
Hexavalent Chromium Disrupts Oocyte Development in Rats by Elevating Oxidative Stress, DNA Double-Strand Breaks, Microtubule Disruption, and Aberrant Segregation of Chromosomes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
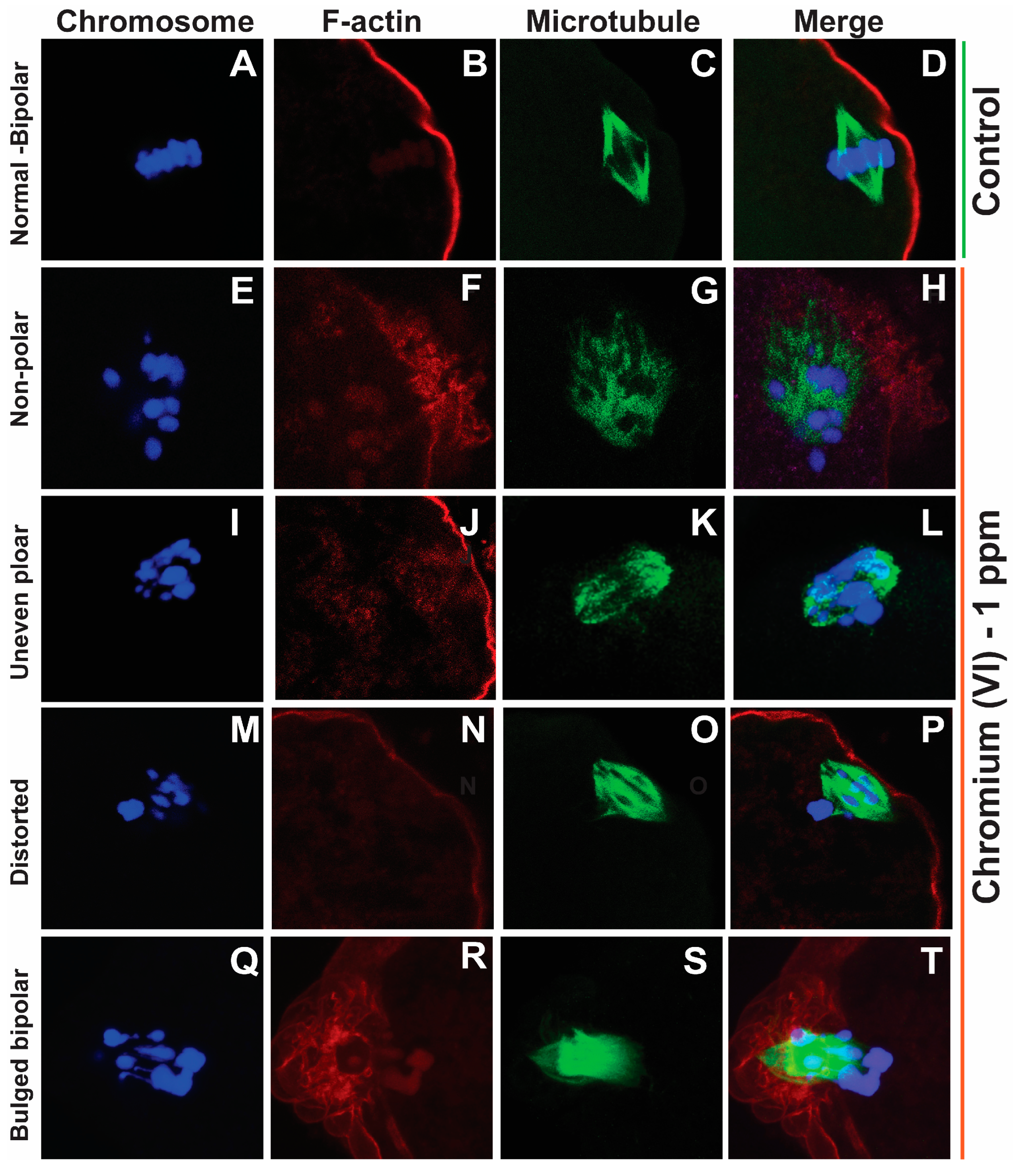
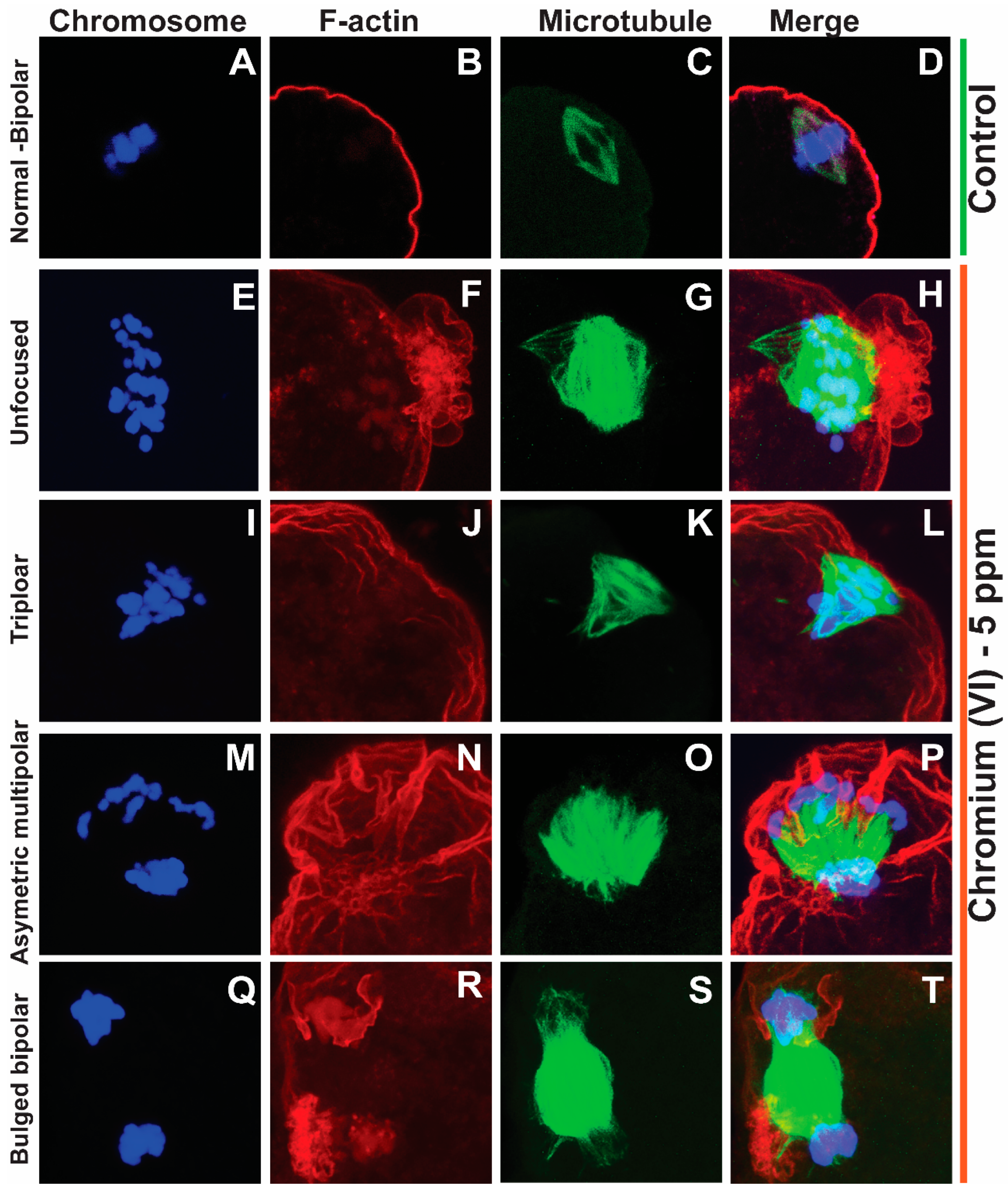
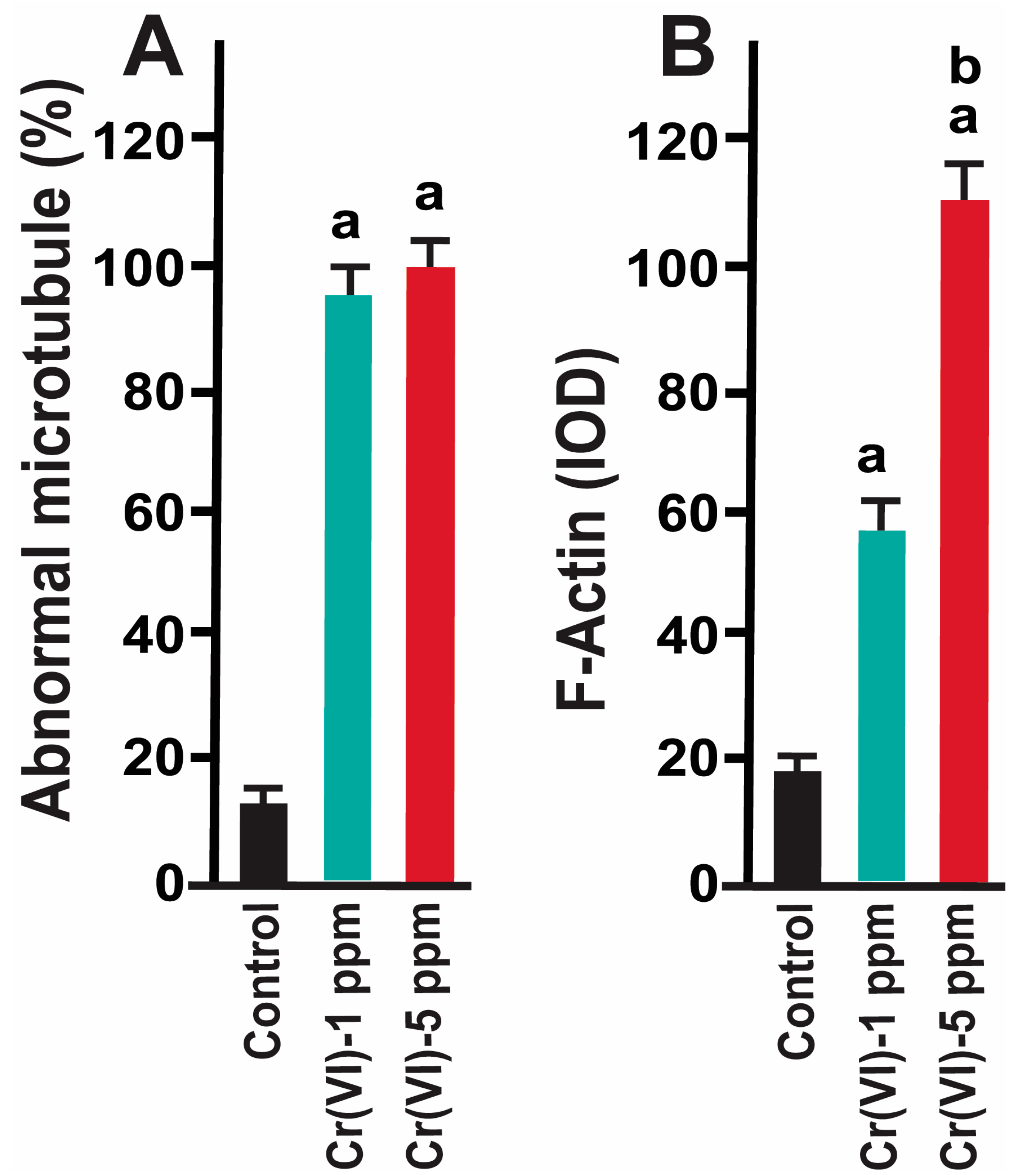
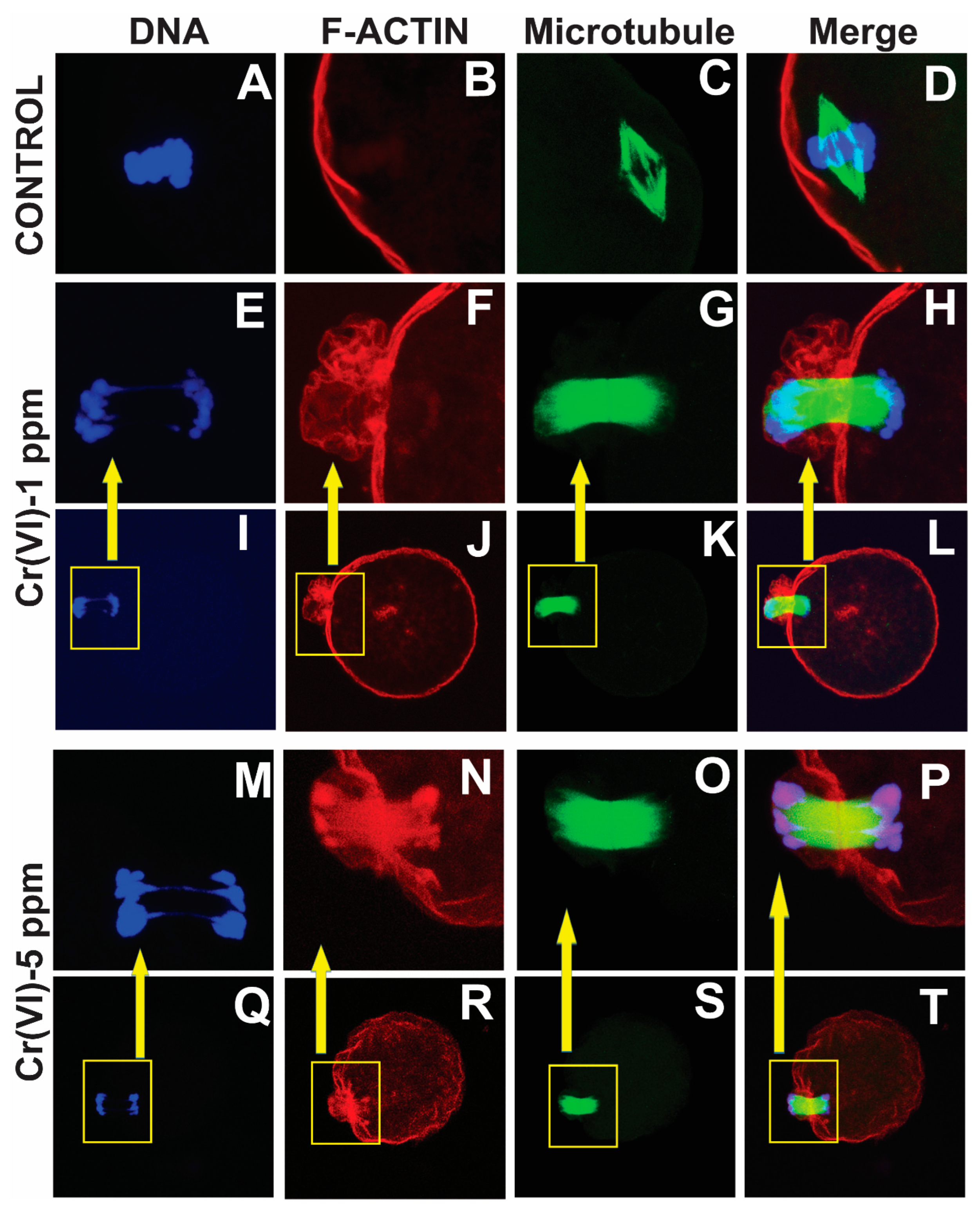
2.1. Exposure to Cr(VI) Distorted Microtubule Structure and Chromosome Arrangement in Metaphase II Oocytes
2.2. Exposure to Cr(VI) Delayed Polar Body Extrusion and Resulted in Incomplete Cytokinesis in MII Oocytes
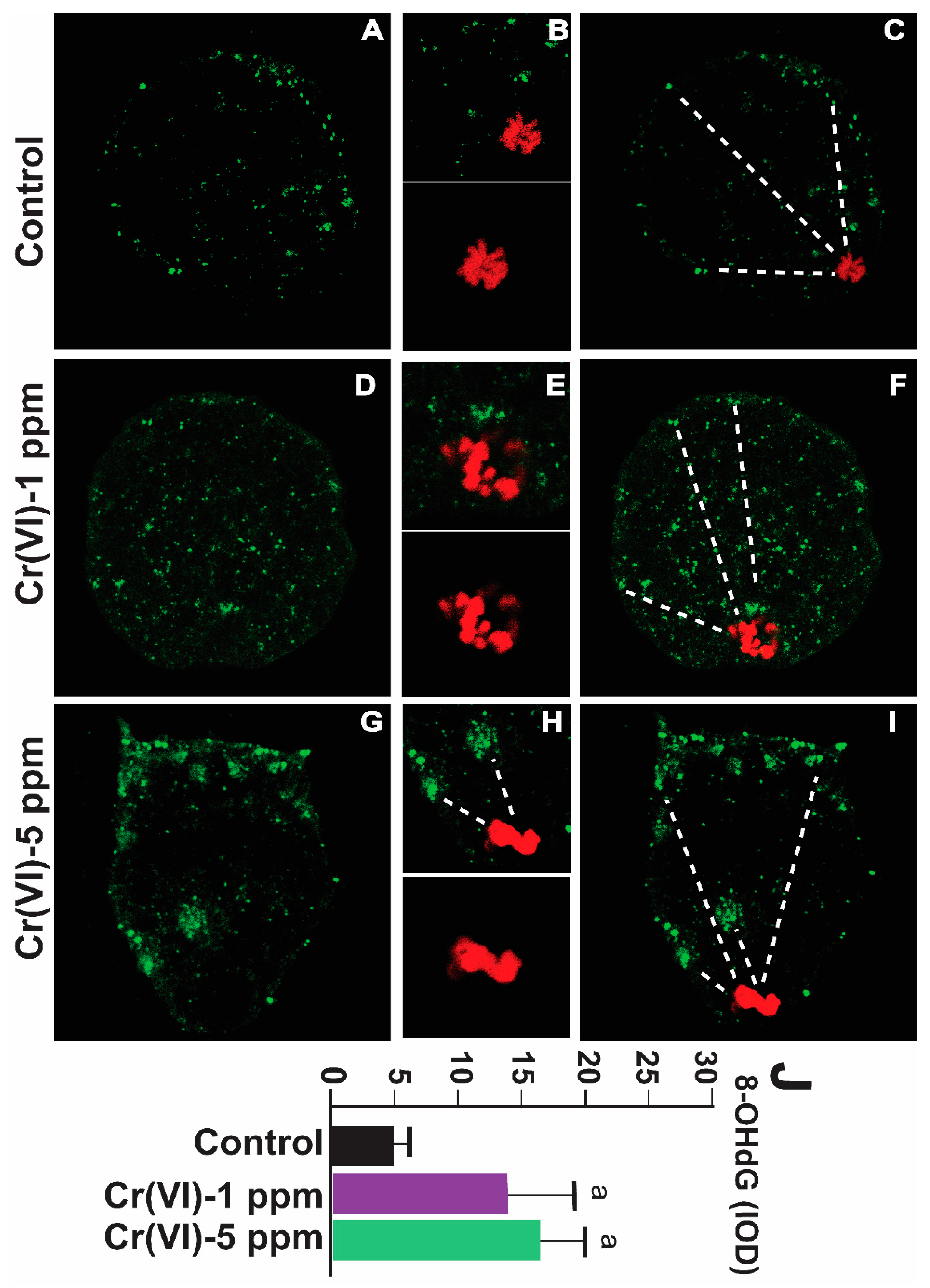
2.3. Exposure to Cr(VI) Increased Oxidative DNA Damage in the MII Oocytes
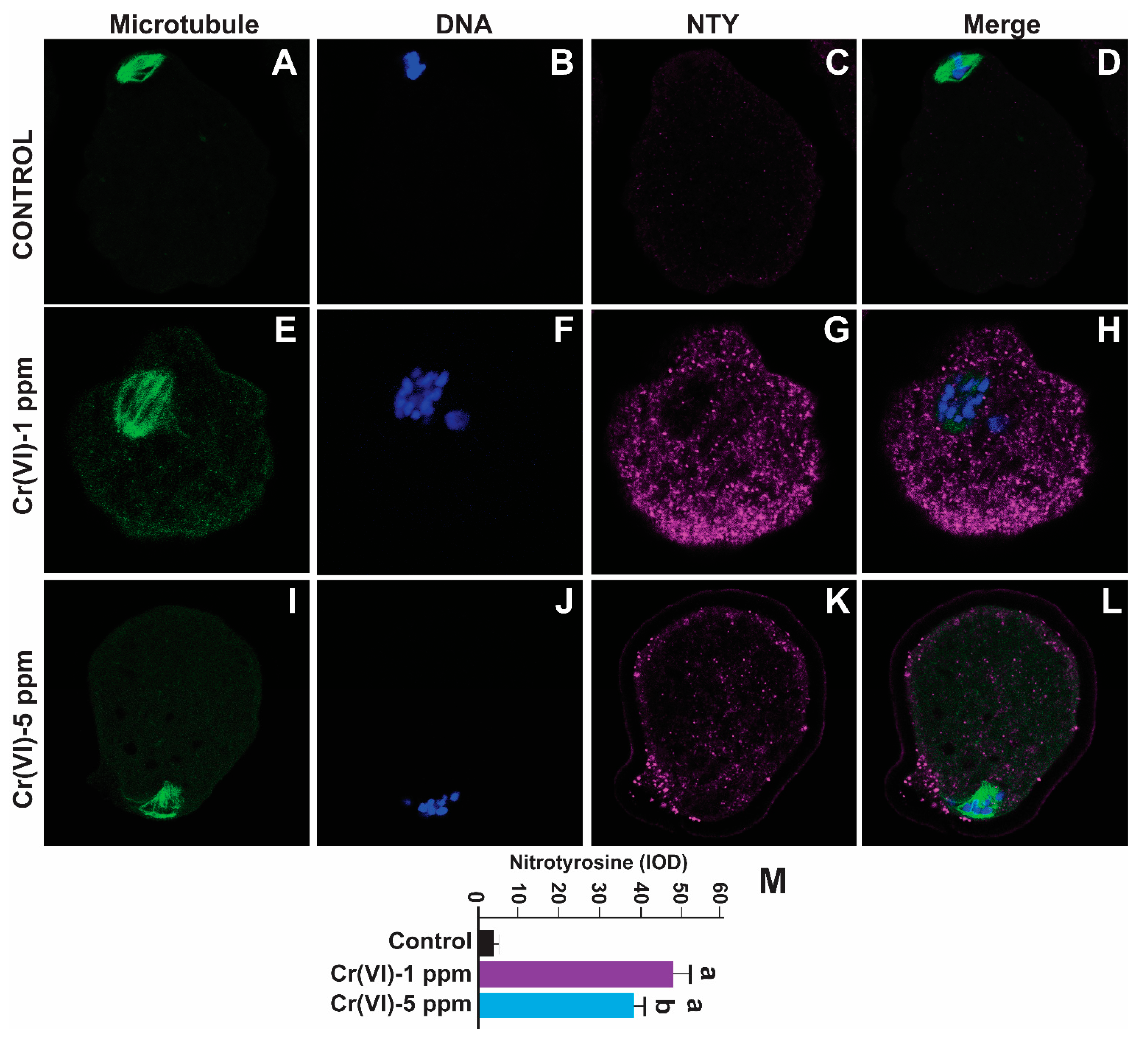
2.4. Exposure to Cr(VI) Increased Oxidative Protein Damage in the MII Oocytes
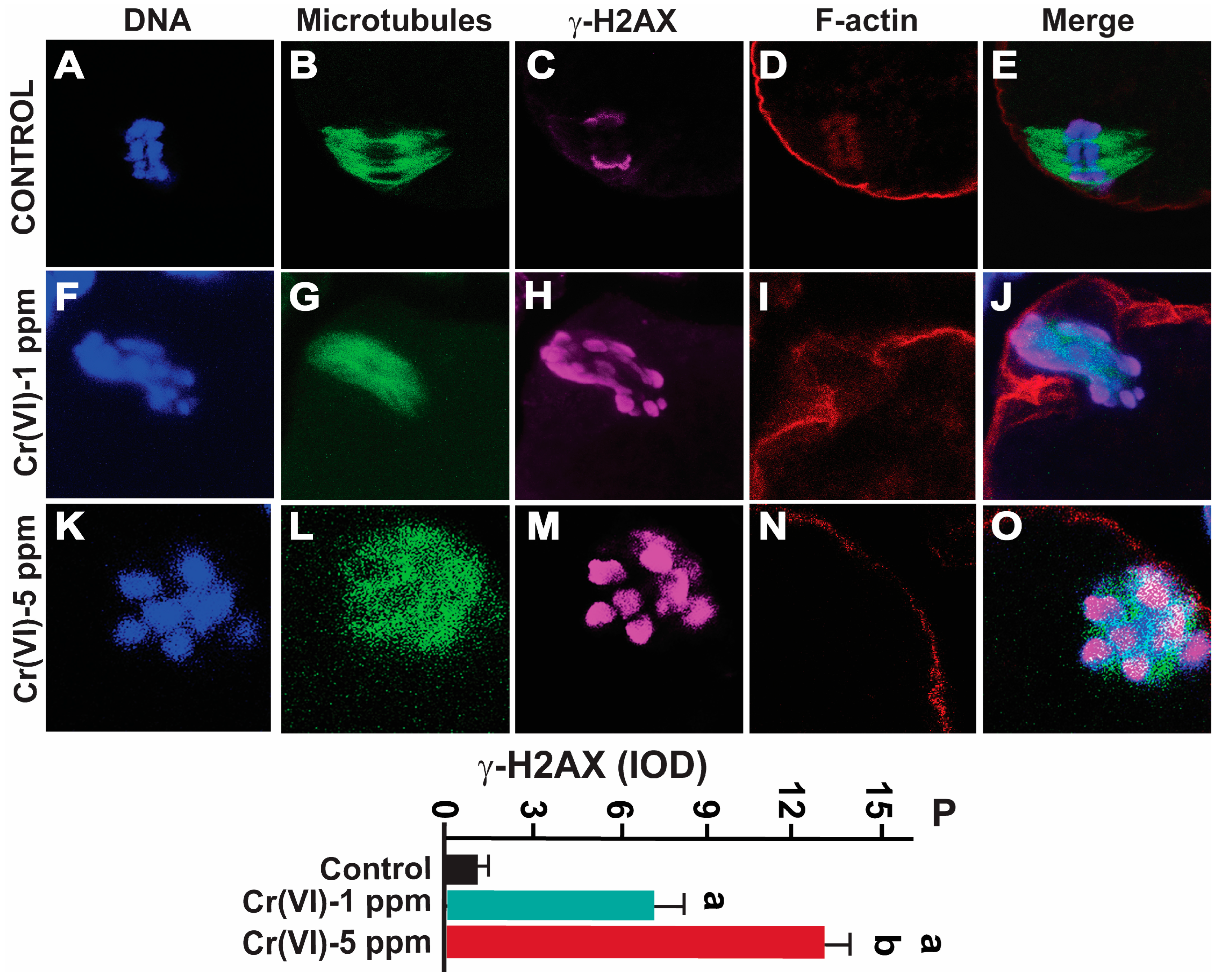
2.5. Exposure to Cr(VI) Increased DNA Double-Strand Breaks in MII Oocytes
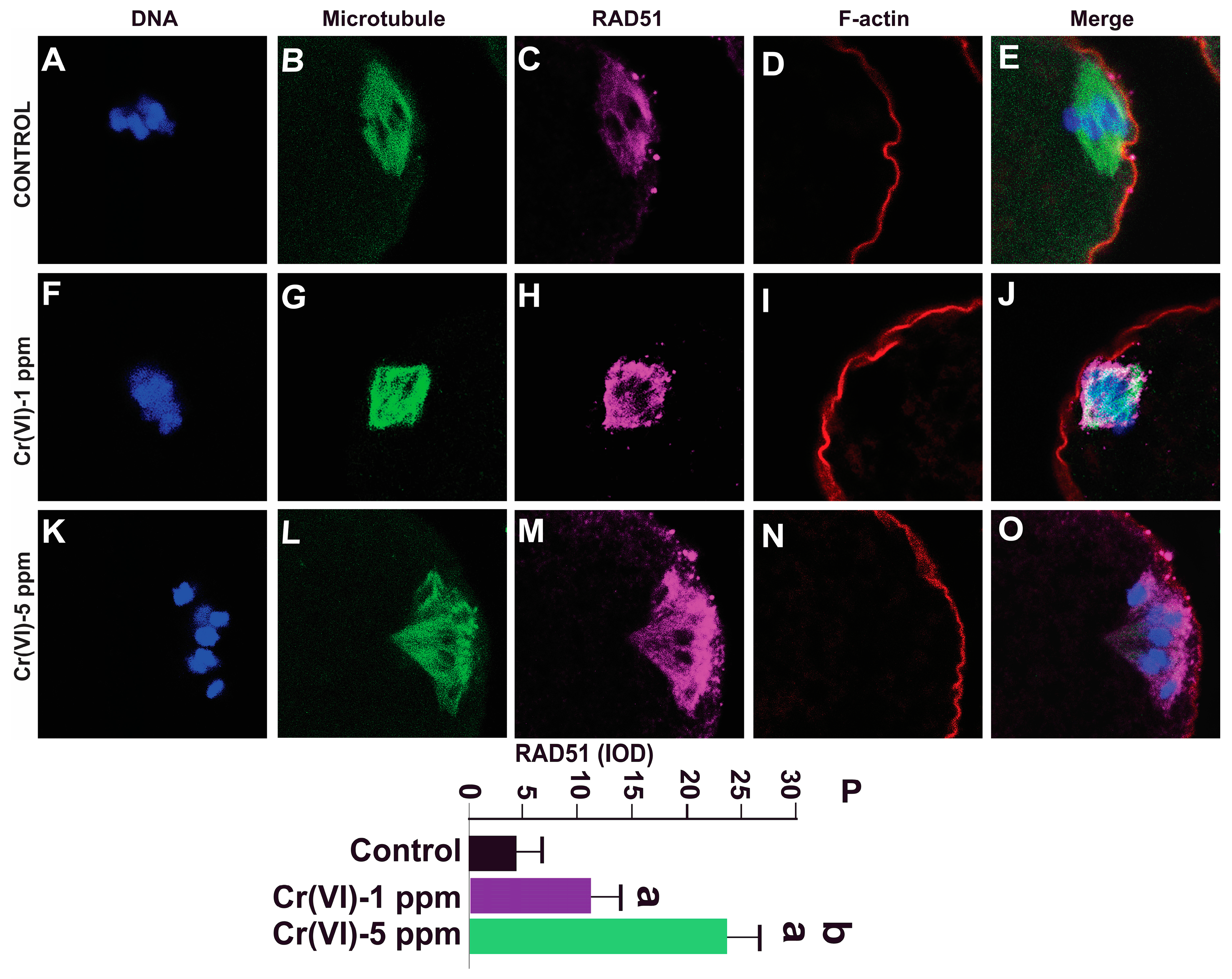
2.6. Exposure to Cr(VI) Increased RAD51 Expression in MII Oocytes
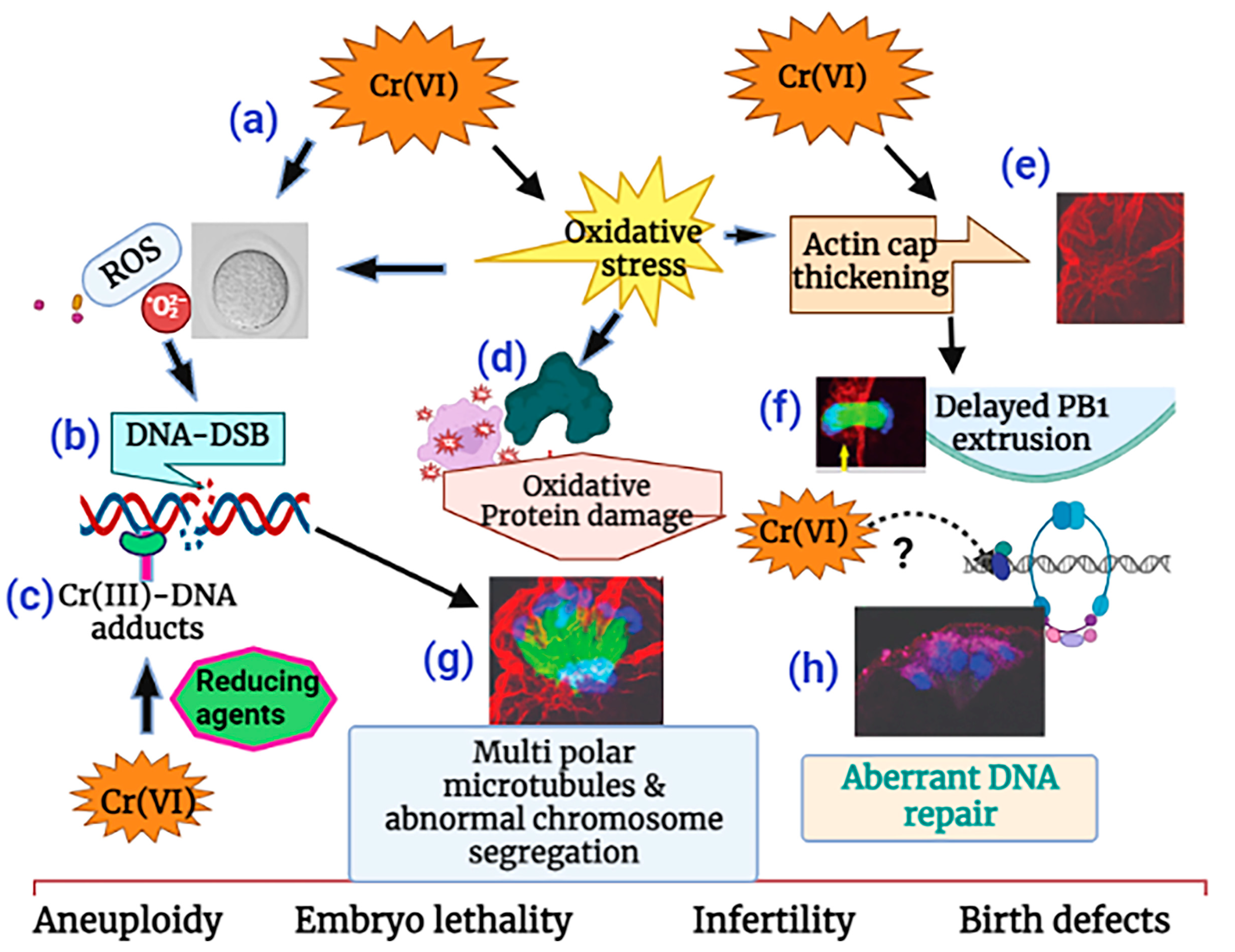
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animals and Treatments
4.3. Superovulation and Oocyte Collection
