
Metabolic Mechanisms and Potential Therapeutic Targets for Prevention of Ovarian Aging: Data from Up-to-Date Experimental Studies





Abstract
1. Introduction
2. Metabolic and Molecular Mechanisms of Ovarian Aging
3. General Factors Involved in Ovarian Aging
3.1. Genetic Factors Involved in Ovarian Aging
3.2. DNA Damage
3.3. Cohesins
3.4. Telomere Length
3.5. Oxidative Damage
4. Potential Molecular/Metabolic Treatments for Prevention of Ovarian Aging
4.1. Pharmaceuticals
4.1.1. Metformin
4.1.2. Visfatin
4.1.3. Sphingosine 1-Phosphate
4.2. Stem Cells
4.2.1. Human Amniotic Mesenchymal Stem Cells (hAMSC)
4.2.2. Umbilical Cord Mesenchymal Stem Cells (UC-MSC)
4.2.3. Human Placenta-Derived Mesenchymal Stem Cells (hPD-MSC)
4.2.4. Menstrual Blood-Derived Mesenchymal Stem Cells
4.2.5. Human Endothelial Progenitor Cells
4.2.6. Adipose-Derived Stem Cell Conditioned Medium (ASC-CM)
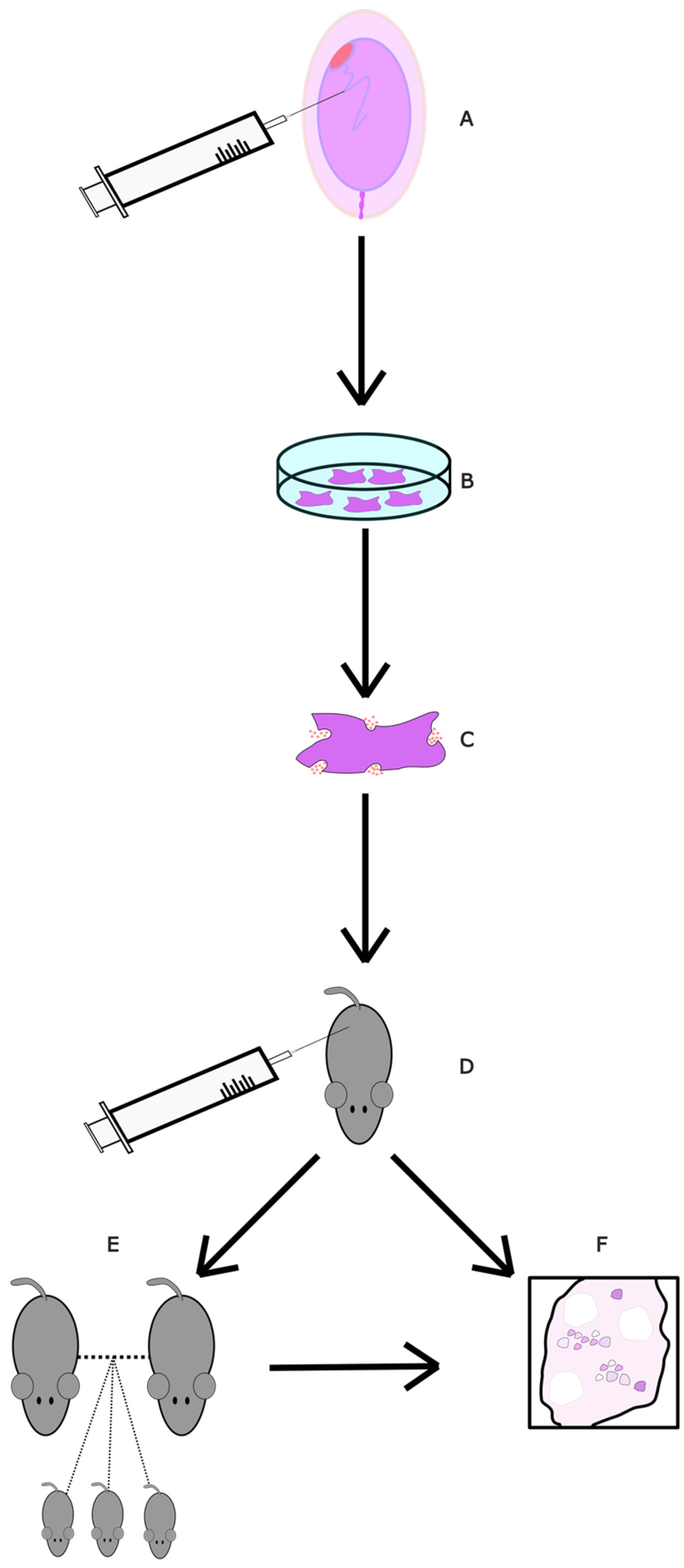
4.2.7. Autologous Mitochondrial Transfer
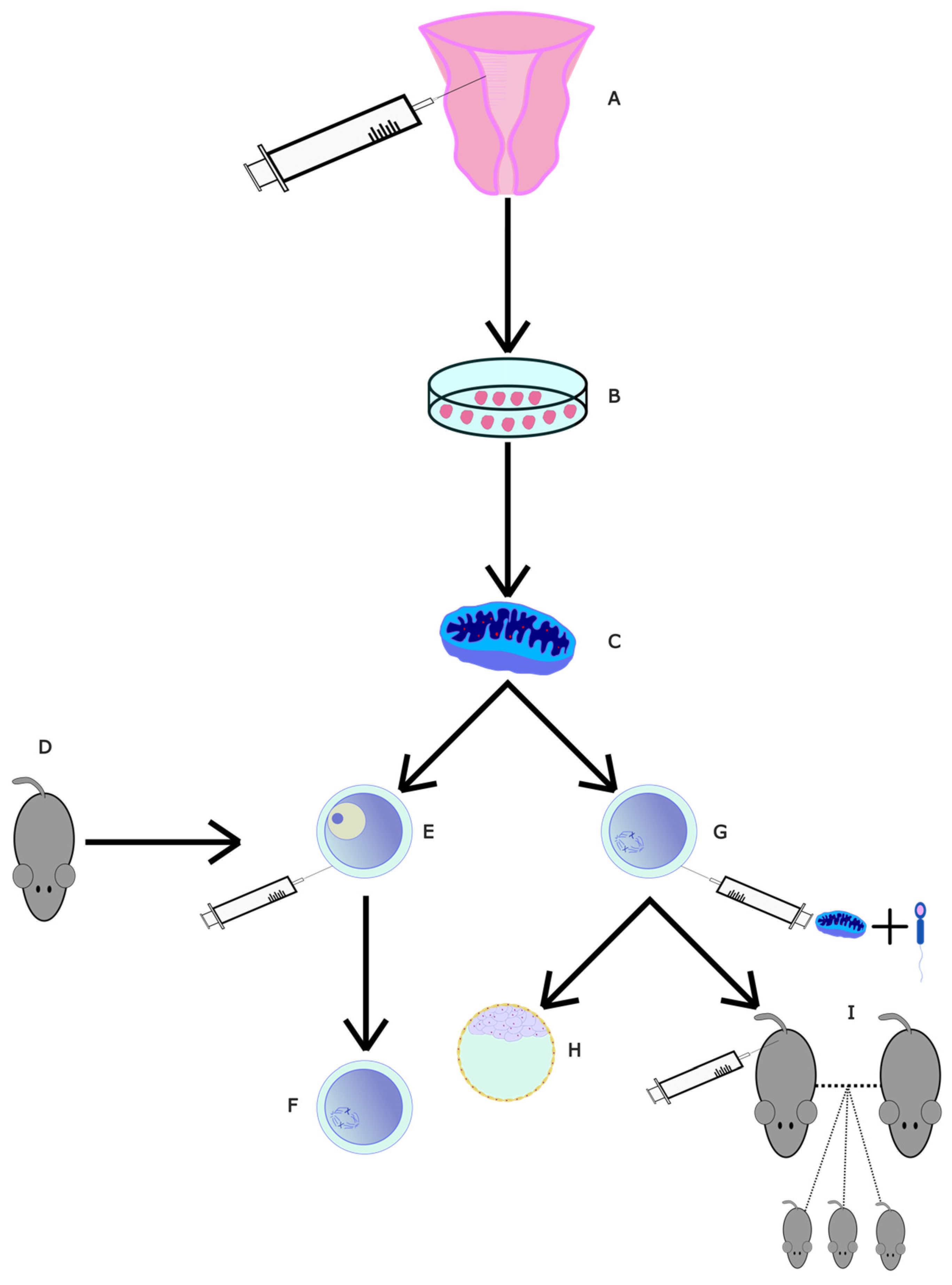
4.2.8. Mitochondrial Transfer from Endothelial Mesenchymal Stem Cells
4.3. Caloric Restriction and Caloric Restriction Mimetics
4.3.1. Caloric Restriction Diets
4.3.2. CR and Rapamycin
4.4. Other Treatments
4.4.1. DHEA
4.4.2. Hyperbaric Oxygen Treatment
4.4.3. Macrophage-Derived Extracellular Vesicles
4.4.4. Cell-Free Fat Extract
4.4.5. TrkB Agonists
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. International Classification of Diseases, GA31 Female Infertility. ICD-11 for Mortality and Morbidity Statistics. Available online: https://icd.who.int/browse11/l-m/en#/http://id.who.int/icd/entity/1237004558 (accessed on 31 March 2022).
