
Anti-Inflammatory Activity of Black Soldier Fly Oil Associated with Modulation of TLR Signaling: A Metabolomic Approach


Abstract
1. Introduction
2. Results
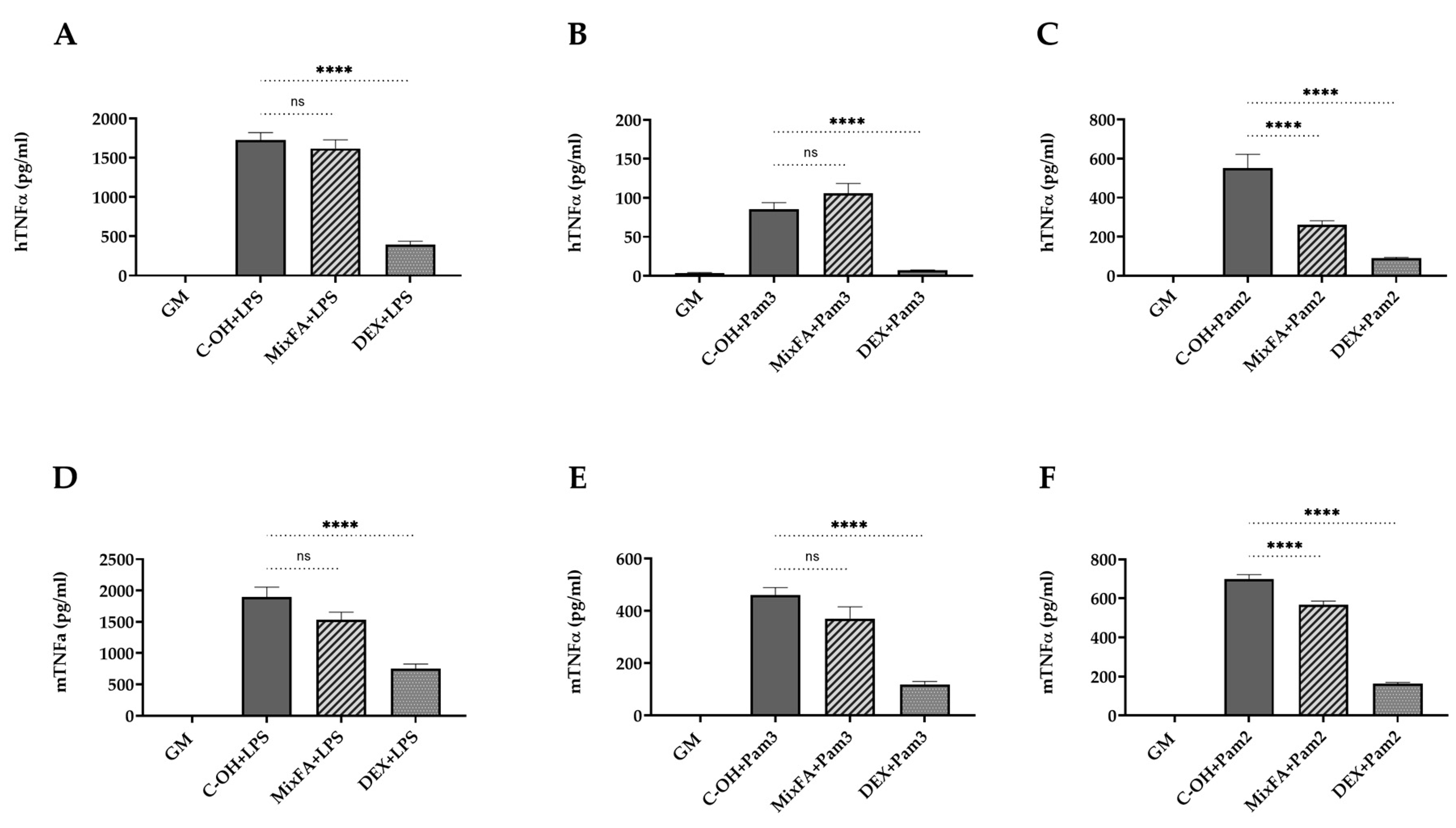
2.1. Suppression of TLR4-Mediated Proinflammatory Cytokines by Modified BSFL Oil (MBSFL) and C12:0 in Macrophages
2.2. MBSFL and C12:0 Reciprocally Modulate TLR2-Mediated Proinflammatory Cytokines Expression and Secretion in Macrophages
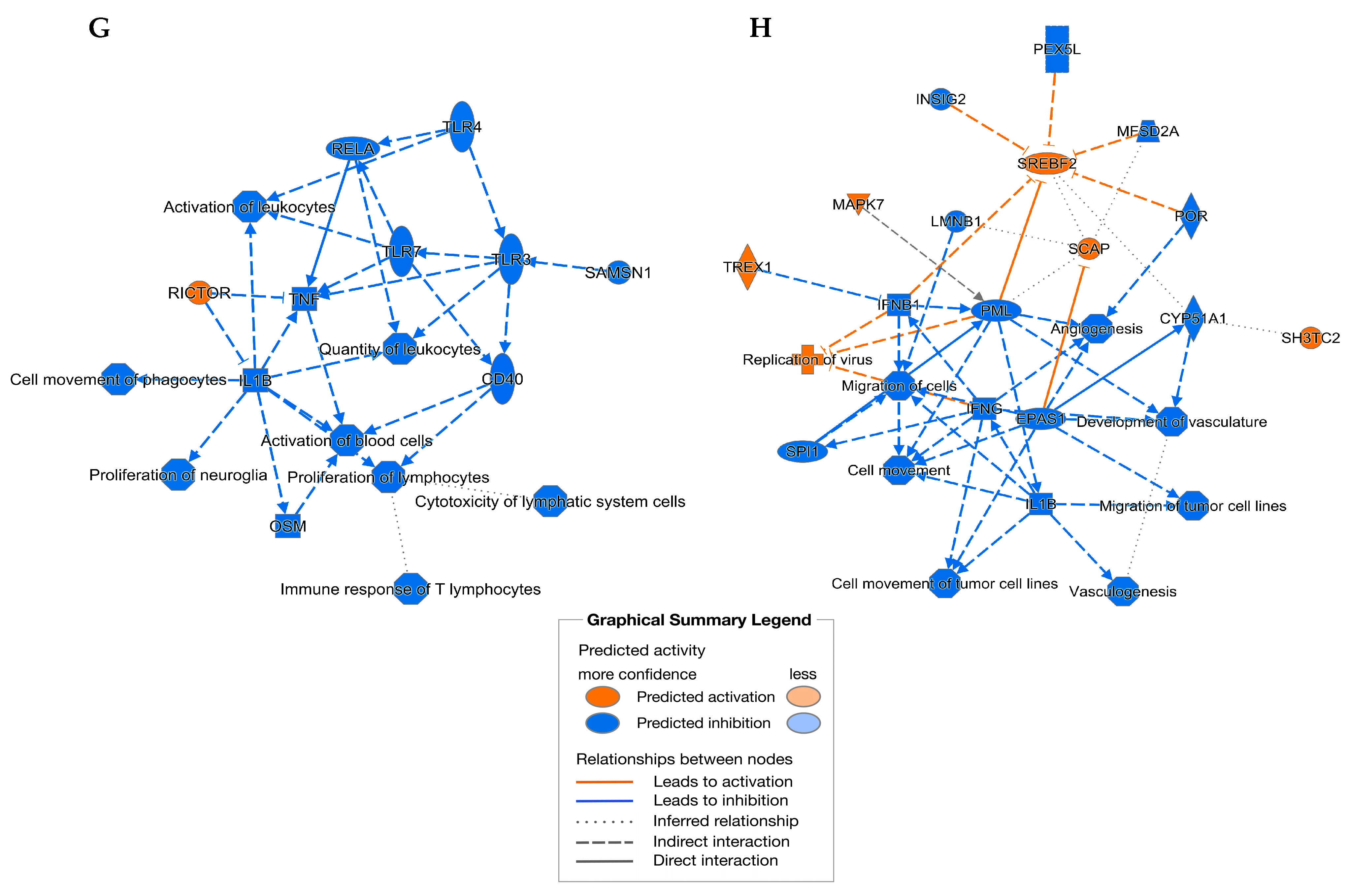
2.3. Treatment with MBSFL or SL Differentially Modifies the Transcriptome Profile of LPS or Pam3-Stimulated THP-1 Cells
2.4. Fatty Acid Composition of the BSFL Oil Is Not Directly Correlated to Its Anti-Inflammatory Effect
2.5. The Ensemble of Anti-Inflammatory Bioactive Lipids Comprising BSFL Oil May Contribute to Its Anti-Inflammatory Activity
2.6. Amelioration of Colitis-Associated Indicators in DSS-Treated Mice by BSFL Oil
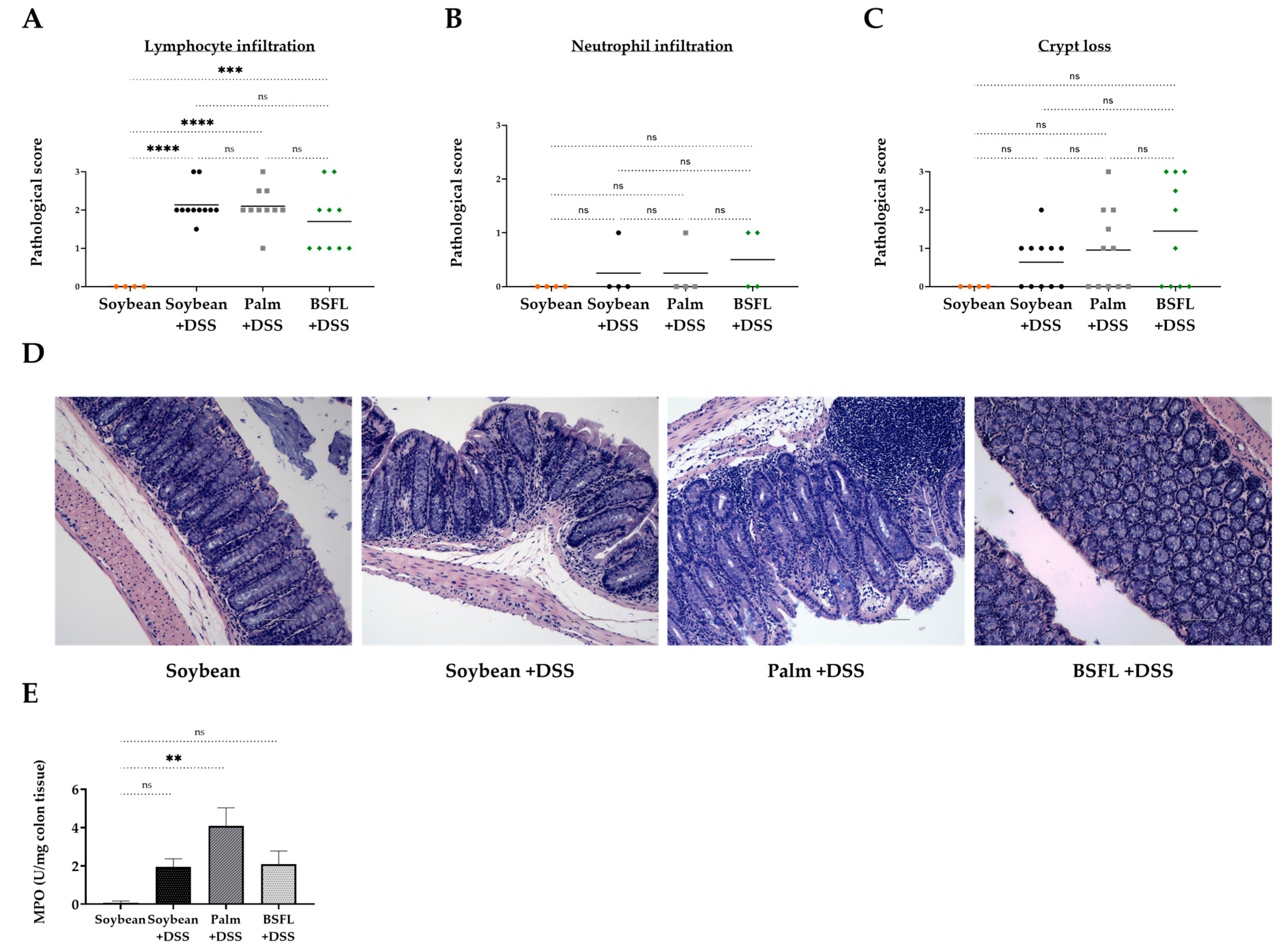
2.7. Histopathological Signs of Colitis in DSS-Treated Mice as a Function of Diet Composition
2.8. BSFL Oil Induces an Enhanced Earlier Expression of Secretory IgA in Response to DSS Treatment
3. Discussion
4. Materials and Methods
4.1. Modified BSFL (MBSFL) Oil
4.2. Fatty Acid–Albumin Complex
4.3. Cell Lines
4.4. Cell Treatment
4.5. Cell Viability Assay
4.6. Enzyme-Linked Immunosorbent Assay (ELISA)
4.7. Reverse Transcription PCR (RT-PCR) and Real-Time PCR (qPCR)
4.8. RNA Sequencing Protocol and Computational Pipeline
4.9. Metabolomics
4.10. Animals and DSS Model
4.11. Fatty Acid Composition
4.12. Disease Activity Index (DAI)
4.13. Histological Analysis
4.14. Myeloperoxidase (MPO) Assay
4.15. Fecal IgA
4.16. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lu, Y.; Li, X.; Liu, S.; Zhang, Y.; Zhang, D. Toll-like Receptors and Inflammatory Bowel Disease. Front. Immunol. 2018, 9, 72. [Google Scholar] [CrossRef] [PubMed]
- Płóciennikowska, A.; Hromada-Judycka, A.; Borzęcka, K.; Kwiatkowska, K. Co-Operation of TLR4 and Raft Proteins in LPS-Induced pro-Inflammatory Signaling. Cell. Mol. Life Sci. 2015, 72, 557–581. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, G.I.; Langley, K.G.; Berglund, N.A.; Kammoun, H.L.; Reibe, S.; Estevez, E.; Weir, J.; Mellett, N.A.; Pernes, G.; Conway, J.R.W.; et al. Evidence That TLR4 Is Not a Receptor for Saturated Fatty Acids but Mediates Lipid-Induced Inflammation by Reprogramming Macrophage Metabolism. Cell Metab. 2018, 27, 1096–1110.e5. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Qian, Y.; Fang, Q.; Zhong, P.; Li, W.; Wang, L.; Fu, W.; Zhang, Y.; Xu, Z.; Li, X.; et al. Saturated Palmitic Acid Induces Myocardial Inflammatory Injuries through Direct Binding to TLR4 Accessory Protein MD2. Nat. Commun. 2017, 8, 13997. [Google Scholar] [CrossRef]
