

Mismatch Repair Protein Msh2 Is Necessary for Macronuclear Stability and Micronuclear Division in Tetrahymena thermophila


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
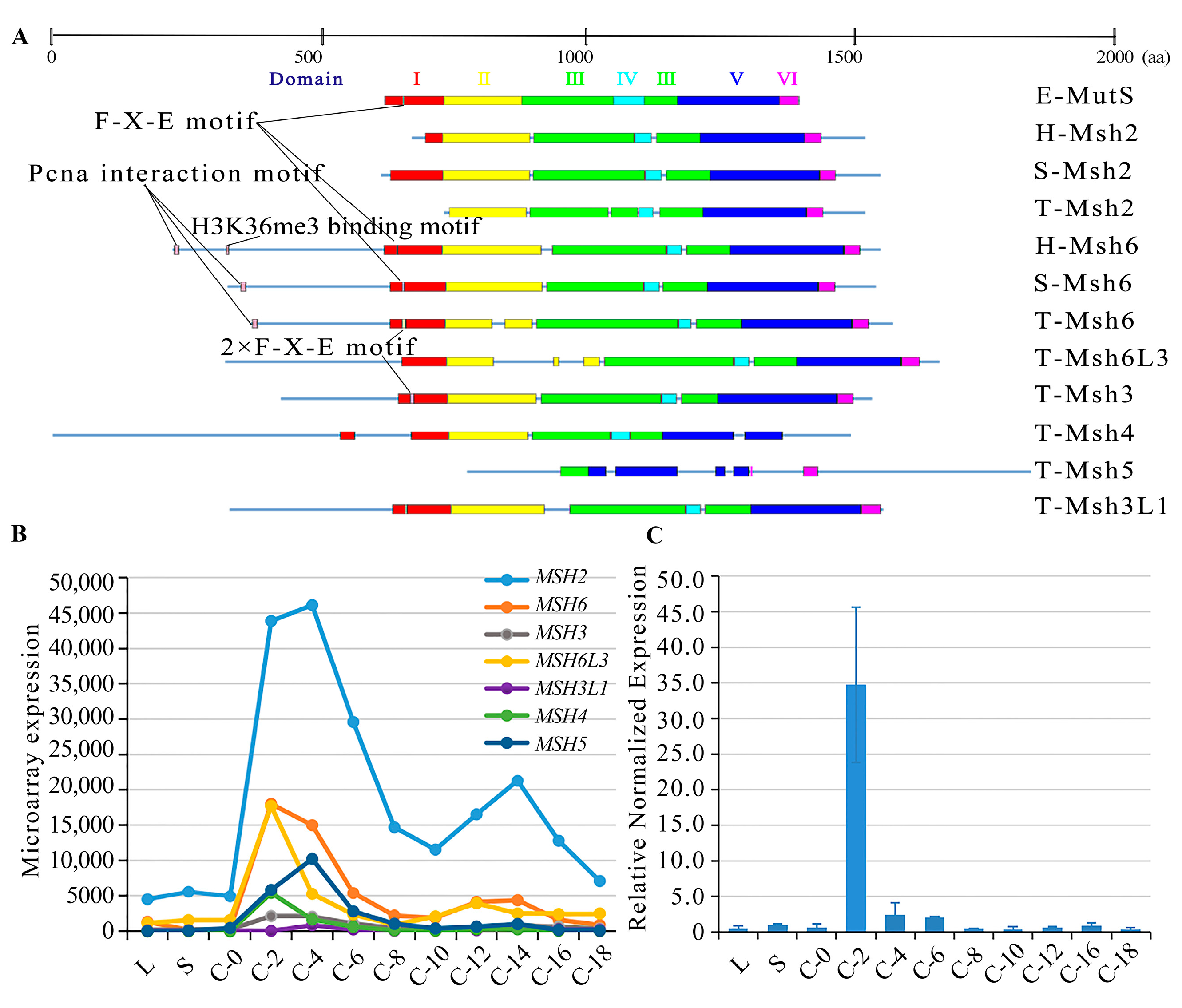
2.1. Characterization of MutS Proteins in T. thermophila
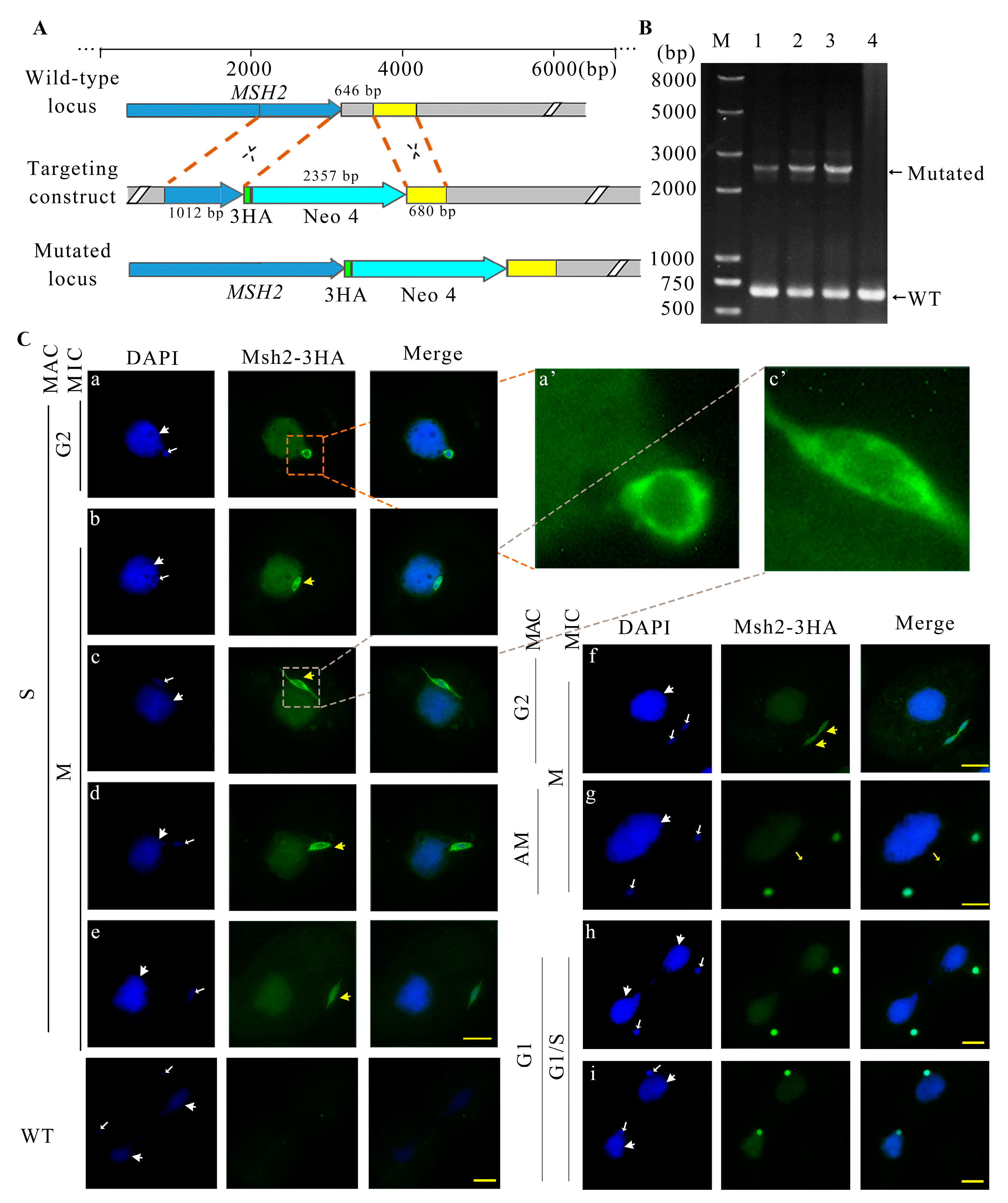
2.2. Msh2-3HA Was Localized in the MAC and MIC during Vegetative Growth
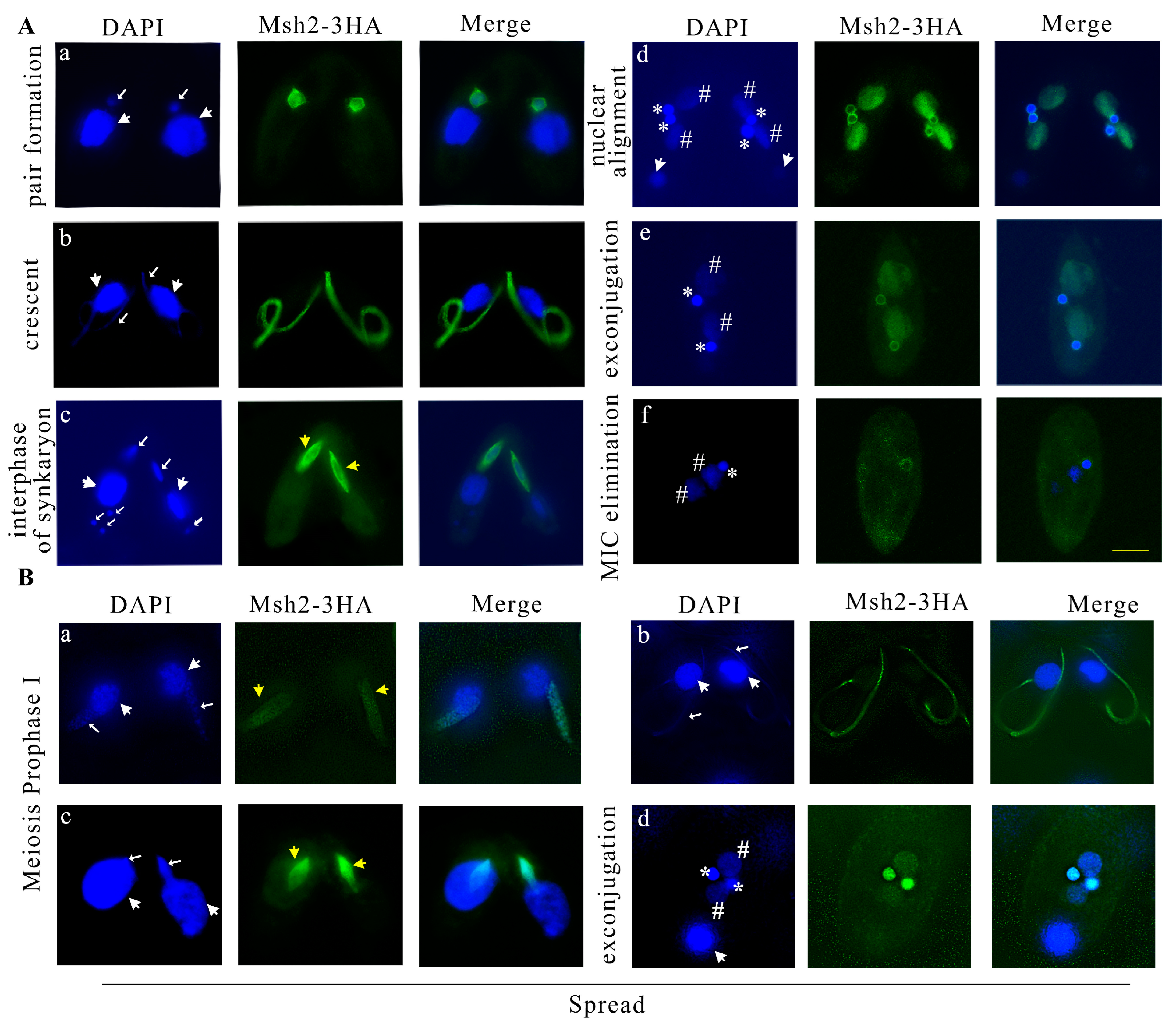
2.3. Msh2 Localized in the MIC and New MAC during Sexual Development
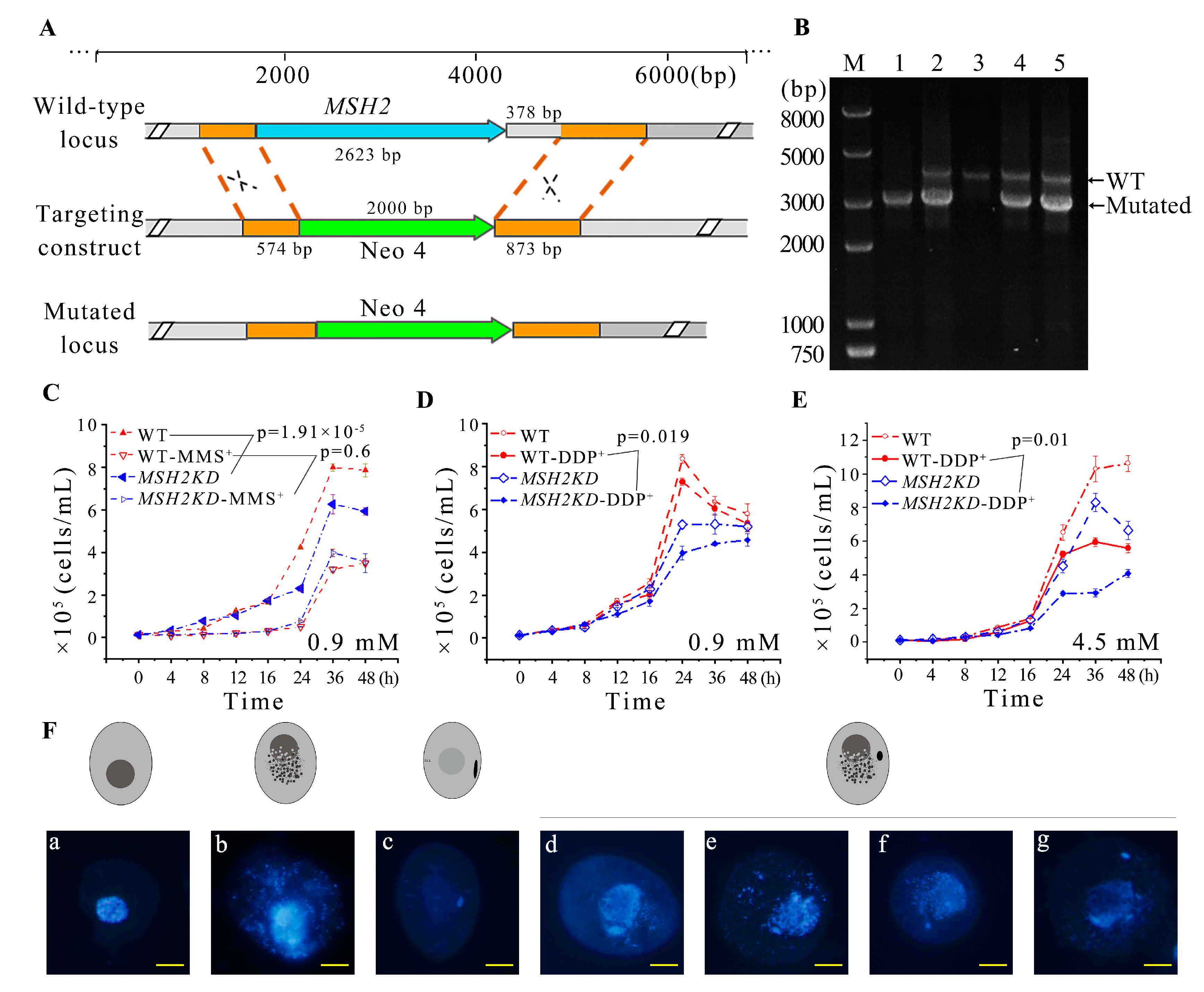
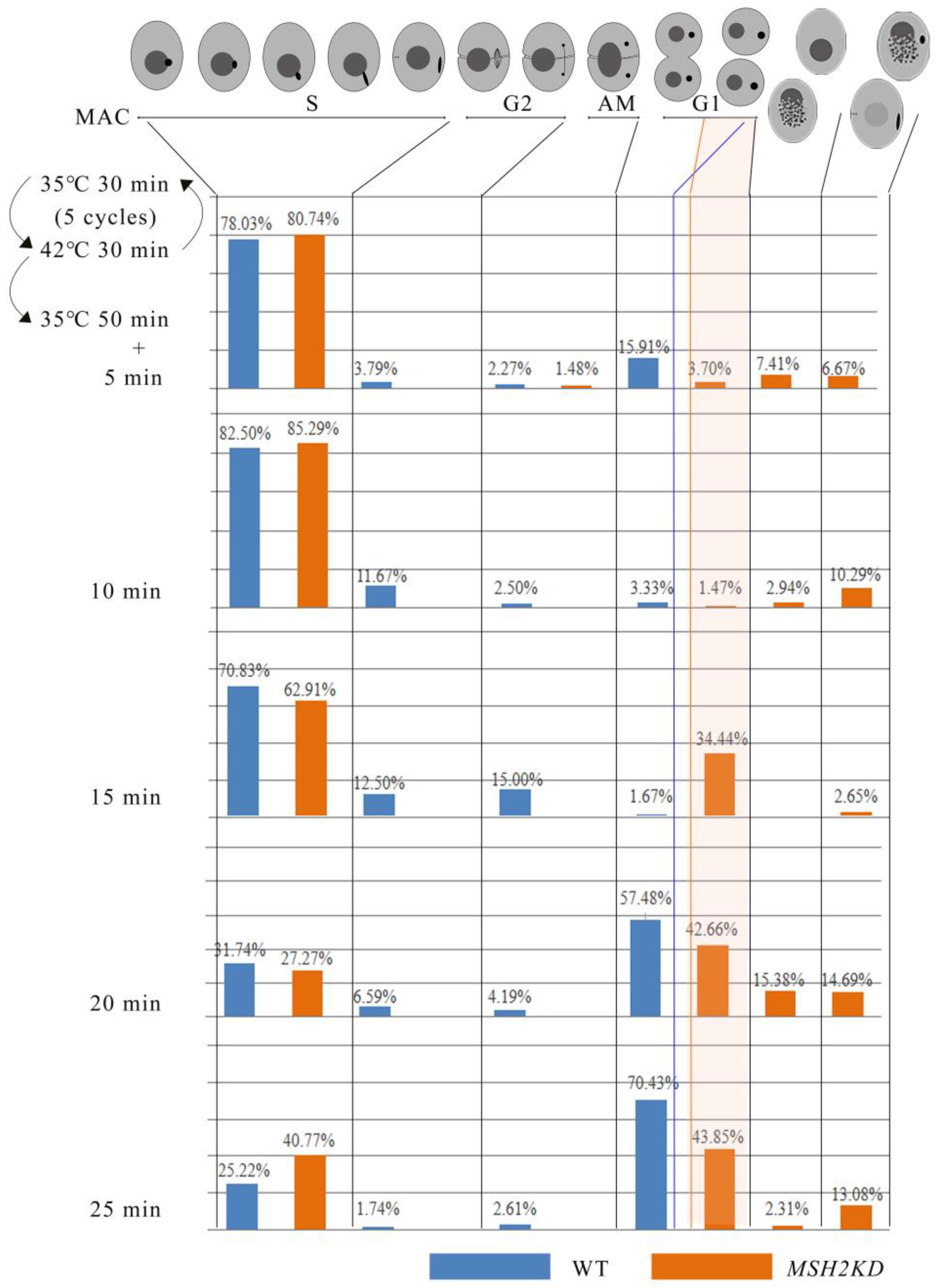
2.4. MSH2 Knocking Down Affected the Division of Nuclei during Vegetative Growing Stage
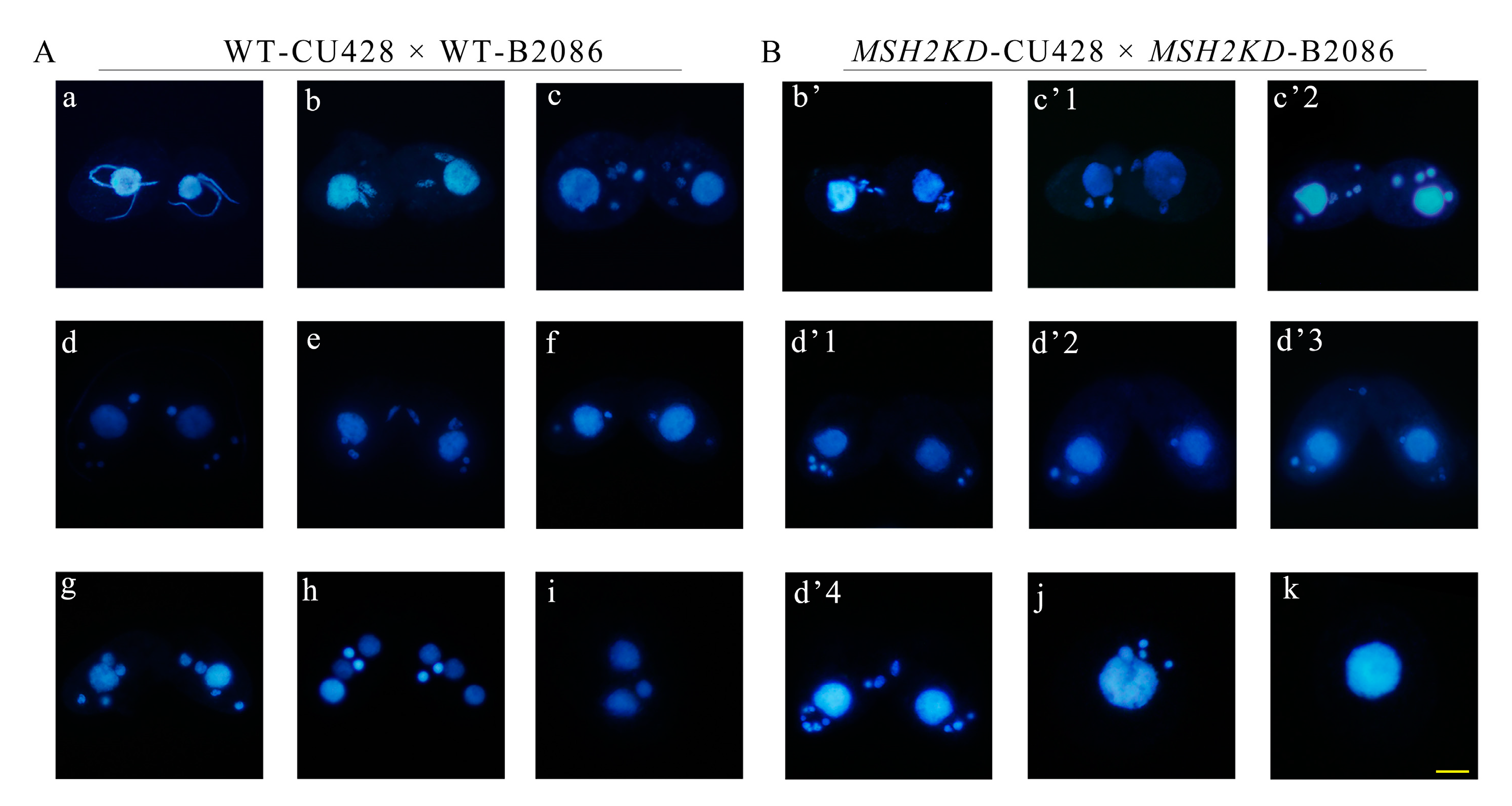
2.5. Knocking Down MSH2 Affected Sexual Development
2.6. Conditional Knockdown of MSH2 Affected Sexual Development during Conjugation
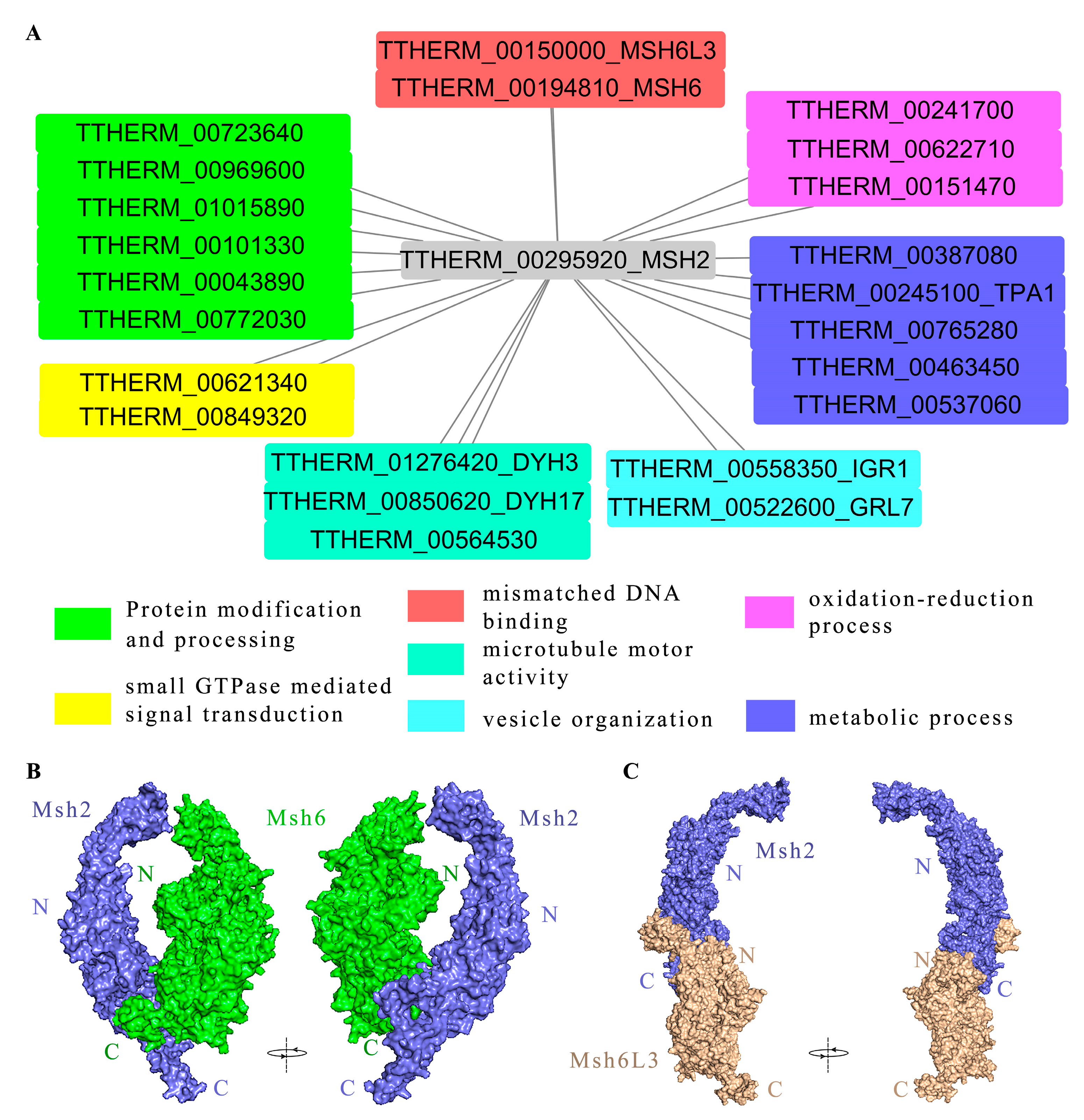
2.7. Msh2 Interacted with MMR-Dependent and MMR-Independent Factors
3. Discussion
4. Materials and Methods
4.1. Culture, Starvation, and Pairing of T. thermophila
4.2. Identification of MutS Homologous Proteins
4.3. Construct of Msh2-3HA Mutant Cell Line
4.4. Immunofluorescent Localization Analysis
4.5. Construction of MSH2 Knockout Mutants
4.6. Synchronization of Cell Division
4.7. Nuclear Development
4.8. Construction of MSH2 Conditional Knockdown Mutants
4.9. qRT-PCR Analysis
4.10. Co-Immunoprecipitation and Mass Spectrometry
4.11. Protein–Protein Docking
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kunkel, T.A.; Erie, D.A. DNA mismatch repair. Annu. Rev. Biochem. 2005, 74, 681–710. [Google Scholar] [CrossRef] [PubMed]
- Fink, D.; Aebi, S.; Howell, S.B. The role of DNA mismatch repair in drug resistance. Clin. Cancer Res. 1998, 4, 1–6. [Google Scholar] [PubMed]
- Hsieh, P.; Yamane, K. DNA mismatch repair: Molecular mechanism, cancer, and ageing. Mech. Ageing Dev. 2008, 129, 391–407. [Google Scholar] [CrossRef]
- Kunkel, T.A.; Erie, D.A. Eukaryotic Mismatch Repair in Relation to DNA Replication. Annu. Rev. Genet. 2015, 49, 291–313. [Google Scholar] [CrossRef] [PubMed]