- Female Infertility. Cleveland Clinic. Available online: https://my.clevelandclinic.org/health/diseases/17774-female-infertility (accessed on 2 April 2022).
- Mascarenhas, M.N.; Flaxman, S.R.; Boerma, T.; Vanderpoel, S.; Stevens, G.A. National, regional, and global trends in infertility prevalence since 1990: A systematic analysis of 277 health surveys. PLoS Med. 2012, 9, e1001356. [Google Scholar] [CrossRef] [PubMed]
- Bellieni, C. The Best Age for Pregnancy and Undue Pressures. J. Fam. Reprod. Health 2016, 10, 104–107. [Google Scholar]
- Martin, J.A.; Hamilton, B.E.; Osterman, M.J.K. Births in the United States, 2016. NCHS Data Brief. 2017, 287, 1–8. [Google Scholar]
- Eurostat. Women in the EU Are Having Their First Child Later. Available online: https://ec.europa.eu/eurostat/web/products-eurostat-news/-/ddn-20210224-1#:~:text=In%202019%2C%20the%20mother’s%20age,and%20Romania%20(26.9%20years) (accessed on 1 April 2022).
- Tesarik, J.; Galán-Lázaro, M.; Mendoza-Tesarik, R. Ovarian Aging: Molecular Mechanisms and Medical Management. Int. J. Mol. Sci. 2021, 22, 1371. [Google Scholar] [CrossRef]
- Park, S.U.; Walsh, L.; Berkowitz, K.M. Mechanisms of ovarian aging. Reproduction 2021, 162, R19–R33. [Google Scholar] [CrossRef]
- Smits, M.A.J.; Janssens, G.E.; Goddijn, M.; Hamer, G.; Houtkooper, R.H.; Mastenbroek, S. Longevity pathways are associated with human ovarian ageing. Hum. Reprod. Open 2021, 2021, hoab020. [Google Scholar] [CrossRef]
- Grabowska, W.; Sikora, E.; Bielak-Zmijewska, A. Sirtuins, a promising target in slowing down the ageing process. Biogerontology 2017, 18, 447–476. [Google Scholar] [CrossRef]
- Kinser, H.E.; Pincus, Z. MicroRNAs as modulators of longevity and the aging process. Hum. Genet. 2020, 139, 291–308. [Google Scholar] [CrossRef]
- Xu, S.; Schaack, S.; Seyfert, A.; Choi, E.; Lynch, M.; Cristescu, M.E. High mutation rates in the mitochondrial genomes of Daphnia pulex. Mol. Biol. Evol. 2012, 29, 763–769. [Google Scholar] [CrossRef]
- Vera, E.; de Jesus, B.B.; Foronda, M.; Flores, J.M.; Blasco, M.A. The Rate of Increase of Short Telomeres Predicts Longevity in Mammals. Cell Rep. 2012, 2, 732–737. [Google Scholar] [CrossRef]
- Venkatesan, S.; Khaw, A.K.; Hande, M.P. Telomere Biology—Insights into an Intriguing Phenomenon. Cells 2017, 6, 15. [Google Scholar] [CrossRef] [PubMed]
- Dorronsoro, A.; Santiago, F.E.; Grassi, D.; Zhang, T.; Lai, R.C.; McGowan, S.J.; Angelini, L.; Lavasani, M.; Corbo, L.; Lu, A.; et al. Mesenchymal stem cell-derived extracellular vesicles reduce senescence and extend health span in mouse models of aging. Aging Cell 2021, 20, e13337. [Google Scholar] [CrossRef]
- Takahashi, A.; Yousif, A.; Hong, L.; Chefetz, I. Premature ovarian insufficiency: Pathogenesis and therapeutic potential of mesenchymal stem cell. J. Mol. Med. 2021, 99, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Zou, Q.; Wang, F.; Wu, H.; Chen, R.; Lv, J.; Ling, M.; Sun, J.; Wang, W.; Li, H.; et al. Human amniotic mesenchymal stem cells improve ovarian function in natural aging through secreting hepatocyte growth factor and epidermal growth factor. Stem Cell Res. Ther. 2018, 9, 55. [Google Scholar] [CrossRef]
- Salha, O.; Abusheika, N.; Sharma, V. Dynamics of human follicular growth and in-vitro oocyte maturation. Hum. Reprod. Updat. 1998, 4, 816–832. [Google Scholar] [CrossRef]
- Baker, T.G. A quantitative and cytological study of germ cells in human ovaries. Proc. R. Soc. London Ser. B Boil. Sci. 1963, 158, 417–433. [Google Scholar] [CrossRef]
- Richardson, S.J.; Senikas, V.; Nelson, J.F. Follicular depletion during the menopausal transition: Evidence for accelerated loss and ultimate exhaustion. J. Clin. Endocrinol. Metab. 1987, 65, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
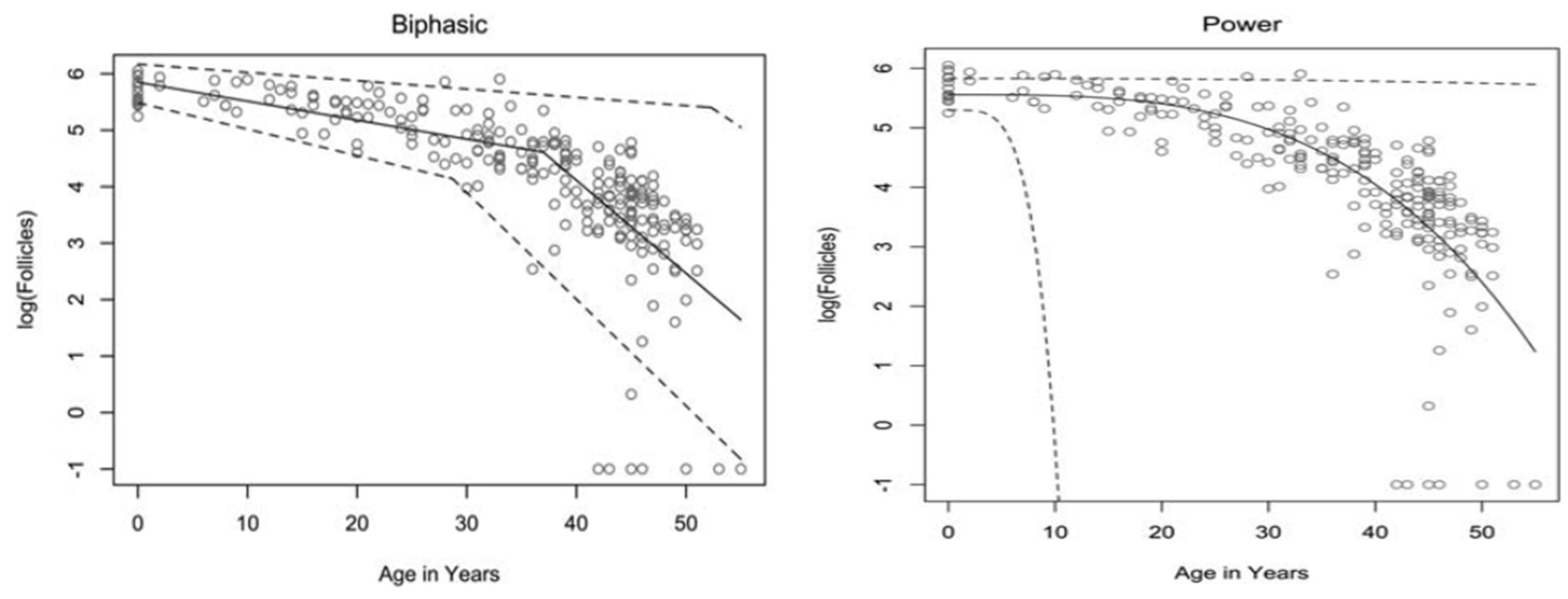
- Hansen, K.R.; Knowlton, N.S.; Thyer, A.C.; Charleston, J.S.; Soules, M.R.; Klein, N.A. A new model of reproductive aging: The decline in ovarian non-growing follicle number from birth to menopause. Hum. Reprod. 2008, 23, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Faddy, M.J.; Gosden, R.G.; Gougeon, A.; Richardson, S.J.; Nelson, J.F. Accelerated disappearance of ovarian follicles in mid-life: Implications for forecasting menopause. Hum. Reprod. 1992, 7, 1342–1346. [Google Scholar] [CrossRef]
- Cox, E.; Takov, V. Embryology; Ovarian Follicle Development; StatPearls: Tampa, FL, USA, 2022. Available online: https://www.ncbi.nlm.nih.gov/books/NBK532300/ (accessed on 22 May 2022).