4.4. Immunocytochemistry of γ-H2AX, RAD 51, 8-OHdG, Nitrotyrosine, and Microtubules
4.5. Image Quantification
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Inhorn, M.C.; Patrizio, P. Infertility around the globe: New thinking on gender, reproductive technologies and global movements in the 21st century. Hum. Reprod. Update 2015, 21, 411–426. [Google Scholar] [CrossRef]
- Stringer, J.M.; Winship, A.; Liew, S.H.; Hutt, K. The capacity of oocytes for DNA repair. Cell. Mol. Life Sci. 2018, 75, 2777–2792. [Google Scholar] [CrossRef]
- Winship, A.L.; Stringer, J.M.; Liew, S.H.; Hutt, K.J. The importance of DNA repair for maintaining oocyte quality in response to anti-cancer treatments, environmental toxins and maternal ageing. Hum. Reprod. Update 2018, 24, 119–134. [Google Scholar] [CrossRef]
- Vessa, B.; Perlman, B.; McGovern, P.G.; Morelli, S.S. Endocrine disruptors and female fertility: A review of pesticide and plasticizer effects. F&S Rep. 2022, 3, 86–90. [Google Scholar] [CrossRef]
- Munisha, M.; Schimenti, J.C. Genome maintenance during embryogenesis. DNA Repair 2021, 106, 103195. [Google Scholar] [CrossRef] [PubMed]
- Robertson, A.B.; Klungland, A.; Rognes, T.; Leiros, I. DNA repair in mammalian cells: Base excision repair: The long and short of it. Cell. Mol. Life Sci. 2009, 66, 981–993. [Google Scholar] [CrossRef] [PubMed]
