
Exploring the Inhibitory Effect of AgBiS2 Nanoparticles on Influenza Viruses

 and
and 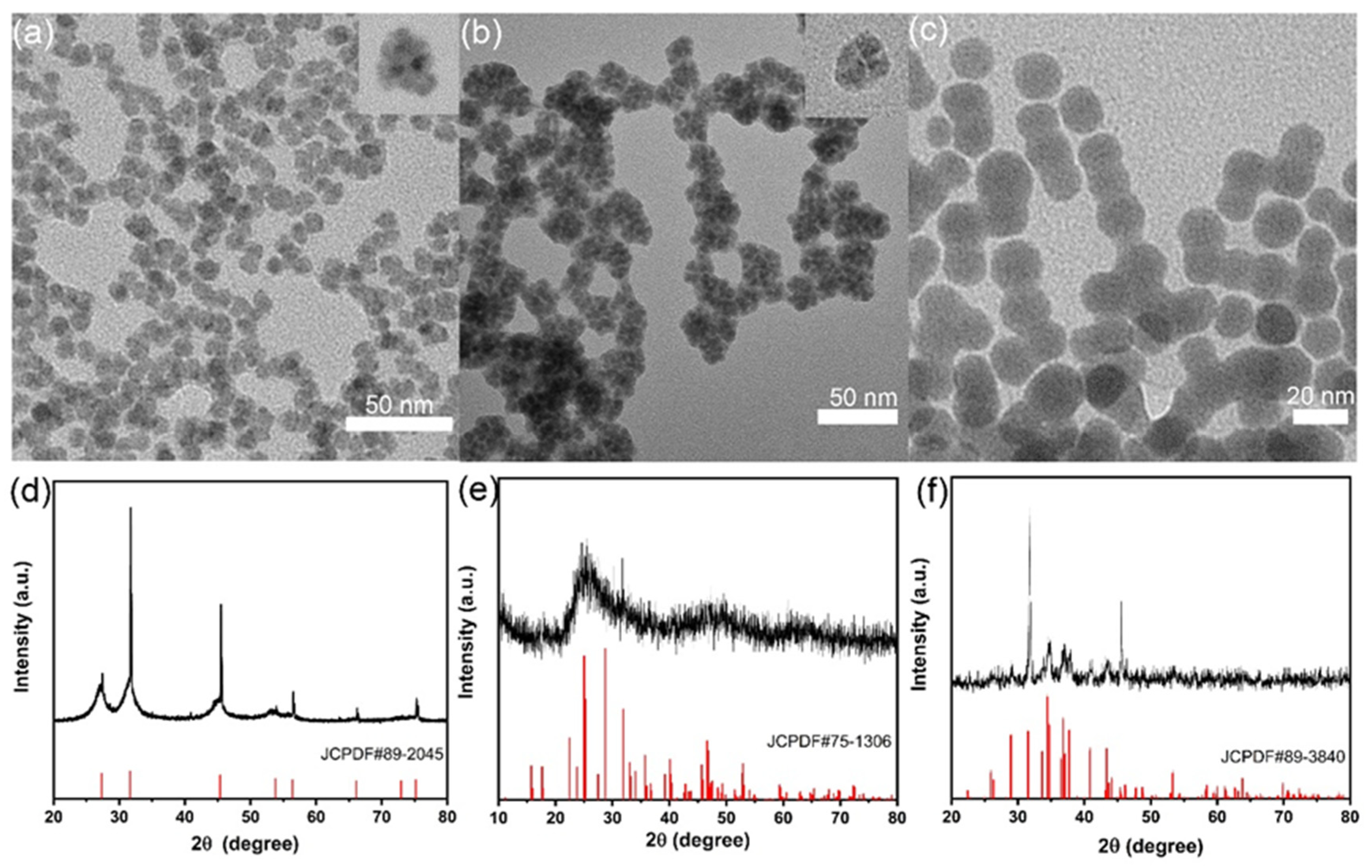{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
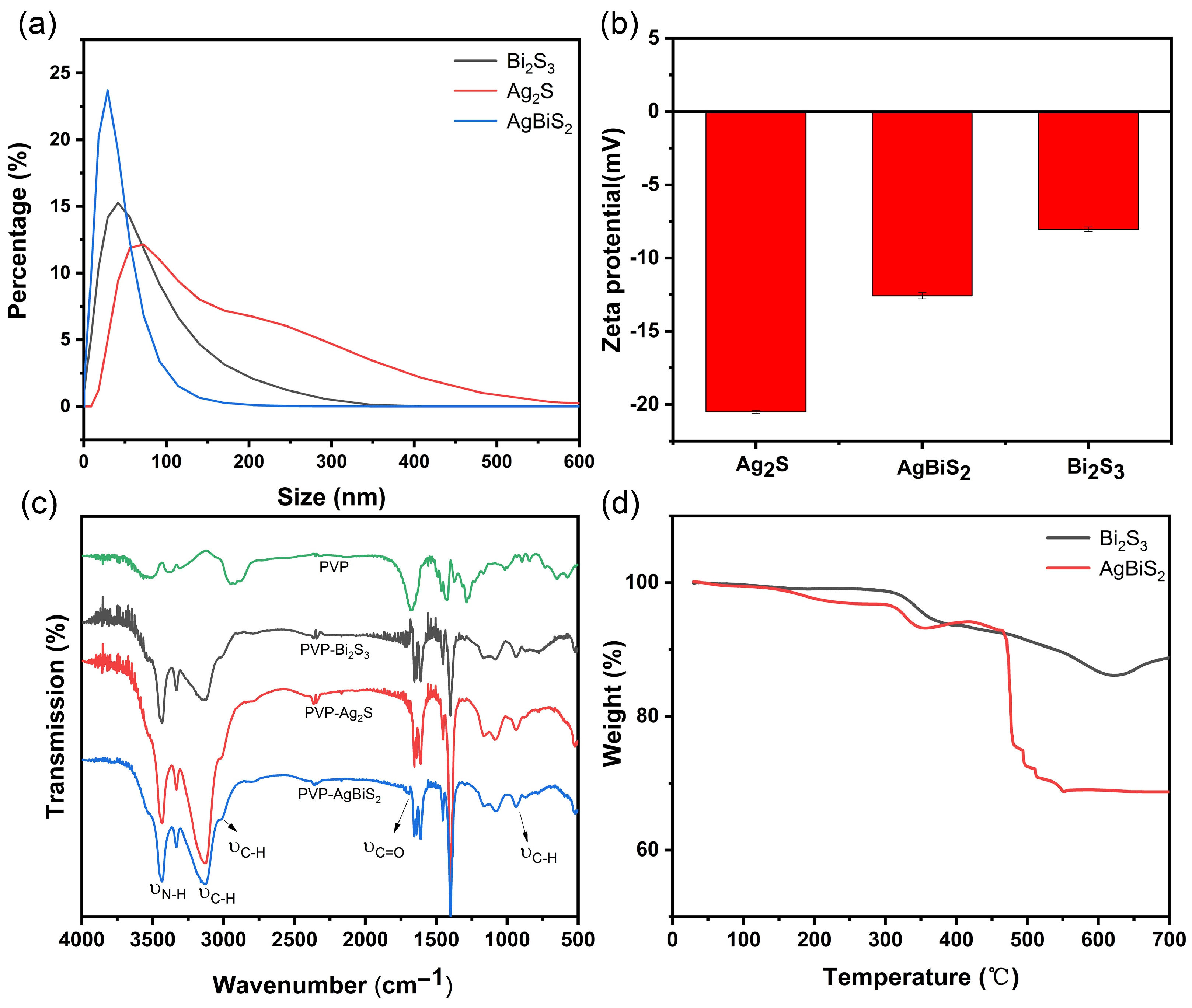
2.1. Material Characterization
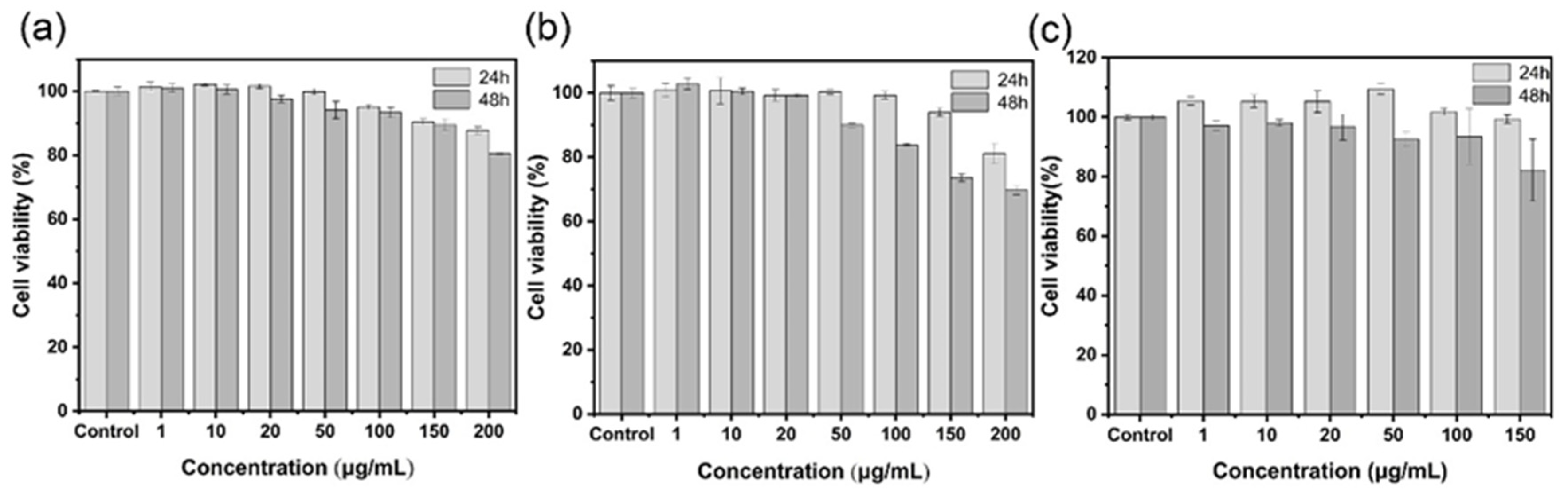
2.2. Cytotoxicity Test
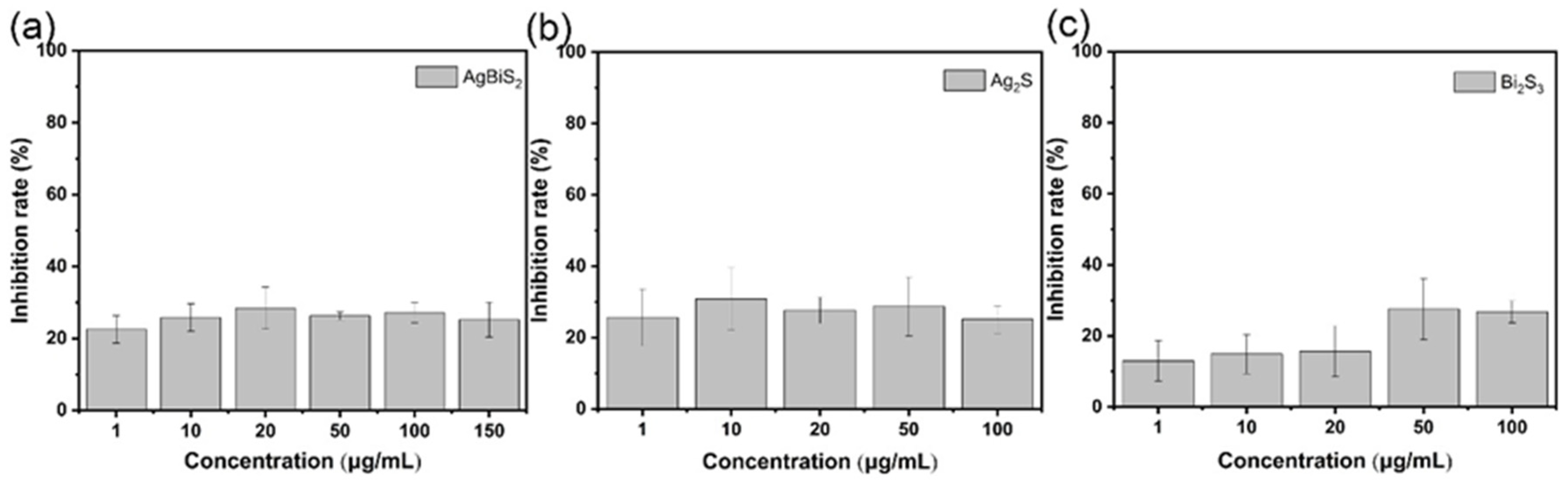
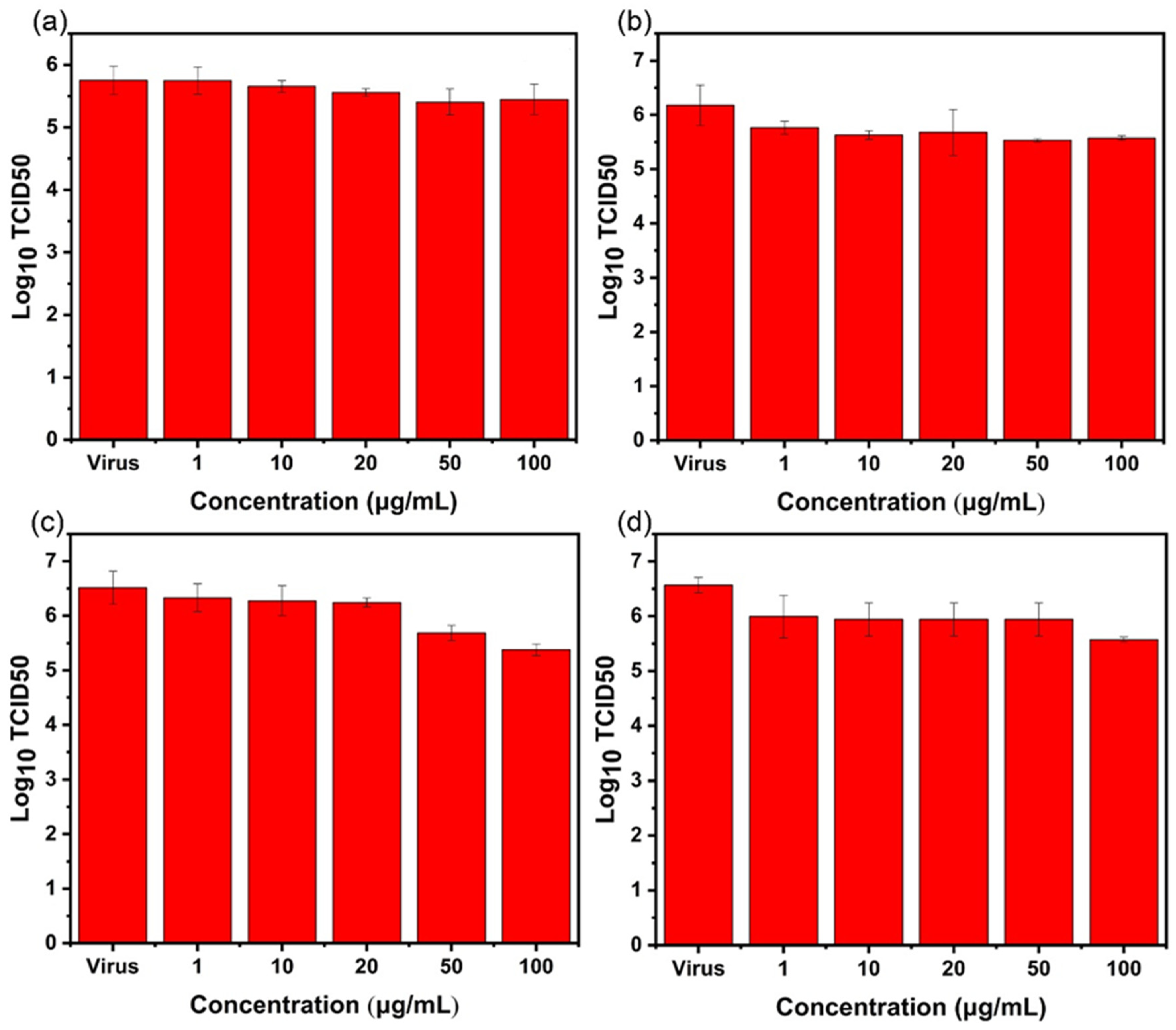
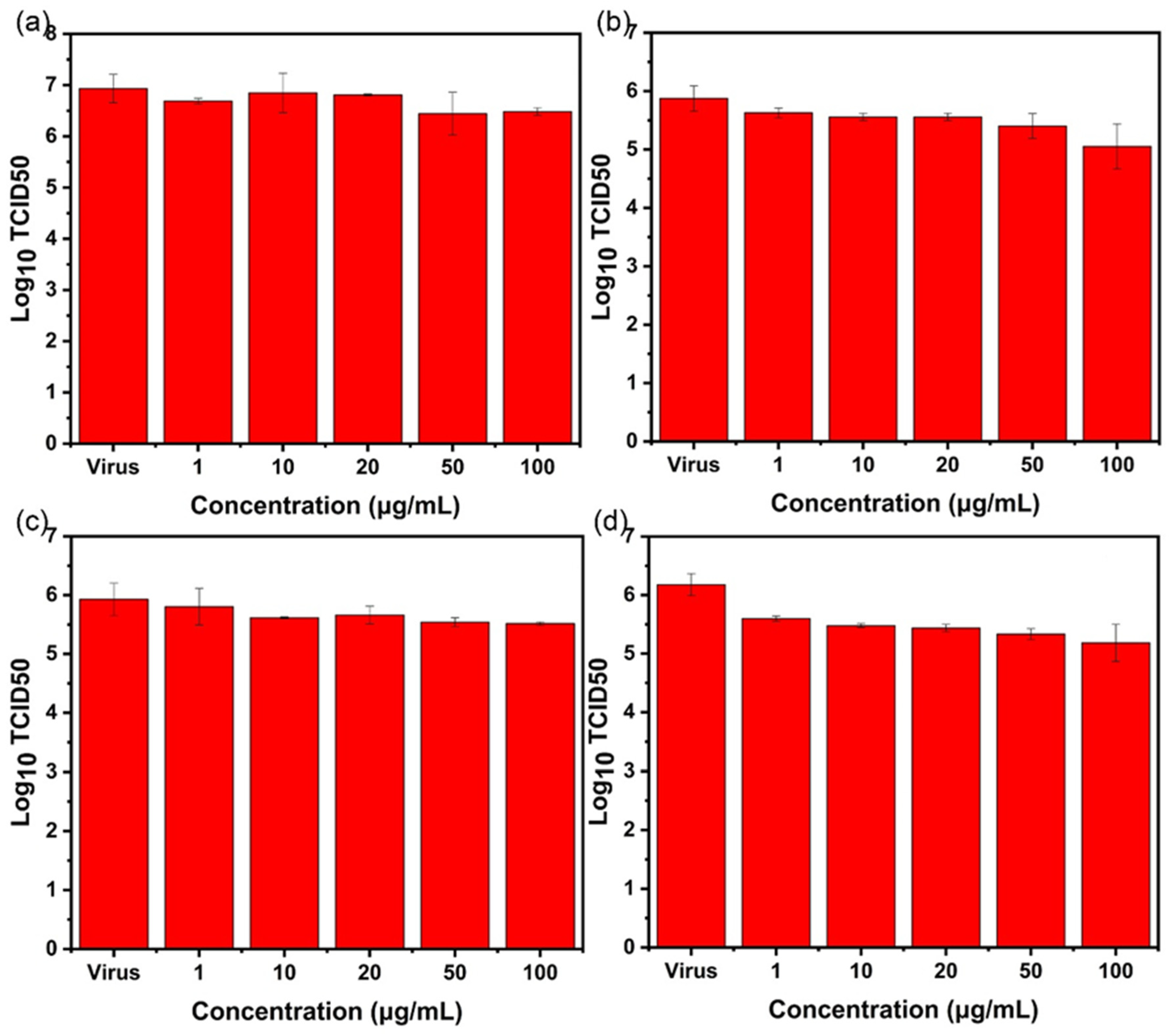
2.3. Antiviral Effect Evaluation Experiment
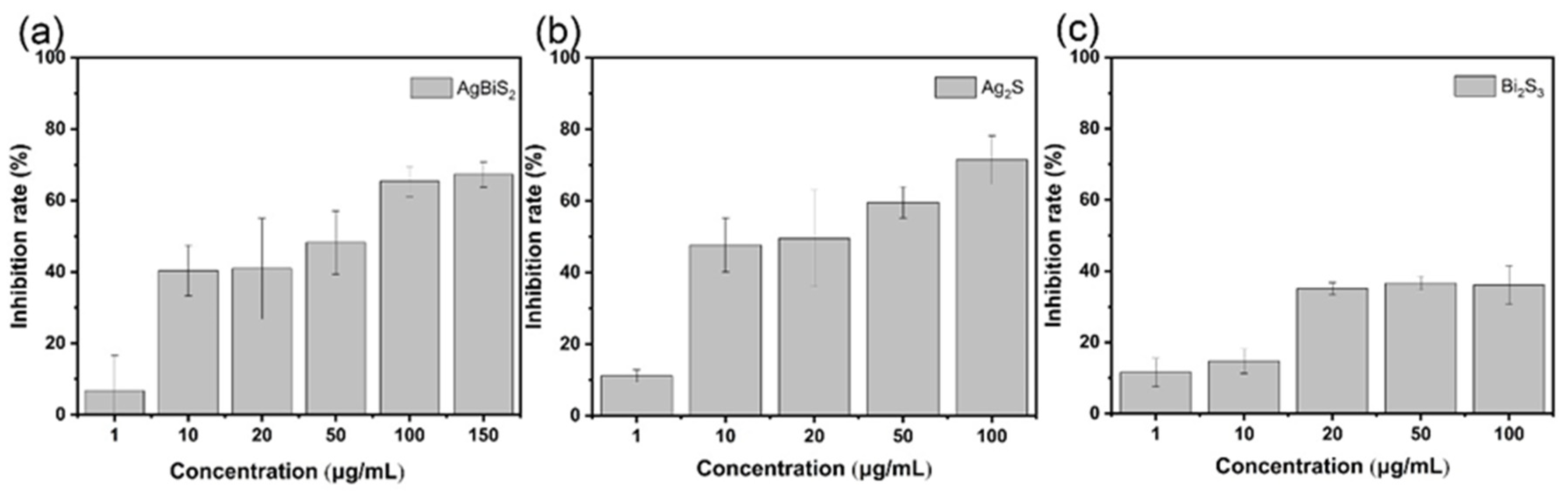
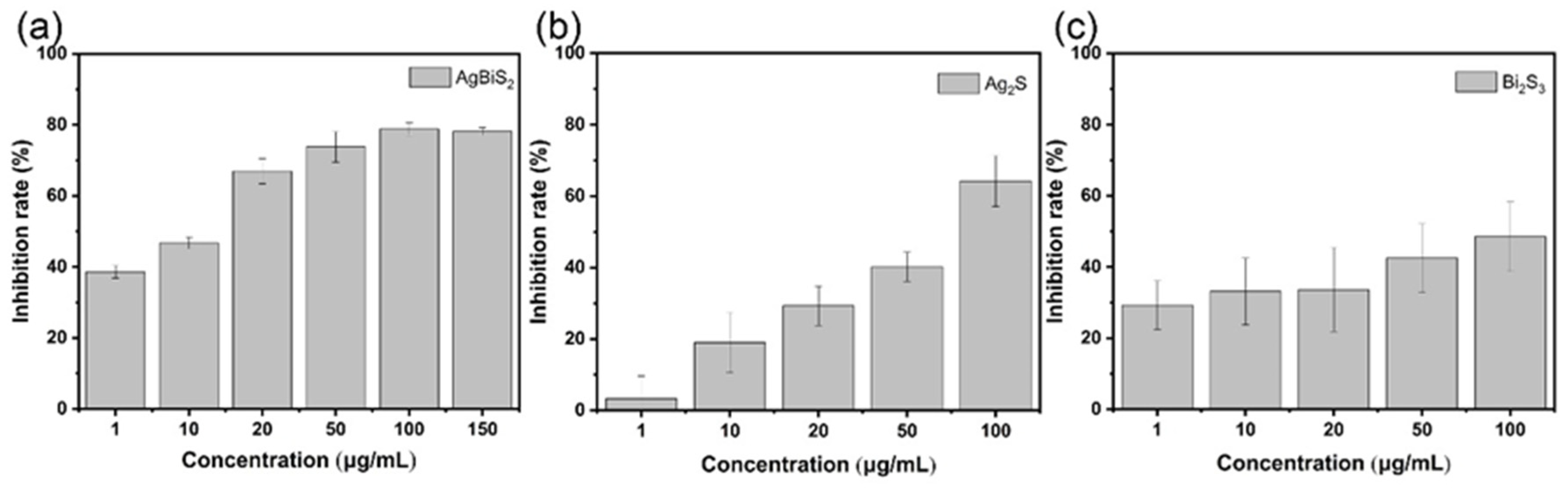
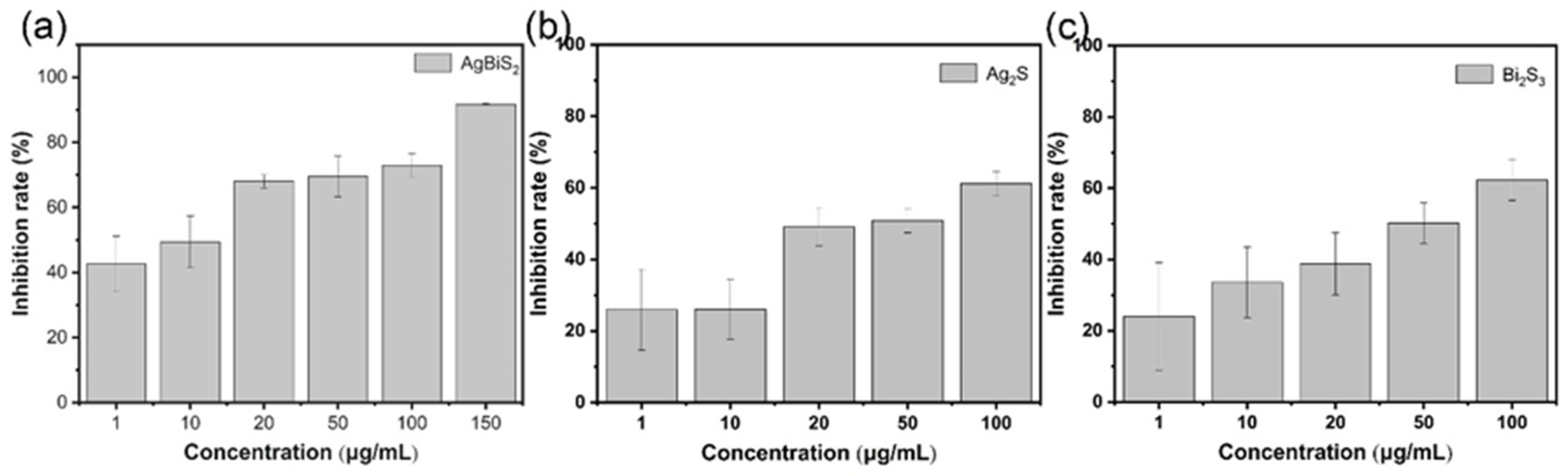
2.3.1. Direct Inactivating Group
2.3.2. Co-Cultivation Inhibition Group
2.3.3. Nanoparticle Preculture Inhibition Group
2.3.4. Virus Post-Infection Inhibition Group
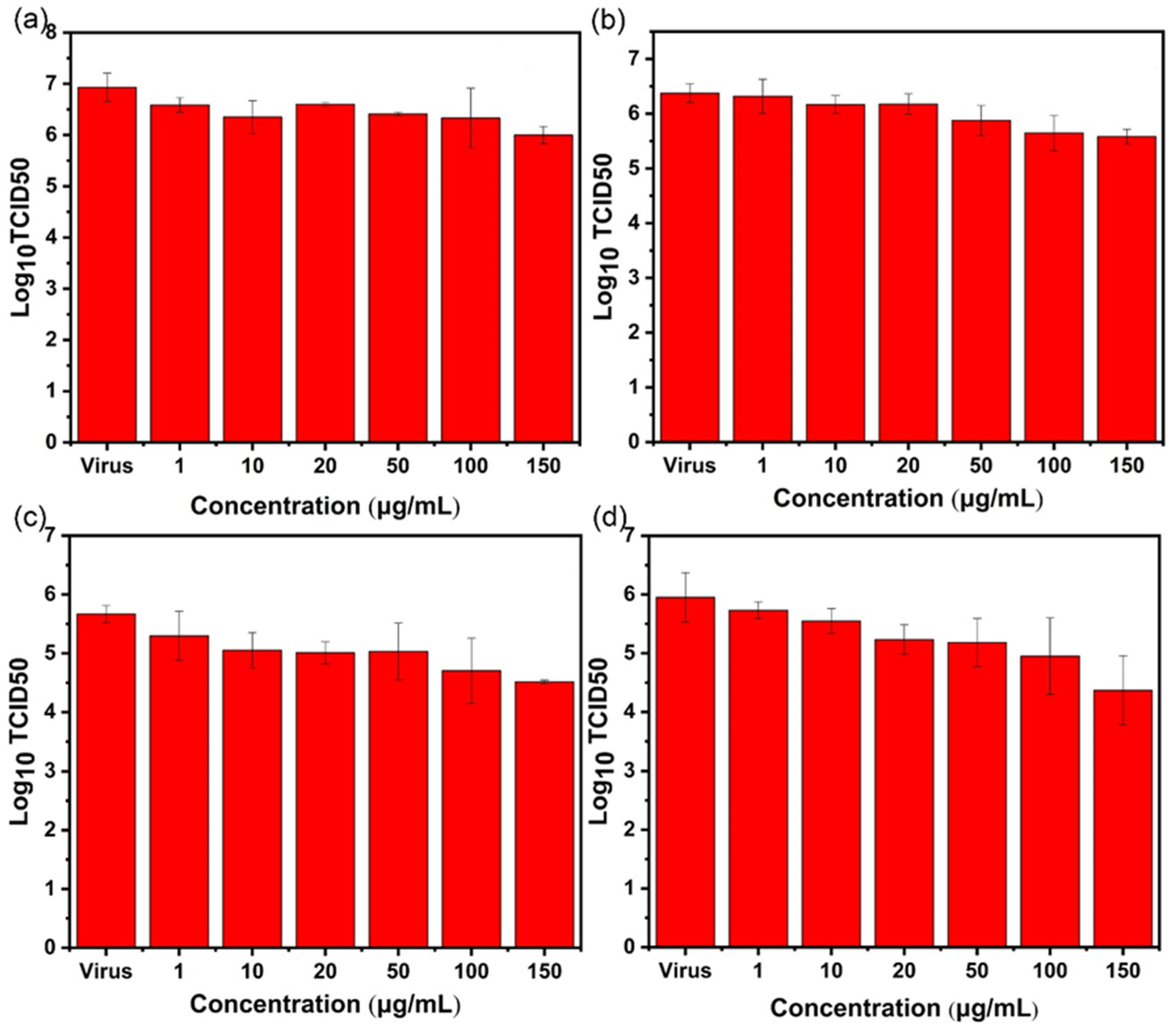
2.4. TCID50 Test for Virus Activity
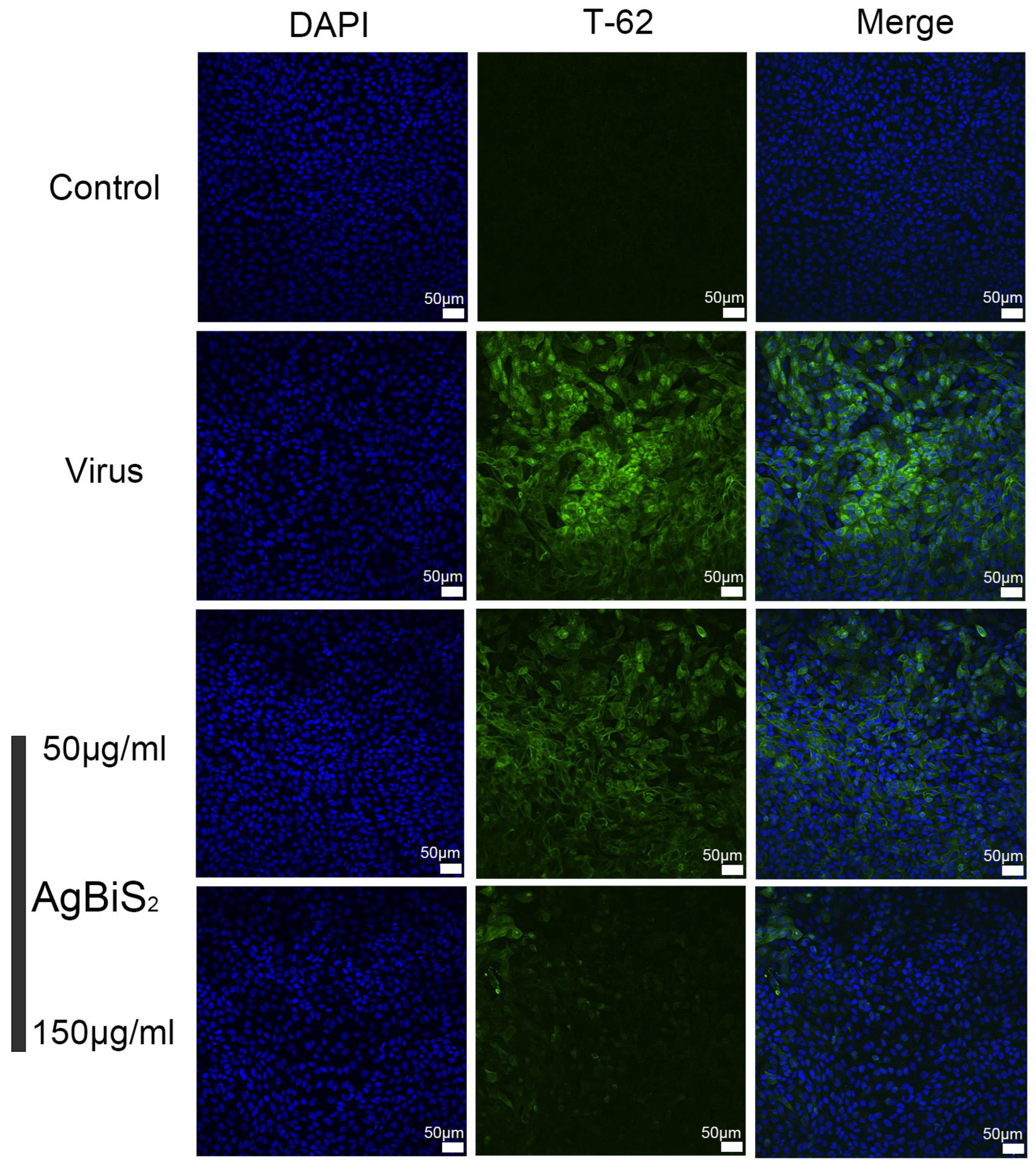
2.5. Indirect Immunofluorescence Experiment
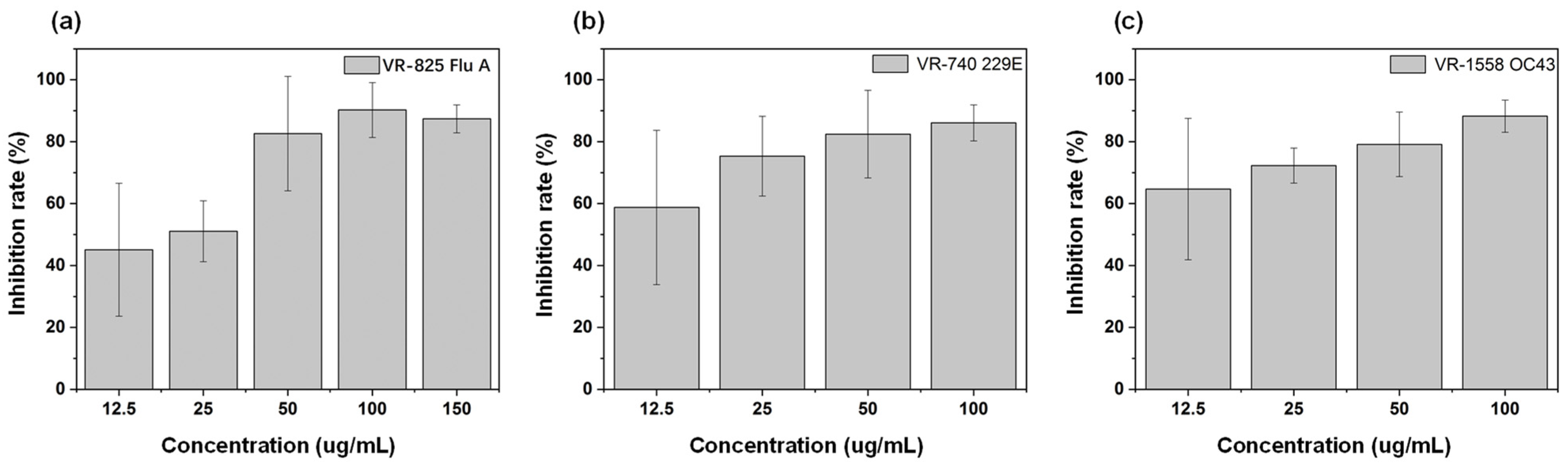
2.6. Inhibitory Effect on Influenza A Virus, α and β Coronaviruses
3. Discussion
4. Materials and Methods
4.1. Synthesis of Nanoparticles
4.1.1. Synthesis of Bi2S3 Nanoparticles
4.1.2. Synthesis of Ag2S Nanoparticles
4.1.3. Synthesis of AgBiS2 Nanoparticles
4.2. Characterization of Nanoparticles
4.3. Cultivation of Cells and Viruses
4.4. Cytotoxicity Test
4.5. TCID50 Assay for Determination of Virus Titer
4.6. Virus Inhibition Test
4.6.1. Direct Inactivating Experiment
