

Pro- and Anti-Inflammatory Prostaglandins and Cytokines in Humans: A Mini Review



Abstract
1. Introduction
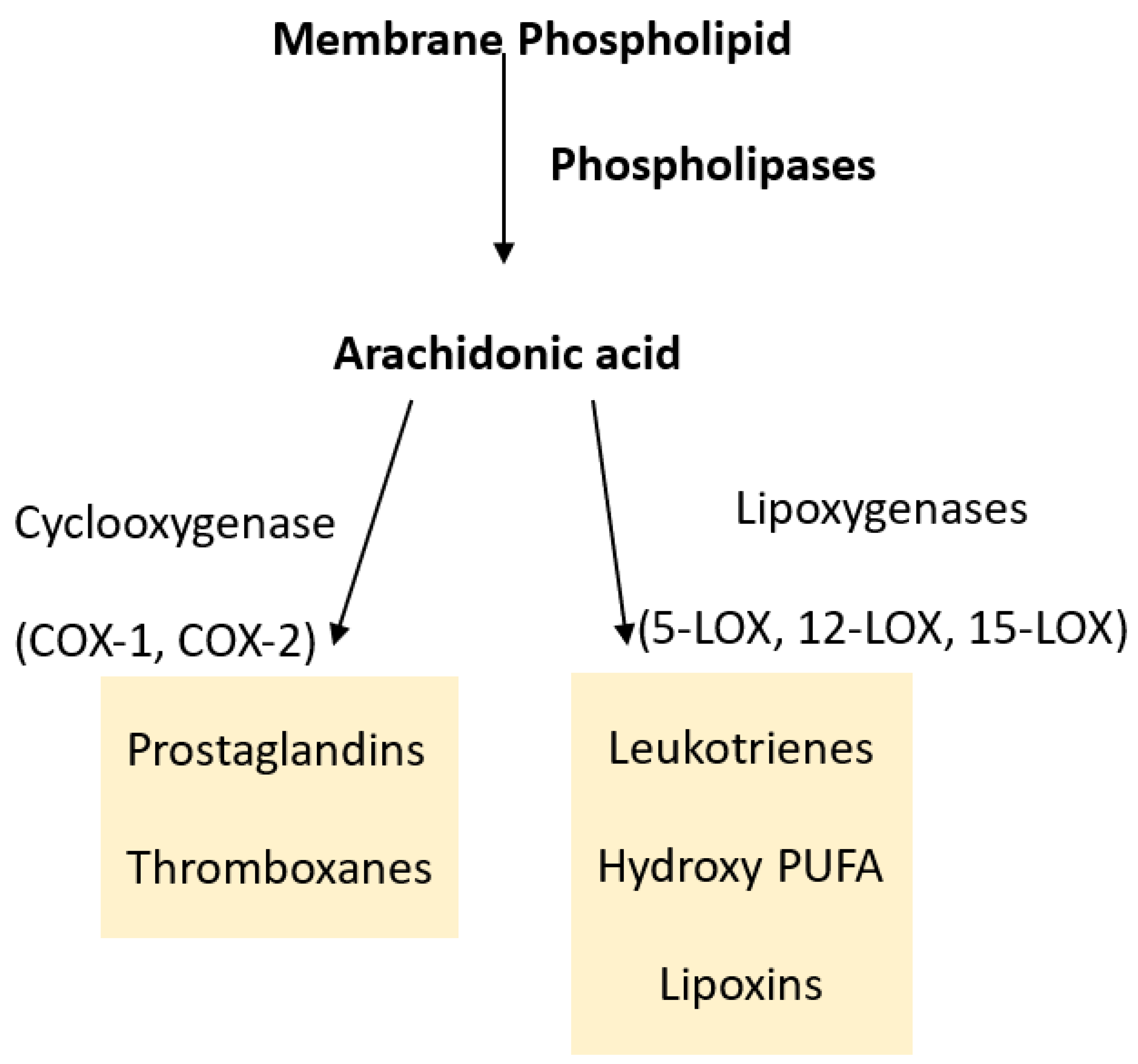
2. Prostaglandins, Leukotrienes and Eicosanoids
2.1. Cyclooxygenases and Prostaglandins
2.2. Lipoxygenases and Leukotrienes
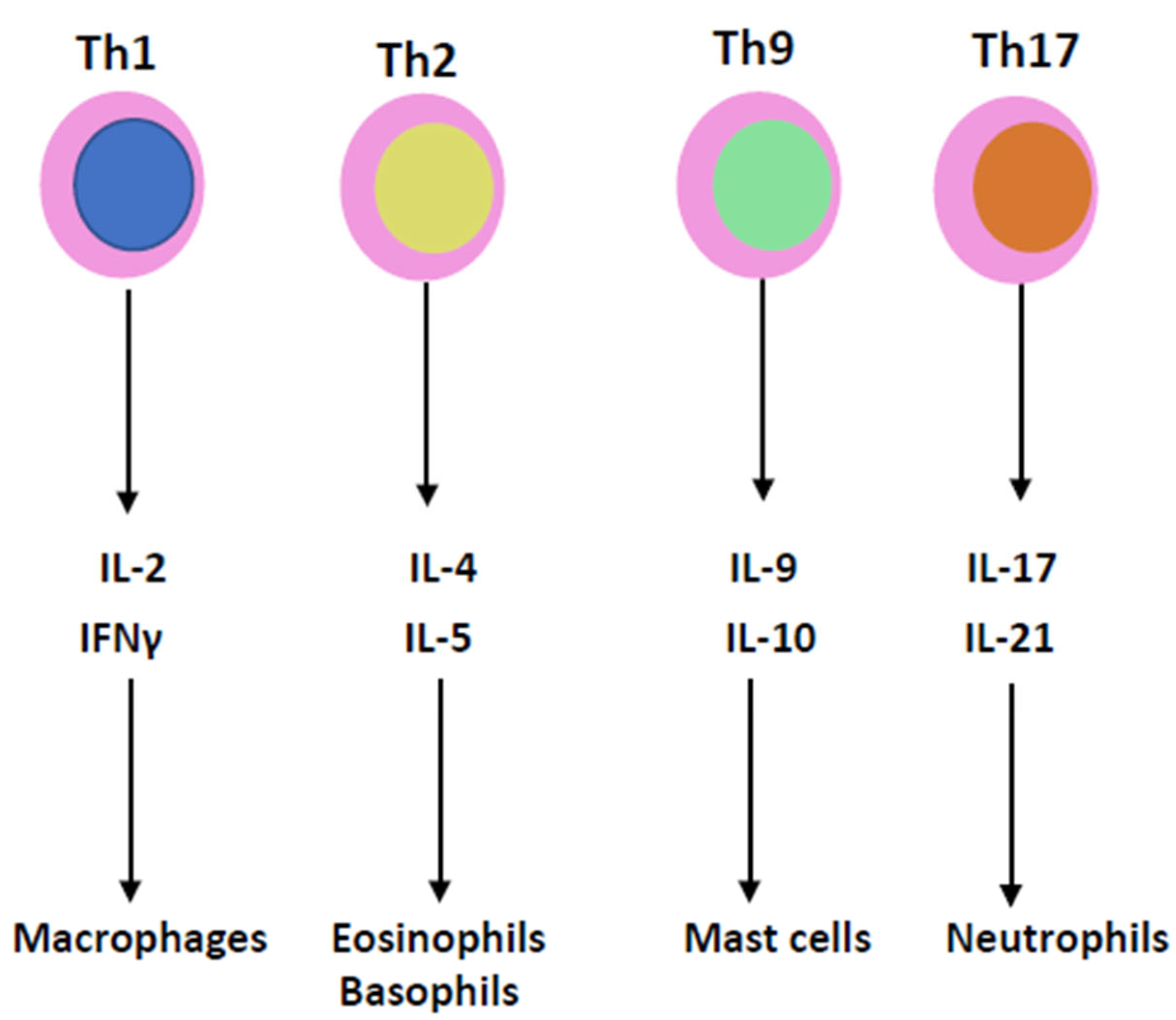
3. Cytokines
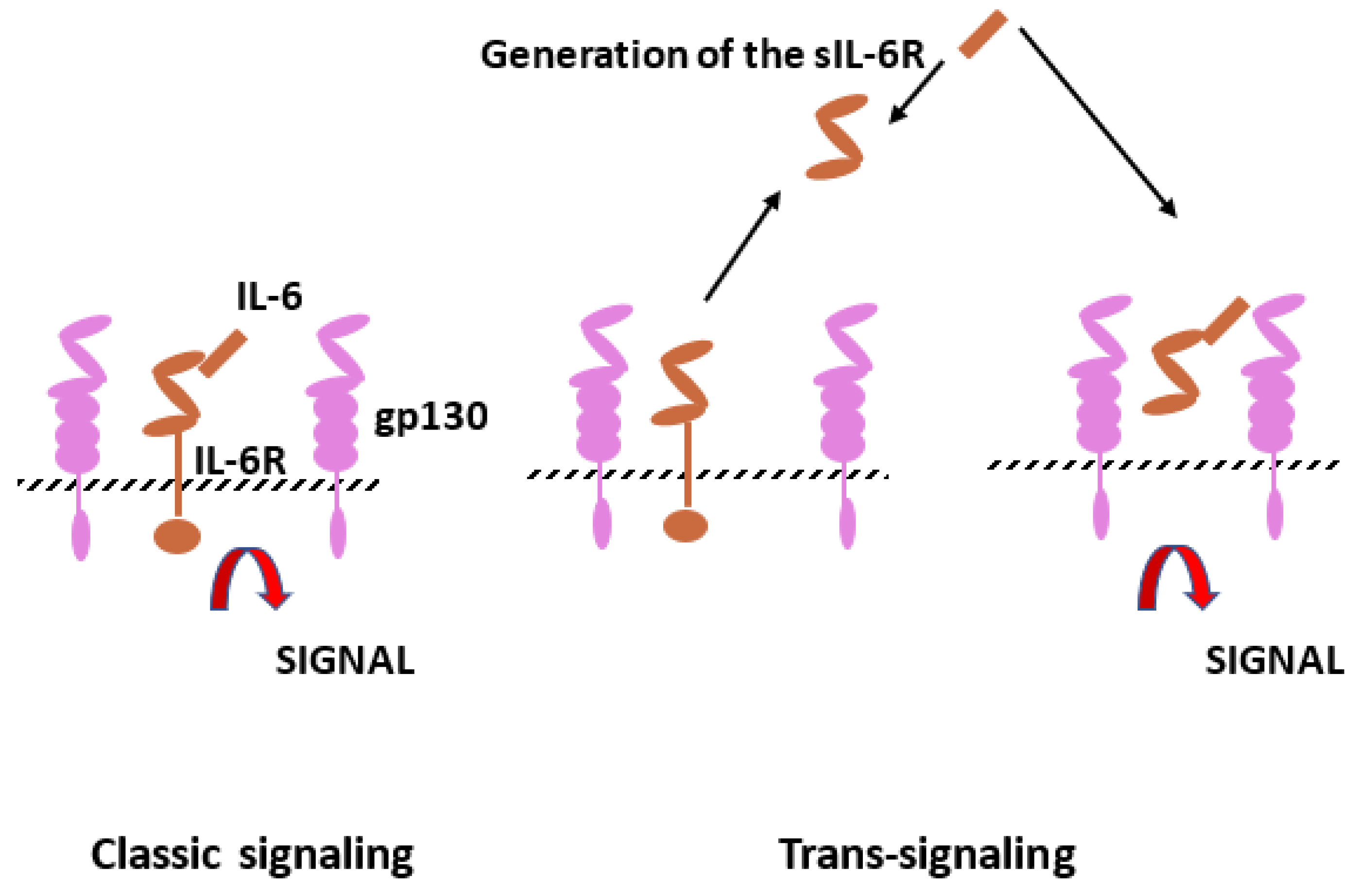
3.1. IL-1 and IL-6 Families
3.2. IL-1 and IL-6 Inhibitors
3.3. TNF Family
3.4. TNF Inhibitors
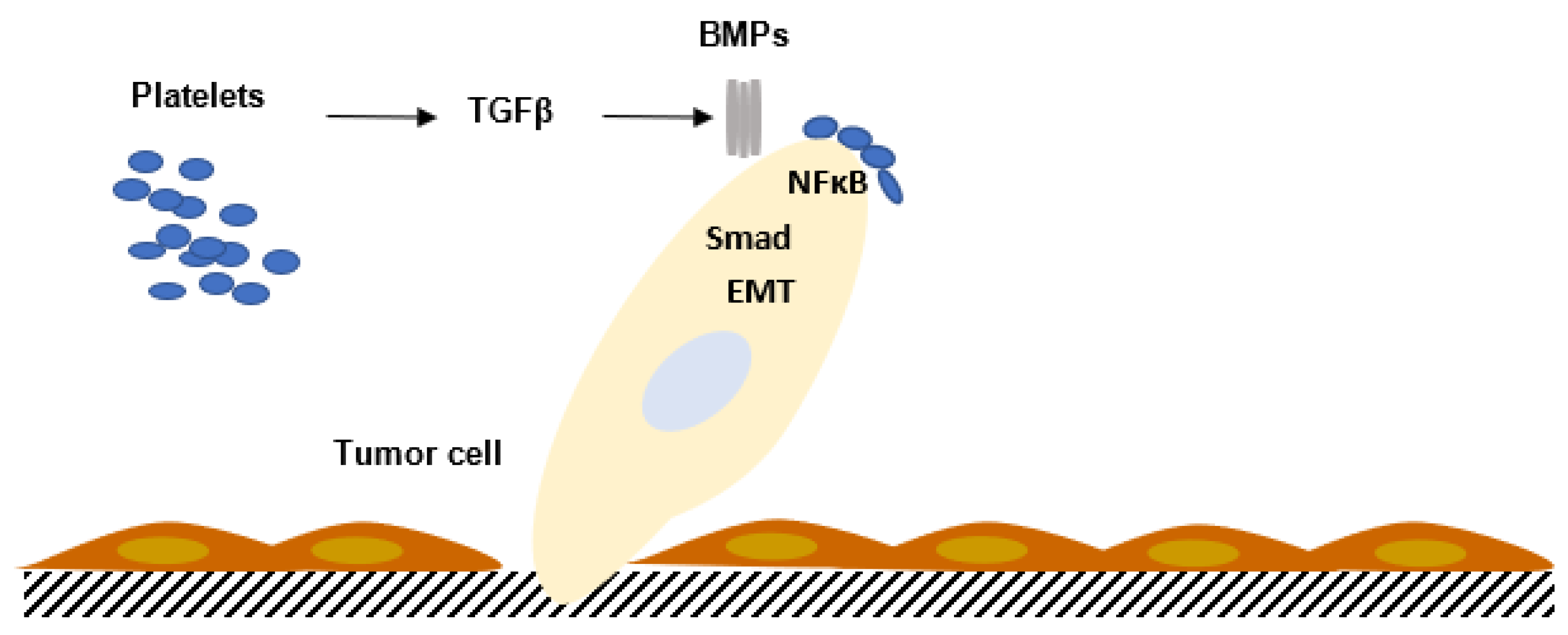
3.5. Transforming Growth Factor Family
3.6. Interferon Family
4. Cytokine Production in Human Pathology
4.1. Pathophysiology
4.2. Inflammatory Anemia
4.3. Cytokine Storm
5. Hematopoietic Growth Factors
5.1. Erythropoietin (EPO)
5.2. The Leukocyte-Stimulating Factors
5.3. Thrombopoietin
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Porter, R.R. Structural studies of immunoglobulins. Science 1973, 180, 713–716. [Google Scholar] [CrossRef] [PubMed]
- Rather, L.J. Disturbance of function (functio laesa): The legendary fifth cardinal sign of inflammation, added by Galen to the four cardinal signs of Celsus. Bull. N. Y. Acad. Med. 1971, 47, 303–322. [Google Scholar] [PubMed]
- Wautier, J.L.; Wautier, M.P. Old and New Blood Markers in Human Colorectal Cancer. Int. J. Mol. Sci. 2022, 23, 12968. [Google Scholar] [CrossRef] [PubMed]
- Maddipati, K.R. Non-inflammatory Physiology of “Inflammatory” Mediators—Unalamation, a New Paradigm. Front. Immunol. 2020, 11, 580117. [Google Scholar] [CrossRef]
- Wautier, J.L.; Wautier, M.P. Cellular and Molecular Aspects of Blood Cell-Endothelium Interactions in Vascular Disorders. Int. J. Mol. Sci. 2020, 21, 5315. [Google Scholar] [CrossRef]
- Dennis, E.A.; Cao, J.; Hsu, Y.H.; Magrioti, V.; Kokotos, G. Phospholipase A2 enzymes: Physical structure, biological function, disease implication, chemical inhibition, and therapeutic intervention. Chem. Rev. 2011, 111, 6130–6185. [Google Scholar] [CrossRef]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef]
- Morteau, O.; Morham, S.G.; Sellon, R.; Dieleman, L.A.; Langenbach, R.; Smithies, O.; Sartor, R.B. Impaired mucosal defense to acute colonic injury in mice lacking cyclooxygenase-1 or cyclooxygenase-2. J. Clin. Investig. 2000, 105, 469–478. [Google Scholar] [CrossRef]