- Nicholas, D.A.; Zhang, K.; Hung, C.; Glasgow, S.; Aruni, A.W.; Unternaehrer, J.; Payne, K.J.; Langridge, W.H.R.; De Leon, M. Palmitic Acid Is a Toll-like Receptor 4 Ligand That Induces Human Dendritic Cell Secretion of IL-1β. PLoS ONE 2017, 12, e0176793. [Google Scholar] [CrossRef]
- Hattor, Y.; Kasai, K.; Akimoto, K.; Thiemermann, C. Induction of NO Synthesis by Lipoteichoic Acid from Staphylococcus aureus in J774 Macrophages: Involvement of a CD14-Dependent Pathway. Biochem. Biophys. Res. Commun. 1997, 233, 375–379. [Google Scholar] [CrossRef]
- Ruysschaert, J.M.; Lonez, C. Role of Lipid Microdomains in TLR-Mediated Signalling. Biochim. Biophys. Acta Biomembr. 2015, 1848, 1860–1867. [Google Scholar] [CrossRef]
- Kang, J.Y.; Nan, X.; Jin, M.S.; Youn, S.J.; Ryu, Y.H.; Mah, S.; Han, S.H.; Lee, H.; Paik, S.G.; Lee, J.O. Recognition of Lipopeptide Patterns by Toll-like Receptor 2-Toll-like Receptor 6 Heterodimer. Immunity 2009, 31, 873–884. [Google Scholar] [CrossRef]
- Acosta-Martinez, M.; Cabail, M.Z. The PI3K/Akt Pathway in Meta-Inflammation. Int. J. Mol. Sci. 2022, 23, 15330. [Google Scholar] [CrossRef]
- Wu, M.M.; Wang, Q.M.; Huang, B.Y.; Mai, C.T.; Wang, C.L.; Wang, T.T.; Zhang, X.J. Dioscin Ameliorates Murine Ulcerative Colitis by Regulating Macrophage Polarization. Pharmacol. Res. 2021, 172, 105796. [Google Scholar] [CrossRef]
- Ganeshan, K.; Chawla, A. Metabolic Regulation of Immune Responses. Annu. Rev. Immunol. 2014, 32, 609–634. [Google Scholar] [CrossRef] [PubMed]
- Gasco, L.; Finke, M.; van Huis, A. Can Diets Containing Insects Promote Animal Health? J. Insects Food Feed 2018, 4, 1–4. [Google Scholar] [CrossRef]
- Papada, E.; Kaliora, A.C.; Gioxari, A.; Papalois, A.; Forbes, A. Anti-Inflammatory Effect of Elemental Diets with Different Fat Composition in Experimental Colitis. Br. J. Nutr. 2014, 111, 1213–1220. [Google Scholar] [CrossRef]
- Geng, S.; Zhu, W.; Xie, C.; Li, X.; Wu, J.; Liang, Z.; Xie, W.; Zhu, J.; Huang, C.; Zhu, M.; et al. Medium-Chain Triglyceride Ameliorates Insulin Resistance and Inflammation in High Fat Diet-Induced Obese Mice. Eur. J. Nutr. 2016, 55, 931–940. [Google Scholar] [CrossRef]
- Bertevello, P.L.; De Nardi, L.; Torrinhas, R.S.; Logullo, A.F.; Waitzberg, D.L. Partial Replacement of ω-6 Fatty Acids with Medium-Chain Triglycerides, but Not Olive Oil, Improves Colon Cytokine Response and Damage in Experimental Colitis. J. Parenter. Enter. Nutr. 2012, 36, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.U.; Aamir, K.; Jusuf, P.R.; Sethi, G.; Sisinthy, S.P.; Ghildyal, R.; Arya, A. Lauric Acid Ameliorates Lipopolysaccharide (LPS)-Induced Liver Inflammation by Mediating TLR4/MyD88 Pathway in Sprague Dawley (SD) Rats. Life Sci. 2021, 265, 118750. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Tsai, T.H.; Chuang, L.T.; Li, Y.Y.; Zouboulis, C.C.; Tsai, P.J. Anti-Bacterial and Anti-Inflammatory Properties of Capric Acid against Propionibacterium Acnes: A Comparative Study with Lauric Acid. J. Dermatol. Sci. 2014, 73, 232–240. [Google Scholar] [CrossRef]