- Guerrette, S.; Wilson, T.; Gradia, S.; Fishel, R. Interactions of human hMSH2 with hMSH3 and hMSH2 with hMSH6: Examination of mutations found in hereditary nonpolyposis colorectal cancer. Mol. Cell Biol. 1998, 18, 6616–6623. [Google Scholar] [CrossRef]
- Karran, P. Thiopurines, DNA damage, DNA repair and therapy-related cancer. Br. Med. Bull. 2006, 79–80, 153–170. [Google Scholar] [CrossRef]
- Li, G.-M. Mechanisms and functions of DNA mismatch repair. Cell Res. 2008, 18, 85–98. [Google Scholar] [CrossRef]
- Zhang, M.; Xiang, S.; Joo, H.Y.; Wang, L.; Williams, K.A.; Liu, W.; Hu, C.; Tong, D.; Haakenson, J.; Wang, C.; et al. HDAC6 deacetylates and ubiquitinates MSH2 to maintain proper levels of MutSα. Mol. Cell 2014, 55, 31–46. [Google Scholar] [CrossRef]
- Zhang, H.; Xiao, X.; Wei, W.; Huang, C.; Wang, M.; Wang, L.; He, Y.; Sun, J.; Jiang, Y.; Jiang, G.; et al. CircLIFR synergizes with MSH2 to attenuate chemoresistance via MutSα/ATM-p73 axis in bladder cancer. Mol. Cancer 2021, 20, 70. [Google Scholar] [CrossRef]
- Pabla, N.; Ma, Z.; McIlhatton, M.A.; Fishel, R.; Dong, Z. hMSH2 recruits ATR to DNA damage sites for activation during DNA damage-induced apoptosis. J. Biol. Chem. 2011, 286, 10411–10418. [Google Scholar] [CrossRef]
- Lin, D.P.; Wang, Y.; Scherer, S.J.; Clark, A.B.; Yang, K.; Avdievich, E.; Jin, B.; Werling, U.; Parris, T.; Kurihara, N.; et al. An Msh2 point mutation uncouples DNA mismatch repair and apoptosis. Cancer Res. 2004, 64, 517–522. [Google Scholar] [CrossRef]
- Chakraborty, U.; Shen, Z.J.; Tyler, J. Chaperoning histones at the DNA repair dance. DNA Repair 2021, 108, 103240. [Google Scholar] [CrossRef] [PubMed]
- Kadyrova, L.Y.; Blanko, E.R.; Kadyrov, F.A. CAF-I-dependent control of degradation of the discontinuous strands during mismatch repair. Proc. Natl. Acad. Sci. USA 2011, 108, 2753–2758. [Google Scholar] [CrossRef]
- Rodriges Blanko, E.; Kadyrova, L.Y.; Kadyrov, F.A. DNA Mismatch Repair Interacts with CAF-1 and ASF1A-H3-H4-dependent Histone (H3-H4)2 Tetramer Deposition. J. Biol. Chem. 2016, 291, 9203–9217. [Google Scholar] [CrossRef]