- Funk, C.D. Prostaglandins and leukotrienes: Advances in eicosanoid biology. Science 2001, 294, 1871–1875. [Google Scholar] [CrossRef]
- Legler, D.F.; Bruckner, M.; Uetz-von Allmen, E.; Krause, P. Prostaglandin E2 at new glance: Novel insights in functional diversity offer therapeutic chances. Int. J. Biochem. Cell Biol. 2010, 42, 198–201. [Google Scholar] [CrossRef]
- Montine, T.J.; Milatovic, D.; Gupta, R.C.; Valyi-Nagy, T.; Morrow, J.D.; Breyer, R.M. Neuronal oxidative damage from activated innate immunity is EP2 receptor-dependent. J. Neurochem. 2002, 83, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Reinold, H.; Ahmadi, S.; Depner, U.B.; Layh, B.; Heindl, C.; Hamza, M.; Pahl, A.; Brune, K.; Narumiya, S.; Müller, U.; et al. Spinal inflammatory hyperalgesia is mediated by prostaglandin E receptors of the EP2 subtype. J. Clin. Investig. 2005, 115, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Brenneis, C.; Coste, O.; Schmidt, R.; Angioni, C.; Popp, L.; Nüsing, R.M.; Becker, W.; Scholich, K.; Geisslinger, G. Consequences of altered eicosanoid patterns for nociceptive processing in mPGES-1-deficient mice. J. Cell. Mol. Med. 2008, 12, 639–648. [Google Scholar] [CrossRef]
- Park, J.Y.; Pillinger, M.H.; Abramson, S.B. Prostaglandin E2 synthesis and secretion: The role of PGE2 synthases. Clin. Immunol. 2006, 119, 229–240. [Google Scholar] [CrossRef]
- Yao, C.; Sakata, D.; Esaki, Y.; Li, Y.; Matsuoka, T.; Kuroiwa, K.; Sugimoto, Y.; Narumiya, S. Prostaglandin E2-EP4 signaling promotes immune inflammation through Th1 cell differentiation and Th17 cell expansion. Nat. Med. 2009, 15, 633–640. [Google Scholar] [CrossRef]
- Wu, K.K.; Liou, J.Y. Cellular and molecular biology of prostacyclin synthase. Biochem. Biophys. Res. Commun. 2005, 338, 45–52. [Google Scholar] [CrossRef]
- Urade, Y.; Ujihara, M.; Horiguchi, Y.; Ikai, K.; Hayaishi, O. The major source of endogenous prostaglandin D2 production is likely antigen-presenting cells. Localization of glutathione-requiring prostaglandin D synthetase in histiocytes, dendritic, and Kupffer cells in various rat tissues. J. Immunol. 1989, 143, 2982–2989. [Google Scholar] [CrossRef]
