
Iron Availability Influences Protein Carbonylation in Arabidopsis thaliana Plants


Abstract
1. Introduction
2. Results
2.1. Iron Homeostasis Influences the Carbonylation of Proteins in A. thaliana In Vivo under Normal Growth Conditions
2.2. Iron Homeostasis Impacts on the Occurrence of Protein Carbonylation under Stress
2.3. Iron Excess and Iron-deficient Conditions Influence Protein Carbonylation in the A. thaliana Seedlings
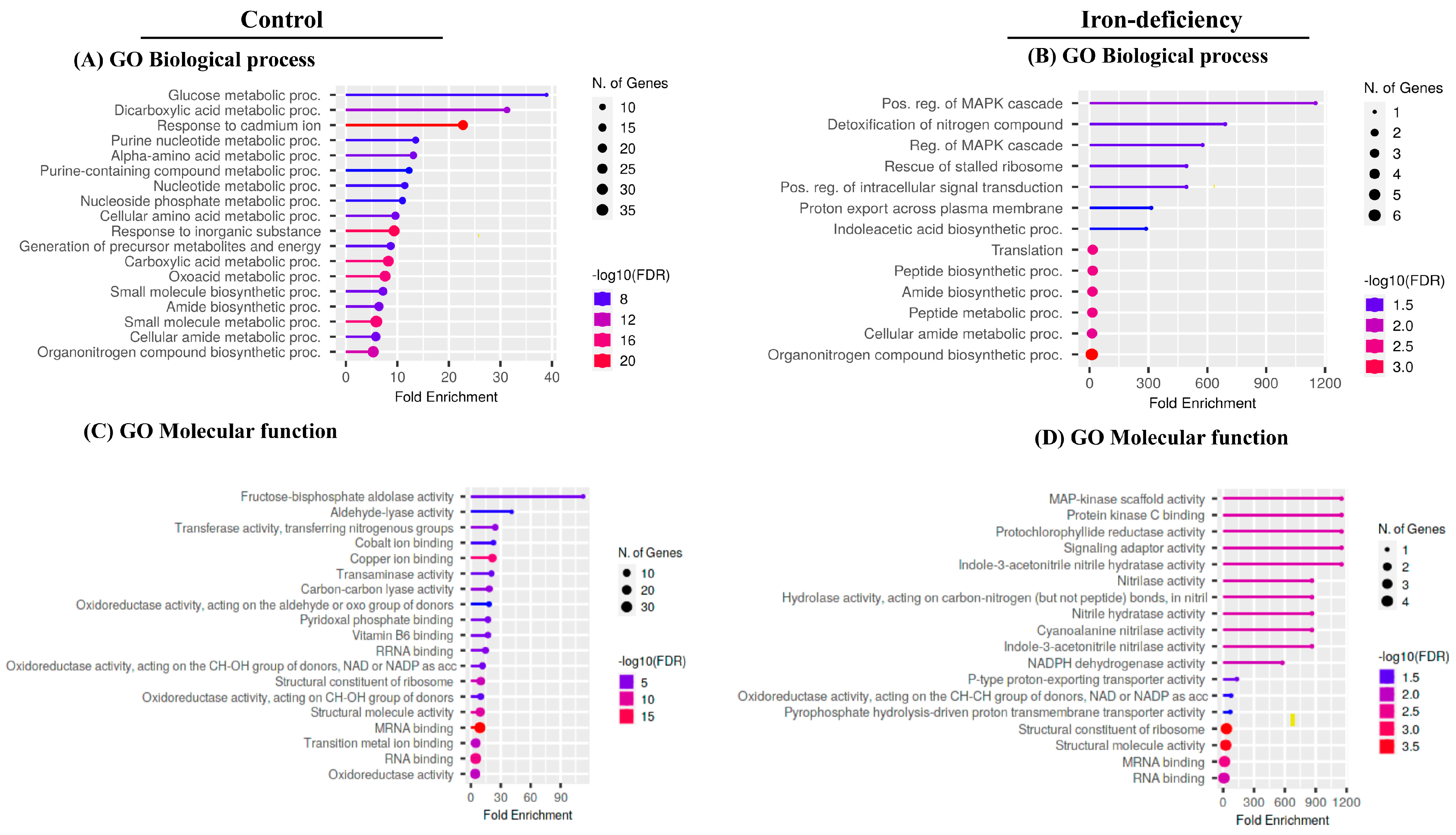
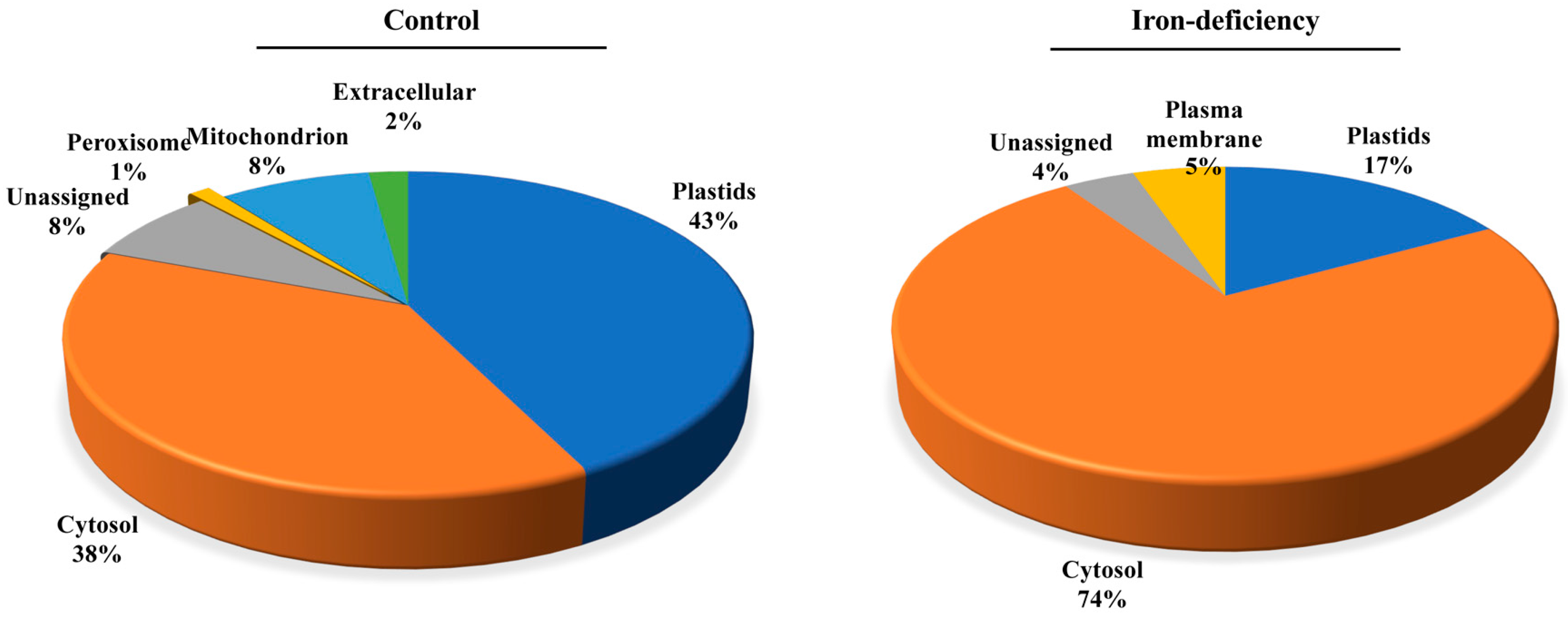
2.4. Identification of Carbonylated Proteins in WT in Response to Iron Deficiency by LC-MS/MS Analysis
2.5. Protein Carbonylation Targeted Different Processes under Control and Iron-deficient Conditions
3. Discussion
3.1. Ferritins are More Relevant to the Steady State Occurrence of Protein Carbonylation under Normal Growth Conditions in Plant Organs
3.2. The Induction of Protein Carbonylation by Intracellular Iron Is Independent of An Oxidative Stress
3.3. Protein Carbonylation Appears as a Candidate Mechanism for the Control of Iron Deficiency Response in A. thaliana
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Profiling of Total Carbonylation Levels at Different Growth Phases and in Different Organs of Arabidopsis Plants
4.3. Heat Stress Treatment
4.4. Iron Treatments
4.5. H2O2 Content Estimation
4.6. DAB and NBT Staining Procedures
4.7. Lipid Peroxidation Assay
4.8. Antioxidant Enzyme Activity of Catalase (CAT) and Superoxide Dismutase (SOD)
4.9. Protein Extraction
4.10. Derivatization of Total Protein with Fluorescent Hydrazide Dye (Cy5.5 and Cy7.5)
4.11. Gel Electrophoresis of Fluorescent Labeled Proteins (1D-PAGE)
4.12. In-gel Fluorescence-based Detection of the Total Proteins and the Carbonylated Proteins
4.13. Determination of Iron (Fe) Content
4.14. Identification of Carbonylated Proteins and Carbonylated Sites
4.15. Database Searching and Criteria Used for Protein Identification and Data Analysis
4.16. Gene Ontology Enrichment Analysis (GO Analysis)
4.17. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Madian, A.G.; Regnier, F.E. Proteomic Identification of Carbonylated Proteins and Their Oxidation Sites. J. Proteome Res. 2010, 9, 3766–3780. [Google Scholar] [CrossRef]
- Tola, A.J.; Jaballi, A.; Missihoun, T.D. Protein Carbonylation: Emerging Roles in Plant Redox Biology and Future Prospects. Plants 2021, 10, 1451. [Google Scholar] [CrossRef]
- Ben-Nissan, G.; Sharon, M. Regulating the 20S Proteasome Ubiquitin-Independent Degradation Pathway. Biomolecules 2014, 4, 862–884. [Google Scholar] [CrossRef] [PubMed]
- Reichmann, D.; Voth, W.; Jakob, U. Maintaining a Healthy Proteome during Oxidative Stress. Mol. Cell 2018, 69, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Dalle-Donne, I.; Aldini, G.; Carini, M.; Colombo, R.; Rossi, R.; Milzani, A.D.G. Protein carbonylation, cellular dysfunction, and disease progression. J. Cell. Mol. Med. 2006, 10, 389–406. [Google Scholar] [CrossRef] [PubMed]
- Møller, I.M.; Rogowska-Wrzesinska, A.; Rao, R. Protein carbonylation and metal-catalyzed protein oxidation in a cellular perspective. J. Proteom. 2011, 74, 2228–2242. [Google Scholar] [CrossRef]
- Biswas, S.; Mano, J. Lipid Peroxide-Derived Short-Chain Carbonyls Mediate Hydrogen Peroxide-Induced and Salt-Induced Programmed Cell Death in Plants. Plant Physiol. 2015, 168, 885–898. [Google Scholar] [CrossRef]
- Islam, M.; Ye, W.; Matsushima, D.; Munemasa, S.; Okuma, E.; Nakamura, Y.; Biswas, S.; Mano, J.; Murata, Y. Reactive Carbonyl Species Mediate ABA Signaling in Guard Cells. Plant Cell Physiol. 2016, 57, 2552–2563. [Google Scholar] [CrossRef]