- Campbell, M.R.; Wang, Y.; Andrew, S.E.; Liu, Y. Msh2 deficiency leads to chromosomal abnormalities, centrosome amplification, and telomere capping defect. Oncogene 2006, 25, 2531–2536. [Google Scholar] [CrossRef] [PubMed]
- Stambuk, S.; Radman, M. Mechanism and control of interspecies recombination in Escherichia coli. I. Mismatch repair, methylation, recombination and replication functions. Genetics 1998, 150, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Honda, M.; Okuno, Y.; Hengel, S.R.; Martín-López, J.V.; Cook, C.P.; Amunugama, R.; Soukup, R.J.; Subramanyam, S.; Fishel, R.; Spies, M. Mismatch repair protein hMSH2-hMSH6 recognizes mismatches and forms sliding clamps within a D-loop recombination intermediate. Proc. Natl. Acad. Sci. USA 2014, 111, E316–E325. [Google Scholar] [CrossRef]
- de los Santos, T.; Hunter, N.; Lee, C.; Larkin, B.; Loidl, J.; Hollingsworth, N.M. The Mus81/Mms4 endonuclease acts independently of double-Holliday junction resolution to promote a distinct subset of crossovers during meiosis in budding yeast. Genetics 2003, 164, 81–94. [Google Scholar] [CrossRef]
- Snowden, T.; Acharya, S.; Butz, C.; Berardini, M.; Fishel, R. hMSH4-hMSH5 recognizes Holliday Junctions and forms a meiosis-specific sliding clamp that embraces homologous chromosomes. Mol. Cell 2004, 15, 437–451. [Google Scholar] [CrossRef]
- Shodhan, A.; Lukaszewicz, A.; Novatchkova, M.; Loidl, J. Msh4 and Msh5 function in SC-independent chiasma formation during the streamlined meiosis of Tetrahymena. Genetics 2014, 198, 983–993. [Google Scholar] [CrossRef]
- Oh, J.M.; Kang, Y.; Park, J.; Sung, Y.; Kim, D.; Seo, Y.; Lee, E.A.; Ra, J.S.; Amarsanaa, E.; Park, Y.U.; et al. MSH2-MSH3 promotes DNA end resection during homologous recombination and blocks polymerase theta-mediated end-joining through interaction with SMARCAD1 and EXO1. Nucleic. Acids. Res. 2023; online ahead of print. [Google Scholar] [CrossRef]
- Barroso-González, J.; García-Expósito, L.; Galaviz, P.; Lynskey, M.L.; Allen, J.A.M.; Hoang, S.; Watkins, S.C.; Pickett, H.A.; O’Sullivan, R.J. Anti-recombination function of MutSα restricts telomere extension by ALT-associated homology-directed repair. Cell Rep. 2021, 37, 110088. [Google Scholar] [CrossRef] [PubMed]