- Herlong, J.L.; Scott, T.R. Positioning prostanoids of the D and J series in the immunopathogenic scheme. Immunol. Lett. 2006, 102, 121–131. [Google Scholar] [CrossRef]
- Fitzpatrick, F.A.; Wynalda, M.A. Albumin-catalyzed metabolism of prostaglandin D2. Identification of products formed in vitro. J. Biol. Chem. 1983, 258, 11713–11718. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Kondo, M.; Osawa, T.; Shibata, N.; Kobayashi, M.; Uchida, K. 15-deoxy-delta 12,14-prostaglandin J2. A prostaglandin D2 metabolite generated during inflammatory processes. J. Biol. Chem. 2002, 277, 10459–10466. [Google Scholar] [CrossRef] [PubMed]
- Forman, B.M.; Tontonoz, P.; Chen, J.; Brun, R.P.; Spiegelman, B.M.; Evans, R.M. 15-Deoxy-delta 12, 14-prostaglandin J2 is a ligand for the adipocyte determination factor PPAR gamma. Cell 1995, 83, 803–812. [Google Scholar] [CrossRef]
- Roberts, L.J., 2nd; Sweetman, B.J.; Lewis, R.A.; Austen, K.F.; Oates, J.A. Increased production of prostaglandin D2 in patients with systemic mastocytosis. N. Engl. J. Med. 1980, 303, 1400–1404. [Google Scholar] [CrossRef]
- Hammad, H.; de Heer, H.J.; Soullie, T.; Hoogsteden, H.C.; Trottein, F.; Lambrecht, B.N. Prostaglandin D2 inhibits airway dendritic cell migration and function in steady state conditions by selective activation of the D prostanoid receptor 1. J. Immunol. 2003, 171, 3936–3940. [Google Scholar] [CrossRef]
- Nagoshi, H.; Uehara, Y.; Kanai, F.; Maeda, S.; Ogura, T.; Goto, A.; Toyo-Oka, T.; Esumi, H.; Shimizu, T.; Omata, M. Prostaglandin D2 inhibits inducible nitric oxide synthase expression in rat vascular smooth muscle cells. Circ. Res. 1998, 82, 204–209. [Google Scholar] [CrossRef]
- Negoro, H.; Shin, W.S.; Hakamada-Taguchi, R.; Eguchi, N.; Urade, Y.; Goto, A.; Toyo-Oka, T.; Fujita, T.; Omata, M.; Uehara, Y. Endogenous prostaglandin D2 synthesis reduces an increase in plasminogen activator inhibitor-1 following interleukin stimulation in bovine endothelial cells. J. Hypertens. 2002, 20, 1347–1354. [Google Scholar] [CrossRef] [PubMed]