- Rothkamm, K.; Lobrich, M. Evidence for a lack of DNA double-strand break repair in human cells exposed to very low x-ray doses. Proc. Natl. Acad. Sci. USA 2003, 100, 5057–5062. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.W.; Xu, D.Q.; Feng, X.Z. The toxic effects and possible mechanisms of glyphosate on mouse oocytes. Chemosphere 2019, 237, 124435. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, Y.L.; Chen, M.H.; Zhang, Z.; Xu, B.H.; Liu, R.; Xu, L.; He, S.W.; Li, F.P.; Qi, Z.Q.; et al. Methoxychlor exposure induces oxidative stress and affects mouse oocyte meiotic maturation. Mol. Reprod. Dev. 2016, 83, 768–779. [Google Scholar] [CrossRef] [PubMed]
- Raghunath, A.; Sundarraj, K.; Nagarajan, R.; Arfuso, F.; Bian, J.; Kumar, A.P.; Sethi, G.; Perumal, E. Antioxidant response elements: Discovery, classes, regulation and potential applications. Redox Biol. 2018, 17, 297–314. [Google Scholar] [CrossRef]
- Song, Y.; Li, R. Effects of environment and lifestyle factors on anovulatory disorder. Environ. Female Reprod. Health 2021, 1300, 113–136. [Google Scholar]