- Lebbe, M.; Woodruff, T.K. Involvement of androgens in ovarian health and disease. Mol. Hum. Reprod. 2013, 19, 828–837. [Google Scholar] [CrossRef]
- Franks, S.; Hardy, K. Androgen Action in the Ovary. Front. Endocrinol. 2018, 9, 452. [Google Scholar] [CrossRef]
- Drummond, A.E. The role of steroids in follicular growth. Reprod. Biol. Endocrinol. 2006, 4, 16. [Google Scholar] [CrossRef] [PubMed]
- Orr, B.; Godek, K.M.; Compton, D. Aneuploidy. Curr. Biol. 2015, 25, R538–R542. [Google Scholar] [CrossRef] [PubMed]
- Coxworth, J.E.; Hawkes, K. Ovarian follicle loss in humans and mice: Lessons from statistical model comparison. Hum. Reprod. 2010, 25, 1796–1805. [Google Scholar] [CrossRef]
- Holton, R.A.; Harris, A.M.; Mukerji, B.; Singh, T.; Dia, F.; Berkowitz, K.M. CHTF18 ensures the quantity and quality of the ovarian reserve. Biol. Reprod. 2020, 103, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Turan, V.; Oktay, K. BRCA-related ATM-mediated DNA double-strand break repair and ovarian aging. Hum. Reprod. Updat. 2020, 26, 43–57. [Google Scholar] [CrossRef]
- Lin, W.; Titus, S.; Moy, F.; Ginsburg, E.S.; Oktay, K. Ovarian Aging in Women with BRCA Germline Mutations. J. Clin. Endocrinol. Metab. 2017, 102, 3839–3847. [Google Scholar] [CrossRef]
- Laven, J.S.E.; Visser, J.A.; Uitterlinden, A.G.; Vermeij, W.P.; Hoeijmakers, J.H.J. Menopause: Genome stability as new paradigm. Maturitas 2016, 92, 15–23. [Google Scholar] [CrossRef]
- Titus, S.; Li, F.; Stobezki, R.; Akula, K.; Unsal, E.; Jeong, K.; Dickler, M.; Robson, M.; Moy, F.; Goswami, S.; et al. Impairment of BRCA1-related DNA double-strand break repair leads to ovarian aging in mice and humans. Sci. Transl. Med. 2013, 5, 172ra21. [Google Scholar] [CrossRef]
- Tiwari, V.; Wilson, D.M., 3rd. DNA Damage and Associated DNA Repair Defects in Disease and Premature Aging. Am. J. Hum. Genet. 2019, 105, 237–257. [Google Scholar] [CrossRef]
- Nagaoka, S.I.; Hassold, T.J.; Hunt, P.A. Human aneuploidy: Mechanisms and new insights into an age-old problem. Nat. Rev. Genet. 2012, 13, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Hodges, C.A.; Revenkova, E.; Jessberger, R.; Hassold, T.J.; A Hunt, P. SMC1β-deficient female mice provide evidence that cohesins are a missing link in age-related nondisjunction. Nat. Genet. 2005, 37, 1351–1355. [Google Scholar] [CrossRef] [PubMed]
- Revenkova, E.; Herrmann, K.; Adelfalk, C.; Jessberger, R. Oocyte Cohesin Expression Restricted to Predictyate Stages Provides Full Fertility and Prevents Aneuploidy. Curr. Biol. 2010, 20, 1529–1533. [Google Scholar] [CrossRef]
- Cheng, J.-M.; Liu, Y.-X. Age-Related Loss of Cohesion: Causes and Effects. Int. J. Mol. Sci. 2017, 18, 1578. [Google Scholar] [CrossRef]
- Herbert, M.; Kalleas, D.; Cooney, D.; Lamb, M.; Lister, L. Meiosis and Maternal Aging: Insights from Aneuploid Oocytes and Trisomy Births. Cold Spring Harb. Perspect. Biol. 2015, 7, a017970. [Google Scholar] [CrossRef]
- Chiang, T.; Duncan, F.E.; Schindler, K.; Schultz, R.M.; Lampson, M.A. Evidence that Weakened Centromere Cohesion Is a Leading Cause of Age-Related Aneuploidy in Oocytes. Curr. Biol. 2010, 20, 1522–1528. [Google Scholar] [CrossRef]
- Ciosk, R.; Shirayama, M.; Shevchenko, A.; Tanaka, T.; Toth, A.; Shevchenko, A.; Nasmyth, K. Cohesin’s Binding to Chromosomes Depends on a Separate Complex Consisting of Scc2 and Scc4 Proteins. Mol. Cell 2000, 5, 243–254. [Google Scholar] [CrossRef]
- Xu, X.; Chen, X.; Zhang, X.; Liu, Y.; Wang, Z.; Wang, P.; Du, Y.; Qin, Y.; Chen, Z.-J. Impaired telomere length and telomerase activity in peripheral blood leukocytes and granulosa cells in patients with biochemical primary ovarian insufficiency. Hum. Reprod. 2017, 32, 201–207. [Google Scholar] [CrossRef]
- Miranda-Furtado, C.L.; Luchiari, H.R.; Chielli Pedroso, D.C.; Kogure, G.S.; Caetano, L.C.; Santana, B.A.; Santana, V.P.; Benetti-Pinto, C.L.; Reis, F.M.; Maciel, M.A.; et al. Skewed X-chromosome inactivation and shorter telomeres associate with idiopathic premature ovarian insufficiency. Fertil. Steril. 2018, 110, 476–485.e1. [Google Scholar] [CrossRef]
- Butts, S.; Riethman, H.; Ratcliffe, S.; Shaunik, A.; Coutifaris, C.; Barnhart, K. Correlation of telomere length and telomerase activity with occult ovarian insufficiency. J. Clin. Endocrinol. Metab. 2009, 94, 4835–4843. [Google Scholar] [CrossRef]
- Wang, S.; Zheng, Y.; Li, J.; Yu, Y.; Zhang, W.; Song, M.; Liu, Z.; Min, Z.; Hu, H.; Jing, Y.; et al. Single-Cell Transcriptomic Atlas of Primate Ovarian Aging. Cell 2020, 180, 585–600.e19. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Trimarchi, J.R.; Smith, P.J.S.; Keefe, D.L. Mitochondrial dysfunction leads to telomere attrition and genomic instability. Aging Cell 2002, 1, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, H.; Hamatani, T.; Kamijo, S.; Iwai, M.; Kobanawa, M.; Ogawa, S.; Miyado, K.; Tanaka, M. Impact of Oxidative Stress on Age-Associated Decline in Oocyte Developmental Competence. Front. Endocrinol. 2019, 10, 811. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Keilty, D.; Zhang, Z.; Chian, R. Mitochondria in oocyte aging: Current understanding. Facts Views Vis. ObGyn 2017, 9, 29–38. [Google Scholar] [PubMed]
- Sallevelt, S.C.E.H.; Dreesen, J.C.F.M.; Drüsedau, M.; Spierts, S.; Coonen, E.; van Tienen, F.H.J.; van Golde, R.J.T.; de Coo, I.F.M.; Geraedts, J.P.M.; Die-Smulders, C.E.M.D.; et al. Preimplantation genetic diagnosis in mitochondrial DNA disorders: Challenge and success. J. Med. Genet. 2013, 50, 125–132. [Google Scholar] [CrossRef]
- Qin, X.; Du, D.; Chen, Q.; Wu, M.; Wu, T.; Wen, J.; Jin, Y.; Zhang, J.; Wang, S. Metformin prevents murine ovarian aging. Aging 2019, 11, 3785–3794. [Google Scholar] [CrossRef]
- Park, B.-K.; Park, M.J.; Kim, H.G.; Han, S.E.; Kim, C.-W.; Joo, B.S.; Lee, K.-S. Role of Visfatin in Restoration of Ovarian Aging and Fertility in the Mouse Aged 18 Months. Reprod. Sci. 2020, 27, 681–689. [Google Scholar] [CrossRef]
- Guzel, Y.; Bildik, G.; Oktem, O. Sphingosine-1-phosphate protects human ovarian follicles from apoptosis in vitro. Eur. J. Obstet. Gynecol. Reprod. Biol. 2018, 222, 19–24. [Google Scholar] [CrossRef]
- Mumusoglu, S.; Turan, V.; Uckan, H.; Suzer, A.; Sokmensuer, L.K.; Bozdag, G. The Impact of a Long-Acting Oral Sphingosine-1-Phosphate Analogue on Ovarian Aging in a Rat Model. Reprod. Sci. 2018, 25, 1330–1335. [Google Scholar] [CrossRef]
- Mohammed, I.; Hollenberg, M.D.; Ding, H.; Triggle, C.R. A Critical Review of the Evidence That Metformin Is a Putative Anti-Aging Drug That Enhances Healthspan and Extends Lifespan. Front. Endocrinol. 2021, 12, 718942. [Google Scholar] [CrossRef]