- Vane, J.R. Inhibition of prostaglandin synthesis as a mechanism of action for aspirin-like drugs. Nat. New Biol. 1971, 231, 232–235. [Google Scholar] [CrossRef]
- Kulesza, A.; Paczek, L.; Burdzinska, A. The Role of COX-2 and PGE2 in the Regulation of Immunomodulation and Other Functions of Mesenchymal Stromal Cells. Biomedicines 2023, 11, 445. [Google Scholar] [CrossRef] [PubMed]
- Moncada, S.; Higgs, E.A. Molecular mechanisms and therapeutic strategies related to nitric oxide. FASEB J. 1995, 9, 1319–1330. [Google Scholar] [CrossRef] [PubMed]
- Jo-Watanabe, A.; Okuno, T.; Yokomizo, T. The Role of Leukotrienes as Potential Therapeutic Targets in Allergic Disorders. Int. J. Mol. Sci. 2019, 20, 3580. [Google Scholar] [CrossRef]
- Back, M. Leukotriene signaling in atherosclerosis and ischemia. Cardiovasc. Drugs Ther. 2009, 23, 41–48. [Google Scholar] [CrossRef]
- Fajgenbaum, D.C.; June, C.H. Cytokine Storm. N. Engl. J. Med. 2020, 383, 2255–2273. [Google Scholar] [CrossRef]
- Hu, D. Role of Anti-inflammatory Cytokines IL-35 and IL-37 in Asthma. Inflammation 2017, 40, 697–707. [Google Scholar] [CrossRef]
- Mora, J.; Weigert, A. IL-1 family cytokines in cancer immunity—A matter of life and death. Biol. Chem. 2016, 397, 1125–1134. [Google Scholar] [CrossRef] [PubMed]
- Palomo, J.; Dietrich, D.; Martin, P.; Palmer, G.; Gabay, C. The interleukin (IL)-1 cytokine family—Balance between agonists and antagonists in inflammatory diseases. Cytokine 2015, 76, 25–37. [Google Scholar] [CrossRef]
- Mantovani, A.; Dinarello, C.A.; Molgora, M.; Garlanda, C. Interleukin-1 and Related Cytokines in the Regulation of Inflammation and Immunity. Immunity 2019, 50, 778–795. [Google Scholar] [CrossRef] [PubMed]
- Rose-John, S. Interleukin-6 Family Cytokines. Cold Spring Harb. Perspect. Biol. 2018, 10, a028415. [Google Scholar] [CrossRef] [PubMed]
- Scheller, J.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S. The pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochim. Biophys. Acta 2011, 1813, 878–888. [Google Scholar] [CrossRef] [PubMed]
- Naka, T.; Nishimoto, N.; Kishimoto, T. The paradigm of IL-6: From basic science to medicine. Arthritis Res. 2002, 4 (Suppl. 3), S233–S242. [Google Scholar] [CrossRef]