- Dabrowska, N.; Wiczkowski, A. Analytics of oxidative stress markers in the early diagnosis of oxygen DNA damage. Adv. Clin. Exp. Med. 2017, 26, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Can oxidative DNA damage be used as a biomarker of cancer risk in humans? Problems, resolutions and preliminary results from nutritional supplementation studies. Free Radic. Res. 1998, 29, 469–486. [Google Scholar] [CrossRef]
- Lim, J.; Luderer, U. Oxidative damage increases and antioxidant gene expression decreases with aging in the mouse ovary. Biol. Reprod. 2011, 84, 775–782. [Google Scholar] [CrossRef]
- Wojsiat, J.; Korczyński, J.; Borowiecka, M.; Żbikowska, H.M. The role of oxidative stress in female infertility and in vitro fertilization. Postep. Hig. I Med. Dosw. 2017, 71, 359–366. [Google Scholar] [CrossRef]
- Al-Saleh, I.; Coskun, S.; Al-Rouqi, R.; Al-Rajudi, T.; Eltabache, C.; Abduljabbar, M.; Al-Hassan, S. Oxidative stress and DNA damage status in couples undergoing in vitro fertilization treatment. Reprod. Fertil. 2021, 2, 117–139. [Google Scholar] [CrossRef]
- Banu, S.K.; Stanley, J.A.; Sivakumar, K.K.; Arosh, J.A.; Burghardt, R.C. Resveratrol protects the ovary against chromium-toxicity by enhancing endogenous antioxidant enzymes and inhibiting metabolic clearance of estradiol. Toxicol. Appl. Pharmacol. 2016, 303, 65–78. [Google Scholar] [CrossRef]