4.6.2. Co-Cultivation Inhibition Experiment
4.6.3. Nanoparticle Preculture Inhibition Experiment
4.6.4. Virus Post-Infection Inhibition Experiment
4.7. Real-Time Fluorescent Quantitative PCR (RT-qPCR) Experiment
4.8. Indirect Immunofluorescence (IFA) Assay
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Toledo-Rueda, W.; Rosas-Murrieta, N.H.; Munoz-Medina, J.E.; Gonzalez-Bonilla, C.R.; Reyes-Leyva, J.; Santos-Lopez, G. Antiviral resistance markers in influenza virus sequences in Mexico, 2000–2017. Infect. Drug Resist. 2018, 11, 1751–1756. [Google Scholar] [CrossRef]
- Ng, L.Y.; Mohammad, A.W.; Leo, C.P.; Hilal, N. Polymeric membranes incorporated with metal/metal oxide nanoparticles: A comprehensive review. Desalination 2013, 308, 15–33. [Google Scholar] [CrossRef]
- Sang, C.; Ma, L.; Luo, D.; Liu, H.; Li, D.; Chen, T. Designing bioresponsive metal azolate framework-based nanosystem for efficient cancer therapy. Chem. Eng. J. 2019, 371, 301–305. [Google Scholar] [CrossRef]
- Delaviz, N.; Gill, P.; Ajami, A.; Aarabi, M. Aptamer-conjugated magnetic nanoparticles for the efficient removal of HCV particles from human plasma samples. RSC Adv. 2015, 5, 79433–79439. [Google Scholar] [CrossRef]
- Allahverdiyev, A.M.; Abamor, E.S.; Bagirova, M.; Rafailovich, M. Antimicrobial effects of TiO2 and Ag2O nanoparticles against drug-resistant bacteria and leishmania parasites. Future Microbiol. 2011, 6, 933–940. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, S.; Lauster, D.; Bardua, M.; Ludwig, K.; Angioletti-Uberti, S.; Popp, N.; Hoffmann, U.; Paulus, F.; Budt, M.; Stadtmuller, M.; et al. Linear polysialoside outperforms dendritic analogs for inhibition of influenza virus infection in vitro and in vivo. Biomaterials 2017, 138, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, Y.; Xu, K.; Qi, Y.; Zhong, J.; Zhang, K.; Li, J.; Wang, E.; Wu, Z.; Kang, Z. Broad-spectrum antiviral property of polyoxometalate localized on a cell surface. ACS Appl. Mater. Interfaces 2014, 6, 9785–9789. [Google Scholar] [CrossRef]
- Sun, H.; Li, H.; Harvey, I.; Sadler, P.J. Interactions of bismuth complexes with metallothionein(II). J. Biol. Chem. 1999, 274, 29094–29101. [Google Scholar] [CrossRef]
- Andrews, P.C.; Ferrero, R.L.; Forsyth, C.M.; Junk, P.C.; Maclellan, J.G.; Peiris, R.M. Bismuth(III) Saccharinate and Thiosaccharinate Complexes and the Effect of Ligand Substitution on Their Activity against Helicobacter pylori. Organometallics 2011, 30, 6283–6291. [Google Scholar] [CrossRef]
- Li, H.; Wang, R.; Sun, H. Systems Approaches for Unveiling the Mechanism of Action of Bismuth Drugs: New Medicinal Applications beyond Helicobacter pylori Infection. Acc. Chem. Res. 2019, 52, 216–227. [Google Scholar] [CrossRef]
- Yang, N.; Tanner, J.A.; Zheng, B.J.; Watt, R.M.; He, M.L.; Lu, L.Y.; Jiang, J.Q.; Shum, K.T.; Lin, Y.P.; Wong, K.L.; et al. Bismuth complexes inhibit the SARS coronavirus. Angew. Chem. Int. Ed. Engl. 2007, 46, 6464–6468. [Google Scholar] [CrossRef]