- Dienz, O.; Eaton, S.M.; Bond, J.P.; Neveu, W.; Moquin, D.; Noubade, R.; Briso, E.M.; Charland, C.; Leonard, W.J.; Ciliberto, G.; et al. The induction of antibody production by IL-6 is indirectly mediated by IL-21 produced by CD4+ T cells. J. Exp. Med. 2009, 206, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Liote, F.; Boval-Boizard, B.; Weill, D.; Kuntz, D.; Wautier, J.L. Blood monocyte activation in rheumatoid arthritis: Increased monocyte adhesiveness, integrin expression, and cytokine release. Clin. Exp. Immunol. 1996, 106, 13–19. [Google Scholar] [CrossRef]
- Su, D.L.; Lu, Z.M.; Shen, M.N.; Li, X.; Sun, L.Y. Roles of pro- and anti-inflammatory cytokines in the pathogenesis of SLE. J. Biomed. Biotechnol. 2012, 2012, 347141. [Google Scholar] [CrossRef] [PubMed]
- Lindenlaub, T.; Sommer, C. Cytokines in sural nerve biopsies from inflammatory and non-inflammatory neuropathies. Acta Neuropathol. 2003, 105, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Interleukin-1 in the pathogenesis and treatment of inflammatory diseases. Blood 2011, 117, 3720–3732. [Google Scholar] [CrossRef]
- Zizzo, G.; Tamburello, A.; Castelnovo, L.; Laria, A.; Mumoli, N.; Faggioli, P.M.; Stefani, I.; Mazzone, A. Immunotherapy of COVID-19: Inside and Beyond IL-6 Signalling. Front. Immunol. 2022, 13, 795315. [Google Scholar] [CrossRef] [PubMed]
- Ridker, P.M.; Rane, M. Interleukin-6 Signaling and Anti-Interleukin-6 Therapeutics in Cardiovascular Disease. Circ. Res. 2021, 128, 1728–1746. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Gupta, S.C.; Kim, J.H. Historical perspectives on tumor necrosis factor and its superfamily: 25 years later, a golden journey. Blood 2012, 119, 651–665. [Google Scholar] [CrossRef]
- Berk, B.C.; Abe, J.I.; Min, W.; Surapisitchat, J.; Yan, C. Endothelial atheroprotective and anti-inflammatory mechanisms. Ann. N. Y. Acad. Sci. 2001, 947, 93–109; discussion 109–110. [Google Scholar] [CrossRef]
- Natoli, G.; Costanzo, A.; Moretti, F.; Fulco, M.; Balsano, C.; Levrero, M. Tumor necrosis factor (TNF) receptor 1 signaling downstream of TNF receptor-associated factor 2. Nuclear factor kappaB (NFkappaB)-inducing kinase requirement for activation of activating protein 1 and NFkappaB but not of c-Jun N-terminal kinase/stress-activated protein kinase. J. Biol. Chem. 1997, 272, 26079–26082. [Google Scholar]