- Muss, H.B.; Smitherman, A.; Wood, W.A.; Nyrop, K.; Tuchman, S.; Randhawa, P.K.; Entwistle, A.R.; Mitin, N.; Shachar, S.S. p16 a biomarker of aging and tolerance for cancer therapy. Transl. Cancer Res. 2020, 9, 5732–5742. [Google Scholar] [CrossRef] [PubMed]
- Sethi, J.K. Is PBEF/visfatin/Nampt an authentic adipokine relevant to the metabolic syndrome? Curr. Hypertens. Rep. 2007, 9, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.N.; Chang, E.M. Primordial follicle activation as new treatment for primary ovarian insufficiency. Clin. Exp. Reprod. Med. 2019, 46, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Adya, R.; Tan, B.K.; Punn, A.; Chen, J.; Randeva, H.S. Visfatin induces human endothelial VEGF and MMP-2/9 production via MAPK and PI3K/Akt signalling pathways: Novel insights into visfatin-induced angiogenesis. Cardiovasc. Res. 2008, 78, 356–365. [Google Scholar] [CrossRef]
- Park, J.-W.; Kim, W.-H.; Shin, S.-H.; Kim, J.Y.; Yun, M.R.; Park, K.J.; Park, H.-Y. Visfatin exerts angiogenic effects on human umbilical vein endothelial cells through the mTOR signaling pathway. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2011, 1813, 763–771. [Google Scholar] [CrossRef]
- Cuvillier, O.; Pirianov, G.; Kleuser, B.; Vanek, P.G.; Coso, O.A.; Gutkind, J.S.; Spiegel, S. Suppression of ceramide-mediated programmed cell death by sphingosine-1-phosphate. Nature 1996, 381, 800–803. [Google Scholar] [CrossRef]
- Yang, W.; Zhang, J.; Xu, B.; He, Y.; Liu, W.; Li, J.; Zhang, S.; Lin, X.; Su, D.; Wu, T.; et al. HucMSC-Derived Exosomes Mitigate the Age-Related Retardation of Fertility in Female Mice. Mol. Ther. 2020, 28, 1200–1213. [Google Scholar] [CrossRef]
- Jiao, W.; Mi, X.; Yang, Y.; Liu, R.; Liu, Q.; Yan, T.; Chen, Z.-J.; Qin, Y.; Zhao, S. Mesenchymal stem cells combined with autocrosslinked hyaluronic acid improve mouse ovarian function by activating the PI3K-AKT pathway in a paracrine manner. Stem Cell Res. Ther. 2022, 13, 49. [Google Scholar] [CrossRef]
- Kim, K.-H.; Kim, E.-Y.; Kim, G.J.; Ko, J.-J.; Cha, K.Y.; Koong, M.K.; Lee, K.-A. Human placenta-derived mesenchymal stem cells stimulate ovarian function via miR-145 and bone morphogenetic protein signaling in aged rats. Stem Cell Res. Ther. 2020, 11, 472. [Google Scholar] [CrossRef]
- Kim, G.A.; Lee, Y.; Kim, H.J.; Oh, H.J.; Kang, S.K.; Ra, J.C.; Lee, B.C. Intravenous human endothelial progenitor cell administration into aged mice enhances embryo development and oocyte quality by reducing inflammation, endoplasmic reticulum stress and apoptosis. J. Vet. Med. Sci. 2018, 80, 1905–1913. [Google Scholar] [CrossRef]
- Ra, K.; Oh, H.J.; Kim, G.A.; Kang, S.K.; Ra, J.C.; Lee, B.C. High Frequency of Intravenous Injection of Human Adipose Stem Cell Conditioned Medium Improved Embryo Development of Mice in Advanced Maternal Age through Antioxidant Effects. Animals 2020, 10, 978. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Mei, Q.; Xie, Q.; Li, H.; Su, P.; Zhang, L.; Li, K.; Ma, D.; Chen, G.; Li, J.; et al. A comparative study of Mesenchymal Stem Cells transplantation approach to antagonize age-associated ovarian hypofunction with consideration of safety and efficiency. J. Adv. Res. 2022, 38, 245–259. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Hao, J.; Liu, B.; Ouyang, Y.; Guo, J.; Dong, M.; Wang, Z.; Gao, F.; Yao, Y. Supplementation of mitochondria from endometrial mesenchymal stem cells improves oocyte quality in aged mice. Cell Prolif. 2023, 56, e13372. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, M.; Zamah, A.; Conti, M. Epidermal Growth Factor-Like Growth Factors in the Follicular Fluid: Role in Oocyte Development and Maturation. Semin. Reprod. Med. 2009, 27, 052–061. [Google Scholar] [CrossRef] [PubMed]
- Uzumcu, M.; Pan, Z.; Chu, Y.; Kuhn, P.E.; Zachow, R. Immunolocalization of the hepatocyte growth factor (HGF) system in the rat ovary and the anti-apoptotic effect of HGF in rat ovarian granulosa cells in vitro. Reproduction 2006, 132, 291–299. [Google Scholar] [CrossRef]
- Taniguchi, F.; Harada, T.; Deura, I.; Iwabe, T.; Tsukihara, S.; Terakawa, N. Hepatocyte growth factor promotes cell proliferation and inhibits progesterone secretion via PKA and MAPK pathways in a human granulosa cell line. Mol. Reprod. Dev. 2004, 68, 335–344. [Google Scholar] [CrossRef]
- Li, T.; Xia, M.; Gao, Y.; Chen, Y.; Xu, Y. Human umbilical cord mesenchymal stem cells: An overview of their potential in cell-based therapy. Expert Opin. Biol. Ther. 2015, 15, 1293–1306. [Google Scholar] [CrossRef]
- Wang, L.; Yu, C.; Chang, T.; Zhang, M.; Song, S.; Xiong, C.; Su, P.; Xiang, W. In situ repair abilities of human umbilical cord–derived mesenchymal stem cells and autocrosslinked hyaluronic acid gel complex in rhesus monkeys with intrauterine adhesion. Sci. Adv. 2020, 6, eaba6357. [Google Scholar] [CrossRef]
- Lai, R.C.; Yeo, R.W.Y.; Lim, S.K. Mesenchymal stem cell exosomes. Semin. Cell Dev. Biol. 2015, 40, 82–88. [Google Scholar] [CrossRef]
- Kim, J.Y.; Park, S.; Park, S.H.; Lee, D.; Kim, G.H.; Noh, J.E.; Lee, K.J.; Kim, G.J. Overexpression of pigment epithelium-derived factor in placenta-derived mesenchymal stem cells promotes mitochondrial biogenesis in retinal cells. Lab. Investig. 2021, 101, 51–69. [Google Scholar] [CrossRef]
- Medina, R.J.; Barber, C.L.; Sabatier, F.; Dignat-George, F.; Melero-Martin, J.M.; Khosrotehrani, K.; Ohneda, O.; Randi, A.M.; Chan, J.K.Y.; Yamaguchi, T.; et al. Endothelial Progenitors: A Consensus Statement on Nomenclature. Stem Cells Transl. Med. 2017, 6, 1316–1320. [Google Scholar] [CrossRef]
- Patel, J.; Donovan, P.; Khosrotehrani, K. Concise Review: Functional Definition of Endothelial Progenitor Cells: A Molecular Perspective. Stem Cells Transl. Med. 2016, 5, 1302–1306. [Google Scholar] [CrossRef] [PubMed]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human Adipose Tissue Is a Source of Multipotent Stem Cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef]
- Kim, W.-S.; Park, B.-S.; Sung, J.-H. The wound-healing and antioxidant effects of adipose-derived stem cells. Expert Opin. Biol. Ther. 2009, 9, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Pawitan, J.A. Prospect of Stem Cell Conditioned Medium in Regenerative Medicine. BioMed Res. Int. 2014, 2014, 965849. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-B.; Hao, J.-X.; Meng, T.-G.; Guo, L.; Dong, M.-Z.; Fan, L.-H.; Ouyang, Y.-C.; Wang, G.; Sun, Q.-Y.; Ou, X.-H.; et al. Transfer of autologous mitochondria from adipose tissue-derived stem cells rescues oocyte quality and infertility in aged mice. Aging 2017, 9, 2480–2488. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.-L.; Chen, X.-C.; Fu, Y.-C.; Xu, J.-J.; Li, L.; Lin, X.-H.; Xiang, Y.-F.; Zhang, X.-M. The effects of caloric restriction and a high-fat diet on ovarian lifespan and the expression of SIRT1 and SIRT6 proteins in rats. Aging Clin. Exp. Res. 2012, 24, 125–133. [Google Scholar] [CrossRef]