- Shu, T.; Huang, M.; Wu, D.; Ren, Y.; Zhang, X.; Han, Y.; Mu, J.; Wang, R.; Qiu, Y.; Zhang, D.Y.; et al. SARS-Coronavirus-2 Nsp13 Possesses NTPase and RNA Helicase Activities That Can Be Inhibited by Bismuth Salts. Virol. Sin. 2020, 35, 321–329. [Google Scholar] [CrossRef]
- Yuan, S.; Wang, R.; Chan, J.F.-W.; Zhang, A.J.; Cheng, T.; Chik, K.K.-H.; Ye, Z.-W.; Wang, S.; Lee, A.C.-Y.; Jin, L.; et al. Metallodrug ranitidine bismuth citrate suppresses SARS-CoV-2 replication and relieves virus-associated pneumonia in Syrian hamsters. Nat. Microbiol. 2020, 5, 1439–1448. [Google Scholar] [CrossRef]
- Park, S.; Park, H.H.; Kim, S.Y.; Kim, S.J.; Woo, K.; Ko, G. Antiviral Properties of Silver Nanoparticles on a Magnetic Hybrid Colloid. Appl. Environ. Microbiol. 2014, 80, 2343–2350. [Google Scholar] [CrossRef] [PubMed]
- Ilegbusi, O.J.; Trakhtenberg, L. Synthesis and Conductometric Property of Sol-Gel-Derived ZnO/PVP Nano Hybrid Films. J. Mater. Eng. Perform. 2012, 22, 911–915. [Google Scholar] [CrossRef]
- Kumar, M.; Devi, P.; Shivling, V.D. Thermal stability and electrochemical properties of PVP-protected Ru nanoparticles synthesized at room temperature. Mater. Res. Express 2017, 4, 085006. [Google Scholar] [CrossRef]
- Kumar, S.R.; Paulpandi, M.; ManivelRaja, M.; Mangalaraj, D.; Viswanathan, C.; Kannan, S.; Ponpandian, N. An in vitro analysis of H1N1 viral inhibition using polymer coated superparamagnetic Fe3O4 nanoparticles. RSC Adv. 2014, 4, 13409–13418. [Google Scholar] [CrossRef]