- Boyce, E.G.; Halilovic, J.; Stan-Ugbene, O. Golimumab: Review of the efficacy and tolerability of a recently approved tumor necrosis factor-alpha inhibitor. Clin. Ther. 2010, 32, 1681–1703. [Google Scholar] [CrossRef]
- Morikawa, M.; Derynck, R.; Miyazono, K. TGF-beta and the TGF-beta Family: Context-Dependent Roles in Cell and Tissue Physiology. Cold Spring Harb. Perspect. Biol. 2016, 8, a021873. [Google Scholar] [CrossRef]
- Moses, H.L.; Yang, E.Y.; Pietenpol, J.A. TGF-beta stimulation and inhibition of cell proliferation: New mechanistic insights. Cell 1990, 63, 245–247. [Google Scholar] [CrossRef] [PubMed]
- Jian, H.; Shen, X.; Liu, I.; Semenov, M.; He, X.; Wang, X.F. Smad3-dependent nuclear translocation of beta-catenin is required for TGF-beta1-induced proliferation of bone marrow-derived adult human mesenchymal stem cells. Genes Dev. 2006, 20, 666–674. [Google Scholar] [CrossRef] [PubMed]
- Krieglstein, K.; Zheng, F.; Unsicker, K.; Alzheimer, C. More than being protective: Functional roles for TGF-beta/activin signaling pathways at central synapses. Trends Neurosci. 2011, 34, 421–429. [Google Scholar] [CrossRef]
- Walter, M.R. The Role of Structure in the Biology of Interferon Signaling. Front. Immunol. 2020, 11, 606489. [Google Scholar] [CrossRef] [PubMed]
- Pestka, S.; Krause, C.D.; Sarkar, D.; Walter, M.R.; Shi, Y.; Fisher, P.B. Interleukin-10 and related cytokines and receptors. Annu. Rev. Immunol. 2004, 22, 929–979. [Google Scholar] [CrossRef]
- Ouyang, W.; Rutz, S.; Crellin, N.K.; Valdez, P.A.; Hymowitz, S.G. Regulation and functions of the IL-10 family of cytokines in inflammation and disease. Annu. Rev. Immunol. 2011, 29, 71–109. [Google Scholar] [CrossRef]
- Muhl, H.; Pfeilschifter, J. Anti-inflammatory properties of pro-inflammatory interferon-gamma. Int. Immunopharmacol. 2003, 3, 1247–1255. [Google Scholar] [CrossRef]
- Opal, S.M.; DePalo, V.A. Anti-inflammatory cytokines. Chest 2000, 117, 1162–1172. [Google Scholar] [CrossRef]