- Kono, H.; Fujii, H.; Asakawa, M.; Maki, A.; Amemiya, H.; Hirai, Y.; Matsuda, M.; Yamamoto, M. Medium-Chain Triglycerides Enhance Secretory IgA Expression in Rat Intestine after Administration of Endotoxin. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 286, 1081–1089. [Google Scholar] [CrossRef]
- Okai, S.; Usui, F.; Yokota, S.; Hori-i, Y.; Hasegawa, M.; Nakamura, T.; Kurosawa, M.; Okada, S.; Yamamoto, K.; Nishiyama, E.; et al. High-Affinity Monoclonal IgA Regulates Gut Microbiota and Prevents Colitis in Mice. Nat. Microbiol. 2016, 1, 16103. [Google Scholar] [CrossRef]
- Machate, D.J.; Figueiredo, P.S.; Marcelino, G.; de Cássia Avellaneda Guimarães, R.; Hiane, P.A.; Bogo, D.; Pinheiro, V.A.Z.; Silva de Oliveira, L.C.; Pott, A. Fatty Acid Diets: Regulation of Gut Microbiota Composition and Obesity and Its Related Metabolic Dysbiosis. Int. J. Mol. Sci. 2020, 21, 4093. [Google Scholar] [CrossRef]
- Tam, J.S.Y. Toll-like Receptor 4 (TLR4) Antagonists as Potential Therapeutics for Intestinal Inflammation. Indian J. Gastroenterol. 2021, 40, 5–21. [Google Scholar] [CrossRef] [PubMed]
- Shmuel-Galia, L.; Aychek, T.; Fink, A.; Porat, Z.; Zarmi, B.; Bernshtein, B.; Brenner, O.; Jung, S.; Shai, Y. Neutralization of Pro-inflammatory Monocytes by Targeting TLR2 Dimerization Ameliorates Colitis. EMBO J. 2016, 35, 685–698. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhao, Y.; Hou, T.; Zeng, H.; Kalambhe, D.; Wang, B.; Shen, X.; Huang, Y. Macrophage-Based Nanotherapeutic Strategies in Ulcerative Colitis. J. Control. Release 2020, 320, 363–380. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, Y.; Moriyama, M.; Kawabe, K.; Satoh, H.; Takano, K.; Azuma, Y.T.; Nakamura, Y. Lauric Acid Alleviates Neuroinflammatory Responses by Activated Microglia: Involvement of the GPR40-Dependent Pathway. Neurochem. Res. 2018, 43, 1723–1735. [Google Scholar] [CrossRef]
- Lee, J.Y.; Zhao, L.; Youn, H.S.; Weatherill, A.R.; Tapping, R.; Feng, L.; Lee, W.H.; Fitzgerald, K.A.; Hwang, D.H. Saturated Fatty Acid Activates but Polyunsaturated Fatty Acid Inhibits Toll-like Receptor 2 Dimerized with Toll-like Receptor 6 or 1. J. Biol. Chem. 2004, 279, 16971–16979. [Google Scholar] [CrossRef]
- Lee, J.Y.; Ye, J.; Gao, Z.; Youn, H.S.; Lee, W.H.; Zhao, L.; Sizemore, N.; Hwang, D.H. Reciprocal Modulation of Toll-like Receptor-4 Signaling Pathways Involving MyD88 and Phosphatidylinositol 3-Kinase/AKT by Saturated and Polyunsaturated Fatty Acids. J. Biol. Chem. 2003, 278, 37041–37051. [Google Scholar] [CrossRef]