- Young, S.J.; Sebald, M.; Shah Punatar, R.; Larin, M.; Masino, L.; Rodrigo-Brenni, M.C.; Liang, C.C.; West, S.C. MutSβ Stimulates Holliday Junction Resolution by the SMX Complex. Cell Rep. 2020, 33, 108289. [Google Scholar] [CrossRef] [PubMed]
- Kolodner, R.D. A personal historical view of DNA mismatch repair with an emphasis on eukaryotic DNA mismatch repair. DNA Repair 2016, 38, 3–13. [Google Scholar] [CrossRef]
- Loidl, J. Tetrahymena meiosis: Simple yet ingenious. PLoS Genet. 2021, 17, e1009627. [Google Scholar] [CrossRef]
- Cole, E.; Sugai, T. Chapter 7—Developmental progression of Tetrahymena through the cell cycle and conjugation. In Methods in Cell Biology; Collins, K., Ed.; Academic Press: Cambridge, MA, USA, 2012; Volume 109, pp. 177–236. [Google Scholar]
- Noto, T.; Mochizuki, K. Whats, hows and whys of programmed DNA elimination in Tetrahymena. Open Biol. 2017, 7, 170–172. [Google Scholar] [CrossRef]
- Orias, E.; Cervantes, M.D.; Hamilton, E.P. Tetrahymena thermophila, a unicellular eukaryote with separate germline and somatic genomes. Res. Microbiol. 2011, 162, 578–586. [Google Scholar] [CrossRef]
- Lukaszewicz, A.; Howard-Till, R.A.; Novatchkova, M.; Mochizuki, K.; Loidl, J. MRE11 and COM1/SAE2 are required for double-strand break repair and efficient chromosome pairing during meiosis of the protist Tetrahymena. Chromosoma 2010, 119, 505–518. [Google Scholar] [CrossRef]
- Loidl, J.; Lorenz, A. DNA double-strand break formation and repair in Tetrahymena meiosis. Semin. Cell Dev. Biol. 2016, 54, 126–134. [Google Scholar] [CrossRef]
- Loidl, J.; Mochizuki, K. Tetrahymena meiotic nuclear reorganization is induced by a checkpoint kinase-dependent response to DNA damage. Mol. Biol. Cell 2009, 20, 2428–2437. [Google Scholar] [CrossRef] [PubMed]
- Lukaszewicz, A.; Howard-Till, R.A.; Loidl, J. Mus81 nuclease and Sgs1 helicase are essential for meiotic recombination in a protist lacking a synaptonemal complex. Nucleic Acids Res. 2013, 41, 9296–9309. [Google Scholar] [CrossRef] [PubMed]
