
Antigenic Cartography Indicates That the Omicron BA.1 and BA.4/BA.5 Variants Remain Antigenically Distant to Ancestral SARS-CoV-2 after Sputnik V Vaccination Followed by Homologous (Sputnik V) or Heterologous (Comirnaty) Revaccination

 and
and
Abstract
1. Introduction
2. Results
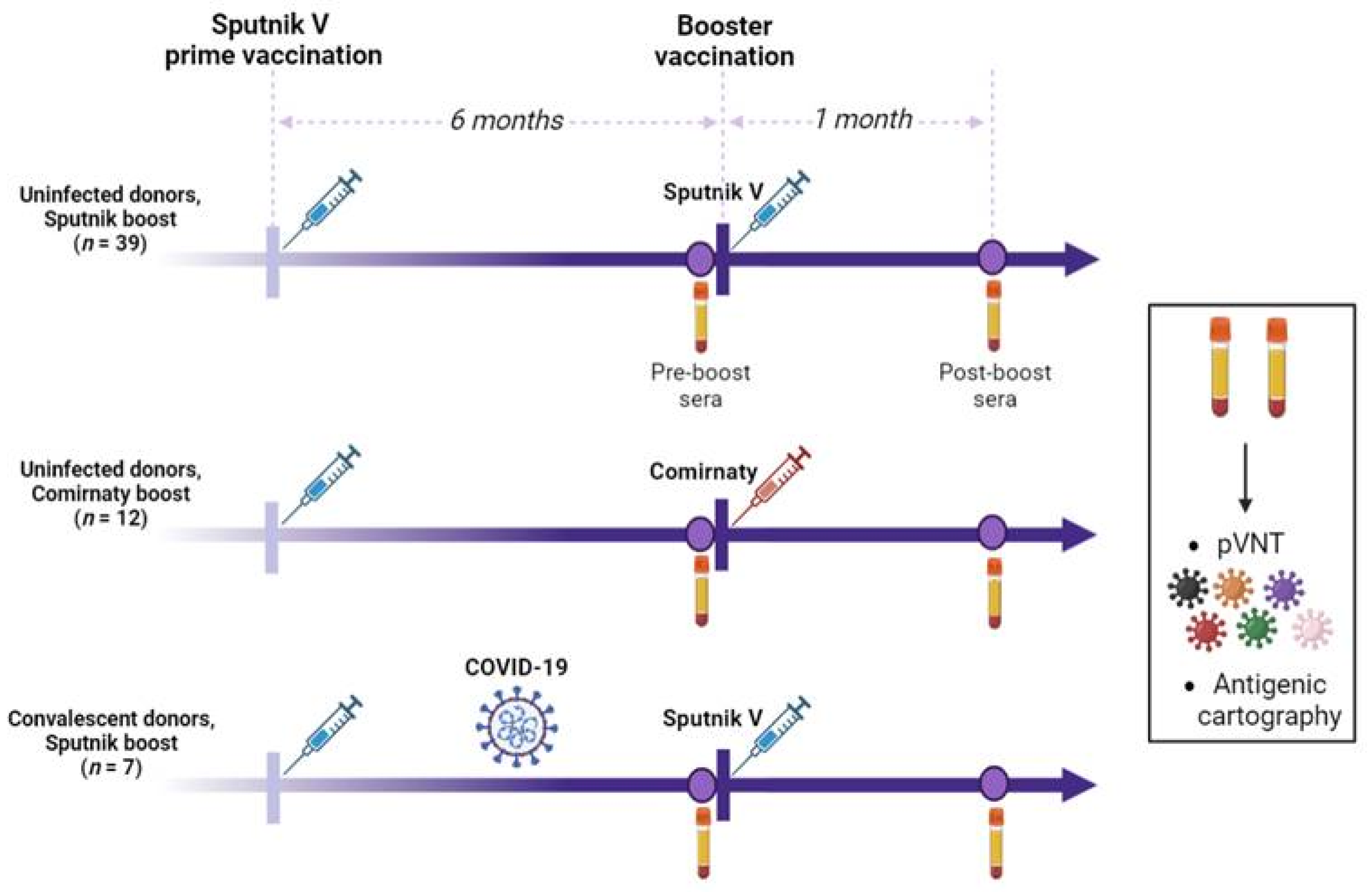
2.1. Study Design
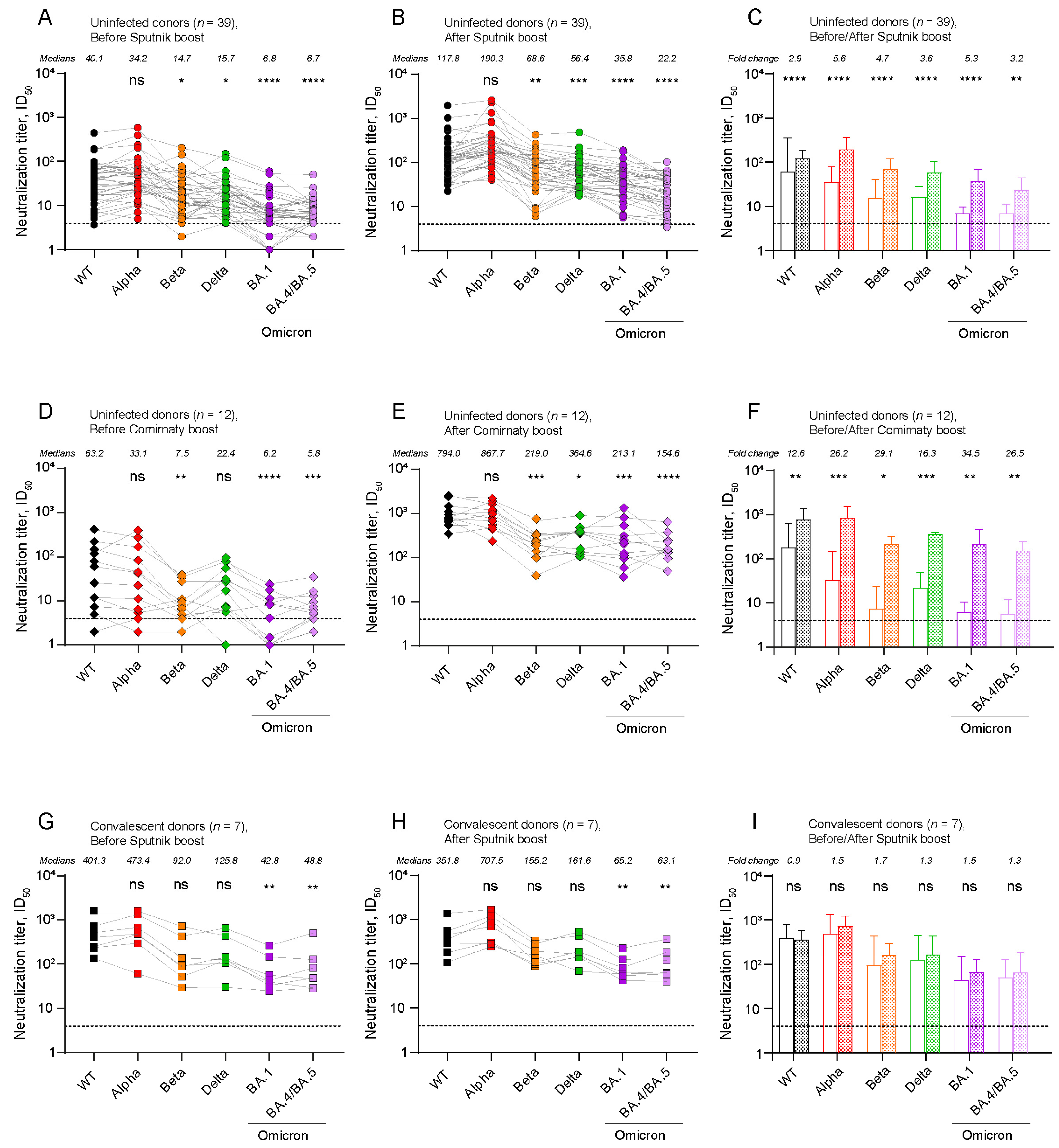
2.2. The Level of Post-Boost SARS-CoV-2 Neutralizing Antibodies (NAbs) Depends on the Type of Booster Vaccine and the History of Exposure to SARS-CoV-2
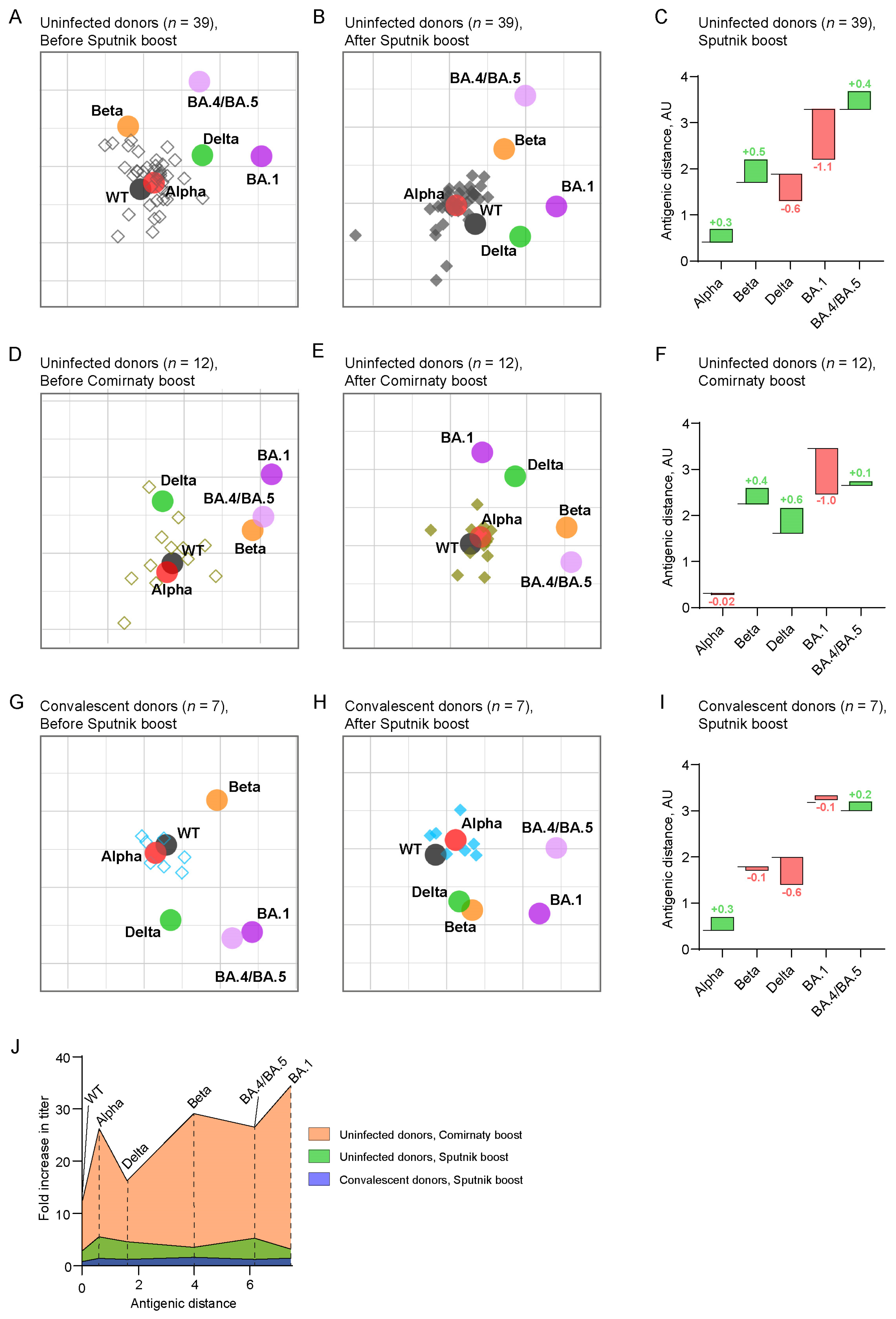
2.3. Antigenic Maps Reveal the Emergence of Booster-Induced Cross-Neutralizing Antibodies
3. Discussion
4. Materials and Methods
4.1. Ethics
4.2. Volunteers and Samples Collection
4.3. SARS-CoV-2 Pseudoviral Particles Production
4.4. Pseudotyped Virus Neutralization Assay
4.5. Antigenic Cartography
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gattinger, P.; Ohradanova-Repic, A.; Valenta, R. Importance, Applications and Features of Assays Measuring SARS-CoV-2 Neutralizing Antibodies. Int. J. Mol. Sci. 2023, 24, 5352. [Google Scholar] [CrossRef]
- Sholukh, A.M.; Fiore-Gartland, A.; Ford, E.S.; Miner, M.D.; Hou, Y.J.; Tse, L.V.; Kaiser, H.; Zhu, H.; Lu, J.; Madarampalli, B.; et al. Evaluation of Cell-Based and Surrogate SARS-CoV-2 Neutralization Assays. J. Clin. Microbiol. 2021, 59, e00527-21. [Google Scholar] [CrossRef]
- Smith, D.J.; Lapedes, A.S.; De Jong, J.C.; Bestebroer, T.M.; Rimmelzwaan, G.F.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Mapping the Antigenic and Genetic Evolution of Influenza Virus. Science 2004, 305, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Fonville, J.M.; Wilks, S.H.; James, S.L.; Fox, A.; Ventresca, M.; Aban, M.; Xue, L.; Jones, T.C.; Le, N.M.H.; Pham, Q.T.; et al. Antibody Landscapes after Influenza Virus Infection or Vaccination. Science 2014, 346, 996–1000. [Google Scholar] [CrossRef] [PubMed]