- Huang, S.; Rutkowsky, J.M.; Snodgrass, R.G.; Ono-Moore, K.D.; Schneider, D.A.; Newman, J.W.; Adams, S.H.; Hwang, D.H. Saturated Fatty Acids Activate TLR-Mediated Proinflammatory Signaling Pathways. J. Lipid Res. 2012, 53, 2002–2013. [Google Scholar] [CrossRef]
- Nakatomi, K.; Ueno, H.; Ishikawa, Y.; Salim, R.C.; Mori, Y.; Kanemoto, I.; Tancharoen, S.; Kikuchi, K.; Miura, N.; Omori, T.; et al. Tlr4/Md-2 Is a Receptor for Extracellular Nucleophosmin 1. Biomed. Rep. 2020, 14, 21. [Google Scholar] [CrossRef]
- Lin, X.; Yang, Y.; Guo, Y.; Liu, H.; Jiang, J.; Zheng, F.; Wu, B. PTTG1 Is Involved in TNF-α-Related Hepatocellular Carcinoma via the Induction of c-Myc. Cancer Med. 2019, 8, 5702–5715. [Google Scholar] [CrossRef]
- Scuruchi, M.; D’Ascola, A.; Avenoso, A.; Mandraffino, G.G.; Campo, S.S.; Campo, G.M. Serglycin as Part of IL-1β Induced Inflammation in Human Chondrocytes. Arch. Biochem. Biophys. 2019, 669, 80–86. [Google Scholar] [CrossRef]
- El Khoury, W.; Nasr, Z. Deregulation of Ribosomal Proteins in Human Cancers. Biosci. Rep. 2021, 41, BSR20211577. [Google Scholar] [CrossRef]
- Rosario, F.J.; Powell, T.L.; Gupta, M.B.; Cox, L.; Jansson, T. MTORC1 Transcriptional Regulation of Ribosome Subunits, Protein Synthesis, and Molecular Transport in Primary Human Trophoblast Cells. Front. Cell Dev. Biol. 2020, 8, 583801. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Fu, X.; Xu, S.; Qiu, P.; Lv, Z.; Cui, M.; Zhang, Q.; Xu, Y. OLR1 Is a Prognostic Factor and Correlated with Immune Infiltration in Breast Cancer. Int. Immunopharmacol. 2021, 101, 108275. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhang, L.; Liechty, C.; Zgheib, C.; Hodges, M.M.; Liechty, K.W.; Xu, J. Long Noncoding RNA GAS5 Regulates Macrophage Polarization and Diabetic Wound Healing. J. Investig. Dermatol. 2020, 140, 1629–1638. [Google Scholar] [CrossRef] [PubMed]
- Bai, D.; Zhao, Y.; Zhu, Q.; Zhou, Y.; Zhao, Y.; Zhang, T.; Guo, Q.; Lu, N. LZ205, a Newly Synthesized Flavonoid Compound, Exerts Anti-Inflammatory Effect by Inhibiting M1 Macrophage Polarization through Regulating PI3K/AKT/MTOR Signaling Pathway. Exp. Cell Res. 2018, 364, 84–94. [Google Scholar] [CrossRef]
- Tanaka, T.; Tahara-Hanaoka, S.; Nabekura, T.; Ikeda, K.; Jiang, S.; Tsutsumi, S.; Inagaki, T.; Magoori, K.; Higurashi, T.; Takahashi, H.; et al. PPARβ 2/Δactivation of CD300a Controls Intestinal Immunity. Sci. Rep. 2014, 4, 5412. [Google Scholar] [CrossRef]
- Lefere, S.; Puengel, T.; Hundertmark, J.; Penners, C.; Frank, A.K.; Guillot, A.; de Muynck, K.; Heymann, F.; Adarbes, V.; Defrêne, E.; et al. Differential Effects of Selective- and Pan-PPAR Agonists on Experimental Steatohepatitis and Hepatic Macrophages. J. Hepatol. 2020, 73, 757–770. [Google Scholar] [CrossRef]
- Varga, T.; Czimmerer, Z.; Nagy, L. PPARs Are a Unique Set of Fatty Acid Regulated Transcription Factors Controlling Both Lipid Metabolism and Inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2011, 1812, 1007–1022. [Google Scholar] [CrossRef]
