

Preconditioned Chorionic Villus Mesenchymal Stem/Stromal Cells (CVMSCs) Minimize the Invasive Phenotypes of Breast Cancer Cell Line MDA231 In Vitro

Abstract
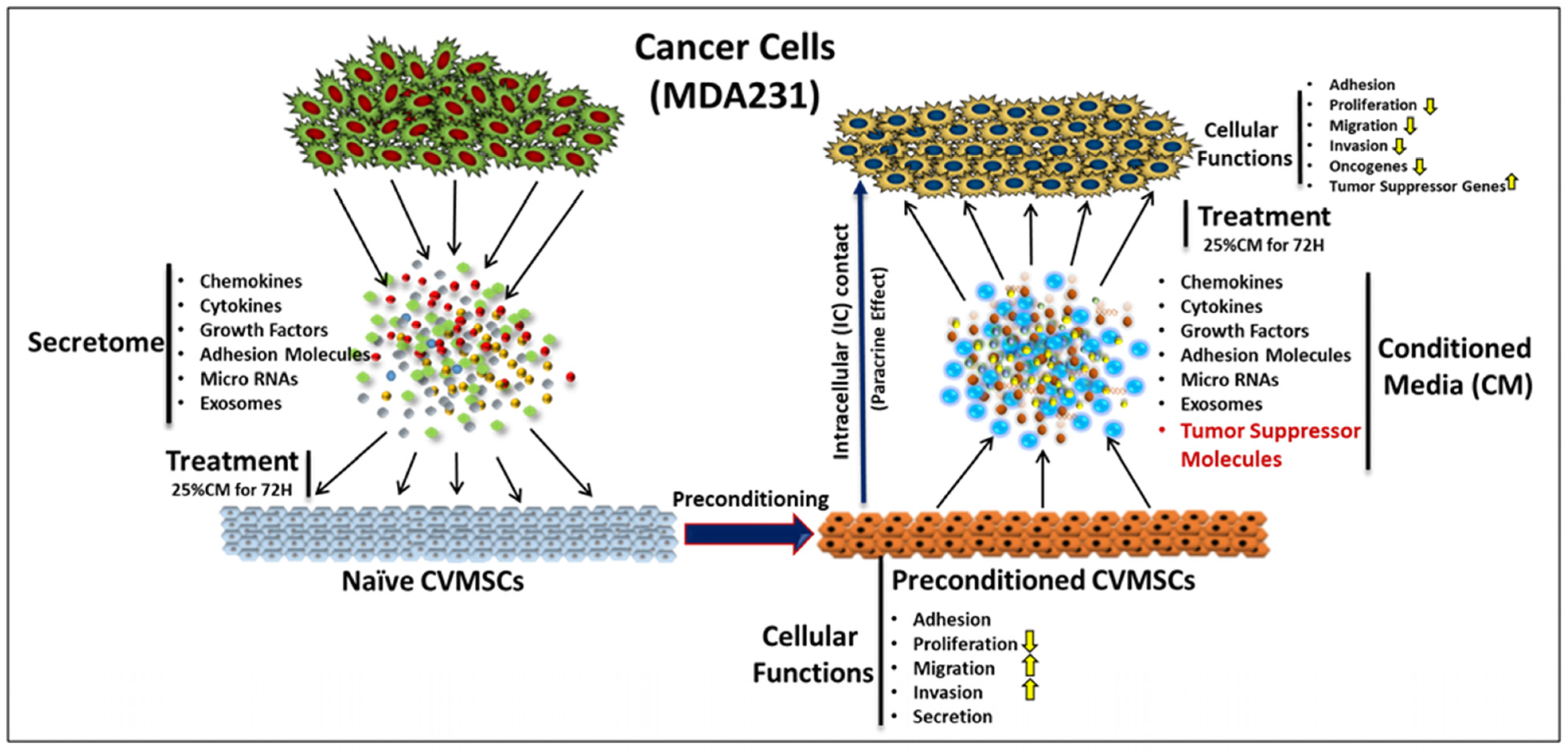
1. Introduction
2. Results
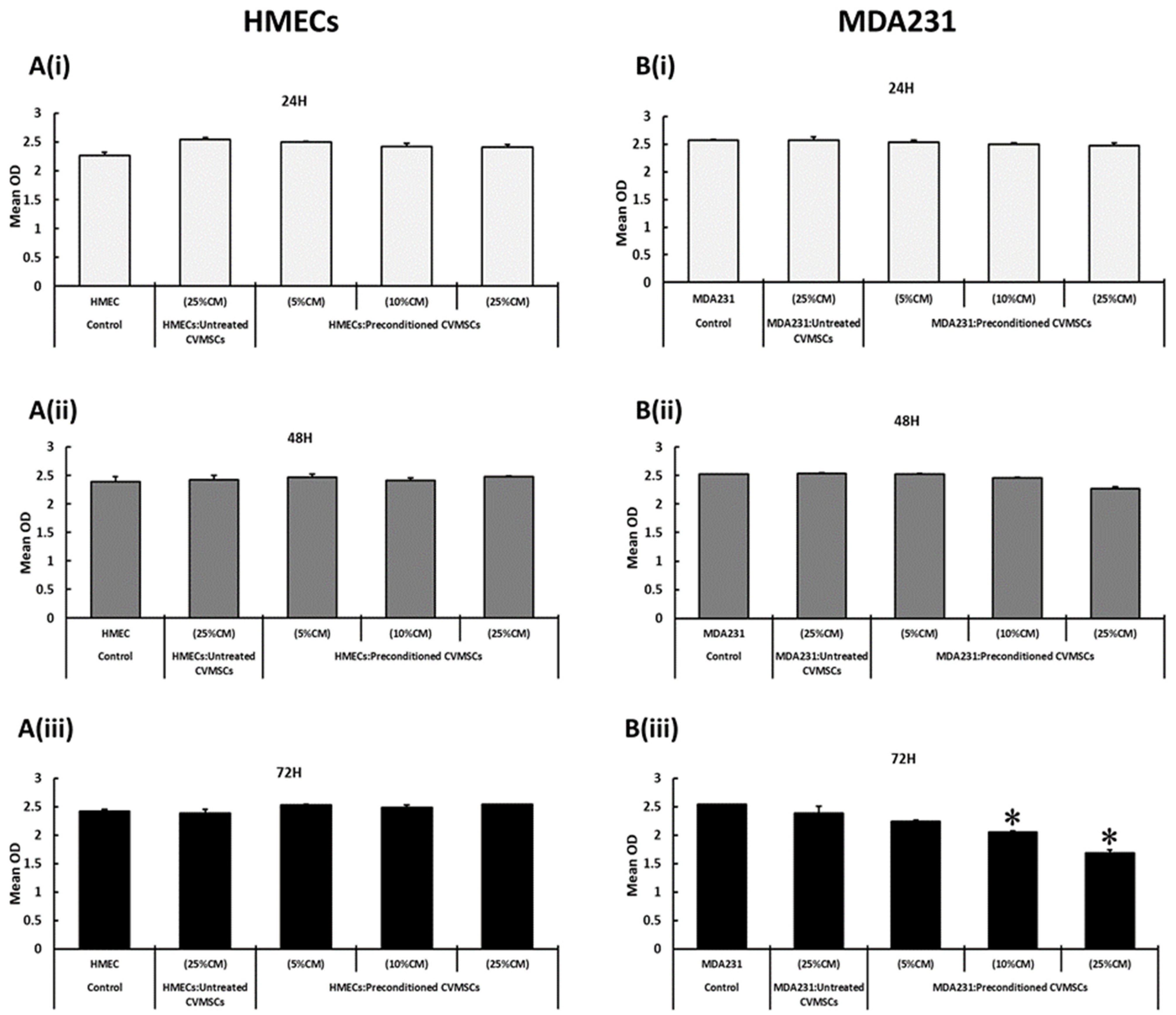
2.1. The Standardization of CM Concentration, Cellular Ratios, and Preconditioning Time
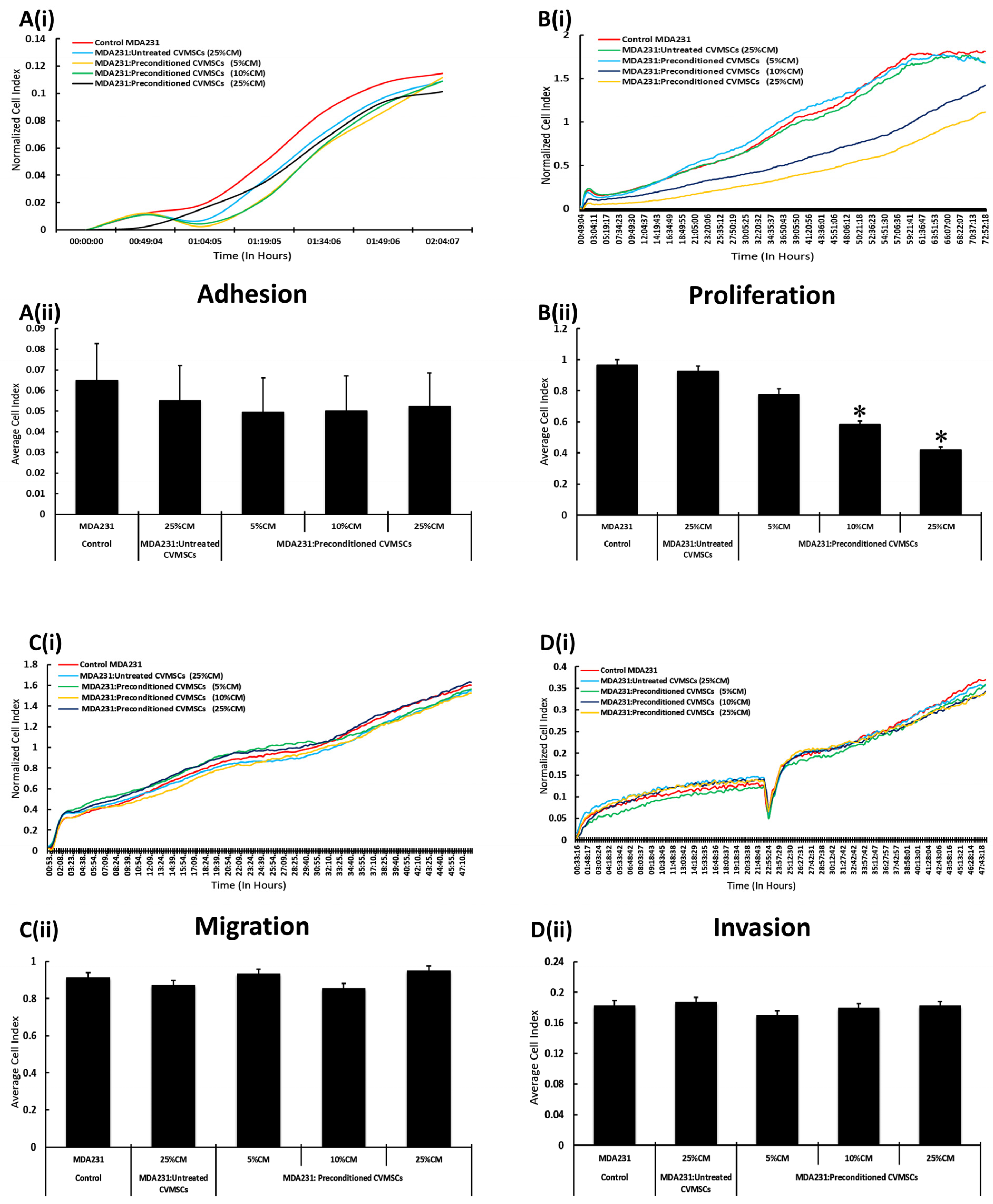
2.2. CM-CVMSCs Enhance Proliferation but No Other Phenotype of MDA231 Cells
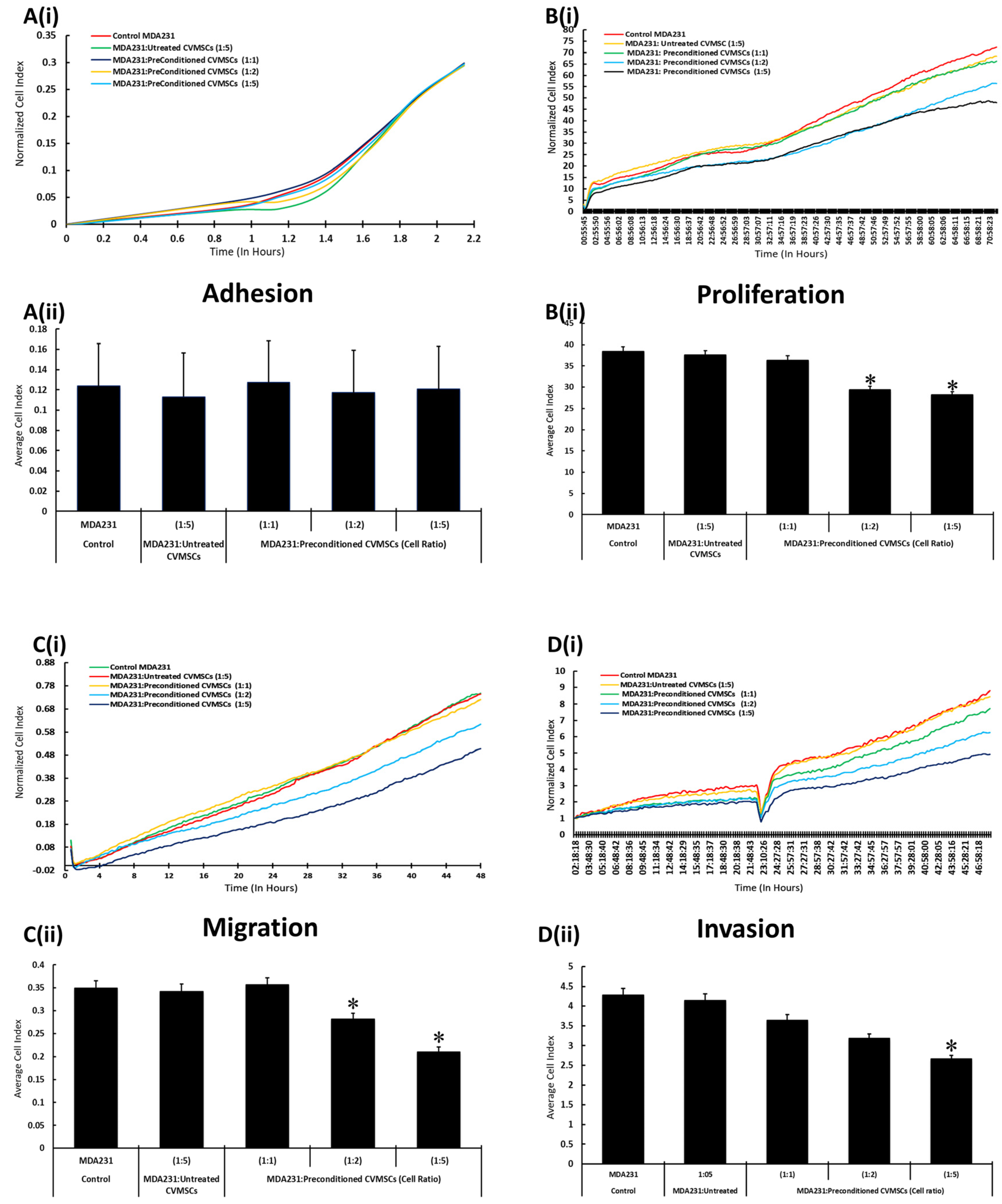
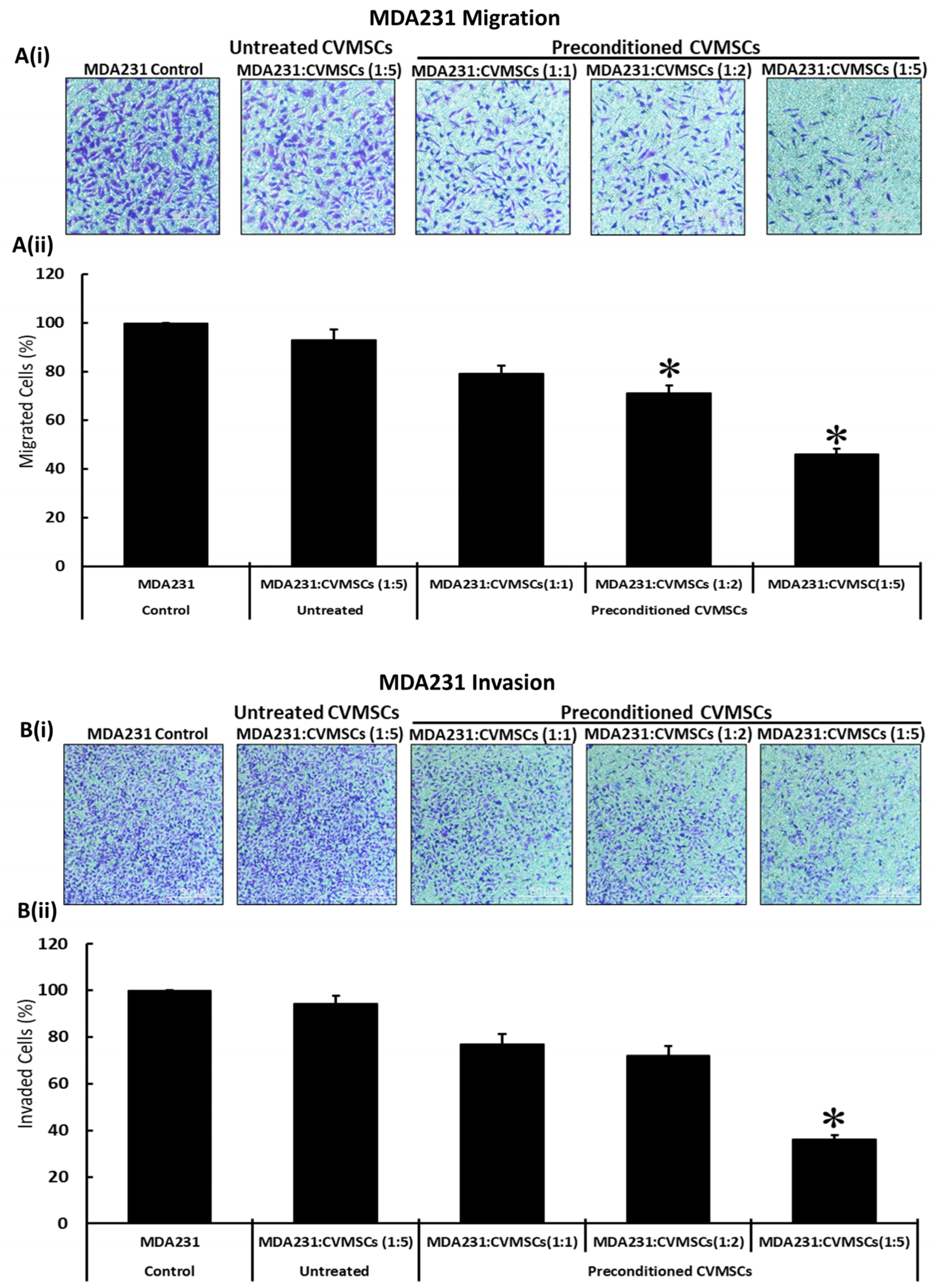
2.3. Preconditioned CVMSCs Suppress the Invasive Phenotype of MDA231 Cells
2.4. CVMSCs Modulate the Expression of Genes Responsible for Breast Cancer Oncogenesis and Metastasis in MDA231 Cells
3. Discussion
4. Materials and Methods
4.1. Ethical Approval and Placenta Collection
4.2. Reagents and Cell Lines
4.3. Isolation, Culture, and Maintenance of CVMSCs and Human Umbilical Vein Endothelial Cells (HUVECs)
4.4. Conditioned Media (CM) Collection and CVMSCs Preconditioning
4.5. MDA231 and HMEC Treatment with CM and Preconditioned CVMSCs
4.6. MTS Cell Proliferation Assay
4.7. Real Time Cell Analysis (RTCA) for Cellular Functions
4.8. In Vitro Cellular Migration and Invasion Assays
4.9. Flow Cytometry
4.10. RNA Isolation and Real-Time PCR (RT-PCR)
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Goffin, J.; Lacchetti, C.; Ellis, P.M.; Ung, Y.C.; Evans, W.K. First-Line Systemic Chemotherapy in the Treatment of Advanced Non-Small Cell Lung Cancer: A Systematic Review. J. Thorac. Oncol. 2010, 5, 260–274. [Google Scholar] [CrossRef]
- Chen, Z.; Fillmore, C.M.; Hammerman, P.S.; Kim, C.F.; Wong, K.-K. Non-Small-Cell Lung Cancers: A Heterogeneous Set of Diseases. Nat. Rev. Cancer 2014, 14, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Sermer, D.; Brentjens, R. CAR T-Cell Therapy: Full Speed Ahead. Hematol. Oncol. 2019, 37, 95–100. [Google Scholar] [CrossRef]
- Sipp, D.; Robey, P.G.; Turner, L. Clear up This Stem-Cell Mess. Nature 2018, 561, 455–457. [Google Scholar] [CrossRef]
- Gomes, J.P.A.; Assoni, A.F.; Pelatti, M.; Coatti, G.; Okamoto, O.K.; Zatz, M. Deepening a Simple Question: Can MSCs Be Used to Treat Cancer? Anticancer Res. 2017, 37, 4747–4758. [Google Scholar] [CrossRef]
- Javan, M.R.; Khosrojerdi, A.; Moazzeni, S.M. New Insights Into Implementation of Mesenchymal Stem Cells in Cancer Therapy: Prospects for Anti-Angiogenesis Treatment. Front. Oncol. 2019, 9, 840. [Google Scholar] [CrossRef]
- Orbay, H.; Tobita, M.; Mizuno, H. Mesenchymal Stem Cells Isolated from Adipose and Other Tissues: Basic Biological Properties and Clinical Applications. Stem Cells Int. 2012, 2012, 461718. [Google Scholar] [CrossRef] [PubMed]
- Phinney, D.G.; Sensebé, L. Mesenchymal Stromal Cells: Misconceptions and Evolving Concepts. Cytotherapy 2013, 15, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Oswald, J.; Boxberger, S.; Jørgensen, B.; Feldmann, S.; Ehninger, G.; Bornhäuser, M.; Werner, C. Mesenchymal Stem Cells Can Be Differentiated into Endothelial Cells in Vitro. Stem Cells 2004, 22, 377–384. [Google Scholar] [CrossRef]