- Biswas, S.; Fukaki, H.; Mori, I.C.; Nakahara, K.; Mano, J. Reactive oxygen species and reactive carbonyl species constitute a feed-forward loop in auxin signaling for lateral root formation. Plant J. 2019, 100, 536–548. [Google Scholar] [CrossRef] [PubMed]
- Møller, I.M.; Jensen, P.E.; Hansson, A. Oxidative Modifications to Cellular Components in Plants. Annu. Rev. Plant Biol. 2007, 58, 459–481. [Google Scholar] [CrossRef]
- Møller, I.M.; Sweetlove, L.J. ROS signalling—specificity is required. Trends Plant Sci. 2010, 15, 370–374. [Google Scholar] [CrossRef]
- Jaballi, A.; Missihoun, T.D. The phytohormone abscisic acid modulates protein carbonylation in Arabidopsis thaliana. Physiol. Plant. 2022, 174, e13658. [Google Scholar] [CrossRef]
- Kristensen, B.K.; Askerlund, P.; Bykova, N.V.; Egsgaard, H.; Møller, I.M. Identification of oxidised proteins in the matrix of rice leaf mitochondria by immunoprecipitation and two-dimensional liquid chromatography-tandem mass spectrometry. Phytochemistry 2004, 65, 1839–1851. [Google Scholar] [CrossRef]
- Sweetlove, L.J.; Møller, I.M. Chapter 1 Oxidation of Proteins in Plants—Mechanisms and Consequences. Adv. Bot. Res. 2009, 52, 1–23. [Google Scholar] [CrossRef]
- Bollineni, R.C.; Fedorova, M.; Hoffmann, R. Qualitative and quantitative evaluation of derivatization reagents for different types of protein-bound carbonyl groups. Analyst 2013, 138, 5081–5088. [Google Scholar] [CrossRef]
- Kehrer, J.P. The Haber-Weiss reaction and mechanisms of toxicity. Toxicology 2000, 149, 43–50. [Google Scholar] [CrossRef]
- Mène-Saffrané, L.; Davoine, C.; Stolz, S.; Majcherczyk, P.; Farmer, E.E. Genetic Removal of Tri-unsaturated Fatty Acids Suppresses Developmental and Molecular Phenotypes of an Arabidopsis Tocopherol-deficient Mutant: Whole-Body Mapping of Malondialdehyde Pools in a Complex Eukaryote. J. Biol. Chem. 2007, 282, 35749–35756. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C. Toxicity of iron and hydrogen peroxide: The Fenton reaction. Toxicol. Lett. 1995, 82–83, 969–974. [Google Scholar] [CrossRef]
- Jeong, J.; Guerinot, M.L. Homing in on iron homeostasis in plants. Trends Plant Sci. 2009, 14, 280–285. [Google Scholar] [CrossRef]
- Broadley, M.; Brown, P.; Cakmak, I.; Rengel, Z.; Zhao, F. Function of Nutrients. Marschner’s Miner. Nutr. High. Plants Third Ed. 2012, 191–248. [Google Scholar] [CrossRef]
- Vert, G.; Grotz, N.; Dédaldéchamp, F.; Gaymard, F.; Guerinot, M.L.; Briat, J.F.; Curie, C. IRT1, an arabidopsis transporter essential for iron uptake from the soil and for plant growth. Plant Cell 2002, 14, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Roschzttardtz, H.; Conéjéro, G.; Divol, F.; Alcon, C.; Verdeil, J.-L.; Curie, C.; Mari, S. New insights into Fe localization in plant tissues. Front. Plant Sci. 2013, 4, 350. [Google Scholar] [CrossRef]
- Mahender, A.; Swamy, B.P.M.; Anandan, A.; Ali, J. Tolerance of Iron-Deficient and -Toxic Soil Conditions in Rice. Plants 2019, 8, 31. [Google Scholar] [CrossRef] [PubMed]
- Becker, M.; Asch, F. Iron toxicity in rice—conditions and management concepts. J. Plant Nutr. Soil Sci. 2005, 168, 558–573. [Google Scholar] [CrossRef]
- Kobayashi, T.; Nishizawa, N.K. Iron Uptake, Translocation, and Regulation in Higher Plants. Annu. Rev. Plant Biol. 2012, 63, 131–152. [Google Scholar] [CrossRef]