- Mykytyn, A.Z.; Rissmann, M.; Kok, A.; Rosu, M.E.; Schipper, D.; Breugem, T.I.; van den Doel, P.B.; Chandler, F.; Bestebroer, T.; de Wit, M.; et al. Antigenic Cartography of SARS-CoV-2 Reveals That Omicron BA.1 and BA.2 Are Antigenically Distinct. Sci. Immunol. 2022, 7, eabq4450. [Google Scholar] [CrossRef] [PubMed]
- Rössler, A.; Netzl, A.; Knabl, L.; Schäfer, H.; Wilks, S.H.; Bante, D.; Falkensammer, B.; Borena, W.; von Laer, D.; Smith, D.J.; et al. BA.2 and BA.5 Omicron Differ Immunologically from Both BA.1 Omicron and Pre-Omicron Variants. Nat. Commun. 2022, 13, 7701. [Google Scholar] [CrossRef] [PubMed]
- Wilks, S.H.; Mühlemann, B.; Shen, X.; Türeli, S.; LeGresley, E.B.; Netzl, A.; Caniza, M.A.; Chacaltana-Huarcaya, J.N.; Corman, V.M.; Daniell, X.; et al. Mapping SARS-CoV-2 Antigenic Relationships and Serological Responses. bioRxiv Prepr. Serv. Biol. 2022, 2022, 1–18. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, J.; Zou, J.; Kalveram, B.; Machado, R.R.G.; Ren, P.; Türeli, S.; Smith, D.J.; Weaver, S.C.; Xie, X.; et al. Cross-Neutralization and Cross-Protection among SARS-CoV-2 Viruses Bearing Different Variant Spikes. Signal Transduct. Target. Ther. 2022, 7, 2020–2023. [Google Scholar] [CrossRef] [PubMed]
- Lusvarghi, S.; Pollett, S.D.; Neerukonda, S.N.; Wang, W.; Wang, R.; Vassell, R.; Epsi, N.J.; Fries, A.C.; Agan, B.K.; Lindholm, D.A.; et al. SARS-CoV-2 BA.1 Variant Is Neutralized by Vaccine Booster–Elicited Serum but Evades Most Convalescent Serum and Therapeutic Antibodies. Sci. Transl. Med. 2022, 14, 8543. [Google Scholar] [CrossRef]
- Wang, W.; Lusvarghi, S.; Subramanian, R.; Epsi, N.J.; Wang, R.; Goguet, E.; Fries, A.C.; Echegaray, F.; Vassell, R.; Coggins, S.A.; et al. Antigenic Cartography of Well-Characterized Human Sera Shows SARS-CoV-2 Neutralization Differences Based on Infection and Vaccination History. Cell Host. Microbe 2022, 30, 1745–1758.e7. [Google Scholar] [CrossRef] [PubMed]