- Ghaffari, H.; Tavakoli, A.; Moradi, A.; Tabarraei, A.; Bokharaei-Salim, F.; Zahmatkeshan, M.; Farahmand, M.; Javanmard, D.; Kiani, S.J.; Esghaei, M.; et al. Inhibition of H1N1 influenza virus infection by zinc oxide nanoparticles: Another emerging application of nanomedicine. J. Biomed. Sci. 2019, 26, 70. [Google Scholar] [CrossRef] [PubMed]
- Du, T.; Liang, J.; Dong, N.; Lu, J.; Fu, Y.; Fang, L.; Xiao, S.; Han, H. Glutathione-Capped Ag2S Nanoclusters Inhibit Coronavirus Proliferation through Blockage of Viral RNA Synthesis and Budding. ACS Appl. Mater. Interfaces 2018, 10, 4369–4378. [Google Scholar] [CrossRef]
- Jeremiah, S.S.; Miyakawa, K.; Morita, T.; Yamaoka, Y.; Ryo, A. Potent antiviral effect of silver nanoparticles on SARS-CoV-2. Biochem. Biophys. Res. Commun. 2020, 533, 195–200. [Google Scholar] [CrossRef]
- Tavakoli, A.; Hashemzadeh, M.S. Inhibition of herpes simplex virus type 1 by copper oxide nanoparticles. J. Virol. Methods 2020, 275, 113688. [Google Scholar] [CrossRef]
- Kolahalam, L.A.; Kasi Viswanath, I.V.; Diwakar, B.S.; Govindh, B.; Reddy, V.; Murthy, Y.L.N. Review on nanomaterials: Synthesis and applications. Mater. Today Proc. 2019, 18, 2182–2190. [Google Scholar] [CrossRef]
