
Hypoxia with or without Treadmill Exercises Affects Slow-Twitch Muscle Atrophy and Joint Destruction in a Rat Model of Rheumatoid Arthritis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Changes in Body Weight and Paw Volume over Time
2.2. The Effects of Treadmill Running in a Hypoxic Environment on Articular Cartilages
2.3. The Effects of Treadmill Running in a Hypoxic Environment on Skeletal Muscles
2.3.1. Muscle Weight on Day 44
2.3.2. Histological Evaluation of the Soleus Muscle on Day 44
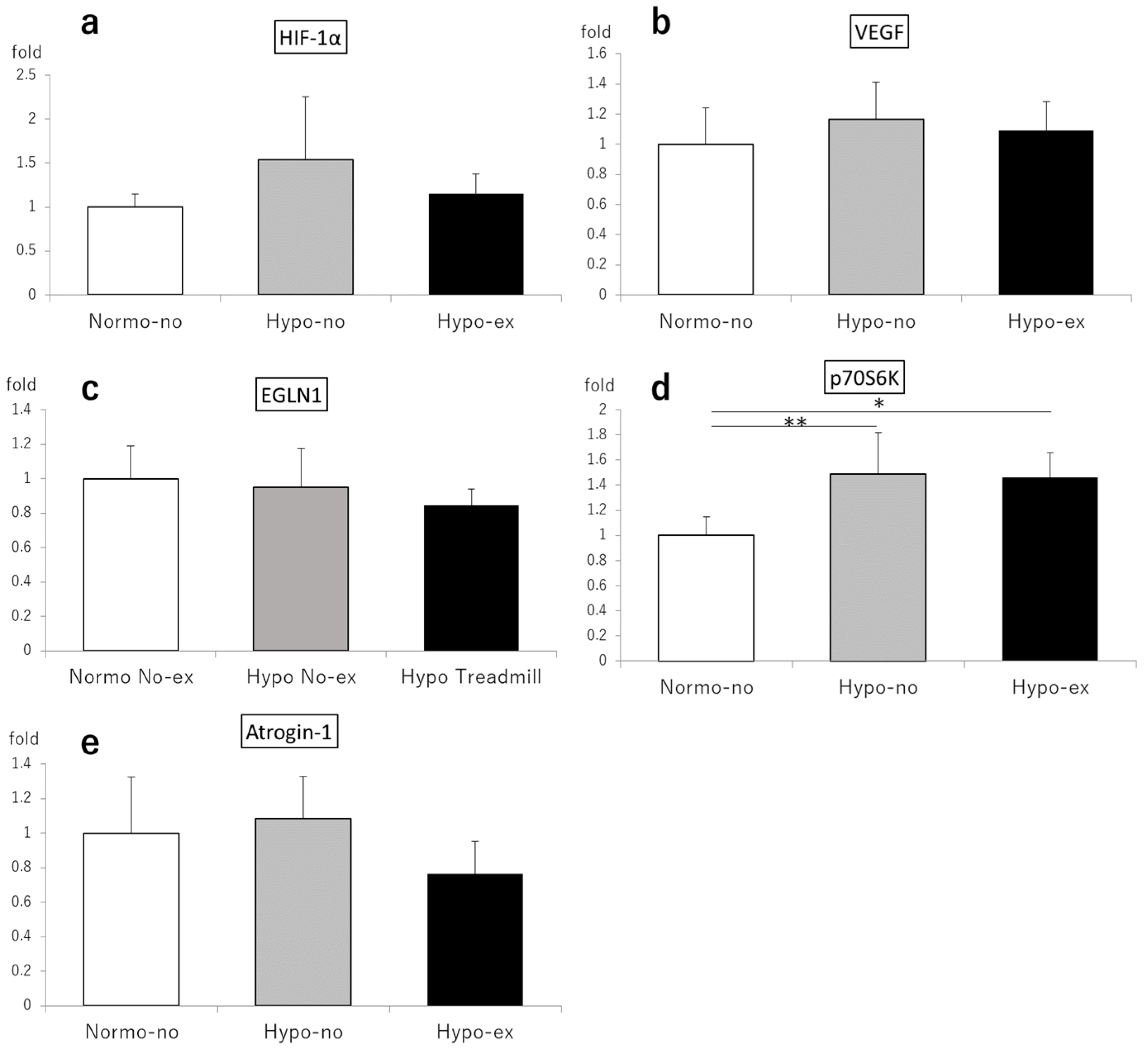
2.3.3. Gene Expression on Days 2 and 44
3. Discussion
4. Materials and Methods
4.1. Collagen-Induced Arthritis Model
4.2. Body Weight, Paw Volume, and Clinical Score
4.3. Evaluation of the EDL and Soleus Muscle Weights
4.4. Rearing in the Hypoxic Chamber and Treadmill Running
4.5. Treadmill Running Protocol
4.6. Histochemical Analysis (Articular Cartilage)
4.7. Muscle Preparation
4.8. Muscle Histochemistry
4.9. Real-Time Polymerase Chain Reaction (PCR)
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aletaha, D.; Neogi, T.; Silman, A.J.; Funovits, J.; Felson, D.T.; Bingham, C.O., 3rd; Birnbaum, N.S.; Burmester, G.R.; Bykerk, V.P.; Cohen, M.D. 2010 Rheumatoid arthritis classification criteria: An American College of Rheumatology/European League against Rheumatism collaborative initiative. Arthritis Rheum. 2010, 62, 2569–2581. [Google Scholar] [CrossRef] [PubMed]
- McInnes, I.B.; Schett, G. The pathogenesis of rheumatoid arthritis. N. Engl. J. Med. 2011, 365, 2205–2219. [Google Scholar] [CrossRef]
- Tekgoz, E.; Colak, S.; Ozalp Ates, F.S.; Sonaeren, I.; Yilmaz, S.; Cinar, M. Sarcopenia in rheumatoid arthritis: Is it a common manifestation? Int. J. Rheum. Dis. 2020, 23, 1685–1691. [Google Scholar] [CrossRef] [PubMed]
- Kamada, Y.; Toyama, S.; Arai, Y.; Inoue, H.; Nakagawa, S.; Fujii, Y.; Kaihara, K.; Kishida, T.; Mazda, O.; Takahashi, K. Treadmill running prevents atrophy differently in fast-versus slow-twitch muscles in a rat model of rheumatoid arthritis. J. Muscle Res. Cell Motil. 2021, 42, 429–441. [Google Scholar] [CrossRef]
- Maini, R.; St Clair, E.W.; Breedveld, F.; Furst, D.; Kalden, J.; Weisman, M.; Smolen, J.; Emery, P.; Harriman, G.; Feldmann, M.; et al. Infliximab (chimeric anti-tumour necrosis factor alpha monoclonal antibody) versus placebo in rheumatoid arthritis patients receiving concomitant methotrexate: A randomised phase III trial. ATTRACT Study Group. Lancet 1999, 354, 1932–1939. [Google Scholar] [CrossRef]
- Lipsky, P.E.; van der Heijde, D.M.; St Clair, E.W.; Furst, D.E.; Breedveld, F.C.; Kalden, J.R.; Smolen, J.S.; Weisman, M.; Emery, P.; Feldmann, M.; et al. Infliximab and methotrexate in the treatment of rheumatoid arthritis. Anti-tumor Necrosis Factor Trial in Rheumatoid Arthritis with Concomitant Therapy Study Group. N. Engl. J. Med. 2000, 343, 1594–1602. [Google Scholar] [CrossRef] [PubMed]