- Atiya, H.; Frisbie, L.; Pressimone, C.; Coffman, L. Mesenchymal Stem Cells in the Tumor Microenvironment. Adv. Exp. Med. Biol. 2020, 1234, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Xu, Z.; Zhao, T.; Zhao, Z.; Shi, M.; Zhao, R.C.; Ye, L.; Zhang, X. Suppression of Tumorigenesis by Human Mesenchymal Stem Cells in a Hepatoma Model. Cell Res. 2008, 18, 500–507. [Google Scholar] [CrossRef]
- Maestroni, G.J.; Hertens, E.; Galli, P. Factor(s) from Nonmacrophage Bone Marrow Stromal Cells Inhibit Lewis Lung Carcinoma and B16 Melanoma Growth in Mice. Cell. Mol. Life Sci. 1999, 55, 663–667. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Ito, Y.; Kawano, Y.; Kurozumi, K.; Kobune, M.; Tsuda, H.; Bizen, A.; Honmou, O.; Niitsu, Y.; Hamada, H. Antitumor Effect of Genetically Engineered Mesenchymal Stem Cells in a Rat Glioma Model. Gene Ther. 2004, 11, 1155–1164. [Google Scholar] [CrossRef]
- Qiao, C.; Xu, W.; Zhu, W.; Hu, J.; Qian, H.; Yin, Q.; Jiang, R.; Yan, Y.; Mao, F.; Yang, H.; et al. Human Mesenchymal Stem Cells Isolated from the Umbilical Cord. Cell Biol. Int. 2008, 32, 8–15. [Google Scholar] [CrossRef]
- Otsu, K.; Das, S.; Houser, S.D.; Quadri, S.K.; Bhattacharya, S.; Bhattacharya, J. Concentration-Dependent Inhibition of Angiogenesis by Mesenchymal Stem Cells. Blood 2009, 113, 4197–4205. [Google Scholar] [CrossRef]
- Gondi, C.S.; Veeravalli, K.K.; Gorantla, B.; Dinh, D.H.; Fassett, D.; Klopfenstein, J.D.; Gujrati, M.; Rao, J.S. Human Umbilical Cord Blood Stem Cells Show PDGF-D-Dependent Glioma Cell Tropism in Vitro and in Vivo. Neuro. Oncol. 2010, 12, 453–465. [Google Scholar] [CrossRef][Green Version]
- Akimoto, K.; Kimura, K.; Nagano, M.; Takano, S.; To’a Salazar, G.; Yamashita, T.; Ohneda, O. Umbilical Cord Blood-Derived Mesenchymal Stem Cells Inhibit, but Adipose Tissue-Derived Mesenchymal Stem Cells Promote, Glioblastoma Multiforme Proliferation. Stem Cells Dev. 2013, 22, 1370–1386. [Google Scholar] [CrossRef]
- Abomaray, F.M.; Al Jumah, M.A.; Alsaad, K.O.; Jawdat, D.; Al Khaldi, A.; Alaskar, A.S.; Al Harthy, S.; Al Subayyil, A.M.; Khatlani, T.; Alawad, A.O.; et al. Phenotypic and Functional Characterization of Mesenchymal Stem/Multipotent Stromal Cells from Decidua Basalis of Human Term Placenta. Stem Cells Int. 2016, 2016, 5184601. [Google Scholar] [CrossRef] [PubMed]
- Abumaree, M.H.; Al Jumah, M.A.; Kalionis, B.; Jawdat, D.; Al Khaldi, A.; AlTalabani, A.A.; Knawy, B.A. Phenotypic and Functional Characterization of Mesenchymal Stem Cells from Chorionic Villi of Human Term Placenta. Stem Cell Rev. Rep. 2013, 9, 16–31. [Google Scholar] [CrossRef]
- Abumaree, M.H.; Abomaray, F.M.; Alshehri, N.A.; Almutairi, A.; AlAskar, A.S.; Kalionis, B.; Al Jumah, M.A. Phenotypic and Functional Characterization of Mesenchymal Stem/Multipotent Stromal Cells From Decidua Parietalis of Human Term Placenta. Reprod. Sci. 2016, 23, 1193–1207. [Google Scholar] [CrossRef] [PubMed]