- Chen, C.; Xu, H.; Peng, Y.; Luo, H.; Huang, G.X.; Wu, X.J.; Dai, Y.C.; Luo, H.L.; Zhang, J.A.; Zheng, B.Y.; et al. Elevation in the counts of IL-35-producing B cells infiltrating into lung tissue in mycobacterial infection is associated with the downregulation of Th1/Th17 and upregulation of Foxp3(+)Treg. Sci. Rep. 2020, 10, 13212. [Google Scholar] [CrossRef]
- Fraenkel, P.G. Anemia of Inflammation. Med. Clin. N. Am. 2016, 101, 285–296. [Google Scholar] [CrossRef]
- Nemeth, E.; Ganz, T. Anemia of inflammation. Hematol. Oncol. Clin. N. Am. 2014, 28, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.W.; Gardner, R.A.; Porter, D.L.; Louis, C.U.; Ahmed, N.; Jensen, M.C.; Grupp, S.A.; Mackall, C.L. Current concepts in the diagnosis and management of cytokine release syndrome. Blood 2014, 124, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Grupp, S.A.; Kalos, M.; Barrett, D.; Aplenc, R.; Porter, D.L.; Rheingold, S.R.; Teachey, D.T.; Chew, A.; Hauck, B.; Wright, J.F.; et al. Chimeric antigen receptor-modified T cells for acute lymphoid leukemia. N. Engl. J. Med. 2013, 368, 1509–1518. [Google Scholar] [CrossRef]
- Rossert, J.; Froissart, M.; Jacquot, C. Anemia management and chronic renal failure progression. Kidney Int. Suppl. 2005, 68, S76–S81. [Google Scholar] [CrossRef] [PubMed]
- Shander, A.; Kaufman, M.; Goodnough, L.T. How I treat anemia in the perisurgical setting. Blood 2020, 136, 814–822. [Google Scholar] [CrossRef]
- Metcalf, D. The colony-stimulating factors and cancer. Cancer Immunol. Res. 2013, 1, 351–356. [Google Scholar] [CrossRef]
- Ghanima, W.; Cooper, N.; Rodeghiero, F.; Godeau, B.; Bussel, J.B. Thrombopoietin receptor agonists: Ten years later. Haematologica 2019, 104, 1112–1123. [Google Scholar] [CrossRef]
- Erickson-Miller, C.L.; Delorme, E.; Tian, S.-S.; Hopson, C.B.; Landis, A.J.; Valoret, E.I.; Sellers, T.S.; Rosen, J.; Miller, S.G.; Luengo, J.I.; et al. Preclinical activity of eltrombopag (SB-497115), an oral, nonpeptide thrombopoietin receptor agonist. Stem Cells 2009, 27, 424–430. [Google Scholar] [CrossRef]
- Broudy, V.C.; Lin, N.L. AMG531 stimulates megakaryopoiesis in vitro by binding to Mpl. Cytokine 2004, 25, 52–60. [Google Scholar] [CrossRef]