- Lei, G.J.; Zhu, X.F.; Wang, Z.W.; Dong, F.; Dong, N.Y.; Zheng, S.J. Abscisic acid alleviates iron deficiency by promoting root iron reutilization and transport from root to shoot in Arabidopsis. Plant Cell Environ. 2014, 37, 852–863. [Google Scholar] [CrossRef]
- Séguéla, M.; Briat, J.-F.; Vert, G.; Curie, C. Cytokinins negatively regulate the root iron uptake machinery in Arabidopsis through a growth-dependent pathway. Plant J. 2008, 55, 289–300. [Google Scholar] [CrossRef]
- Maurer, F.; Müller, S.; Bauer, P. Suppression of Fe deficiency gene expression by jasmonate. Plant Physiol. Biochem. 2011, 49, 530–536. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Iwamoto, M.; Maekawa, M.; Saito, A.; Higo, H.; Higo, K. Evolutionary relationship of plant catalase genes inferred from exon-intron structures: Isozyme divergence after the separation of monocots and dicots. Theor. Appl. Genet. 1998, 97, 9–19. [Google Scholar] [CrossRef]
- Caverzan, A.; Passaia, G.; Rosa, S.B.; Ribeiro, C.W.; Lazzarotto, F.; Margis-Pinheiro, M. Plant responses to stresses: Role of ascorbate peroxidase in the antioxidant protection. Genet. Mol. Biol. 2012, 35, 1011–1019. [Google Scholar] [CrossRef] [PubMed]
- Vogelsang, L.; Dietz, K.-J. Plant thiol peroxidases as redox sensors and signal transducers in abiotic stress acclimation. Free. Radic. Biol. Med. 2022, 193, 764–778. [Google Scholar] [CrossRef] [PubMed]
- König, J.; Jung, T.; Grune, T. Protein Carbonylation in Aging and Senescence. Protein Carbonylation Princ. Anal. Biol. Implic. 2017, 272–290. [Google Scholar] [CrossRef]
- Massenet, O.; Lobreaux, S.; Briat, J.-F. Iron induces ferritin synthesis in maize plantlets. Plant Mol. Biol. 1992, 19, 563–575. [Google Scholar] [CrossRef]
- Briat, J.-F.; Ravet, K.; Arnaud, N.; Duc, C.; Boucherez, J.; Touraine, B.; Cellier, F.; Gaymard, F. New insights into ferritin synthesis and function highlight a link between iron homeostasis and oxidative stress in plants. Ann. Bot. 2010, 105, 811–822. [Google Scholar] [CrossRef]
- Balla, G.; Jacob, H.S.; Balla, J.; Rosenberg, M.; Nath, K.; Apple, F.; Eaton, J.W.; Vercellotti, G.M. Ferritin: A cytoprotective antioxidant strategem of endothelium. J. Biol. Chem. 1992, 267, 18148–18153. [Google Scholar] [CrossRef]
- Zancani, M.; Peresson, C.; Biroccio, A.; Federici, G.; Urbani, A.; Murgia, I.; Soave, C.; Micali, F.; Vianello, A.; Macrì, F. Evidence for the presence of ferritin in plant mitochondria. Eur. J. Biol. Chem. 2004, 271, 3657–3664. [Google Scholar] [CrossRef]
- Tarantino, D.; Casagrande, F.; Soave, C.; Murgia, I. Knocking out of the mitochondrial AtFer4 ferritin does not alter response of Arabidopsis plants to abiotic stresses. J. Plant Physiol. 2010, 167, 453–460. [Google Scholar] [CrossRef]
- Theil, E.C. FERRITIN: STRUCTURE, GENE REGULATION, AND CELLULAR FUNCTION IN ANIMALS, PLANTS, AND MICROORGANISMS. Annu. Rev. Biochem. 1987, 56, 289–315. [Google Scholar] [CrossRef]
- Briat, J.; Lobreaux, S. Iron transport and storage in plants. Trends Plant Sci. 1997, 2, 187–193. [Google Scholar] [CrossRef]
- Curie, C.; Briat, J.-F. Iron Transport and Signaling in Plants. Annu. Rev. Plant Biol. 2003, 54, 183–206. [Google Scholar] [CrossRef]
- Harrison, P.M.; Arosio, P. The ferritins: Molecular properties, iron storage function and cellular regulation. Biochim. Biophys. Acta Bioenerg. 1996, 1275, 161–203. [Google Scholar] [CrossRef] [PubMed]
- Theil, E.C.; Matzapetakis, M.; Liu, X. Ferritins: Iron/oxygen biominerals in protein nanocages. JBIC J. Biol. Inorg. Chem. 2006, 11, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Gaymard, F.; Boucherez, J.; Briat, J.-F. Characterization of a ferritin mRNA from Arabidopsis thaliana accumulated in response to iron through an oxidative pathway independent of abscisic acid. Biochem. J. 1996, 318, 67–73. [Google Scholar] [CrossRef]