- Abumaree, M.H.; Abomaray, F.M.; Alshabibi, M.A.; AlAskar, A.S.; Kalionis, B. Immunomodulatory Properties of Human Placental Mesenchymal Stem/Stromal Cells. Placenta 2017, 59, 87–95. [Google Scholar] [CrossRef]
- Basmaeil, Y.; Al Subayyil, A.; Abumaree, M.; Khatlani, T. Conditions Mimicking the Cancer Microenvironment Modulate the Functional Outcome of Human Chorionic Villus Mesenchymal Stem/Stromal Cells in Vitro. Front. Cell Dev. Biol. 2021, 9, 650125. [Google Scholar] [CrossRef]
- Chapel, A.; Bertho, J.M.; Bensidhoum, M.; Fouillard, L.; Young, R.G.; Frick, J.; Demarquay, C.; Cuvelier, F.; Mathieu, E.; Trompier, F.; et al. Mesenchymal Stem Cells Home to Injured Tissues When Co-Infused with Hematopoietic Cells to Treat a Radiation-Induced Multi-Organ Failure Syndrome. J. Gene Med. 2003, 5, 1028–1038. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I. Why Are MSCs Therapeutic? New Data: New Insight. J. Pathol. 2009, 217, 318–324. [Google Scholar] [CrossRef]
- Le Blanc, K.; Mougiakakos, D. Multipotent Mesenchymal Stromal Cells and the Innate Immune System. Nat. Rev. Immunol. 2012, 12, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Singer, N.G.; Caplan, A.I. Mesenchymal Stem Cells: Mechanisms of Inflammation. Annu. Rev. Pathol. 2011, 6, 457–478. [Google Scholar] [CrossRef]
- Bronckaers, A.; Hilkens, P.; Martens, W.; Gervois, P.; Ratajczak, J.; Struys, T.; Lambrichts, I. Mesenchymal Stem/Stromal Cells as a Pharmacological and Therapeutic Approach to Accelerate Angiogenesis. Pharmacol. Ther. 2014, 143, 181–196. [Google Scholar] [CrossRef]
- Frenette, P.S.; Pinho, S.; Lucas, D.; Scheiermann, C. Mesenchymal Stem Cell: Keystone of the Hematopoietic Stem Cell Niche and a Stepping-Stone for Regenerative Medicine. Annu. Rev. Immunol. 2013, 31, 285–316. [Google Scholar] [CrossRef]
- Al Jumah, M.A.; Abumaree, M.H. The Immunomodulatory and Neuroprotective Effects of Mesenchymal Stem Cells (MSCs) in Experimental Autoimmune Encephalomyelitis (EAE): A Model of Multiple Sclerosis (MS). Int. J. Mol. Sci. 2012, 13, 9298–9331. [Google Scholar] [CrossRef]
- Abumaree, M.H.; Al Jumah, M.A.; Kalionis, B.; Jawdat, D.; Al Khaldi, A.; Abomaray, F.M.; Fatani, A.S.; Chamley, L.W.; Knawy, B.A. Human Placental Mesenchymal Stem Cells (PMSCs) Play a Role as Immune Suppressive Cells by Shifting Macrophage Differentiation from Inflammatory M1 to Anti-Inflammatory M2 Macrophages. Stem Cell Rev. Rep. 2013, 9, 620–641. [Google Scholar] [CrossRef]
- Abumaree, M.H.; Hakami, M.; Abomaray, F.M.; Alshabibi, M.A.; Kalionis, B.; Al Jumah, M.A.; AlAskar, A.S. Human Chorionic Villous Mesenchymal Stem/Stromal Cells Modify the Effects of Oxidative Stress on Endothelial Cell Functions. Placenta 2017, 59, 74–86. [Google Scholar] [CrossRef]
- Basmaeil, Y.S.; Al Subayyil, A.M.; Khatlani, T.; Bahattab, E.; Al-Alwan, M.; Abomaray, F.M.; Kalionis, B.; Alshabibi, M.A.; Alaskar, A.S.; Abumaree, M.H. Human Chorionic Villous Mesenchymal Stem/Stromal Cells Protect Endothelial Cells from Injury Induced by High Level of Glucose. Stem Cell Res. Ther. 2018, 9, 238. [Google Scholar] [CrossRef] [PubMed]