- Ridker, P.M.; Everett, B.M.; Thuren, T.; MacFadyen, J.G.; Chang, W.H.; Ballantyne, C.; Fonseca, F.; Nicolau, J.; Koenig, W.; Anker, S.D.; et al. Antiinflammatory Therapy with Canakinumab for Atherosclerotic Disease. N. Engl. J. Med. 2017, 377, 1119–1131. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Interleukin-1 (IL-1) Family [34,35] | |||
| Cytokines | Pro-Infl | Anti-Infl | Cell Source |
| IL-1α | + | Epithelial, endothelial, stromal cells | |
| IL-1β | + | Monocytes/macrophages | |
| IL-18 | + | Kupffer cells, intestinal epithelial cells | |
| IL-33 | + | Keratinocytes, endothelial cells, epithelial cells, fibroblast-like cells | |
| IL-36α | + | Epithelial cells | |
| IL-36β | + | Epithelial cells | |
| IL-36γ | + | Epithelial cells | |
| Interleukin-6 (IL-6) Family [36] | |||
| Cytokines | Pro-Infl | Anti-Infl | Cell Source |
| IL-6 | + | + | B cells, T cells, monocytes, fibroblasts, endothelial cells |
| IL-11 | + | Stroma cells, fibroblasts | |
| CNF | + | + | Astrocytes |
| LIF | + | + | Pleiotropic |
| OSM | + | + | Pleiotropic |
| CT-1 | + | Myocytes | |
| CLC | + | + | Lymphocytes |
| IL-27 | + | T cells | |
| IL-31 | + | Monocytes/macrophages, dendritic cells, T cells | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wautier, J.-L.; Wautier, M.-P. Pro- and Anti-Inflammatory Prostaglandins and Cytokines in Humans: A Mini Review. Int. J. Mol. Sci. 2023, 24, 9647. https://doi.org/10.3390/ijms24119647
Wautier J-L, Wautier M-P. Pro- and Anti-Inflammatory Prostaglandins and Cytokines in Humans: A Mini Review. International Journal of Molecular Sciences. 2023; 24(11):9647. https://doi.org/10.3390/ijms24119647
Chicago/Turabian StyleWautier, Jean-Luc, and Marie-Paule Wautier. 2023. "Pro- and Anti-Inflammatory Prostaglandins and Cytokines in Humans: A Mini Review" International Journal of Molecular Sciences 24, no. 11: 9647. https://doi.org/10.3390/ijms24119647
APA StyleWautier, J.-L., & Wautier, M.-P. (2023). Pro- and Anti-Inflammatory Prostaglandins and Cytokines in Humans: A Mini Review. International Journal of Molecular Sciences, 24(11), 9647. https://doi.org/10.3390/ijms24119647