- Fishel, R.; Lescoe, M.K.; Rao, M.R.; Copeland, N.G.; Jenkins, N.A.; Garber, J.; Kane, M.; Kolodner, R. The human mutator gene homolog MSH2 and its association with hereditary nonpolyposis colon cancer. Cell 1993, 75, 1027–1038. [Google Scholar] [CrossRef]
- Kolodner, R.D.; Marsischky, G.T. Eukaryotic DNA mismatch repair. Curr. Opin. Genet. Dev. 1999, 9, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Iyer, R.R.; Pluciennik, A.; Burdett, V.; Modrich, P.L. DNA mismatch repair: Functions and mechanisms. Chem. Rev. 2006, 106, 302–323. [Google Scholar] [CrossRef]
- Svetlanov, A.; Cohen, P.E. Mismatch repair proteins, meiosis, and mice: Understanding the complexities of mammalian meiosis. Exp. Cell Res. 2004, 296, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.Y.; Culligan, K.; Lamers, M.; Hays, J. Dissimilar mispair-recognition spectra of Arabidopsis DNA-mismatch-repair proteins MSH2*MSH6 (MutSalpha) and MSH2*MSH7 (MutSgamma). Nucleic Acids Res. 2003, 31, 6027–6034. [Google Scholar] [CrossRef]
- Novak, J.E.; Ross-Macdonald, P.B.; Roeder, G.S. The budding yeast Msh4 protein functions in chromosome synapsis and the regulation of crossover distribution. Genetics 2001, 158, 1013–1025. [Google Scholar] [CrossRef]
- Reyes, G.X.; Schmidt, T.T.; Kolodner, R.D.; Hombauer, H. New insights into the mechanism of DNA mismatch repair. Chromosoma 2015, 124, 443–462. [Google Scholar] [CrossRef]
- Lamers, M.H.; Perrakis, A.; Enzlin, J.H.; Winterwerp, H.H.; de Wind, N.; Sixma, T.K. The crystal structure of DNA mismatch repair protein MutS binding to a G x T mismatch. Nature 2000, 407, 711–717. [Google Scholar] [CrossRef]
- Parfrey, L.W.; Grant, J.; Tekle, Y.I.; Lasek-Nesselquist, E.; Morrison, H.G.; Sogin, M.L.; Patterson, D.J.; Katz, L.A. Broadly sampled multigene analyses yield a well-resolved eukaryotic tree of life. Syst. Biol. 2010, 59, 518–533. [Google Scholar] [CrossRef]
- Li, F.; Tian, L.; Gu, L.; Li, G.M. Evidence that nucleosomes inhibit mismatch repair in eukaryotic cells. J. Biol. Chem. 2009, 284, 33056–33061. [Google Scholar] [CrossRef] [PubMed]
- Gorman, J.; Plys, A.J.; Visnapuu, M.L.; Alani, E.; Greene, E.C. Visualizing one-dimensional diffusion of eukaryotic DNA repair factors along a chromatin lattice. Nat. Struct. Mol. Biol. 2010, 17, 932–938. [Google Scholar] [CrossRef] [PubMed]