- Hall, E.D.; Detloff, M.R.; Johnson, K.; Kupina, N.C. Peroxynitrite-mediated protein nitration and lipid peroxidation in a mouse model of traumatic brain injury. J. Neurotrauma 2004, 21, 9–20. [Google Scholar] [CrossRef]
- Headlam, H.A.; Davies, M.J. Markers of protein oxidation: Different oxidants give rise to variable yields of bound and released carbonyl products. Free. Radic. Biol. Med. 2004, 36, 1175–1184. [Google Scholar] [CrossRef]
- Banu, S.K.; Stanley, J.A.; Taylor, R.J.; Sivakumar, K.K.; Arosh, J.A.; Zeng, L.; Pennathur, S.; Padmanabhan, V. Sexually dimorphic impact of chromium accumulation on human placental oxidative stress and apoptosis. Toxicol. Sci. 2018, 161, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-H.; Zhang, X.; Wang, X.-C.; Jin, L.-F.; Yang, Z.-P.; Jiang, C.-X.; Chen, Q.; Ren, X.-B.; Cao, J.-Z.; Wang, Q. Chronic occupational exposure to hexavalent chromium causes DNA damage in electroplating workers. BMC Public Health 2011, 11, 224. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Itankar, N.; Patil, Y. Biomanagement of hexavalent chromium: Current trends and promising perspectives. J. Environ. Manag. 2021, 279, 111547. [Google Scholar] [CrossRef] [PubMed]