- Amanat, F.; Strohmeier, S.; Meade, P.; Dambrauskas, N.; Mühlemann, B.; Smith, D.J.; Vigdorovich, V.; Noah Sather, D.; Coughlan, L.; Krammer, F. Vaccination with SARS-CoV-2 Variants of Concern Protects Mice from Challenge with Wild-Type Virus. PLoS ONE 2021, 19, e3001384. [Google Scholar] [CrossRef]
- Tuekprakhon, A.; Nutalai, R.; Dijokaite-Guraliuc, A.; Zhou, D.; Ginn, H.M.; Selvaraj, M.; Liu, C.; Mentzer, A.J.; Supasa, P.; Duyvesteyn, H.M.E.; et al. Antibody Escape of SARS-CoV-2 Omicron BA.4 and BA.5 from Vaccine and BA.1 Serum. Cell 2022, 185, 2422–2433.e13. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Yeung, J.; Kalveram, B.; Bills, C.J.; Chen, J.Y.C.; Kurhade, C.; Zou, J.; Widen, S.G.; Mann, B.R.; Kondor, R.; et al. Cross-Neutralization and Viral Fitness of SARS-CoV-2 Omicron Sublineages. Emerg. Microbes Infect. 2023, 12, e2161422. [Google Scholar] [CrossRef]
- Neerukonda, S.N.; Vassell, R.; Lusvarghi, S.; Wang, R.; Echegaray, F.; Bentley, L.; Eakin, A.E.; Erlandson, K.J.; Katzelnick, L.C.; Weiss, C.D.; et al. SARS-CoV-2 Delta Variant Displays Moderate Resistance to Neutralizing Antibodies and Spike Protein Properties of Higher Soluble ACE2 Sensitivity, Enhanced Cleavage and Fusogenic Activity. Viruses 2021, 13, 2485. [Google Scholar] [CrossRef] [PubMed]
- van der Straten, K.; Guerra, D.; van Gils, M.J.; Bontjer, I.; Caniels, T.G.; van Willigen, H.D.G.; Wynberg, E.; Poniman, M.; Burger, J.A.; Bouhuijs, J.H.; et al. Antigenic Cartography Using Sera from Sequence-Confirmed SARS-CoV-2 Variants of Concern Infections Reveals Antigenic Divergence of Omicron. Immunity 2022, 55, 1725–1731.e4. [Google Scholar] [CrossRef]
- Gushchin, V.A.; Pochtovyi, A.A.; Kustova, D.D.; Ogarkova, D.A.; Tarnovetskii, I.Y.; Belyaeva, E.D.; Divisenko, E.V.; Vasilchenko, L.A.; Shidlovskaya, E.V.; Kuznetsova, N.A.; et al. Dynamics of SARS-CoV-2 Major Genetic Lineages in Moscow in the Context of Vaccine Prophylaxis. Int. J. Mol. Sci. 2022, 23, 14670. [Google Scholar] [CrossRef]