- Joshi, V.A.; Joshi, G.M.; Shukla, S.K. Synthesis, properties and applications of nanomaterials: A mini review. In Functionalized Nanomaterials Based Devices for Environmental Applications; Hussain, C.M., Shukla, S.K., Joshi, G.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 61–75. [Google Scholar]
- Bordiwala, R.V. Green synthesis and Applications of Metal Nanoparticles—A Review Article. Results Chem. 2023, 5, 100832. [Google Scholar] [CrossRef]
- Baram-Pinto, D.; Shukla, S.; Perkas, N.; Gedanken, A.; Sarid, R. Inhibition of Herpes Simplex Virus Type 1 Infection by Silver Nanoparticles Capped with Mercaptoethane Sulfonate. Bioconjugate Chem. 2009, 20, 1497–1502. [Google Scholar] [CrossRef]
- Li, Z.; Hu, Y.; Miao, Z.; Xu, H.; Li, C.; Zhao, Y.; Li, Z.; Chang, M.; Ma, Z.; Sun, Y.; et al. Dual-Stimuli Responsive Bismuth Nanoraspberries for Multimodal Imaging and Combined Cancer Therapy. Nano Lett. 2018, 18, 6778–6788. [Google Scholar] [CrossRef]
- Cui, H.; Jiang, J.; Gu, W.; Sun, C.; Wu, D.; Yang, T.; Yang, G. Photocatalytic inactivation efficiency of anatase Nano-TiO2 Sol on the H9N2 avian influenza virus. Photochem. Photobiol. 2010, 86, 1135–1139. [Google Scholar] [CrossRef] [PubMed]
- Imai, K.; Ogawa, H.; Bui, V.N.; Inoue, H.; Fukuda, J.; Ohba, M.; Yamamoto, Y.; Nakamura, K. Inactivation of high and low pathogenic avian influenza virus H5 subtypes by copper ions incorporated in zeolite-textile materials. Antivir. Res. 2012, 93, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Nayak, M.; Sahoo, G.C.; Pandey, K.; Sarkar, M.C.; Ansari, Y.; Das, V.N.R.; Topno, R.K.; Bhawna; Madhukar, M.; et al. Iron oxide nanoparticles based antiviral activity of H1N1 influenza A virus. J. Infect. Chemother. 