- Yadav, K.; Patel, P.; Srivastava, A.K.; Ganapathi, T.R. Overexpression of native ferritin gene MusaFer1 enhances iron content and oxidative stress tolerance in transgenic banana plants. PLoS ONE 2017, 12, e0188933. [Google Scholar] [CrossRef]
- Petit, J.-M.; Briat, J.-F.; Lobréaux, S. Structure and differential expression of the four members of the Arabidopsis thaliana ferritin gene family. Biochem. J. 2001, 359, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Ravet, K.; Touraine, B.; Boucherez, J.; Briat, J.-F.; Gaymard, F.; Cellier, F. Ferritins control interaction between iron homeostasis and oxidative stress in Arabidopsis. Plant J. 2009, 57, 400–412. [Google Scholar] [CrossRef]
- Kim, J.-H.; Sedlak, M.; Gao, Q.; Riley, C.P.; Regnier, F.E.; Adamec, J. Oxidative Stress Studies in Yeast with a Frataxin Mutant: A Proteomics Perspective. J. Proteome Res. 2010, 9, 730–736. [Google Scholar] [CrossRef] [PubMed]
- Busi, M.V.; Maliandi, M.V.; Valdez, H.; Clemente, M.; Zabaleta, E.J.; Araya, A.; Gomez-Casati, D.F. Deficiency of Arabidopsis thaliana frataxin alters activity of mitochondrial Fe-S proteins and induces oxidative stress. Plant J. 2006, 48, 873–882. [Google Scholar] [CrossRef]
- Boyes, D.C.; Zayed, A.M.; Ascenzi, R.; McCaskill, A.J.; Hoffman, N.E.; Davis, K.R.; Görlach, J. Growth Stage-Based Phenotypic Analysis of Arabidopsis: A Model for High Throughput Functional Genomics in Plants. Plant Cell 2001, 13, 1499–1510. [Google Scholar] [CrossRef]
- Gautam, C.K.; Tsai, H.-H.; Schmidt, W. IRONMAN tunes responses to iron deficiency in concert with environmental pH. Plant Physiol. 2021, 187, 1728–1745. [Google Scholar] [CrossRef]
- Gautam, C.K.; Tsai, H.-H.; Schmidt, W. A Quick Method to Quantify Iron in Arabidopsis Seedlings Iron Transport in Plants View Project PH Sensing in Plants View Project. Bio. Protoc. 2022, 12, e4324. [Google Scholar] [CrossRef]
- Matamoros, M.A.; Kim, A.; Peñuelas, M.; Ihling, C.; Griesser, E.; Hoffmann, R.; Fedorova, M.; Frolov, A.; Becana, M. Protein Carbonylation and Glycation in Legume Nodules. Plant Physiol. 2018, 177, 1510–1528. [Google Scholar] [CrossRef]
- Rojas Echeverri, J.C.; Milkovska-Stamenova, S.; Hoffmann, R. A workflow towards the reproducible identification and quantitation of protein carbonylation sites in human plasma. Antioxidants 2021, 10, 369. [Google Scholar] [CrossRef] [PubMed]
- Chavez, J.; Wu, J.; Han, B.; Chung, W.-G.; Maier, C.S. New Role for an Old Probe: Affinity Labeling of Oxylipid Protein Conjugates by N‘-Aminooxymethylcarbonylhydrazino d-biotin. Anal. Chem. 2006, 78, 6847–6854. [Google Scholar] [CrossRef] [PubMed]
- Bollineni, R.C.; Hoffmann, R.; Fedorova, M. Proteome-wide profiling of carbonylated proteins and carbonylation sites in HeLa cells under mild oxidative stress conditions. Free. Radic. Biol. Med. 2014, 68, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.; Schmidt, W. Environmentally Induced Plasticity of Root Hair Development in Arabidopsis. Plant Physiol. 2004, 134, 409–419. [Google Scholar] [CrossRef]
- Chen, J.-G.; Ullah, H.; Temple, B.; Liang, J.; Guo, J.; Alonso, J.M.; Ecker, J.R.; Jones, A.M. RACK1 mediates multiple hormone responsiveness and developmental processes in Arabidopsis. J. Exp. Bot. 2006, 57, 2697–2708. [Google Scholar] [CrossRef]