- Abumaree, M.H.; Alshehri, N.A.; Almotery, A.; Al Subayyil, A.M.; Bahattab, E.; Abomaray, F.M.; Khatlani, T.; Kalionis, B.; Jawdat, D.; El-Muzaini, M.F.; et al. Preconditioning Human Natural Killer Cells with Chorionic Villous Mesenchymal Stem Cells Stimulates Their Expression of Inflammatory and Anti-Tumor Molecules. Stem Cell Res. Ther. 2019, 10, 50. [Google Scholar] [CrossRef] [PubMed]
- Vicinanza, C.; Lombardi, E.; Da Ros, F.; Marangon, M.; Durante, C.; Mazzucato, M.; Agostini, F. Modified Mesenchymal Stem Cells in Cancer Therapy: A Smart Weapon Requiring Upgrades for Wider Clinical Applications. World J. Stem Cells 2022, 14, 54–75. [Google Scholar] [CrossRef] [PubMed]
- Krampera, M. Mesenchymal Stromal Cell “Licensing”: A Multistep Process. Leukemia 2011, 25, 1408–1414. [Google Scholar] [CrossRef]
- Xiao, Q.; Wang, S.; Tian, H.; Xin, L.; Zou, Z.; Hu, Y.; Chang, C.; Wang, X.; Yin, Q.; Zhang, X.; et al. TNF-α Increases Bone Marrow Mesenchymal Stem Cell Migration to Ischemic Tissues. Cell Biochem. Biophys. 2012, 62, 409–414. [Google Scholar] [CrossRef]
- Ponte, A.L.; Marais, E.; Gallay, N.; Langonné, A.; Delorme, B.; Hérault, O.; Charbord, P.; Domenech, J. The in Vitro Migration Capacity of Human Bone Marrow Mesenchymal Stem Cells: Comparison of Chemokine and Growth Factor Chemotactic Activities. Stem Cells 2007, 25, 1737–1745. [Google Scholar] [CrossRef]
- Li, M.; Zeng, L.; Liu, S.; Dangelmajer, S.; Kahlert, U.D.; Huang, H.; Han, Y.; Chi, X.; Zhu, M.; Lei, T. Transforming Growth Factor-β Promotes Homing and Therapeutic Efficacy of Human Mesenchymal Stem Cells to Glioblastoma. J. Neuropathol. Exp. Neurol. 2019, 78, 315–325. [Google Scholar] [CrossRef]
- Escobar, P.; Bouclier, C.; Serret, J.; Bièche, I.; Brigitte, M.; Caicedo, A.; Sanchez, E.; Vacher, S.; Vignais, M.-L.; Bourin, P.; et al. IL-1β Produced by Aggressive Breast Cancer Cells Is One of the Factors That Dictate Their Interactions with Mesenchymal Stem Cells through Chemokine Production. Oncotarget 2015, 6, 29034–29047. [Google Scholar] [CrossRef] [PubMed]
- Lieblein, J.C.; Ball, S.; Hutzen, B.; Sasser, A.K.; Lin, H.-J.; Huang, T.H.; Hall, B.M.; Lin, J. STAT3 Can Be Activated through Paracrine Signaling in Breast Epithelial Cells. BMC Cancer 2008, 8, 302. [Google Scholar] [CrossRef]
- Freund, A.; Chauveau, C.; Brouillet, J.-P.; Lucas, A.; Lacroix, M.; Licznar, A.; Vignon, F.; Lazennec, G. IL-8 Expression and Its Possible Relationship with Estrogen-Receptor-Negative Status of Breast Cancer Cells. Oncogene 2003, 22, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Bachmeier, B.E.; Nerlich, A.G.; Lichtinghagen, R.; Sommerhoff, C.P. Matrix Metalloproteinases (MMPs) in Breast Cancer Cell Lines of Different Tumorigenicity. Anticancer Res. 2001, 21, 3821–3828. [Google Scholar] [PubMed]
- Shen, J.; Xu, S.; Zhou, H.; Liu, H.; Jiang, W.; Hao, J.; Hu, Z. IL-1β Induces Apoptosis and Autophagy via Mitochondria Pathway in Human Degenerative Nucleus Pulposus Cells. Sci. Rep. 2017, 7, 41067. [Google Scholar] [CrossRef]