- Horton, D.L.; McElhinney, L.M.; Marston, D.A.; Wood, J.L.N.; Russell, C.A.; Lewis, N.; Kuzmin, I.V.; Fouchier, R.A.M.; Osterhaus, A.D.M.E.; Fooks, A.R.; et al. Quantifying Antigenic Relationships among the Lyssaviruses. J. Virol. 2010, 84, 11841–11848. [Google Scholar] [CrossRef]
- Lorusso, A.; Marini, V.; Di Gennaro, A.; Ronchi, G.F.; Casaccia, C.; Carelli, G.; Passantino, G.; D’Alterio, N.; D’Innocenzo, V.; Savini, G.; et al. Antigenic Relationship among Zoonotic Flaviviruses from Italy. Infect. Genet. Evol. 2019, 68, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Katzelnick, L.C.; Fonville, J.M.; Gromowski, G.D.; Arriaga, J.B.; Green, A.; James, S.L.; Lau, L.; Montoya, M.; Wang, C.; Vanblargan, L.A.; et al. Dengue Viruses Cluster Antigenically but Not as Discrete Serotypes. Science 2015, 349, 1338–1343. [Google Scholar] [CrossRef]
- Schaefer-Babajew, D.; Wang, Z.; Muecksch, F.; Cho, A.; Loewe, M.; Cipolla, M.; Raspe, R.; Johnson, B.; Canis, M.; DaSilva, J.; et al. Antibody Feedback Regulates Immune Memory after SARS-CoV-2 mRNA Vaccination. Nature 2023, 613, 735–742. [Google Scholar] [CrossRef]
- Bekliz, M.; Adea, K.; Vetter, P.; Eberhardt, C.S.; Hosszu-Fellous, K.; Vu, D.L.; Puhach, O.; Essaidi-Laziosi, M.; Waldvogel-Abramowski, S.; Stephan, C.; et al. Neutralization Capacity of Antibodies Elicited through Homologous or Heterologous Infection or Vaccination against SARS-CoV-2 VOCs. Nat. Commun. 2022, 13, 1–10. [Google Scholar] [CrossRef]
- Fabricius, D.; Ludwig, C.; Scholz, J.; Rode, I.; Tsamadou, C.; Jacobsen, E.M.; Winkelmann, M.; Grempels, A.; Lotfi, R.; Janda, A.; et al. mRNA Vaccines Enhance Neutralizing Immunity against SARS-CoV-2 Variants in Convalescent and ChAdOx1-Primed Subjects. Vaccines 2021, 9, 918. [Google Scholar] [CrossRef]