2019, 25, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Abo-Zeid, Y.; Ismail, N.S.M.; McLean, G.R.; Hamdy, N.M. A molecular docking study repurposes FDA approved iron oxide nanoparticles to treat and control COVID-19 infection. Eur. J. Pharm. Sci. 2020, 153, 105465. [Google Scholar] [CrossRef] [PubMed]
- Vonnemann, J.; Sieben, C.; Wolff, C.; Ludwig, K.; Böttcher, C.; Herrmann, A.; Haag, R. Virus inhibition induced by polyvalent nanoparticles of different sizes. Nanoscale 2014, 6, 2353. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yue, L.; Yang, Z.; Miao, Y.; Ouyang, R.; Hu, Y. Metal-Based Nanomaterials: Work as Drugs and Carriers against Viral Infections. Nanomaterials 2021, 11, 2129. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Gu, Z.; Yan, L.; Yong, Y.; Yi, X.; Zhang, X.; Liu, J.; Wu, R.; Ge, C.; Chen, C.; et al. Poly(Vinylpyrollidone)- and Selenocysteine-Modified Bi2Se3 Nanoparticles Enhance Radiotherapy Efficacy in Tumors and Promote Radioprotection in Normal Tissues. Adv. Mater. 2017, 29, 1701268. [Google Scholar] [CrossRef] [PubMed]
- Tavakoli, A.; Ataei-Pirkooh, A.; Mm Sadeghi, G.; Bokharaei-Salim, F.; Sahrapour, P.; Kiani, S.J.; Monavari, S.H. Polyethylene glycol-coated zinc oxide nanoparticle an efficient nanoweapon to fight against herpes simplex virus type 1. Nanomedicine 2018, 13, 2675–2690. [Google Scholar] [CrossRef] [PubMed]