- Eisele, P.S.; Furrer, R.; Beer, M.; Handschin, C. The PGC-1 Coactivators Promote an Anti-Inflammatory Environment in Skeletal Muscle in Vivo. Biochem. Biophys. Res. Commun. 2015, 464, 692–697. [Google Scholar] [CrossRef]
- Cherry, A.D.; Suliman, H.B.; Bartz, R.R.; Piantadosi, C.A. Peroxisome Proliferator-Activated Receptor γ Co-Activator 1-α as a Critical Co-Activator of the Murine Hepatic Oxidative Stress Response and Mitochondrial Biogenesis in Staphylococcus aureus Sepsis. J. Biol. Chem. 2014, 289, 41–52. [Google Scholar] [CrossRef]
- Nautiyal, J.; Christian, M.; Parker, M.G. Distinct Functions for RIP140 in Development, Inflammation, and Metabolism. Trends Endocrinol. Metab. 2013, 24, 451–459. [Google Scholar] [CrossRef]
- Guo, Z.; Shen, Y.; Zhong, J.; Li, Z.; Guo, Q.; Yao, X.; Wang, Y.; Wu, W. RIP140-Mediated NF-ΚB Inflammatory Pathway Promotes Metabolic Dysregulation in Retinal Pigment Epithelium Cells. Curr. Issues Mol. Biol. 2022, 44, 5788–5801. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Bensinger, S.J. Reprogramming Cholesterol Metabolism in Macrophages and Its Role in Host Defense against Cholesterol-Dependent Cytolysins. Cell. Mol. Immunol. 2022, 19, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Gilroy, D.W.; Edin, M.L.; De Maeyer, R.P.H.; Bystrom, J.; Newson, J.; Lih, F.B.; Stables, M.; Zeldin, D.C.; Bishop-Bailey, D. CYP450-Derived Oxylipins Mediate Inflammatory Resolution. Proc. Natl. Acad. Sci. USA 2016, 113, E3240–E3249. [Google Scholar] [CrossRef] [PubMed]
- Hildreth, K.; Kodani, S.D.; Hammock, B.D.; Zhao, L. Cytochrome P450-Derived Linoleic Acid Metabolites EpOMEs and DiHOMEs: A Review of Recent Studies. J. Nutr. Biochem. 2020, 86, 108484. [Google Scholar] [CrossRef]
- Pauls, S.D.; Rodway, L.A.; Winter, T.; Taylor, C.G.; Zahradka, P.; Aukema, H.M. Anti-Inflammatory Effects of α-Linolenic Acid in M1-like Macrophages Are Associated with Enhanced Production of Oxylipins from α-Linolenic and Linoleic Acid. J. Nutr. Biochem. 2018, 57, 121–129. [Google Scholar] [CrossRef]
- Kasatkina, L.A.; Heinemann, A.; Hudz, Y.A.; Thomas, D.; Sturm, E.M. Stearoylethanolamide Interferes with Retrograde Endocannabinoid Signalling and Supports the Blood-Brain Barrier Integrity under Acute Systemic Inflammation. Biochem. Pharmacol. 2020, 174, 113783. [Google Scholar] [CrossRef]
- Ezzili, C.; Otrubova, K.; Boger, D.L. Fatty Acid Amide Signaling Molecules. Bioorg. Med. Chem. Lett. 2010, 20, 5959–5968. [Google Scholar] [CrossRef]
- Schlosburg, J.E.; Kinsey, S.G.; Lichtman, A.H. Targeting Fatty Acid Amide Hydrolase (FAAH) to Treat Pain and Inflammation. AAPS J. 2009, 11, 39–44. [Google Scholar] [CrossRef]