- Hillus, D.; Schwarz, T.; Tober-Lau, P.; Vanshylla, K.; Hastor, H.; Thibeault, C.; Jentzsch, S.; Helbig, E.T.; Lippert, L.J.; Tscheak, P.; et al. Safety, Reactogenicity, and Immunogenicity of Homologous and Heterologous Prime-Boost Immunisation with ChAdOx1 NCoV-19 and BNT162b2: A Prospective Cohort Study. Lancet Respir. Med. 2021, 9, 1255–1265. [Google Scholar] [CrossRef] [PubMed]
- Godkov, M.A.; Ogarkova, D.A.; Gushchin, V.A.; Kleymenov, D.A.; Mazunina, E.P.; Bykonia, E.N.; Pochtovyi, A.A.; Shustov, V.V.; Shcheblyakov, D.V.; Komarov, A.G.; et al. Revaccination in Age-Risk Groups with Sputnik V Is Immunologically Effective and Depends on the Initial Neutralizing SARS-CoV-2 IgG Antibodies Level. Vaccines 2023, 11, 90. [Google Scholar] [CrossRef]
- Bates, T.A.; Leier, H.C.; McBride, S.K.; Schoen, D.; Lyski, Z.L.; Lee, D.X.; Messer, W.B.; Curlin, M.E.; Tafesse, F.G. An Extended Interval between Vaccination and Infection Enhances Hybrid Immunity against SARS-CoV-2 Variants. JCI Insight 2023, 8, e165265. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.J.; Rouphael, N.G.; Widge, A.T.; Jackson, L.A.; Roberts, P.C.; Makhene, M.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; Pruijssers, A.J.; et al. Safety and Immunogenicity of SARS-CoV-2 mRNA-1273 Vaccine in Older Adults. N. Engl. J. Med. 2020, 383, 2427–2438. [Google Scholar] [CrossRef] [PubMed]
- Walsh, E.E.; Frenck, R.W.; Falsey, A.R.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M.J.; Bailey, R.; et al. Safety and Immunogenicity of Two RNA-Based COVID-19 Vaccine Candidates. N. Engl. J. Med. 2020, 383, 2439–2450. [Google Scholar] [CrossRef]
- Logunov, D.Y.; Dolzhikova, I.V.; Zubkova, O.V.; Tukhvatullin, A.I.; Shcheblyakov, D.V.; Dzharullaeva, A.S.; Grousova, D.M.; Erokhova, A.S.; Kovyrshina, A.V.; Botikov, A.G.; et al. Safety and Immunogenicity of an rAd26 and rAd5 Vector-Based Heterologous Prime-Boost COVID-19 Vaccine in Two Formulations: Two Open, Non-Randomised Phase 1/2 Studies from Russia. Lancet 2020, 396, 887–897. [Google Scholar] [CrossRef]