- Guo, J.; Wang, J.; Xi, L.; Huang, W.-D.; Liang, J.; Chen, J.-G. RACK1 is a negative regulator of ABA responses in Arabidopsis. J. Exp. Bot. 2009, 60, 3819–3833. [Google Scholar] [CrossRef]
- Ge, S.; Jung, D.; Yao, R. ShinyGO: A Graphical Gene-Set Enrichment Tool for Animals and Plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef]
- Hooper, C.M.; Castleden, I.R.; Tanz, S.; Aryamanesh, N.; Millar, A.H. SUBA4: The interactive data analysis centre for Arabidopsis subcellular protein locations. Nucleic Acids Res. 2017, 45, D1064–D1074. [Google Scholar] [CrossRef] [PubMed]
- Johansson, E.; Olsson, O.; Nyström, T. Progression and Specificity of Protein Oxidation in the Life Cycle of Arabidopsis thaliana. J. Biol. Chem. 2004, 279, 22204–22208. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Contento, A.L.; Bassham, D.C. Disruption of Autophagy Results in Constitutive Oxidative Stress in Arabidopsis. Autophagy 2007, 3, 257–258. [Google Scholar] [CrossRef] [PubMed]
- Reyt, G.; Boudouf, S.; Boucherez, J.; Gaymard, F.; Briat, J.-F. Iron- and Ferritin-Dependent Reactive Oxygen Species Distribution: Impact on Arabidopsis Root System Architecture. Mol. Plant 2015, 8, 439–453. [Google Scholar] [CrossRef]
- Briat, J.-F.; Duc, C.; Ravet, K.; Gaymard, F. Ferritins and iron storage in plants. Biochim. Biophys. Acta Gen. Subj. 2010, 1800, 806–814. [Google Scholar] [CrossRef]
- Tissot, N.; Robe, K.; Gao, F.; Grant-Grant, S.; Boucherez, J.; Bellegarde, F.; Maghiaoui, A.; Marcelin, R.; Izquierdo, E.; Benhamed, M.; et al. Transcriptional integration of the responses to iron availability in Arabidopsis by the bHLH factor ILR3. New Phytol. 2019, 223, 1433–1446. [Google Scholar] [CrossRef]
- Fodor, E.; Szabó-Nagy, A.; Erdei, L. The Effects of Cadmium on the Fluidity and H+-ATPase Activity of Plasma Membrane from Sunflower and Wheat Roots. J. Plant Physiol. 1995, 147, 87–92. [Google Scholar] [CrossRef]
- Schaller, G.E.; Sussman, M.R. Phosphorylation of the plasma-membrane H+-ATPase of oat roots by a calcium-stimulated protein kinase. Planta 1988, 173, 509–518. [Google Scholar] [CrossRef]
- Portillo, F. Regulation of plasma membrane H+-ATPase in fungi and plants. Biochim. et Biophys. Acta (BBA) - Rev. Biomembr. 2000, 1469, 31–42. [Google Scholar] [CrossRef]
- Wang, J.; Lan, P.; Gao, H.; Zheng, L.; Li, W.; Schmidt, W. Expression changes of ribosomal proteins in phosphate- and iron-deficient Arabidopsis roots predict stress-specific alterations in ribosome composition. BMC Genom. 2013, 14, 783. [Google Scholar] [CrossRef]
- Gallo, S.; Ricciardi, S.; Manfrini, N.; Pesce, E.; Oliveto, S.; Calamita, P.; Mancino, M.; Maffioli, E.; Moro, M.; Crosti, M.; et al. RACK1 Specifically Regulates Translation through Its Binding to Ribosomes. Mol. Cell. Biol. 2018, 38, e00230-18. [Google Scholar] [CrossRef]
- Guo, J.; Wang, S.; Valerius, O.; Hall, H.; Zeng, Q.; Li, J.-F.; Weston, D.J.; Ellis, B.E.; Chen, J.-G. Involvement of Arabidopsis RACK1 in Protein Translation and Its Regulation by Abscisic Acid. Plant Physiol. 2011, 155, 370–383. [Google Scholar] [CrossRef]
- Romero, A.M.; Ramos-Alonso, L.; Alepuz, P.; Puig, S.; Martínez-Pastor, M.T. Global translational repression induced by iron deficiency in yeast depends on the Gcn2/eIF2α pathway. Sci. Rep. 2020, 10, 233. [Google Scholar] [CrossRef]
- Long, T.A.; Tsukagoshi, H.; Busch, W.; Lahner, B.; Salt, D.E.; Benfey, P.N. The bHLH Transcription Factor POPEYE Regulates Response to Iron Deficiency in Arabidopsis Roots. Plant Cell 2010, 22, 2219–2236. [Google Scholar] [CrossRef]