- Boughton-Smith, N.K.; Wallace, J.L.; Morris, G.P.; Whittle, B.J.R. The Effect of Anti-inflammatory Drugs on Eicosanoid Formation in a Chronic Model of Inflammatory Bowel Disease in the Rat. Br. J. Pharmacol. 1988, 94, 65–72. [Google Scholar] [CrossRef]
- Shen, H.; Liu, X.; Jiang, M.; Luo, G.; Wu, Z.; Chen, B.; Li, J.; Liu, L.; Chen, S. Anti-Inflammatory Cembrane-Type Diterpenoids and Prostaglandins from Soft Coral Lobophytum Sarcophytoides. Mar. Drugs 2019, 17, 481. [Google Scholar] [CrossRef]
- Fitc, F.; Tritc, R. Anti-Inflammatory Cyclopentenone Prostaglandins Are Direct Inhibitors of IkB Kinase. Nature 2000, 403, 103–108. [Google Scholar] [CrossRef]
- Cárdeno, A.; Aparicio-Soto, M.; Montserrat-de la Paz, S.; Bermudez, B.; Muriana, F.J.G.; Alarcón-de-la-Lastra, C. Squalene Targets Pro- and Anti-Inflammatory Mediators and Pathways to Modulate over-Activation of Neutrophils, Monocytes and Macrophages. J. Funct. Foods 2015, 14, 779–790. [Google Scholar] [CrossRef]
- Araldi, E.; Fernández-Fuertes, M.; Canfrán-Duque, A.; Tang, W.; Cline, G.W.; Madrigal-Matute, J.; Pober, J.S.; Lasunción, M.A.; Wu, D.; Fernández-Hernando, C.; et al. Lanosterol Modulates TLR4-Mediated Innate Immune Responses in Macrophages. Cell Rep. 2017, 19, 2743–2755. [Google Scholar] [CrossRef]
- Snodgrass, R.G.; Benatzy, Y.; Schmid, T.; Namgaladze, D.; Mainka, M.; Schebb, N.H.; Lütjohann, D.; Brüne, B. Efferocytosis Potentiates the Expression of Arachidonate 15-Lipoxygenase (ALOX15) in Alternatively Activated Human Macrophages through LXR Activation. Cell Death Differ. 2021, 28, 1301–1316. [Google Scholar] [CrossRef] [PubMed]
- Joseph, S.B.; Castrillo, A.; Laffitte, B.A.; Mangelsdorf, D.J.; Tontonoz, P. Reciprocal Regulation of Inflammation and Lipid Metabolism by Liver X Receptors. Nat. Med. 2003, 9, 213–219. [Google Scholar] [CrossRef]
- Kang, Y.; Yang, G.; Zhang, S.; Ross, C.F.; Zhu, M.J. Goji Berry Modulates Gut Microbiota and Alleviates Colitis in IL-10-Deficient Mice. Mol. Nutr. Food Res. 2018, 62, 1800535. [Google Scholar] [CrossRef]
- Moon, C.; Baldridge, M.T.; Wallace, M.A.; Burnham, C.-A.D.; Virgin, H.W.; Stappenbeck, T.S. Vertically Transmitted Fecal IgA Levels Distinguish Extra-Chromosomal Phenotypic Variation. Nature 2015, 521, 90–93. [Google Scholar] [CrossRef]
- Richter, H.; Steinberg, S.; Baron, Y.; Inbart, A. Modified Black Soldier Fly Larvae Oil with Modified Lauric Acid for Treatment against Biofilm Formation and Microorganism Growth. Internationl Application. WO2020234884A1, 21 May 2020. [Google Scholar]
- Kohjima, M.; Enjoji, M.; Higuchi, N.; Kato, M.; Kotoh, K.; Yoshimoto, T.; Fujino, T.; Yada, M.; Yada, R.; Harada, N.; et al. Re-Evaluation of Fatty Acid Metabolism-Related Gene Expression in Nonalcoholic Fatty Liver Disease. Int. J. Mol. Med. 2007, 20, 351–358. [Google Scholar] [CrossRef]