- Fabbi, M.; Carbotti, G.; Ferrini, S. Dual Roles of IL-27 in Cancer Biology and Immunotherapy. Mediat. Inflamm. 2017, 2017, 3958069. [Google Scholar] [CrossRef]
- Zhuang, W.Z.; Lin, Y.H.; Su, L.J.; Wu, M.S.; Jeng, H.Y.; Chang, H.C.; Huang, Y.H.; Ling, T.Y. Mesenchymal Stem/Stromal Cell-Based Therapy: Mechanism, Systemic Safety and Biodistribution for Precision Clinical Applications. J. Biomed. Sci. 2021, 28, 28. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Shao, J.-Z.; Xiang, L.-X.; Dong, X.-J.; Zhang, G.-R. Mesenchymal Stem Cells: A Promising Candidate in Regenerative Medicine. Int. J. Biochem. Cell Biol. 2008, 40, 815–820. [Google Scholar] [CrossRef]
- da Silva Meirelles, L.; Fontes, A.M.; Covas, D.T.; Caplan, A.I. Mechanisms Involved in the Therapeutic Properties of Mesenchymal Stem Cells. Cytokine Growth Factor Rev. 2009, 20, 419–427. [Google Scholar] [CrossRef]
- Linero, I.; Chaparro, O. Paracrine Effect of Mesenchymal Stem Cells Derived from Human Adipose Tissue in Bone Regeneration. PLoS ONE 2014, 9, e107001. [Google Scholar] [CrossRef]
- Hocking, A.M.; Gibran, N.S. Mesenchymal Stem Cells: Paracrine Signaling and Differentiation during Cutaneous Wound Repair. Exp. Cell Res. 2010, 316, 2213–2219. [Google Scholar] [CrossRef]
- Ankrum, J.; Karp, J.M. Mesenchymal Stem Cell Therapy: Two Steps Forward, One Step Back. Trends Mol. Med. 2010, 16, 203–209. [Google Scholar] [CrossRef]
- Horie, M.; Choi, H.; Lee, R.H.; Reger, R.L.; Ylostalo, J.; Muneta, T.; Sekiya, I.; Prockop, D.J. Intra-Articular Injection of Human Mesenchymal Stem Cells (MSCs) Promote Rat Meniscal Regeneration by Being Activated to Express Indian Hedgehog That Enhances Expression of Type II Collagen. Osteoarthr. Cartil. 2012, 20, 1197–1207. [Google Scholar] [CrossRef] [PubMed]
- Moh, M.C.; Shen, S. The Roles of Cell Adhesion Molecules in Tumor Suppression and Cell Migration: A New Paradox. Cell Adh. Migr. 2009, 3, 334–336. [Google Scholar] [CrossRef] [PubMed]
- Gumbiner, B.M. Cell Adhesion: The Molecular Basis of Tissue Architecture and Morphogenesis. Cell 1996, 84, 345–357. [Google Scholar] [CrossRef]
- Yang, C.; Lei, D.; Ouyang, W.; Ren, J.; Li, H.; Hu, J.; Huang, S. Conditioned Media from Human Adipose Tissue-Derived Mesenchymal Stem Cells and Umbilical Cord-Derived Mesenchymal Stem Cells Efficiently Induced the Apoptosis and Differentiation in Human Glioma Cell Lines in Vitro. Biomed Res. Int. 2014, 2014, 109389. [Google Scholar] [CrossRef] [PubMed]
- Velpula, K.K.; Dasari, V.R.; Tsung, A.J.; Gondi, C.S.; Klopfenstein, J.D.; Mohanam, S.; Rao, J.S. Regulation of Glioblastoma Progression by Cord Blood Stem Cells Is Mediated by Downregulation of Cyclin D1. PLoS ONE 2011, 6, e18017. [Google Scholar] [CrossRef]
- Kološa, K.; Motaln, H.; Herold-Mende, C.; Koršič, M.; Lah, T.T. Paracrine Effects of Mesenchymal Stem Cells Induce Senescence and Differentiation of Glioblastoma Stem-like Cells. Cell Transplant. 2015, 24, 631–644. [Google Scholar] [CrossRef]
