-Aguilera JF. New Insights and Potential Therapeutic Interventions in Metabolic Diseases. International Journal of Molecular Sciences. 2023; 24(13):10672. https://doi.org/10.3390/ijms241310672
Chicago/Turabian StyleClemente-Suárez, Vicente Javier, Alexandra Martín-Rodríguez, Laura Redondo-Flórez, Clara López-Mora, Rodrigo Yáñez-Sepúlveda, and José Francisco Tornero-Aguilera. 2023. "New Insights and Potential Therapeutic Interventions in Metabolic Diseases" International Journal of Molecular Sciences 24, no. 13: 10672. https://doi.org/10.3390/ijms241310672
APA StyleClemente-Suárez, V. J., Martín-Rodríguez, A., Redondo-Flórez, L., López-Mora, C., Yáñez-Sepúlveda, R., & Tornero-Aguilera, J. F. (2023). New Insights and Potential Therapeutic Interventions in Metabolic Diseases. International Journal of Molecular Sciences, 24(13), 10672. https://doi.org/10.3390/ijms241310672