- Smolen, J.S.; Landewé, R.B.M.; Bijlsma, J.W.J.; Burmester, G.R.; Dougados, M.; Kerschbaumer, A.; McInnes, I.B.; Sepriano, A.; van Vollenhoven, R.F.; de Wit, M.; et al. EULAR recommendations for the management of rheumatoid arthritis with synthetic and biological disease-modifying antirheumatic drugs: 2019 update. Ann. Rheum. Dis. 2020, 79, 685–699. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.A.; Wells, G.A.; Christensen, R.; Ghogomu, E.T.; Maxwell, L.; Macdonald, J.K.; Filippini, G.; Skoetz, N.; Francis, D.; Lopes, L.C.; et al. Adverse effects of biologics: A network meta-analysis and Cochrane overview. Cochrane Database Syst. Rev. 2011, 2011, CD008794. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.F.; Liu, J.R.; Chang, S.H.; Kuo, C.F.; See, L.C. Trends of adverse events and mortality after DMARDs in patients with rheumatoid arthritis: Interrupted time-series analysis. Immun. Inflamm. Dis. 2022, 10, e630. [Google Scholar] [CrossRef]
- Hurkmans, E.; van der Giesen, F.J.; Vliet Vlieland, T.P.; Schoones, J.; Van den Ende, E.C. Dynamic exercise programs (aerobic capacity and/or muscle strength training) in patients with rheumatoid arthritis. Cochrane Database Syst. Rev. 2009, 4, CD006853. [Google Scholar] [CrossRef]
- Williams, M.A.; Srikesavan, C.; Heine, P.J.; Bruce, J.; Brosseau, L.; Hoxey-Thomas, N.; Lamb, S.E. Exercise for rheumatoid arthritis of the hand. Cochrane Database Syst. Rev. 2018, 7, CD003832. [Google Scholar] [CrossRef] [PubMed]
- Baillet, A.; Vaillant, M.; Guinot, M.; Juvin, R.; Gaudin, P. Efficacy of resistance exercises in rheumatoid arthritis: Meta-analysis of randomized controlled trials. Rheumatology 2012, 51, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Metsios, G.S.; Kitas, G.D. Physical activity, exercise and rheumatoid arthritis: Effectiveness, mechanisms and implementation. Best Pract. Res. Clin. Rheumatol. 2018, 32, 669–682. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Wang, S.; Zhang, J.; Xu, T. Effects of body weight-supported treadmill training on cartilage-subchondral bone unit in the rat model of posttraumatic osteoarthritis. J. Orthop. Res. 2021, 39, 1227–1235. [Google Scholar] [CrossRef]
- Shimomura, S.; Inoue, H.; Arai, Y.; Nakagawa, S.; Fujii, Y.; Kishida, T.; Ichimaru, S.; Tsuchida, S.; Shirai, T.; Ikoma, K.; et al. Treadmill running ameliorates destruction of articular cartilage and subchondral bone, not only synovitis, in a rheumatoid arthritis rat model. Int. J. Mol. Sci. 2018, 19, 1653. [Google Scholar] [CrossRef]
- Zhang, F.J.; Luo, W.; Lei, G.H. Role of HIF-1α and HIF-2α in osteoarthritis. Jt. Bone Spine 2015, 82, 144–147. [Google Scholar] [CrossRef]
- Shimomura, S.; Inoue, H.; Arai, Y.; Nakagawa, S.; Fujii, Y.; Kishida, T.; Shin-Ya, M.; Ichimaru, S.; Tsuchida, S.; Mazda, O.; et al. Mechanical stimulation of chondrocytes regulates HIF-1α under hypoxic conditions. Tissue Cell 2021, 71, 101574. [Google Scholar] [CrossRef]