- Isola, J.V.V.; Zanini, B.M.; Hense, J.D.; Alvarado-Rincón, J.A.; Garcia, D.N.; Pereira, G.C.; Vieira, A.D.; Oliveira, T.L.; Collares, T.; Gasperin, B.G.; et al. Mild calorie restriction, but not 17α-estradiol, extends ovarian reserve and fertility in female mice. Exp. Gerontol. 2022, 159, 111669. [Google Scholar] [CrossRef]
- Garcia, D.N.; Saccon, T.D.; Pradiee, J.; Rincón, J.A.A.; Andrade, K.R.S.; Rovani, M.T.; Mondadori, R.G.; Cruz, L.A.X.; Barros, C.C.; Masternak, M.M.; et al. Effect of caloric restriction and rapamycin on ovarian aging in mice. Geroscience 2019, 41, 395–408. [Google Scholar] [CrossRef]
- Flanagan, E.W.; Most, J.; Mey, J.T.; Redman, L.M. Calorie Restriction and Aging in Humans. Annu. Rev. Nutr. 2020, 40, 105–133. [Google Scholar] [CrossRef] [PubMed]
- Llarena, N.; Hine, C. Reproductive Longevity and Aging: Geroscience Approaches to Maintain Long-Term Ovarian Fitness. J. Gerontol. Ser. A 2021, 76, 1551–1560. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, Q.; Du, D.; Wu, T.; Wen, J.; Wu, M.; Zhang, Y.; Yan, W.; Zhou, S.; Li, Y.; et al. Can ovarian aging be delayed by pharmacological strategies? Aging 2019, 11, 817–832. [Google Scholar] [CrossRef]
- Li, J.; Kim, S.G.; Blenis, J. Rapamycin: One drug, many effects. Cell Metab. 2014, 19, 373–379. [Google Scholar] [CrossRef]
- Pelosi, E.; Omari, S.; Michel, M.; Ding, J.; Amano, T.; Forabosco, A.; Schlessinger, D.; Ottolenghi, C. Constitutively active Foxo3 in oocytes preserves ovarian reserve in mice. Nat. Commun. 2013, 4, 1843. [Google Scholar] [CrossRef]
- John, G.B.; Shirley, L.J.; Gallardo, T.D.; Castrillon, D.H. Specificity of the requirement for Foxo3 in primordial follicle activation. Reproduction 2007, 133, 855–863. [Google Scholar] [CrossRef]
- Chu, N.; Gui, Y.; Qiu, X.; Zhang, N.; Li, L.; Li, D.; Tang, W.; Gober, H.-J.; Zhang, B.; Wang, L. The effect of DHEA on apoptosis and cohesin levels in oocytes in aged mice. Biosci. Trends 2017, 11, 427–438. [Google Scholar] [CrossRef]
- Ma, Y.; Zhong, Y.; Chen, X.; Liu, H.; Shi, Y.; Zhang, X.; Sun, H. Hyperbaric Oxygen Treatment Ameliorates the Decline in Oocyte Quality and Improves the Fertility of Aged Female Mice. Reprod. Sci. 2022, 30, 1834–1840. [Google Scholar] [CrossRef]
- Xiao, Y.; Peng, X.; Peng, Y.; Zhang, C.; Liu, W.; Yang, W.; Dou, X.; Jiang, Y.; Wang, Y.; Yang, S.; et al. Macrophage-derived extracellular vesicles regulate follicular activation and improve ovarian function in old mice by modulating local environment. Clin. Transl. Med. 2022, 12, e1071. [Google Scholar] [CrossRef]
- Liu, M.; Li, W.; Zhou, X.; Zhou, M.; Zhang, W.; Liu, Q.; Zhang, A.; Xu, B. Cell-Free Fat Extract Improves Ovarian Function and Fertility in Mice with Advanced Age. Front. Endocrinol. 2022, 13, 912648. [Google Scholar] [CrossRef]
- Qin, X.; Zhao, Y.; Zhang, T.; Yin, C.; Qiao, J.; Guo, W.; Lu, B. TrkB agonist antibody ameliorates fertility deficits in aged and cyclophosphamide-induced premature ovarian failure model mice. Nat. Commun. 2022, 13, 914. [Google Scholar] [CrossRef]
- Rutkowski, K.; Sowa, P.; Rutkowska-Talipska, J.; Kuryliszyn-Moskal, A.; Rutkowski, R. Dehydroepiandrosterone (DHEA): Hypes and Hopes. Drugs 2014, 74, 1195–1207. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Kang, B.; Cai, Y.; Chen, C.; Yu, Z.; Li, W.; Zhang, W. Cell-free fat extract attenuates osteoarthritis via chondrocytes regeneration and macrophages immunomodulation. Stem Cell Res. Ther. 2022, 13, 133. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.; Wang, X.; Yu, Z.; Cai, Y.; Liu, W.; Zhou, G.; Wang, X.; Yu, Z.; Zhang, W.J. Cell-free fat extract promotes tissue regeneration in a tissue expansion model. Stem Cell Res. Ther. 2020, 11, 50. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xu, M.; Kan, T.; Wang, Y.; Zhang, S.; Deng, M.; Zhang, W.; Li, H.; Yu, Z. Fat Extract Modulates Calcium Signaling and Protects against Hyperactive Osteoclastogenesis in Bone Remodeling with Antioxidant Capacity. bioRxiv 2022. [Google Scholar] [CrossRef]
- Ivashkevich, A.N.; Martin, O.A.; Smith, A.J.; Redon, C.E.; Bonner, W.M.; Martin, R.F.; Lobachevsky, P.N. γH2AX foci as a measure of DNA damage: A computational approach to automatic analysis. Mutat. Res. Mol. Mech. Mutagen. 2011, 711, 49–60. [Google Scholar] [CrossRef]
- Ojeda, S.R.; Romero, C.; Tapia, V.; Dissen, G.A. Neurotrophic and cell–cell dependent control of early follicular development. Mol. Cell. Endocrinol. 2000, 163, 67–71. [Google Scholar] [CrossRef]
- Chang, H.-M.; Wu, H.-C.; Sun, Z.-G.; Lian, F.; Leung, P.C.K. Neurotrophins and glial cell line-derived neurotrophic factor in the ovary: Physiological and pathophysiological implications. Hum. Reprod. Updat. 2019, 25, 224–242. [Google Scholar] [CrossRef]
- Ziegenhorn, A.A.; Schulte-Herbrüggen, O.; Danker-Hopfe, H.; Malbranc, M.; Hartung, H.-D.; Anders, D.; Lang, U.E.; Steinhagen-Thiessen, E.; Schaub, R.T.; Hellweg, R. Serum neurotrophins—A study on the time course and influencing factors in a large old age sample. Neurobiol. Aging 2007, 28, 1436–1445. [Google Scholar] [CrossRef]
- Pelosi, E.; Simonsick, E.; Forabosco, A.; Garcia-Ortiz, J.E.; Schlessinger, D. Dynamics of the Ovarian Reserve and Impact of Genetic and Epidemiological Factors on Age of Menopause1. Biol. Reprod. 2015, 92, 127381. [Google Scholar] [CrossRef]
- Bressler, L.H.; Steiner, A. Anti-Müllerian hormone as a predictor of reproductive potential. Curr. Opin. Endocrinol. Diabetes 2018, 25, 385–390. [Google Scholar] [CrossRef]
- Tsoukalas, D.; Fragkiadaki, P.; Docea, A.O.; Alegakis, A.K.; Sarandi, E.; Thanasoula, M.; Spandidos, D.A.; Tsatsakis, A.; Razgonova, M.P.; Calina, D. Discovery of potent telomerase activators: Unfolding new therapeutic and anti-aging perspectives. Mol. Med. Rep. 2019, 20, 3701–3708. [Google Scholar] [CrossRef] [PubMed]
- Sadowska-Bartosz, I.; Bartosz, G. Effect of Antioxidants Supplementation on Aging and Longevity. BioMed Res. Int. 2014, 2014, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Godman, C.A.; Joshi, R.; Giardina, C.; Perdrizet, G.; Hightower, L.E. Hyperbaric oxygen treatment induces antioxidant gene expression. Ann. N. Y. Acad. Sci. 2010, 1197, 178–183. [Google Scholar] [CrossRef]