- Ju, X.; Zenke, M.; Hart, D.N.J.; Clark, G.J. CD300a/c Regulate Type i Interferon and TNF-α Secretion by Human Plasmacytoid Dendritic Cells Stimulated with TLR7 and TLR9 Ligands. Blood 2008, 112, 1184–1194. [Google Scholar] [CrossRef]
- Hasumi, H.; Baba, M.; Hong, S.B.; Hasumi, Y.; Huang, Y.; Yao, M.; Valera, V.A.; Linehan, W.M.; Schmidt, L.S. Identification and Characterization of a Novel Folliculin-Interacting Protein FNIP2. Gene 2008, 415, 60–67. [Google Scholar] [CrossRef]
- Hor, Y.T.; Voon, D.C.C.; Koo, J.K.W.; Wang, H.; Lau, W.M.; Ashktorab, H.; Chan, S.L.; Ito, Y. A Role for RUNX3 in Inflammation-Induced Expression of IL23A in Gastric Epithelial Cells. Cell Rep. 2014, 8, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Yamagata, K.; Tusruta, C.; Ohtuski, A.; Tagami, M. Docosahexaenoic Acid Decreases TNF-α-Induced Lectin-like Oxidized Low-Density Lipoprotein Receptor-1 Expression in THP-1 Cells. Prostaglandins Leukot. Essent. Fat. Acids 2014, 90, 125–132. [Google Scholar] [CrossRef]
- Jaitin, D.A.; Kenigsberg, E.; Keren-Shaul, H.; Elefant, N.; Paul, F.; Zaretsky, I.; Mildner, A.; Cohen, N.; Jung, S.; Tanay, A.; et al. Massively Parallel Single-Cell RNA-Seq for Marker-Free Decomposition of Tissues into Cell Types. Science 2014, 343, 772–776. [Google Scholar] [CrossRef]
- Keren-Shaul, H.; Kenigsberg, E.; Jaitin, D.A.; David, E.; Paul, F.; Tanay, A.; Amit, I. MARS-Seq2.0: An Experimental and Analytical Pipeline for Indexed Sorting Combined with Single-Cell RNA Sequencing. Nat. Protoc. 2019, 14, 1841–1862. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet.J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq-A Python Framework to Work with High-Throughput Sequencing Data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Köster, J.; Rahmann, S. Snakemake—A Scalable Bioinformatics Workflow Engine. Bioinformatics 2012, 28, 2520–2522. [Google Scholar] [CrossRef] [PubMed]
- Watrous, J.D.; Niiranen, T.J.; Lagerborg, K.A.; Henglin, M.; Xu, Y.J.; Rong, J.; Sharma, S.; Vasan, R.S.; Larson, M.G.; Armando, A.; et al. Directed Non-Targeted Mass Spectrometry and Chemical Networking for Discovery of Eicosanoids and Related Oxylipins. Cell Chem. Biol. 2019, 26, 433–442.e4. [Google Scholar] [CrossRef]
- Jeengar, M.K.; Thummuri, D.; Magnusson, M.; Naidu, V.G.M. Uridine Ameliorates Dextran Sulfate Sodium (DSS)-Induced Colitis in Mice. Sci. Rep. 2017, 7, 3924. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Shajib, M.S.; Manocha, M.M.; Khan, W.I. Investigating Intestinal Inflammation in DSS-Induced Model of IBD. J. Vis. Exp. 2012, 3678. [Google Scholar] [CrossRef]
- Cao, A.T.; Yao, S.; Gong, B.; Elson, C.O.; Cong, Y. Th17 Cells Upregulate Polymeric Ig Receptor and Intestinal IgA and Contribute to Intestinal Homeostasis. J. Immunol. 2012, 189, 4666–4673. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Compound | Concentration (pg/µL) |