- Cirillo, F.; Resmini, G.; Ghiroldi, A.; Piccoli, M.; Bergante, S.; Tettamanti, G.; Anastasia, L. Activation of the hypoxia-inducible factor 1α promotes myogenesis through the noncanonical Wnt pathway, leading to hypertrophic myotubes. FASEB J. 2017, 31, 2146–2156. [Google Scholar] [CrossRef]
- Kaihara, K.; Nakagawa, S.; Arai, Y.; Inoue, H.; Tsuchida, S.; Fujii, Y.; Kamada, Y.; Kishida, T.; Mazda, O.; Takahashi, K. Sustained hypoxia suppresses joint destruction in a rat model of rheumatoid arthritis via negative feedback of hypoxia inducible factor-1α. Int. J. Mol. Sci. 2021, 22, 3898. [Google Scholar] [CrossRef]
- Lund-Olesen, K. Oxygen tension in synovial fluids. Arthritis Rheum. 1970, 13, 769–776. [Google Scholar] [CrossRef]
- Hu, L.; Liu, R.; Zhang, L. Advance in bone destruction participated by JAK/STAT in rheumatoid arthritis and therapeutic effect of JAK/STAT inhibitors. Int. Immunopharmacol. 2022, 111, 109095. [Google Scholar] [CrossRef]
- Bartalucci, A.; Ferrucci, M.; Fulceri, F.; Lazzeri, G.; Lenzi, P.; Toti, L.; Serpiello, F.R.; La Torre, A.; Gesi, M. High-intensity exercise training produces morphological and biochemical changes in adrenal gland of mice. Histol. Histopathol. 2012, 27, 753–769. [Google Scholar] [PubMed]
- Toti, L.; Bartalucci, A.; Ferrucci, M.; Fulceri, F.; Lazzeri, G.; Lenzi, P.; Soldani, P.; Gobbi, P.; La Torre, A.; Gesi, M. High-intensity exercise training induces morphological and biochemical changes in skeletal muscles. Biol. Sport 2013, 30, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.X.; Zhan, Y.X.; You, H.N.; You, Y.Y.; Li, W.Y.; Jiang, S.S.; Zheng, M.F.; Zhang, L.Z.; Chen, K.; Chen, Q.X. Effects of lower body positive pressure treadmill on functional improvement in knee osteoarthritis: A randomized clinical trial study. World J. Clin. Cases 2021, 9, 10604–10615. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Cao, H.; Wang, M.; Zou, J.; Wu, W. Moderate-intensity treadmill running relieves motion-induced post-traumatic osteoarthritis mice by up-regulating the expression of lncRNA H19. Biomed. Eng. Online 2021, 20, 111. [Google Scholar] [CrossRef]
- González-Chávez, S.A.; López-Loeza, S.M.; Acosta-Jiménez, S.; Cuevas-Martínez, R.; Pacheco-Silva, C.; Chaparro-Barrera, E.; Pacheco-Tena, C. Low-intensity physical exercise decreases inflammation and joint damage in the preclinical phase of a rheumatoid arthritis murine model. Biomolecules 2023, 13, 488. [Google Scholar] [CrossRef]
- Takarada, Y.; Nakamura, Y.; Aruga, S.; Onda, T.; Miyazaki, S.; Ishii, N. Rapid increase in plasma growth hormone after low-intensity resistance exercise with vascular occlusion. J. Appl. Physiol. 2000, 88, 61–65. [Google Scholar] [CrossRef]