- Hachmo, Y.; Hadanny, A.; Mendelovic, S.; Hillman, P.; Shapira, E.; Landau, G.; Gattegno, H.; Zrachya, A.; Daniel-Kotovsky, M.; Catalogna, M.; et al. The effect of hyperbaric oxygen therapy on the pathophysiology of skin aging: A prospective clinical trial. Aging 2021, 13, 24500–24510. [Google Scholar] [CrossRef] [PubMed]
- Hachmo, Y.; Hadanny, A.; Hamed, R.A.; Daniel-Kotovsky, M.; Catalogna, M.; Fishlev, G.; Lang, E.; Polak, N.; Doenyas, K.; Friedman, M.; et al. Hyperbaric oxygen therapy increases telomere length and decreases immunosenescence in isolated blood cells: A prospective trial. Aging 2020, 12, 22445. [Google Scholar] [CrossRef] [PubMed]
- Bray, G.; Edelstein, S.; Crandall, J. Long-Term Safety, Tolerability, and Weight Loss Associated With Metformin in the Diabetes Prevention Program Outcomes Study. Diabetes Care 2021, 35, 731–737. [Google Scholar] [CrossRef]
- Davis, S.R.; Panjari, M.; Stanczyk, F.Z. DHEA Replacement for Postmenopausal Women. J. Clin. Endocrinol. Metab. 2021, 96, 1642–1653. [Google Scholar] [CrossRef]
- Chern, C.U.; Tsui, K.H.; Vitale, S.G.; Chen, S.N.; Wang, P.H.; Cianci, A.; Tsai, H.W.; Wen, Z.H.; Lin, L.T. Dehydroepiandrosterone (DHEA) supplementation improves in vitro fertilization outcomes of poor ovarian responders, especially in women with low serum concentration of DHEA-S: A retrospective cohort study. Reprod. Biol. Endocrinol. 2018, 16, 90. [Google Scholar] [CrossRef]
- Samaras, N.; Samaras, D.; Frangos, E.; Forster, A.; Philippe, J. A Review of Age-Related Dehydroepiandrosterone Decline and Its Association with Well-Known Geriatric Syndromes: Is Treatment Beneficial? Rejuven. Res. 2013, 16, 285–294. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author | Pharmaceutical | Design and Method | Ovarian Effect | Conclusions |
|---|---|---|---|---|
| Qin et al., 2019 [50] | Metformin | Mouse study: Female 28-month-old mice given 100 mg/kg metformin (MET) for 6 months, repeated with a control group | MET had higher primordial follicle count (MET ~10 vs. control ~5, p < 0.01), primary follicle count (MET ~60 vs. control ~40, p < 0.05) and SIRT-1 levels (MET ~1.6 vs. control 1.0, p < 0.05). MET had lower 8-OHdG, (MET ~30 vs. control ~40, p < 0.05), 4-HNE (MET ~30 vs. control ~50, p < 0.05) and p16 (MET ~30 vs. control ~40) | Metformin delays ovarian aging in mice |
| Park et al., 2020 [51] | Visfatin | Mouse study: 18-month-old mice were injected with 0.1 mL 500 ng/mL Visfatin or 1000 ng/mL three times at two-day intervals. Experiment 1: 12 mice split into 0 (control), 500 or 1000 ng/mL Visfatin groups. Experiment 2: 12 18-month-old female mice split into two groups: 500 ng/mL Visfatin and 1000 ng/mL. The mice were mated with males | Experiment 1: number of primary, secondary and early antral follicles higher than control in Visfatin groups. For 500 ng/mL: (primary 26.0 ± 1.5 vs. control 14.0 ± 1.5, p < 0.05), (secondary 11.0 ± 0.6 vs. control 6.0± 1.0, p < 0.05, antral 6.0 ± 0.5 vs. control 1.0 ± 0.6, p < 0.01). For 1000 ng/mL: (primary 26.0 ± 2.1 vs. control 14.0 ± 1.5, p < 0.05), (secondary 10.0 ± 0.6 vs. control 6.0 ± 1.0, p < 0.05, antral 4.0 ± 0.6 vs. control 1.0 ± 0.6, p < 0.05). Atretic follicles lower than control for Visfatin groups (500 ng/mL 6.0 vs. 13.0 control, p < 0.05, 1000 ng/mL 7.0 vs. 13.0 control, p < 0.05). Experiment 2: 500 ng/mL had more pups per female (2.20 ± 1.30 vs. control 0, p < 0.05) | In mice, Visfatin may recover ovarian function and fertility for post-menopausal aged mice |
| Guzel et al., 2018 [52] | gS1P | In vitro study: Ovarian cortical tissue from patients. The samples were incubated with S1P concentrations of either 0 (control), 200 or 400 μM for 4 days | Primordial and secondary follicle counts higher in 400 μM and 200 μM vs. control. For 400 μM: (primordial: 2.3 ± 0, primordial control 0.8 ± 0.2, p < 0.01), (secondary: 0.7 ± 0.1, control 0.1 ± 0.05, p < 0.05). For 200 μM: (primordial 200 μM 1.7 ± 0.2, control, 0.8 ± 0.2, p < 0.05), (secondary 200 μM 0.4 ± 0.1 vs. control 0.1 ± 0.05, p < 0.05). Number of follicles with positive cleaved caspase 3 marker lower in 200 μM than control (0.8 ± 0.05 vs. control 1.2 ± 0.1, p < 0.05) and in 400 μM as well (0.2 ± 0.0.5 vs. control 1.2 ± 0.1, p < 0.01) | Under in vitro conditions, S1P inhibits follicular apoptosis and improves survivability in the human ovary |
| Mumusoglu et al., 2018 [53] | S1P | Rat study: 30 female 10-month-old rats divided into three groups: (A) 0.1 mg/kg Fingolimod (S1P analogue), (B) 1 mg/kg Fingolimod, (C) control. Duration: 60 days | B had higher AMH values (Group B 5.72 ± 0.61 ng/mL vs. Group C 4.81 ± 0.85 ng/mL, p = 0.05). A had a higher non-apoptotic ratio (Group A 67.0% ± 16.4% vs. Group C 29.9% ± 19.5%, p < 0.001). B had a higher non-apoptotic follicle ratio (Group B; 51.1% ± 11.5% vs. Group C 29.9% ± 19.5%, p = 0.023) | S1P may decrease spontaneous follicular apoptosis given its inhibition of the ceramide-induced death pathway |
| Author | Stem Cell Type | Method | Ovarian Effect | Conclusions |
|---|---|---|---|---|
| Ding et al., 2018 [17] | hAMSC | Mouse study: female mice 12–14 months of age were split into four groups (n = 15 per group and injected with hepatocyte growth factor (HGF), epidermal growth factor (EGF), both or PBS (control). Duration: 4 weeks | The hAMSC group had higher primordial (88% higher, p < 0.001), primary (89% higher, p < 0.001), secondary (86% higher, p < 0.01) and antral follicle counts (81% higher, p < 0.001) and AMH than the control (92% higher, p < 0.001) | hAMSCs inhibit ovarian aging through HGF and EGF secretion |
| Yang et al., 2020 [61] | Human umbilical cord mesenchymal stem cell exosomes (UC-MSCexos) | Mouse study: 10-month-old mice were injected with UC-MSCexos. Three weeks later, the ovaries were collected. Another 10-month-old group was injected with either UC-MSCexos or PBS (control) | Primordial follicle % lower in exo group (exo ~14%, control ~17%, p < 0.05), late antral % higher in exo group (exo ~12%, ~9% control, p < 0.05). Exo group had 2.5× mean number of pups per mouse vs. control (4.5 vs. 2). Exo group % of aberrant spindles 20%, control ~38%, p < 0.01. ROS fluorescence intensity in exo group ~20 vs. control ~25, p < 0.05. Mitochondrial fluorescence intensity in exo group ~1.6 vs. control 1.0, p < 0.01 | UC-MSCexos induce follicular maturation and could aid in restoring fertility |