- Jiao, H.; Guan, F.; Yang, B.; Li, J.; Shan, H.; Song, L.; Hu, X.; Du, Y. Human Umbilical Cord Blood-Derived Mesenchymal Stem Cells Inhibit C6 Glioma via Downregulation of Cyclin D1. Neurol. India 2011, 59, 241–247. [Google Scholar] [CrossRef]
- Ganta, C.; Chiyo, D.; Ayuzawa, R.; Rachakatla, R.; Pyle, M.; Andrews, G.; Weiss, M.; Tamura, M.; Troyer, D. Rat Umbilical Cord Stern Cells Completely Abolish Rat Mammary Carcinomas with No Evidence of Metastasis or Recurrence 100 Days Post-Tumor Cell Inoculation. Cancer Res. 2009, 69, 1815–1820. [Google Scholar] [CrossRef] [PubMed]
- Gauthaman, K.; Yee, F.C.; Cheyyatraivendran, S.; Biswas, A.; Choolani, M.; Bongso, A. Human Umbilical Cord Wharton’s Jelly Stem Cell (HWJSC) Extracts Inhibit Cancer Cell Growth in Vitro. J. Cell. Biochem. 2012, 113, 2027–2039. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhou, C.; Chen, X.; Tao, C.; Cheng, H.; Lu, X. Suppression of Tumor Cell Proliferation and Migration by Human Umbilical Cord Mesenchymal Stem Cells: A Possible Role for Apoptosis and Wnt Signaling. Oncol. Lett. 2018, 15, 8536–8544. [Google Scholar] [CrossRef] [PubMed]
- Khalil, C.; Moussa, M.; Azar, A.; Tawk, J.; Habbouche, J.; Salameh, R.; Ibrahim, A.; Alaaeddine, N. Anti-Proliferative Effects of Mesenchymal Stem Cells (MSCs) Derived from Multiple Sources on Ovarian Cancer Cell Lines: An in-Vitro Experimental Study. J. Ovarian Res. 2019, 12, 70. [Google Scholar] [CrossRef] [PubMed]
- Dzobo, K.; Vogelsang, M.; Thomford, N.E.; Dandara, C.; Kallmeyer, K.; Pepper, M.S.; Parker, M.I. Wharton’s Jelly-Derived Mesenchymal Stromal Cells and Fibroblast-Derived Extracellular Matrix Synergistically Activate Apoptosis in a P21-Dependent Mechanism in WHCO1 and MDA MB 231 Cancer Cells In Vitro. Stem Cells Int. 2016, 2016, 4842134. [Google Scholar] [CrossRef] [PubMed]
- Koh, D.-I.; Han, D.; Ryu, H.; Choi, W.-I.; Jeon, B.-N.; Kim, M.-K.; Kim, Y.; Kim, J.Y.; Parry, L.; Clarke, A.R.; et al. KAISO, a Critical Regulator of P53-Mediated Transcription of CDKN1A and Apoptotic Genes. Proc. Natl. Acad. Sci. USA 2014, 111, 15078–15083. [Google Scholar] [CrossRef]
- Larson, P.S.; Schlechter, B.L.; King, C.-L.; Yang, Q.; Glass, C.N.; Mack, C.; Pistey, R.; de las Morenas, A.; Rosenberg, C.L. CDKN1C/P57kip2is a Candidate Tumor Suppressor Gene in Human Breast Cancer. BMC Cancer 2008, 8, 68. [Google Scholar] [CrossRef]
- Jorgovanovic, D.; Song, M.; Wang, L.; Zhang, Y. Roles of IFN-γ in Tumor Progression and Regression: A Review. Biomark. Res. 2020, 8, 49. [Google Scholar] [CrossRef]
- Corso, G.; Veronesi, P.; Sacchini, V.; Galimberti, V. Prognosis and Outcome in CDH1-Mutant Lobular Breast Cancer. Eur. J. cancer Prev. 2018, 27, 237–238. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.A.; Robinson, T.J.; Liu, J.C.; Shrestha, M.; Voisin, V.; Ju, Y.; Chung, P.E.D.; Pellecchia, G.; Fell, V.L.; Bae, S.; et al. RB1 Deficiency in Triple-Negative Breast Cancer Induces Mitochondrial Protein Translation. J. Clin. Investig. 2016, 126, 3739–3757. [Google Scholar] [CrossRef]