- Kawada, S.; Ishii, N. Skeletal muscle hypertrophy after chronic restriction of venous blood flow in rats. Med. Sci. Sports Exerc. 2005, 37, 1144–1150. [Google Scholar] [CrossRef]
- Nakajima, T.; Yasuda, T.; Koide, S.; Yamasoba, T.; Obi, S.; Toyoda, S.; Sato, Y.; Inoue, T.; Kano, Y. Repetitive restriction of muscle blood flow enhances mTOR signaling pathways in a rat model. Heart Vessel. 2016, 31, 1685–1695. [Google Scholar] [CrossRef]
- von Maltzahn, J.; Bentzinger, C.F.; Rudnicki, M.A. Wnt7a-Fzd7 signalling directly activates the Akt/mTOR anabolic growth pathway in skeletal muscle. Nat. Cell Biol. 2011, 14, 186–191. [Google Scholar] [CrossRef]
- Ameln, H.; Gustafsson, T.; Sundberg, C.J.; Okamoto, K.; Jansson, E.; Poellinger, L.; Makino, Y. Physiological activation of hypoxia inducible factor-1 in human skeletal muscle. FASEB J. 2005, 19, 1009–1011. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Ma, N.; Li, X.; Kang, M.; Guo, M.; Song, L. Application of GC/MS-based metabonomic profiling in studying the therapeutic effects of Aconitum carmichaeli with Ampelopsis japonica extract on collagen-induced arthritis in rats. Molecules 2019, 24, 1934. [Google Scholar] [CrossRef] [PubMed]
- Weinberger, A.; Halpern, M.; Zahalka, M.A.; Quintana, F.; Traub, L.; Moroz, C. Placental immunomodulator ferritin, a novel immunoregulator, suppresses experimental arthritis. Arthritis Rheum. 2003, 48, 846–853. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamada, Y.; Arai, Y.; Toyama, S.; Inoue, A.; Nakagawa, S.; Fujii, Y.; Kaihara, K.; Cha, R.; Mazda, O.; Takahashi, K. Hypoxia with or without Treadmill Exercises Affects Slow-Twitch Muscle Atrophy and Joint Destruction in a Rat Model of Rheumatoid Arthritis. Int. J. Mol. Sci. 2023, 24, 9761. https://doi.org/10.3390/ijms24119761
Kamada Y, Arai Y, Toyama S, Inoue A, Nakagawa S, Fujii Y, Kaihara K, Cha R, Mazda O, Takahashi K. Hypoxia with or without Treadmill Exercises Affects Slow-Twitch Muscle Atrophy and Joint Destruction in a Rat Model of Rheumatoid Arthritis. International Journal of Molecular Sciences. 2023; 24(11):9761. https://doi.org/10.3390/ijms24119761
Chicago/Turabian StyleKamada, Yoichiro, Yuji Arai, Shogo Toyama, Atsuo Inoue, Shuji Nakagawa, Yuta Fujii, Kenta Kaihara, Ryota Cha, Osam Mazda, and Kenji Takahashi. 2023. "Hypoxia with or without Treadmill Exercises Affects Slow-Twitch Muscle Atrophy and Joint Destruction in a Rat Model of Rheumatoid Arthritis" International Journal of Molecular Sciences 24, no. 11: 9761. https://doi.org/10.3390/ijms24119761
APA StyleKamada, Y., Arai, Y., Toyama, S., Inoue, A., Nakagawa, S., Fujii, Y., Kaihara, K., Cha, R., Mazda, O., & Takahashi, K. (2023). Hypoxia with or without Treadmill Exercises Affects Slow-Twitch Muscle Atrophy and Joint Destruction in a Rat Model of Rheumatoid Arthritis. International Journal of Molecular Sciences, 24(11), 9761. https://doi.org/10.3390/ijms24119761