- Metzger, L. New Report: CPS Energy’s Calaveras Power Station among Nation’s Top Water Polluters; Environment Texas Research & Policy Center: St. Austin, TX, USA, 2022. [Google Scholar]
- Dubey, R.; Verma, P.; Kumar, S. Cr (III) genotoxicity and oxidative stress: An occupational health risk for leather tannery workers of South Asian developing countries. Toxicol. Ind. Health 2022, 38, 112–126. [Google Scholar] [CrossRef]
- Yatera, K.; Morimoto, Y.; Ueno, S.; Noguchi, S.; Kawaguchi, T.; Tanaka, F.; Suzuki, H.; Higashi, T. Cancer risks of hexavalent chromium in the respiratory tract. J. UOEH 2018, 40, 157–172. [Google Scholar] [CrossRef] [PubMed]
- Remy, L.L.; Byers, V.; Clay, T. Reproductive outcomes after non-occupational exposure to hexavalent chromium, Willits California, 1983–2014. Environ. Health 2017, 16, 18. [Google Scholar] [CrossRef]
- Wuri, L.; Arosh, J.A.; Wu, J.Z.; Banu, S.K. Exposure to hexavalent chromium causes infertility by disrupting cytoskeletal machinery and mitochondrial function of the metaphase II oocytes in superovulated rats. Toxicol. Rep. 2022, 9, 219–229. [Google Scholar] [CrossRef] [PubMed]
- OEHHA. Chromium-6 Drinking Water MCL. Available online: https://www.waterboards.ca.gov/drinking_water/certlic/drinkingwater/Chromium6.html (accessed on 18 April 2023).