- Paplomata, E.; O’Regan, R. The PI3K/AKT/MTOR Pathway in Breast Cancer: Targets, Trials and Biomarkers. Ther. Adv. Med. Oncol. 2014, 6, 154–166. [Google Scholar] [CrossRef]
- Barnes, D.M.; Gillett, C.E. Cyclin D1 in Breast Cancer. Breast Cancer Res. Treat. 1998, 52, 1–15. [Google Scholar] [CrossRef]
- Ding, J.; Guo, C.; Hu, P.; Chen, J.; Liu, Q.; Wu, X.; Cao, Y.; Wu, J. CSF1 Is Involved in Breast Cancer Progression through Inducing Monocyte Differentiation and Homing. Int. J. Oncol. 2016, 49, 2064–2074. [Google Scholar] [CrossRef]
- Masuda, H.; Zhang, D.; Bartholomeusz, C.; Doihara, H.; Hortobagyi, G.N.; Ueno, N.T. Role of Epidermal Growth Factor Receptor in Breast Cancer. Breast Cancer Res. Treat. 2012, 136, 331–345. [Google Scholar] [CrossRef]
- Sullivan, N.J.; Sasser, A.K.; Axel, A.E.; Vesuna, F.; Raman, V.; Ramirez, N.; Oberyszyn, T.M.; Hall, B.M. Interleukin-6 Induces an Epithelial–Mesenchymal Transition Phenotype in Human Breast Cancer Cells. Oncogene 2009, 28, 2940–2947. [Google Scholar] [CrossRef] [PubMed]
- Vleugel, M.M.; Greijer, A.E.; Bos, R.; van der Wall, E.; van Diest, P.J. C-Jun Activation Is Associated with Proliferation and Angiogenesis in Invasive Breast Cancer. Hum. Pathol. 2006, 37, 668–674. [Google Scholar] [CrossRef] [PubMed]
- Imamura, T.; Hikita, A.; Inoue, Y. The Roles of TGF-β Signaling in Carcinogenesis and Breast Cancer Metastasis. Breast Cancer 2012, 19, 118–124. [Google Scholar] [CrossRef]
- Chambers, A.F.; Groom, A.C.; MacDonald, I.C. Dissemination and Growth of Cancer Cells in Metastatic Sites. Nat. Rev. Cancer 2002, 2, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, H.; Wyckoff, J.; Condeelis, J. Cell Migration in Tumors. Curr. Opin. Cell Biol. 2005, 17, 559–564. [Google Scholar] [CrossRef]
- Friedl, P.; Wolf, K. Tumour-Cell Invasion and Migration: Diversity and Escape Mechanisms. Nat. Rev. Cancer 2003, 3, 362–374. [Google Scholar] [CrossRef]
- Mittal, V. Epithelial Mesenchymal Transition in Tumor Metastasis. Annu. Rev. Pathol. 2018, 13, 395–412. [Google Scholar] [CrossRef]
- Roche, J. The Epithelial-to-Mesenchymal Transition in Cancer. Cancers 2018, 10, 52. [Google Scholar] [CrossRef]
- Scimeca, M.; Antonacci, C.; Colombo, D.; Bonfiglio, R.; Buonomo, O.C.; Bonanno, E. Emerging Prognostic Markers Related to Mesenchymal Characteristics of Poorly Differentiated Breast Cancers. Tumor Biol. 2016, 37, 5427–5435. [Google Scholar] [CrossRef]
- Micalizzi, D.S.; Ford, H.L. Epithelial-Mesenchymal Transition in Development and Cancer. Future Oncol. 2009, 5, 1129–1143. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhou, B.P. Epithelial-Mesenchymal Transition---A Hallmark of Breast Cancer Metastasis. Cancer Hallm. 2013, 1, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Brabletz, T.; Kalluri, R.; Nieto, M.A.; Weinberg, R.A. EMT in Cancer. Nat. Rev. Cancer 2018, 18, 128–134. [Google Scholar] [CrossRef]