- Schöpf, B.; Bregenhorn, S.; Quivy, J.P.; Kadyrov, F.A.; Almouzni, G.; Jiricny, J. Interplay between mismatch repair and chromatin assembly. Proc. Natl. Acad. Sci. USA 2012, 109, 1895–1900. [Google Scholar] [CrossRef] [PubMed]
- Howard-Till, R.A.; Lukaszewicz, A.; Loidl, J. The recombinases Rad51 and Dmc1 play distinct roles in DNA break repair and recombination partner choice in the meiosis of Tetrahymena. PLoS Genet. 2011, 7, e1001359. [Google Scholar] [CrossRef]
- Tang, Z.; Takahashi, Y. Analysis of Protein–Protein Interaction by Co-IP in Human Cells. In Two-Hybrid Systems: Methods and Protocols; Oñate-Sánchez, L., Ed.; Springer: New York, NY, USA, 2018; pp. 289–296. [Google Scholar]
- Edamatsu, M. Functional characterization of lethal P-loop mutations in Tetrahymena outer arm dynein (Dyh3p). Biochem. Biophys. Res. Commun. 2018, 496, 1382–1388. [Google Scholar] [CrossRef]
- Obmolova, G.; Ban, C.; Hsieh, P.; Yang, W. Crystal structures of mismatch repair protein MutS and its complex with a substrate DNA. Nature 2000, 407, 703–710. [Google Scholar] [CrossRef]
- Mazur, D.J.; Mendillo, M.L.; Kolodner, R.D. Inhibition of Msh6 ATPase activity by mispaired DNA induces a Msh2(ATP)-Msh6(ATP) state capable of hydrolysis-independent movement along DNA. Mol. Cell 2006, 22, 39–49. [Google Scholar] [CrossRef]
- Christmann, M.; Kaina, B. Nuclear translocation of mismatch repair proteins MSH2 and MSH6 as a response of cells to alkylating agents. J. Biol. Chem. 2000, 275, 36256–36262. [Google Scholar] [CrossRef]
- Doerder, F.P.; Debault, L.E. Cytofluorimetric analysis of nuclear DNA during meiosis, fertilization and macronuclear development in the ciliate Tetrahymena pyriformis, syngen 1. J. Cell Sci. 1975, 17, 471–493. [Google Scholar] [CrossRef]
- Knudsen, N.; Andersen, S.D.; Lützen, A.; Nielsen, F.C.; Rasmussen, L.J. Nuclear translocation contributes to regulation of DNA excision repair activities. DNA Repair 2009, 8, 682–689. [Google Scholar] [CrossRef]