| Jiao et al., 2022 [62] | Umbilical cord mesenchymal stem cells (UC-MSC) | Mouse study: 9–10-month-old female mice injected with either a) 5 μL MSC-CM + 0.3 mg/mL HA, b) 5 μL MSC-CM + 0.3 mg/mL HA + 1 μg/mL HGF neutralising antibody, or c) 800 ng/mL HGF + 0.3 mg/mL HA | Primary (p = 0.046), secondary (p = 0.003), antral (p = 0.032) and total follicle counts higher in MSC/HA group than control (p < 0.05). Higher pups/litter than control (3.33 ± 2.02 vs. 1.6 ± 1.41, p = 0.042) | UC-MSC have may prolong natural aging in mice by protecting the follicular reserve |
| Kim K. et al., 2022 [63] | Human placenta-derived mesenchymal stem cells (hPD-MSC) | Rat study: Rats split into four groups. 1: injected with 0.2 mL PUBS containing 5 × 105 hPD-MSCs. 2: injected with the same dosage three times, at 10-day intervals. 3: A group injected with only PBS (positive control). 4: A group not injected (negative control) Each group n = 24 | For single-injection therapy, AMH levels had an increase in week 5 (hPD ~1.5, control 0.75, p < 0.05). In week 5, miR-16-5p, miR-34a-5p and miR-191-5p significantly decreased (p < 0.05) in hPD-MSC group compared to control (miR-16-5p: control ~1 vs. hPD-MSC ~0.5), (miR-34a-5p: control ~1 vs. hPD-MSC ~0.8), (miR-191-5p: control ~1.1 vs. hPD-MSC ~0.8) | hPD-MSC transplantation improves ovarian function in menopausal rats and alleviates symptoms of ovarian aging |
| Kim G. et al., 2018 [64] | Human endothelial progenitor cells (hEPC) | Mouse study: 4- and 6-month-old female mice injected twice with 5 × 104 cells at a 4-day interval. hEPCs from four healthy human donors. After being cultured, the hEPCs were injected in a suspension of PBS into each mouse. PBS-only served as control | Ifn-γ and Il-1β both lower in 4- and 6-month-old mice vs. controls. Ifn-γ: (4-month hEPC ~0.25 vs. control 1.5, p < 0.01), (6-month ~0.5 vs. control ~1.5, p < 0.001). Il-1β: (4-month hEPC ~0.1 vs. control ~1.0, p < 0.05), (6 month ~0.25 vs. control ~1.25, p < 0.05). Tnfα lower in 6-month-old mice (hEPC ~1.5 vs. control ~2.5, p < 0.01). Bcl2 higher in both (4-month ~3 vs. control ~1, p < 0.05) (6-month ~1.5 vs. control ~1, p < 0.01) | hEPCs may improve ovarian quality with aging |
| Ra et al., 2020 [65] | Human adipose stem cell conditioned medium (ASC-CM) | Mouse study: ASCs were isolated from a 39-year-old healthy woman. Then, 4- and 6-month-old female mice were randomly divided into control (PBS) and treatment groups. Treatment group given a medium of ASC, either three times in 8-day intervals (8D-3T) or six times in 4-day intervals (4D-6T) | The 6-month-old mice of the 4D-6T group had a higher number of foetuses than the control (4D-6T ~15 vs. control ~7, p < 0.05). Ovaries of pregnant mice had lower caspase 3 mRNA levels for both interval groups vs. control (3T-8D ~0.3, 6T-4F ~0.5, control 1, p < 0.01 for both) | ASC-CM has antioxidant effects that improve oocyte quality in aging mice |
| Wang et al., 2022 [66] | Menstrual blood-derived mesenchymal stem cells (MenSC) | Mouse study: 8-month-old C57BL/6J mice were divided into three groups: MenSC injected intraperitoneally, in the ovaries and through tail intravenous injection. MenSC concentration: 6 × 105 MSCs in 50 μL normal saline | At 8 weeks post-transplantation, the MenSC group injected directly in the ovaries had significantly lower levels of atresia compared to the control (~28 vs. 45, p < 0.01). The relative mtDNA amount in the ovaries was significantly higher in this group at 40 weeks post-treatment (~2 vs. ~0.8, p < 0.05) compared to the control. The MenSC group also had a significantly higher number of antral follicles (~7 vs. ~4, p < 0.01). In IVF, the MSC group had a significantly higher blastocyst formation rate (~55% vs. ~35%, p < 0.01) | Blood-derived MenSCs may aid in fertility preservation and also assist in IVF as a treatment |
| Jiao et al., 2022 [62] | UC-MSCs and autocrosslinked hyaluronic acid (HA) | Mouse study: 9/10-month-old C57BL/6J female mice were transplanted with 5 μL UC-MSC + 0.3 mg/mL HA + 1 μg/mL HGF. After 14 days observation, mice were either sacrificed for analysis or allowed to mate | The MSC/HA group had a significantly higher number of total follicles compared to age-matched controls (~500 vs. ~300, p < 0.05) and a higher number of primary, secondary and antral follicles (~200 vs. ~100, p < 0.05 for all three). Significantly higher number of pups/litter (~4 vs. ~2, p < 0.05) | Autocrosslinked HA boosts the effect of UC-MSCs in the ovaries. Together they restore ovarian function in naturally aged mice |
| Zhang et al., 2023 [67] | Mitochondria supplementation from endometrial mesenchymal stem cells (EN-MSC) | In vitro and mouse study: 10-month-old mice. Mitochondria from EN-MSCs were injected into germinal vesicle-stage oocytes of aged mice | The % of normal spindles in MII oocytes of aged mice was significantly higher in the study group than control (~60% vs. ~40%, p < 0.05). The rate of aneuploidy was significantly lower (~40% vs. ~60%, p < 0.05). The membrane potential was significantly higher (ΔΨm ~1.25 vs. ~0.4, p < 0.0001). When transplanted into mice via IVF, fertilised oocytes also had significantly higher live birth rates (~25% vs. ~12%, p < 0.05) and significantly higher blastocyst formation rates as well (~70% vs. ~50%, p < 0.05), significantly higher % live birth rates (~25% vs. ~12%, p < 0.05) | Mitochondrial supplementation from sources such as EN-MSCs has the potential to restore ovarian function in aged mice and also improve IVF outcomes |
| Author | Pharmaceutical | Design and Method | Ovarian Effect | Conclusions |
|---|---|---|---|---|
| Luo et al., 2012 [81] | CR | Rat study: 48 female 2-month-old Sprague Dawley rats were randomly divided into four groups: control, 25% CR group, 45% CR group, high-fat diet group (HF) | The 45CR group had higher levels of primary follicles vs. control (control 19.40 ± 3.99, 45CR 56.71 ± 5.60, p < 0.01). The % of primordial follicles was higher for 25CR and 45CR than the control (control 20.5 ± 3.5%, 25CR 41.9 ± 2.9% (p < 0.01), 45CR 44.4 ± 2.2% (p < 0.001)). The % of antral follicles was lower in the 25CR and 45CR groups than the control (control ~30%, 25 CR ~18% (p < 0.05), 45CR ~17% (p < 0.05). SIRT1 was higher in the 25CR group than control (p < 0.001) | Caloric restriction, especially moderate, maintains the follicular reserve and inhibits ovarian aging |
| Isola et al., 2022 [82] | CR | Mouse study: 72 female mice were divided into four groups: control, 17α-E2, 10% caloric restriction (CR10) and 30% caloric restriction (CR30). Duration: 24 weeks | Primordial follicle count was higher in CR10 (follicles/mm2 CR10 ~3.5 vs. control ~2.8, p < 0.05). Transitional follicle count was higher in both CR groups than the control (control ~1.8 vs. CR10 ~2 vs. CR30 ~30). Primary follicle count was higher in CR30 (CR30 ~2.5 vs. control ~1, p < 0.05). CR30 had higher pregnancy rate vs. control (100% vs. 44.44%, p < 0.05) | CR may prolong ovarian reserve and, thus, increase fertility lifespan in mice |