- Baram-Pinto, D.; Shukla, S.; Gedanken, A.; Sarid, R. Inhibition of HSV-1 attachment, entry, and cell-to-cell spread by functionalized multivalent gold nanoparticles. Small 2010, 6, 1044–1050. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]













Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Yue, L.; Shen, B.; Yang, Z.; Shao, J.; Miao, Y.; Ouyang, R.; Hu, Y. Exploring the Inhibitory Effect of AgBiS2 Nanoparticles on Influenza Viruses. Int. J. Mol. Sci. 2023, 24, 10223. https://doi.org/10.3390/ijms241210223
Yang J, Yue L, Shen B, Yang Z, Shao J, Miao Y, Ouyang R, Hu Y. Exploring the Inhibitory Effect of AgBiS2 Nanoparticles on Influenza Viruses. International Journal of Molecular Sciences. 2023; 24(12):10223. https://doi.org/10.3390/ijms241210223
Chicago/Turabian StyleYang, Junlei, Lihuan Yue, Bei Shen, Zhu Yang, Jiang Shao, Yuqing Miao, Ruizhuo Ouyang, and Yihong Hu. 2023. "Exploring the Inhibitory Effect of AgBiS2 Nanoparticles on Influenza Viruses" International Journal of Molecular Sciences 24, no. 12: 10223. https://doi.org/10.3390/ijms241210223
APA StyleYang, J., Yue, L., Shen, B., Yang, Z., Shao, J., Miao, Y., Ouyang, R., & Hu, Y. (2023). Exploring the Inhibitory Effect of AgBiS2 Nanoparticles on Influenza Viruses. International Journal of Molecular Sciences, 24(12), 10223. https://doi.org/10.3390/ijms241210223