- Hayes, A.P.; Sevi, L.A.; Feldt, M.C.; Rose, M.D.; Gammie, A.E. Reciprocal regulation of nuclear import of the yeast MutSalpha DNA mismatch repair proteins Msh2 and Msh6. DNA Repair 2009, 8, 739–751. [Google Scholar] [CrossRef]
- Knudsen, N.O.; Nielsen, F.C.; Vinther, L.; Bertelsen, R.; Holten-Andersen, S.; Liberti, S.E.; Hofstra, R.; Kooi, K.; Rasmussen, L.J. Nuclear localization of human DNA mismatch repair protein exonuclease 1 (hEXO1). Nucleic Acids Res. 2007, 35, 2609–2619. [Google Scholar] [CrossRef]
- Iwamoto, M.; Mori, C.; Osakada, H.; Koujin, T.; Hiraoka, Y.; Haraguchi, T. Nuclear localization signal targeting to macronucleus and micronucleus in binucleated ciliate Tetrahymena thermophila. Genes. Cells 2018, 23, 568–579. [Google Scholar] [CrossRef] [PubMed]
- Vaisman, A.; Varchenko, M.; Umar, A.; Kunkel, T.A.; Risinger, J.I.; Barrett, J.C.; Hamilton, T.C.; Chaney, S.G. The role of hMLH1, hMSH3, and hMSH6 defects in cisplatin and oxaliplatin resistance: Correlation with replicative bypass of platinum-DNA adducts. Cancer Res. 1998, 58, 3579–3585. [Google Scholar]
- Plotz, G.; Piiper, A.; Wormek, M.; Zeuzem, S.; Raedle, J. Analysis of the human MutLalpha.MutSalpha complex. Biochem. Biophys. Res. Commun. 2006, 340, 852–859. [Google Scholar] [CrossRef]
- Gorovsky, M.A.; Yao, M.C.; Keevert, J.B.; Pleger, G.L. Isolation of micro- and macronuclei of Tetrahymena pyriformis. Methods Cell Biol. 1975, 9, 311–327. [Google Scholar] [CrossRef] [PubMed]
- Bruns, P.J.; Brussard, T.B. Pair formation in tetrahymena pyriformis, an inducible developmental system. J Exp Zool. 1974, 188, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, X.; Song, W.; Wang, W.; Gao, S. Cyclin Cyc2p is required for micronuclear bouquet formation in Tetrahymena thermophila. Sci. China. Life. Sci. 2019, 62, 668–680. [Google Scholar] [CrossRef]
- Holz, G.G.; Scherbaum, O.H.; Williams, N. The arrest of mitosis and stomatogenesis during temperature-induction of synchronous division in Tetrahymena pyriformis, mating type 1, variety 1. Exp. Cell Res. 1957, 13, 618–621. [Google Scholar] [CrossRef]