| Garcia et al., 2019 [83] | CR and rapamycin | Mouse study: 36 1-month-old C57BL/6 mice were split into three groups: control, rapamycin (rapa), 30% CR. Mice in the rapamycin group received 4 mg/kg rapamycin every other day for 93 days. Afterwards, mice were euthanised and their ovaries were examined | Number of primordial follicles was higher in rapa (p = 0.04) and CR (p = 0.02) vs. control (rapa: ~12,000 CR: ~13,000, control: ~6000). Number of primary and secondary follicles was lower in both groups vs. control (control ~4000, rapa ~1800 (p < 0.05), CR ~1800 (p < 0.05) and control ~650, rapa ~300 (p < 0.5), CR ~300 (p < 0.05)). Number of tertiary follicles was lower in CR (control ~220, CR ~110, p = 0.005) | Rapamycin and CR both show signs that they might prolong reproductive lifespan by inhibiting primordial follicle activation |
| Author | Pharmaceutical | Design and Method | Ovarian Effect | Conclusions |
|---|---|---|---|---|
| Chu et al., 2017 [90] | DHEA | Mouse study: 15 mice were divided into three groups: E2 group, receiving 100 μg/day β-oestradiol. DHEA group receiving 5 mg/day DHEA. Control group receiving saline. Experiment was carried on for 4 weeks. | DHEA had (p < 0.05) higher primordial (14.77 ± 0.6994 vs. 11.58 ± 1.264), primary (18.42 ± 0.7732 vs. 16.25 ± 0.5383) and secondary (8.583 ± 0.8744 vs. 8.333 ± 0.6195) follicle count vs. control. %TUNEL-positive Follicles lower in DHEA vs. control (40.77 ± 1.382 vs. 45.59 ± 1.653, p < 0.05) as well as %γ H2AX-positive follicles (53.77 ± 1.424 vs. 61.16 ± 2.288, p < 0.05) | DHEA promotes folliculogenesis, inhibits ovarian apoptosis and decreases DSB occurrences in oocytes |
| Ma et al., 2022 [91] | Hyperbaric oxygen treatment (HBOT) | Mouse study: 40-week-old female mice were administered 100% oxygen at 2.5 ATA pressure for 90 min. Duration: 10 consecutive days. Afterwards, mice were sacrificed for oocyte analysis and extraction for IVF | There were significantly higher serum AMH levels vs. control (~550 ng/L vs. ~250, p < 0.05). Significantly lower apoptotic nuclei vs. control (mean density ~55 vs. ~45, p < 0.05). The number of retrieved oocytes and number of blastocyst formation were significantly higher vs. control as well (7 vs. 3, p < 0.05 and 5 vs. 1, p < 0.05, respectively) | HBOT may improve the quality of oocytes |
| Xiao et al., 2022 [92] | Macrophage-derived extracellular vesicles | Mouse study: 10-month-old ICR female mice were injected with either PBS (control), M1 or M2 extracellular vesicles. Injections occurred five times over a period of 5 days | The M2 group had a significantly higher number of primordial cells (~45 vs. ~38, p < 0.05). Growing follicle % was significantly lower in the M2 group vs. control (~35% vs. ~25%). Relative intensity of p-AKT significantly was higher in M1 vs. control (1.5 vs. 1.25, p < 0.05) and significantly lower in M2 vs. control (1 vs. 1.25, p < 0.05). Intensity of p-RSP6 was significantly higher in M1 vs. control (1.2 vs. 1, p < 0.05) and significantly lower in M2 group (0.75 vs. 1, p < 0.05). Post-superovulation, the M2 group had a significantly higher number of retrieved oocytes vs. control (~12 vs. ~7, p < 0.05) | M2 extracellular vesicle injection may alleviate symptoms of ovarian aging through inflammation reduction |
| Liu et al., 2022 [93] | Cell-free fat extract (CEFFE) | Mouse study: Mice were divided into three groups: (A) 8-week-old mice (young control), (B) 10-month-old mice (old control) and (C) 10-month-old mice, given 200 μL CEFFE through the tail every other day. Study duration: 2 weeks | At 8 weeks post-treatment, the study group had significantly higher AMH levels compared to the control (~2300 pg/mL vs. ~2000, p < 0.001). It also had a significantly higher no. of primary (~4 vs. ~2, p < 0.05), secondary (~6 vs. ~2, p < 0.05) and total follicles (~14 vs. ~4, p < 0.01), as well as a significantly greater blastocyst formation rate (41.67 ± 37.27 vs. 3.57 ± 9.45, p < 0.05) and litter number (6.63 ± 2.07 vs. 3.25 ± 2.05 p < 0.01)and significantly lower relative γH2AX staining than the control (~5 vs. ~10, p < 0.01) | While the exact mechanism remains unknown, CEFFE provides and restores function in aged ovaries |
| Qin et al., 2022 [94] | TrkB agonist (Ab4B19) | Mouse study: Six 12-month-old C57BL female mice were divided into two groups, one treated with Ab4B19 and the other with just IgG. Duration: 16 days. The mice were then either mated to analyse fertility or were sacrificed for ovarian examination | There was a significantly (p < 0.05) higher primordial (~500 vs. ~400), preantral (~350 vs. ~250) and antral follicle count (~250 vs. ~150) per ovary vs. vehicle and significantly lower atretic follicle count vs. vehicle (~100 vs. ~150, p < 0.0001). There were significantly more ovulated oocytes per mouse (~28 vs. ~18, p = 0.0284) and a significantly larger mean litter size in the TrkB group vs. vehicle (~8 vs. ~6, p = 0.0017) | TrkB agonist treatment may have potential in prolonging the reproductive window as well as treating ovarian aging |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valtetsiotis, K.; Valsamakis, G.; Charmandari, E.; Vlahos, N.F. Metabolic Mechanisms and Potential Therapeutic Targets for Prevention of Ovarian Aging: Data from Up-to-Date Experimental Studies. Int. J. Mol. Sci. 2023, 24, 9828. https://doi.org/10.3390/ijms24129828
Valtetsiotis K, Valsamakis G, Charmandari E, Vlahos NF. Metabolic Mechanisms and Potential Therapeutic Targets for Prevention of Ovarian Aging: Data from Up-to-Date Experimental Studies. International Journal of Molecular Sciences. 2023; 24(12):9828. https://doi.org/10.3390/ijms24129828
Chicago/Turabian StyleValtetsiotis, Konstantinos, Georgios Valsamakis, Evangelia Charmandari, and Nikolaos F. Vlahos. 2023. "Metabolic Mechanisms and Potential Therapeutic Targets for Prevention of Ovarian Aging: Data from Up-to-Date Experimental Studies" International Journal of Molecular Sciences 24, no. 12: 9828. https://doi.org/10.3390/ijms24129828
APA StyleValtetsiotis, K., Valsamakis, G., Charmandari, E., & Vlahos, N. F. (2023). Metabolic Mechanisms and Potential Therapeutic Targets for Prevention of Ovarian Aging: Data from Up-to-Date Experimental Studies. International Journal of Molecular Sciences, 24(12), 9828. https://doi.org/10.3390/ijms24129828