- Byazrova, M.G.; Kulemzin, S.V.; Astakhova, E.A.; Belovezhets, T.N.; Efimov, G.A.; Chikaev, A.N.; Kolotygin, I.O.; Gorchakov, A.A.; Taranin, A.V.; Filatov, A.V. Memory B Cells Induced by Sputnik V Vaccination Produce SARS-CoV-2 Neutralizing Antibodies Upon Ex Vivo Restimulation. Front. Immunol. 2022, 13, 840707. [Google Scholar] [CrossRef]
- Astakhova, E.A.; Byazrova, M.G.; Yusubalieva, G.M.; Kulemzin, S.V.; Kruglova, N.A.; Prilipov, A.G.; Baklaushev, V.P.; Gorchakov, A.A.; Taranin, A.V.; Filatov, A.V. Functional Profiling of In Vitro Reactivated Memory B Cells Following Natural SARS-CoV-2 Infection and Gam-COVID-Vac Vaccination. Cells 2022, 11, 1991. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Name of Subgroup | Uninfected Donors, Sputnik V Booster | Uninfected Donors, Comirnaty Booster | Convalescent Donors, Sputnik V Booster | |
|---|---|---|---|---|
| Number of participants | 39 | 12 | 7 | |
| Age | Years, median (range) | 27 (18–73) | 25 (19–43) | 23 (19–63) |
| Sex | Female | 19 | 6 | 3 |
| Male | 20 | 6 | 4 | |
| Booster vaccine | Sputnik V | Comirnaty | Sputnik V | |
| Time between prime vaccination and pre-boost serum sampling | Days, median (range) | 195 (176–270) | 275 (261–335) | 207 (183–282) |
| Time between pre- and post-boost serum sampling | Days, median (range) | 33 (29–39) | 38 (30–46) | 31 (30–36) |
| Anti-N IgG antibodies in pre-boost sera | Number of participants with a positive result | 0/39 | 0/12 | 5/7 |
| Anti-S IgG antibodies in pre-boost sera | Relative units (minimum—0, maximum—15.0) | 6.0 (0–10.4) | 7.3 (0–11.8) | 12.7 (11.2–14.4) |
| PCR-confirmed COVID-19 | Number of participants with a positive result | 0/39 | 0/12 | 7/7 |
| Time between recovery from COVID-19 and boost | Days, median (range) | - | - | 78 (42–102) |