- Selote, D.; Samira, R.; Matthiadis, A.; Gillikin, J.W.; Long, T.A. Iron-Binding E3 Ligase Mediates Iron Response in Plants by Targeting Basic Helix-Loop-Helix Transcription Factors. Plant Physiol. 2015, 167, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Selote, D.; Matthiadis, A.; Gillikin, J.W.; Sato, M.H.; Long, T.A. The E3 ligase BRUTUS facilitates degradation of VOZ1/2 transcription factors. Plant Cell Environ. 2018, 41, 2463–2474. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-García, A.; Morales, M.L.; Garrido-García, V.; García-Baquero, I.; Leivas, A.; Carreño-Tarragona, G.; Sánchez, R.; Arenas, A.; Cedena, T.; Ayala, R.M.; et al. Protein Carbonylation in Patients with Myelodysplastic Syndrome: An Opportunity for Deferasirox Therapy. Antioxidants 2019, 8, 508. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Missihoun, T.D.; Bartels, D. The role of Arabidopsis aldehyde dehydrogenase genes in response to high temperature and stress combinations. J. Exp. Bot. 2017, 68, 4295–4308. [Google Scholar] [CrossRef] [PubMed]
- Hindt, M.N.; Akmakjian, G.Z.; Pivarski, K.L.; Punshon, T.; Baxter, I.; Salt, D.E.; Guerinot, M.L. BRUTUS and its paralogs, BTS LIKE1 and BTS LIKE2, encode important negative regulators of the iron deficiency response in Arabidopsis thaliana. Metallomics 2017, 9, 876–890. [Google Scholar] [CrossRef]
- Li, Z.-G. Measurement of Signaling Molecules Calcium Ion, Reactive Sulfur Species, Reactive Carbonyl Species, Reactive Nitrogen Species, and Reactive Oxygen Species in Plants. Plant Signal. Mol. Role Regul. Under Stress. Environ. 2019, 83–103. [Google Scholar] [CrossRef]
- Brumbarova, T.; Bauer, P.; Ivanov, R. Molecular mechanisms governing Arabidopsis iron uptake. Trends Plant Sci. 2015, 20, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ji, T.-T.; Li, T.-T.; Tian, Y.-Y.; Wang, L.-F.; Liu, W.-C. Jasmonic acid promotes leaf senescence through MYC2-mediated repression of CATALASE2 expression in Arabidopsis. Plant Sci. 2020, 299, 110604. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Zhang, J.; Ji, X.; Feng, Z.; Wang, X.; Huang, W.; You, C.; Wang, X.; Hao, Y. Low nitrate alleviates iron deficiency by regulating iron homeostasis in apple. Plant, Cell Environ. 2021, 44, 1869–1884. [Google Scholar] [CrossRef]
- Tewari, R.K.; Hadacek, F.; Sassmann, S.; Lang, I. Iron deprivation-induced reactive oxygen species generation leads to non-autolytic PCD in Brassica napus leaves. Environ. Exp. Bot. 2013, 91, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Yusuf, M.; Singh, P.; Sardar, M.; Sarin, N. Histochemical Detection of Superoxide and H2O2 Accumulation in Brassica juncea Seedlings. Bio-Protocol 2014, 8, e1108. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.B.; Hahn, E.-J.; Paek, K.-Y. Effects of temperature on oxidative stress defense systems, lipid peroxidation and lipoxygenase activity in Phalaenopsis. Plant Physiol. Biochem. 2005, 43, 213–223. [Google Scholar] [CrossRef]
- Chen, T.; Zhang, B. Measurements of Proline and Malondialdehyde Content and Antioxidant Enzyme Activities in Leaves of Drought Stressed Cotton. Bio-Protocol 2016, 6, e1913. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Tamarit, J.; de Hoogh, A.; Obis, E.; Alsina, D.; Cabiscol, E.; Ros, J. Analysis of oxidative stress-induced protein carbonylation using fluorescent hydrazides. J. Proteom. 2012, 75, 3778–3788. [Google Scholar] [CrossRef]
- Fangue-Yapseu, G.Y.; Tola, A.J.; Missihoun, T.D. Proteome-wide analysis of hydrogen peroxide-induced protein carbonylation in Arabidopsis thaliana. Front. Plant Sci. 2022, 13, 4999. [Google Scholar] [CrossRef] [PubMed]