- Malott, V. West County Road 112 ground water plume. EPA Reg. 2014, 6, 1–7. [Google Scholar]
- Roeles, J.; Tsiavaliaris, G. Actin-microtubule interplay coordinates spindle assembly in human oocytes. Nat. Commun. 2019, 10, 4651. [Google Scholar] [CrossRef]
- MacLennan, M.; Crichton, J.H.; Playfoot, C.J.; Adams, I.R. Oocyte development, meiosis and aneuploidy. Semin. Cell Dev. Biol. 2015, 45, 68–76. [Google Scholar] [CrossRef]
- Campbell, A.; Fishel, S.; Bowman, N.; Duffy, S.; Sedler, M.; Thornton, S. Retrospective analysis of outcomes after IVF using an aneuploidy risk model derived from time-lapse imaging without PGS. Reprod. BioMedicine Online 2013, 27, 140–146. [Google Scholar] [CrossRef]
- Cimini, D.; Degrassi, F. Aneuploidy: A matter of bad connections. Trends Cell Biol. 2005, 15, 442–451. [Google Scholar] [CrossRef]
- Chang, H.; Li, J.; Zhang, C.; Qian, W. Octocrylene exposure impairs mouse oocyte quality by inducing spindle defects and mitochondria dysfunction. Toxicology 2022, 479, 153306. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.-X.; Ding, Z.-M.; Ahmad, M.J.; Wang, Y.-S.; Duan, Z.-Q.; Miao, Y.-L.; Xiong, J.-J.; Huo, L.-J. Bisphenol B exposure disrupts mouse oocyte meiotic maturation in vitro through affecting spindle assembly and chromosome alignment. Front. Cell Dev. Biol. 2020, 8, 616771. [Google Scholar] [CrossRef] [PubMed]
- Gumus, E.; Sisko, A.; Abas, B.I.; Demirkan, B.; Cevik, O. Quercetin protects mouse oocytes against chromium-induced damage in vitro and in vivo. J. Trace Elem. Med. Biol. 2023, 75, 127087. [Google Scholar] [CrossRef] [PubMed]
- Henderson, A.L.; Colaiácovo, M.P. Exposure to phthalates: Germline dysfunction and aneuploidy. Prenat. Diagn. 2021, 41, 610–619. [Google Scholar] [CrossRef]
- Hunt, P.A.; Koehler, K.E.; Susiarjo, M.; Hodges, C.A.; Ilagan, A.; Voigt, R.C.; Thomas, S.; Thomas, B.F.; Hassold, T.J. Bisphenol A exposure causes meiotic aneuploidy in the female mouse. Curr. Biol. 2003, 13, 546–553. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-hydroxy-2′-deoxyguanosine (8-OHdG): A critical biomarker of oxidative stress and carcinogenesis. J. Environ. Sci. Health Part C 2009, 27, 120–139. [Google Scholar] [CrossRef]
- Kujjo, L.L.; Ronningen, R.; Ross, P.; Pereira, R.J.; Rodriguez, R.; Beyhan, Z.; Goissis, M.D.; Baumann, T.; Kagawa, W.; Camsari, C.; et al. RAD51 plays a crucial role in halting cell death program induced by ionizing radiation in bovine oocytes. Biol. Reprod. 2012, 86, 76. [Google Scholar] [CrossRef]
- Crain, D.A.; Janssen, S.J.; Edwards, T.M.; Heindel, J.; Ho, S.-m.; Hunt, P.; Iguchi, T.; Juul, A.; McLachlan, J.A.; Schwartz, J. Female reproductive disorders: The roles of endocrine-disrupting compounds and developmental timing. Fertil. Steril. 2008, 90, 911–940. [Google Scholar] [CrossRef]
- Guillette Jr, L.; Crain, D. Endocrine Disrupting Contaminants: An Evolutionary Perspective; Francis and Taylor Inc.: Philadelphia, PA, USA, 2000; p. 355. [Google Scholar]
- Bullach, A.; Trapphoff, T.; Zühlke, S.; Spiteller, M.; Dieterle, S. Impact of Nonylphenols and Polyhalogenated Compounds in Follicular Fluid on the Outcome of Intracytoplasmic Sperm Injection. Reprod. Sci. 2021, 28, 2118–2128. [Google Scholar] [CrossRef] [PubMed]
- Eichenlaub-Ritter, U.; Vogt, E.; Cukurcam, S.; Sun, F.; Pacchierotti, F.; Parry, J. Exposure of mouse oocytes to bisphenol A causes meiotic arrest but not aneuploidy. Mutat. Res. 2008, 651, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Grossman, D.; Kalo, D.; Gendelman, M.; Roth, Z. Effect of di-(2-ethylhexyl) phthalate and mono-(2-ethylhexyl) phthalate on in vitro developmental competence of bovine oocytes. Cell Biol. Toxicol. 2012, 28, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Sunderam, S.; Kissin, D.M.; Crawford, S.B.; Folger, S.G.; Jamieson, D.J.; Warner, L.; Barfield, W.D. Assisted reproductive technology surveillance—United States, 2012. Morb. Mortal. Wkly. Rep. Surveill. Summ. 2015, 64, 1–29. [Google Scholar]
- Gilchrist, R.B.; Lane, M.; Thompson, J.G. Oocyte-secreted factors: Regulators of cumulus cell function and oocyte quality. Hum. Reprod. Update 2008, 14, 159–177. [Google Scholar] [CrossRef]
- Nickens, K.P.; Patierno, S.R.; Ceryak, S. Chromium genotoxicity: A double-edged sword. Chem. Biol. Interact. 2010, 188, 276–288. [Google Scholar] [CrossRef]
- Banu, S.K.; Stanley, J.A.; Sivakumar, K.K.; Arosh, J.A.; Barhoumi, R.; Burghardt, R.C. Identifying a Novel Role for X-prolyl Aminopeptidase (Xpnpep)2 in CrVI-Induced Adverse Effects on Germ Cell Nest Breakdown and Follicle Development. Biol. Reprod. 2015, 92, 67. [Google Scholar] [CrossRef]
- Mogessie, B.; Schuh, M. Actin protects mammalian eggs against chromosome segregation errors. Science 2017, 357, eaal1647. [Google Scholar] [CrossRef]
- Maddox, A.S.; Azoury, J.; Dumont, J. Polar body cytokinesis. Cytoskeleton 2012, 69, 855–868. [Google Scholar] [CrossRef]
- Stanley, J.A.; Sivakumar, K.K.; Arosh, J.A.; Burghardt, R.C.; Banu, S.K. Edaravone mitigates hexavalent chromium-induced oxidative stress and depletion of antioxidant enzymes while estrogen restores antioxidant enzymes in the rat ovary in F1 offspring. Biol. Reprod. 2014, 91, 11–12. [Google Scholar] [CrossRef]
- Banu, S.K.; Stanley, J.A.; Sivakumar, K.K.; Arosh, J.A.; Taylor, R.J.; Burghardt, R.C. Chromium VI− Induced developmental toxicity of placenta is mediated through spatiotemporal dysregulation of cell survival and apoptotic proteins. Reprod. Toxicol. 2017, 68, 171–190. [Google Scholar] [CrossRef]
- Banu, S.K.; Samuel, J.B.; Arosh, J.A.; Burghardt, R.C.; Aruldhas, M.M. Lactational exposure to hexavalent chromium delays puberty by impairing ovarian development, steroidogenesis and pituitary hormone synthesis in developing Wistar rats. Toxicol. Appl. Pharmacol. 2008, 232, 180–189. [Google Scholar] [CrossRef]
- Elizur, S.E.; Lebovitz, O.; Orvieto, R.; Dor, J.; Zan-Bar, T. Reactive oxygen species in follicular fluid may serve as biochemical markers to determine ovarian aging and follicular metabolic age. Gynecol. Endocrinol. 2014, 30, 705–707. [Google Scholar] [CrossRef]