| Infection period | - | - | May–August 2021 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Astakhova, E.A.; Morozov, A.A.; Byazrova, M.G.; Sukhova, M.M.; Mikhailov, A.A.; Minnegalieva, A.R.; Gorchakov, A.A.; Filatov, A.V. Antigenic Cartography Indicates That the Omicron BA.1 and BA.4/BA.5 Variants Remain Antigenically Distant to Ancestral SARS-CoV-2 after Sputnik V Vaccination Followed by Homologous (Sputnik V) or Heterologous (Comirnaty) Revaccination. Int. J. Mol. Sci. 2023, 24, 10493. https://doi.org/10.3390/ijms241310493
Astakhova EA, Morozov AA, Byazrova MG, Sukhova MM, Mikhailov AA, Minnegalieva AR, Gorchakov AA, Filatov AV. Antigenic Cartography Indicates That the Omicron BA.1 and BA.4/BA.5 Variants Remain Antigenically Distant to Ancestral SARS-CoV-2 after Sputnik V Vaccination Followed by Homologous (Sputnik V) or Heterologous (Comirnaty) Revaccination. International Journal of Molecular Sciences. 2023; 24(13):10493. https://doi.org/10.3390/ijms241310493
Chicago/Turabian StyleAstakhova, Ekaterina A., Alexey A. Morozov, Maria G. Byazrova, Maria M. Sukhova, Artem A. Mikhailov, Aygul R. Minnegalieva, Andrey A. Gorchakov, and Alexander V. Filatov. 2023. "Antigenic Cartography Indicates That the Omicron BA.1 and BA.4/BA.5 Variants Remain Antigenically Distant to Ancestral SARS-CoV-2 after Sputnik V Vaccination Followed by Homologous (Sputnik V) or Heterologous (Comirnaty) Revaccination" International Journal of Molecular Sciences 24, no. 13: 10493. https://doi.org/10.3390/ijms241310493
APA StyleAstakhova, E. A., Morozov, A. A., Byazrova, M. G., Sukhova, M. M., Mikhailov, A. A., Minnegalieva, A. R., Gorchakov, A. A., & Filatov, A. V. (2023). Antigenic Cartography Indicates That the Omicron BA.1 and BA.4/BA.5 Variants Remain Antigenically Distant to Ancestral SARS-CoV-2 after Sputnik V Vaccination Followed by Homologous (Sputnik V) or Heterologous (Comirnaty) Revaccination. International Journal of Molecular Sciences, 24(13), 10493. https://doi.org/10.3390/ijms241310493