- Tola, A.J.; Missihoun, T.D. Ammonium sulfate-based prefractionation improved proteome coverage and detection of carbonylated proteins in Arabidopsis thaliana leaf extract. Planta 2023, 257, 1–19. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SN | Protein Name | Accession Number | TAIR Gene Code | Molecular Mass (kDa) | Peptide Number | Localization | Functional Category |
|---|---|---|---|---|---|---|---|
| 1 | Catalase 3 | B9DG18 | AT1G20620 | 56 | 12 | Peroxisome | ROS scavenging/stress defense response/Antioxidant enzymes |
| 2 | Monodehydroascorbate reductase 1 | Q9LFA3 | AT3G52880 | 46 | 7 | Peroxisome | |
| 3 | Thioredoxin superfamily protein | F4JBC9 | AT3G26060 | 24 | 7 | Chloroplast | |
| 4 | NAD(P)H dehydrogenase (quinone) FQR1 | Q9LSQ5 | AT5G54500 | 22 | 2 | Chloroplast | |
| 5 | Alcohol dehydrogenase class-3 | Q96533 | AT5G43940 | 41 | 10 | Cytosol | |
| 6 | V-type proton ATPase subunit B1 | P11574 | AT1G76030 | 54 | 9 | Plasma membrane | Ion channel transport |
| 7 | V-type proton ATPase subunit C | Q9SDS7 | AT1G12840 | 43 | 4 | Plasma membrane | |
| 8 | Heat shock 70 kDa protein 9 | Q8GUM2 | AT4G37910 | 73 | 6 | Mitochondria | Protein folding |
| 9 | Chaperonin 60 subunit beta 1 | P21240 | AT1G55490 | 64 | 15 | Chloroplast | |
| 10 | Peptidyl-prolyl cis-trans isomerase CYP18-4 | Q42406 | AT4G34870 | 18 | 9 | Cytosol | |
| 11 | Ferredoxin--nitrite reductase | Q39161 | AT2G15620 | 66 | 6 | Chloroplast | Nitrate assimilation/Nitrogen metabolism |
| 12 | Glutamate synthase 1 [NADH] | Q9LV03 | AT5G53460 | 242 | 11 | Chloroplast | Amino acid metabolism |
| 13 | Aspartate aminotransferase | P46643 | AT2G30970 | 48 | 7 | Mitochondria | |
| 14 | Glutamate dehydrogenase 1 | Q43314 | AT5G18170 | 45 | 5 | Chloroplast | |
| 15 | Cysteine synthase | P47999 | AT2G43750 | 42 | 7 | Chloroplast | |
| 16 | Imidazole glycerol phosphate synthase hisHF | Q9SZ30 | AT4G26900 | 64 | 6 | Chloroplast | |
| 17 | Phosphoglycerate kinase 3 | Q9SAJ4 | AT1G79550 | 42 | 10 | Cytosol | Carbohydrate metabolism/Calvin cycle/ |
| 18 | Glucose-6-phosphate 1-dehydrogenase | A0A1I9LLZ6 | AT3G27300 | 67 | 3 | Cytosol | |
| 19 | Fructose-bisphosphate aldolase 6 | Q9SJQ9 | AT2G36460 | 38 | 5 | Cytosol | |
| 20 | Triosephosphate isomerase | Q9SKP6 | AT2G21170 | 33 | 8 | Chloroplast | |
| 21 | Malate dehydrogenase 1 | Q9ZP06 | AT1G53240 | 36 | 7 | Mitochondria | TCA cycle/Energy metabolism |
| 22 | Adenylyl cyclase-associated protein | A0A1P8B7W9 | AT4G34490 | 58 | 3 | Chloroplast | Signalling |
| 23 | Metacaspase-4 | O64517 | AT1G79340 | 45 | 2 | Cytosol | Plant defense |
| 24 | Biotin carboxylase | F4JYE0 | AT5G35360 | 61 | 9 | Chloroplast | Fatty acid metabolism |
| 25 | Formate dehydrogenase | A0A1P8B9L1 | AT5G14780 | 41 | 4 | Mitochondria |
| SN | Protein Name | Accession Number | TAIR Gene Code | Molecular Mass (kDa) | Peptide Number | Localization | Functional Category |
|---|---|---|---|---|---|---|---|
| 1 | Receptor for activated C kinase 1 (RACK1) | O24456 | AT1G18080 | 36 | 8 | Cytosol | Signal transduction |
| 2 | 60S Ribosomal protein L15-1 (RPL15) | O23515 | AT4G16720 | 24 | 4 | Ribosome | Translation |
| 3 | 50S Ribosomal protein L5 (RPL5) | O04603 | AT4G01310 | 28 | 6 | Ribosome | Translation |
| 4 | Nitrilase 1 (NIT1) | P32961 | AT3G44310 | 38 | 6 | Cytosol | Indole acetic acid biosynthesis process |
| 5 | Subtilisin-like protease (SBT1) | Q9LVJ1 | AT3G14067 | 82 | 2 | Cytosol | Proteolysis |