|---|---|---|
| ω3 metabolite | 13S-hydroxy-octadecadienoic acid (13S-HOTrE) | 31,505 |
| 17,18-Dihydroxyeicosatetraenoic acid (17,18-DiHETE) | 1654 | |
| 12S-hydroxyeicosapentaenoic acid (12S-HEPE) | 150 | |
| ω6 metabolite | 9,10-dihydroxyoctadecenoic acid (9,10-DiHOME) | 91,118 |
| 9,10-epoxyoctadecenoic acid (9,10-EpOME) | 6390 | |
| 12-hydroxyoctadecadienoic acid (12-HODE) | 12,058 | |
| 11,12-epoxyeicosatrienoic acid (11,12-EET) | 224 | |
| Lipoxin A4 (LXA4) | 1048 | |
| 15S-hydroperoxyeicosatetraenoic acid (15S-HpETE) | 989 | |
| 5S-hydroxyeicosatetraenoic acid (5S-HETE) | 159 | |
| Prostanoid | Prostaglandin A1 (PGA1) | 330 |
| Prostaglandin B2 (PGB2) | 1348 | |
| Prostaglandin E2 (PGE2) | 1338 | |
| 6-keto Prostaglandin F1α (6-keto PGF1α) | 2898 | |
| Thromboxane B1 (TXB1) | 5991 | |
| Leukotriene | Leukotriene B3 (LTB3) | 427 |
| Leukotriene C4 (LTC4) | 222 | |
| N-acylamide | N-Linoleoyl Leucine | 74,687 |
| N-Oleoyl Valine | 13,233 | |
| N-Palmitoyl Glycine (PalGly) | 10,007 | |
| N-acylethanol amine (NAE) | Stearoyl Ethanolamide (SEA) | 16,739 |
| Oleoyl Ethanolamide (OEA) | 2608 | |
| Palmitoyl Ethanolamide (PEA) | 383 | |
| N-acyl serotonin | Oleoyl Serotonin (OA-5-HT) | 7954 |
| Group | Compound | Concentration (nmol/g) |
|---|---|---|
| Sterols | Lanosterol | 20.40 |
| Desmosterol | 2.17 | |
| 7-Dehydrocholesterol (7-DHC) | 0.56 | |
| Dihydrolanosterol | 0.12 | |
| Zymosterol | 2.31 | |
| Zymostenol | 0.83 | |
| Ubiquinones | CoQ-06 | 2.02 |
| CoQ-07 | 2.76 | |
| CoQ-08 | 1.89 | |
| CoQ-09 | 24.19 | |
| CoQ-10 | 0.68 | |
| CoQ-11 | 0.03 | |
| Squalene | 37.82 | |
| Dolichols (13 to 21 isoprene units) | 42.71 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Richter, H.; Gover, O.; Schwartz, B. Anti-Inflammatory Activity of Black Soldier Fly Oil Associated with Modulation of TLR Signaling: A Metabolomic Approach. Int. J. Mol. Sci. 2023, 24, 10634. https://doi.org/10.3390/ijms241310634
Richter H, Gover O, Schwartz B. Anti-Inflammatory Activity of Black Soldier Fly Oil Associated with Modulation of TLR Signaling: A Metabolomic Approach. International Journal of Molecular Sciences. 2023; 24(13):10634. https://doi.org/10.3390/ijms241310634
Chicago/Turabian StyleRichter, Hadas, Ofer Gover, and Betty Schwartz. 2023. "Anti-Inflammatory Activity of Black Soldier Fly Oil Associated with Modulation of TLR Signaling: A Metabolomic Approach" International Journal of Molecular Sciences 24, no. 13: 10634. https://doi.org/10.3390/ijms241310634
APA StyleRichter, H., Gover, O., & Schwartz, B. (2023). Anti-Inflammatory Activity of Black Soldier Fly Oil Associated with Modulation of TLR Signaling: A Metabolomic Approach. International Journal of Molecular Sciences, 24(13), 10634. https://doi.org/10.3390/ijms241310634