- Valko, M.; Morris, H.; Cronin, M. Metals, toxicity and oxidative stress. Curr. Med. Chem. 2005, 12, 1161–1208. [Google Scholar] [CrossRef]
- Carpenter, D.O. Effects of Persistent and Bioactive Organic Pollutants on Human Health; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Amadi, C.N.; Igweze, Z.N.; Orisakwe, O.E. Heavy metals in miscarriages and stillbirths in developing nations. Middle East Fertil. Soc. J. 2017, 22, 91–100. [Google Scholar] [CrossRef]
- Hessel, E.V.S.; Staal, Y.C.M.; Piersma, A.H.; den Braver-Sewradj, S.P.; Ezendam, J. Occupational exposure to hexavalent chromium. Part I. Hazard assessment of non-cancer health effects. Regul. Toxicol. Pharm. 2021, 126, 105048. [Google Scholar] [CrossRef]
- Stanley, J.A.; Lee, J.; Nithy, T.K.; Arosh, J.A.; Burghardt, R.C.; Banu, S.K. Chromium-VI arrests cell cycle and decreases granulosa cell proliferation by down-regulating cyclin-dependent kinases (CDK) and cyclins and up-regulating CDK-inhibitors. Reprod. Toxicol. 2011, 32, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Banu, S.K.; Stanley, J.A.; Lee, J.; Stephen, S.D.; Arosh, J.A.; Hoyer, P.B.; Burghardt, R.C. Hexavalent chromium-induced apoptosis of granulosa cells involves selective sub-cellular translocation of Bcl-2 members, ERK1/2 and p53. Toxicol. Appl. Pharmacol. 2011, 251, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Lord, T.; Aitken, R.J. Fertilization stimulates 8-hydroxy-2′-deoxyguanosine repair and antioxidant activity to prevent mutagenesis in the embryo. Dev. Biol. 2015, 406, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Goud, P.T.; Goud, A.P.; Joshi, N.; Puscheck, E.; Diamond, M.P.; Abu-Soud, H.M. Dynamics of nitric oxide, altered follicular microenvironment, and oocyte quality in women with endometriosis. Fertil. Steril. 2014, 102, 151–159. [Google Scholar] [CrossRef]
- Banerjee, J.; Shaeib, F.; Maitra, D.; Saed, G.M.; Dai, J.; Diamond, M.P.; Abu-Soud, H.M. Peroxynitrite affects the cumulus cell defense of metaphase II mouse oocytes leading to disruption of the spindle structure in vitro. Fertil. Steril. 2013, 100, 578–584.e571. [Google Scholar] [CrossRef]
- Goud, A.P.; Goud, P.T.; Diamond, M.P.; Gonik, B.; Abu-Soud, H.M. Reactive oxygen species and oocyte aging: Role of superoxide, hydrogen peroxide, and hypochlorous acid. Free Radic. Biol. Med. 2008, 44, 1295–1304. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Wise, S.S.; Wise, J.P., Sr. Deficient repair of particulate hexavalent chromium-induced DNA double strand breaks leads to neoplastic transformation. Mutat. Res. 2008, 649, 230–238. [Google Scholar] [CrossRef]
- Namgoong, S.; Kim, N.H. Meiotic spindle formation in mammalian oocytes: Implications for human infertility. Biol. Reprod. 2018, 98, 153–161. [Google Scholar] [CrossRef]
- Ghosh, A.; Tripathy, A.; Ghosh, D. Impact of endocrine disrupting chemicals (EDCs) on reproductive health of human. In Proceedings of the Zoological Society; Springer: Berlin/Heidelberg, Germany; pp. 16–30.
- La Marca, A.; Mastellari, E. Infertility: Epidemiology and etiology. Female Reprod. Dysfunct. 2020, 211–233. [Google Scholar] [CrossRef]
- Ganesan, S.; Keating, A.F. Bisphenol A-Induced Ovotoxicity Involves DNA Damage Induction to Which the Ovary Mounts a Protective Response Indicated by Increased Expression of Proteins Involved in DNA Repair and Xenobiotic Biotransformation. Toxicol. Sci. 2016, 152, 169–180. [Google Scholar] [CrossRef]
- Liu, K.H.; Sun, X.F.; Feng, Y.Z.; Cheng, S.F.; Li, B.; Li, Y.P.; Shen, W.; Li, L. The impact of Zearalenone on the meiotic progression and primordial follicle assembly during early oogenesis. Toxicol. Appl. Pharmacol. 2017, 329, 9–17. [Google Scholar] [CrossRef] [PubMed]
- California Environmental Protection Agency. Public Health Goal for Hexavalent Chromium (CrVI) in Drinking Water; California Environmental Protection Agency: Sacramento, CA, USA, 2011.
- Sivakumar, K.K.; Stanley, J.A.; Arosh, J.A.; Pepling, M.E.; Burghardt, R.C.; Banu, S.K. Prenatal exposure to chromium induces early reproductive senescence by increasing germ cell apoptosis and advancing germ cell cyst breakdown in the F1 offspring. Dev. Biol. 2014, 388, 22–34. [Google Scholar] [CrossRef]
- Lee, J.; McCracken, J.A.; Banu, S.K.; Arosh, J.A. Intrauterine Inhibition of Prostaglandin Transporter Protein Blocks Release of Luteolytic PGF2alpha Pulses Without Suppressing Endometrial Expression of Estradiol or Oxytocin Receptor in Ruminants. Biol. Reprod. 2013, 89, 21–29. [Google Scholar] [CrossRef]
- Wang, C.-J.; Zhou, Z.-G.; Holmqvist, A.; Zhang, H.; Li, Y.; Adell, G.; Sun, X.-F. Survivin expression quantified by Image Pro-Plus compared with visual assessment. Appl. Immunohistochem. Mol. Morphol. 2009, 17, 530–535. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wuri, L.; Burghardt, R.C.; Arosh, J.A.; Long, C.R.; Banu, S.K. Hexavalent Chromium Disrupts Oocyte Development in Rats by Elevating Oxidative Stress, DNA Double-Strand Breaks, Microtubule Disruption, and Aberrant Segregation of Chromosomes. Int. J. Mol. Sci. 2023, 24, 10003. https://doi.org/10.3390/ijms241210003
Wuri L, Burghardt RC, Arosh JA, Long CR, Banu SK. Hexavalent Chromium Disrupts Oocyte Development in Rats by Elevating Oxidative Stress, DNA Double-Strand Breaks, Microtubule Disruption, and Aberrant Segregation of Chromosomes. International Journal of Molecular Sciences. 2023; 24(12):10003. https://doi.org/10.3390/ijms241210003
Chicago/Turabian StyleWuri, Liga, Robert C. Burghardt, Joe A. Arosh, Charles R. Long, and Sakhila K. Banu. 2023. "Hexavalent Chromium Disrupts Oocyte Development in Rats by Elevating Oxidative Stress, DNA Double-Strand Breaks, Microtubule Disruption, and Aberrant Segregation of Chromosomes" International Journal of Molecular Sciences 24, no. 12: 10003. https://doi.org/10.3390/ijms241210003
APA StyleWuri, L., Burghardt, R. C., Arosh, J. A., Long, C. R., & Banu, S. K. (2023). Hexavalent Chromium Disrupts Oocyte Development in Rats by Elevating Oxidative Stress, DNA Double-Strand Breaks, Microtubule Disruption, and Aberrant Segregation of Chromosomes. International Journal of Molecular Sciences, 24(12), 10003. https://doi.org/10.3390/ijms241210003