| 6 | Plasma membrane ATPase (HA2) | F4JPJ7 | AT4G30190 | 108 | 4 | Plasma membrane | Proton (H+) transmembrane transport |
| 7 | 30S Ribosomal protein S2 (RPS2) | P56797 | ATCG00160 | 27 | 4 | Ribosome | Translation |
| 8 | NADPH-protochlorophyllide oxidoreductase (POR) | F4I2F8 | AT1G03630 | 44 | 2 | Chloroplast | Chlorophyll biosynthesis |
| SN | Protein Name | Accession Number | Tair Gene Code | Molecular Mass (kDa) | Peptide Number | Functional Category |
|---|---|---|---|---|---|---|
| 1 | Ascorbate peroxidase | F4HU93 | AT5G02500 | 28 | 9 | Stress defense response/Antioxidant enzymes |
| 2 | Peroxidase 32 | Q9LHB9 | AT3G32980 | 39 | 7 | |
| 3 | Monodehyroascorbate reductase 6 | F41576 | AT1G63940 | 52 | 14 | |
| 4 | Selenium binding protein 2 | Q93WNO | AT4G14040 | 54 | 6 | Metal-binding proteins |
| 5 | Magnesium chelatase subunite ChII-2 | Q5XF33 | AT5G45930 | 46 | 3 | |
| 6 | Glutamate--glyoxylate aminotransferase 1 | Q9LR30 | AT1G23310 | 53 | 18 | Glyoxylate metabolism |
| 7 | Oxalate--CoA ligase | Q9SMT7 | AT3G48990 | 56 | 7 | |
| 8 | Transketolase | F4JBY2 | AT3G60750 | 80 | 19 | Carbohydrate/Energy metabolism |
| 9 | Pyruvate kinase | Q94KE3 | AT3G52990 | 57 | 7 | |
| 10 | Sedoheptulose-1,7-bisphosphatase | P46283 | AT3G55800 | 42 | 10 | |
| 11 | Glucose-1-phosphate aldenyltransferase large subunit 1 | P55229 | AT5G19220 | 58 | 10 | |
| 12 | Phosphoribulokinase | P25697 | AT1G32060 | 44 | 9 | |
| 13 | Cytosolic isocitrate dehydrogenase [NADP] | Q9SRZ6 | AT1G65930 | 46 | 17 | |
| 14 | Ferrodoxin-dependent glutamate synthase 1 | Q9ZNZ7 | AT5G04140 | 177 | 19 | Nitrogen–carbon metabolism |
| 15 | Nitrile specifier protein | Q9SDM9 | AT3G16400 | 52 | 16 | |
| 16 | Glutamate-1-semialdehyde-2-aminomutase 1 | P42799 | AT5G63570 | 50 | 7 | |
| 17 | Glutamine synthetase | Q43127 | AT3G55800 | 47 | 12 | |
| 18 | S-adenoyl-L-methionine-dependent methyltrnsferase | AOA1P8B3H0 | AT4G34050 | 32 | 4 | Amino acid metabolism |
| 19 | S-adenoylmethionine synthase 4 | Q9LUT2 | AT5G20720 | 43 | 10 | |
| 20 | Serine hydroxymethyltransferase 1 | Q9SZJ5 | AT4G37930 | 57 | 10 | |
| 21 | Glycine dehydrogenase (decarboxylating) 1 | Q94B78 | AT4G33010 | 113 | 17 | |
| 22 | Chaperonin 60 subunit alpha 1 | P21238 | AT3G09260 | 62 | 17 | Protein folding |
| 23 | PYK10-binding protein 1 | O04314 | AT4G14880 | 32 | 10 | |
| 24 | Elongation factor Tu | P17745 | AT4G20360 | 52 | 11 | Translation/Protein synthesis |
| 25 | Elongation factor G | Q9SI75 | AT1G62750 | 86 | 9 | |
| 26 | Tubulin alpha-2 chain | B9DGT7 | AT1G50010 | 50 | 12 | Cytoskeleton organization |
| 27 | Actin-7 | P53492 | AT5G09810 | 42 | 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tola, A.J.; Missihoun, T.D. Iron Availability Influences Protein Carbonylation in Arabidopsis thaliana Plants. Int. J. Mol. Sci. 2023, 24, 9732. https://doi.org/10.3390/ijms24119732
Tola AJ, Missihoun TD. Iron Availability Influences Protein Carbonylation in Arabidopsis thaliana Plants. International Journal of Molecular Sciences. 2023; 24(11):9732. https://doi.org/10.3390/ijms24119732
Chicago/Turabian StyleTola, Adesola J., and Tagnon D. Missihoun. 2023. "Iron Availability Influences Protein Carbonylation in Arabidopsis thaliana Plants" International Journal of Molecular Sciences 24, no. 11: 9732. https://doi.org/10.3390/ijms24119732
APA StyleTola, A. J., & Missihoun, T. D. (2023). Iron Availability Influences Protein Carbonylation in Arabidopsis thaliana Plants. International Journal of Molecular Sciences, 24(11), 9732. https://doi.org/10.3390/ijms24119732