- Lian, Y.; Hao, H.; Xu, J.; Bo, T.; Liang, A.; Wang, W. The histone chaperone Nrp1 is required for chromatin stability and nuclear division in Tetrahymena thermophila. Epigenet. Chromatin. 2021, 14, 34. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Carlson, A.; Sinitcyn, P.; Mann, M.; Cox, J. Visualization of LC-MS/MS proteomics data in MaxQuant. Proteomics 2015, 15, 1453–1456. [Google Scholar] [CrossRef] [PubMed]
- Schwanhausser, B.; Busse, D.; Li, N.; Dittmar, G.; Schuchhardt, J.; Wolf, J.; Chen, W.; Selbach, M. Global quantification of mammalian gene expression control. Nature 2011, 473, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Nabeel-Shah, S.; Garg, J.; Saettone, A.; Ashraf, K.; Lee, H.; Wahab, S.; Ahmed, N.; Fine, J.; Derynck, J.; Pu, S.; et al. Functional characterization of RebL1 highlights the evolutionary conservation of oncogenic activities of the RBBP4/7 orthologue in Tetrahymena thermophila. Nucleic Acids Res. 2021, 49, 6196–6212. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, A.K.; Stefans, M.; Christopher, M.Y.; Mark, N.W.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Xue, Y.; Yang, S.; Bo, T.; Xu, J.; Wang, W. Mismatch Repair Protein Msh2 Is Necessary for Macronuclear Stability and Micronuclear Division in Tetrahymena thermophila. Int. J. Mol. Sci. 2023, 24, 10559. https://doi.org/10.3390/ijms241310559
Wang L, Xue Y, Yang S, Bo T, Xu J, Wang W. Mismatch Repair Protein Msh2 Is Necessary for Macronuclear Stability and Micronuclear Division in Tetrahymena thermophila. International Journal of Molecular Sciences. 2023; 24(13):10559. https://doi.org/10.3390/ijms241310559
Chicago/Turabian StyleWang, Lin, Yuhuan Xue, Sitong Yang, Tao Bo, Jing Xu, and Wei Wang. 2023. "Mismatch Repair Protein Msh2 Is Necessary for Macronuclear Stability and Micronuclear Division in Tetrahymena thermophila" International Journal of Molecular Sciences 24, no. 13: 10559. https://doi.org/10.3390/ijms241310559
APA StyleWang, L., Xue, Y., Yang, S., Bo, T., Xu, J., & Wang, W. (2023). Mismatch Repair Protein Msh2 Is Necessary for Macronuclear Stability and Micronuclear Division in Tetrahymena thermophila. International Journal of Molecular Sciences, 24(13), 10559. https://doi.org/10.3390/ijms241310559