- Visweswaran, M.; Arfuso, F.; Dilley, R.J.; Newsholme, P.; Dharmarajan, A. The Inhibitory Influence of Adipose Tissue-Derived Mesenchymal Stem Cell Environment and Wnt Antagonism on Breast Tumour Cell Lines. Int. J. Biochem. Cell Biol. 2018, 95, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Han, G.; Liu, H.; Qin, C. Suppression of Cholangiocarcinoma Cell Growth by Human Umbilical Cord Mesenchymal Stem Cells: A Possible Role of Wnt and Akt Signaling. PLoS ONE 2013, 8, e62844. [Google Scholar] [CrossRef]
- Grunwald, G.B. The Structural and Functional Analysis of Cadherin Calcium-Dependent Cell Adhesion Molecules. Curr. Opin. Cell Biol. 1993, 5, 797–805. [Google Scholar] [CrossRef]
- Lei, H.; Sjöberg-Margolin, S.; Salahshor, S.; Werelius, B.; Jandáková, E.; Hemminki, K.; Lindblom, A.; Vorechovský, I. CDH1 Mutations Are Present in Both Ductal and Lobular Breast Cancer, but Promoter Allelic Variants Show No Detectable Breast Cancer Risk. Int. J. Cancer 2002, 98, 199–204. [Google Scholar] [CrossRef]
- Berx, G.; Cleton-Jansen, A.M.; Nollet, F.; de Leeuw, W.J.; van de Vijver, M.; Cornelisse, C.; van Roy, F. E-Cadherin Is a Tumour/Invasion Suppressor Gene Mutated in Human Lobular Breast Cancers. EMBO J. 1995, 14, 6107–6115. [Google Scholar] [CrossRef]
- Tau, G.Z.; Cowan, S.N.; Weisburg, J.; Braunstein, N.S.; Rothman, P.B. Regulation of IFN-Gamma Signaling Is Essential for the Cytotoxic Activity of CD8(+) T Cells. J. Immunol. 2001, 167, 5574–5582. [Google Scholar] [CrossRef]
- Liu, H.; Shen, Z.; Wang, Z.; Wang, X.; Zhang, H.; Qin, J.; Qin, X.; Xu, J.; Sun, Y. Increased Expression of IDO Associates with Poor Postoperative Clinical Outcome of Patients with Gastric Adenocarcinoma. Sci. Rep. 2016, 6, 21319. [Google Scholar] [CrossRef] [PubMed]
- Manore, S.G.; Doheny, D.L.; Wong, G.L.; Lo, H.-W. IL-6/JAK/STAT3 Signaling in Breast Cancer Metastasis: Biology and Treatment. Front. Oncol. 2022, 12, 866014. [Google Scholar] [CrossRef] [PubMed]
- Sizemore, S.T.; Sizemore, G.M.; Booth, C.N.; Thompson, C.L.; Silverman, P.; Bebek, G.; Abdul-Karim, F.W.; Avril, S.; Keri, R.A. Hypomethylation of the MMP7 Promoter and Increased Expression of MMP7 Distinguishes the Basal-like Breast Cancer Subtype from Other Triple-Negative Tumors. Breast Cancer Res. Treat. 2014, 146, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Al Subayyil, A.; Basmaeil, Y.S.; Alenzi, R.; Khatlani, T. Human Placental Mesenchymal Stem/Stromal Cells (PMSCs) Inhibit Agonist-Induced Platelet Functions Reducing Atherosclerosis and Thrombosis Phenotypes. J. Cell. Mol. Med. 2021, 25, 9268–9280. [Google Scholar] [CrossRef] [PubMed]
- Basmaeil, Y.; Bahattab, E.; Al Subayyil, A.; Kulayb, H.B.; Alrodayyan, M.; Abumaree, M.; Khatlani, T. Decidua Parietalis Mesenchymal Stem/Stromal Cells and Their Secretome Diminish the Oncogenic Properties of MDA231 Cells In Vitro. Cells 2021, 10, 3493. [Google Scholar] [CrossRef] [PubMed]
- Vistejnova, L.; Dvorakova, J.; Hasova, M.; Muthny, T.; Velebny, V.; Soucek, K.; Kubala, L. The Comparison of Impedance-Based Method of Cell Proliferation Monitoring with Commonly Used Metabolic-Based Techniques. Neuro Endocrinol. Lett. 2009, 30 (Suppl. 1), 121–127. [Google Scholar]
- Stefanowicz-Hajduk, J.; Ochocka, J.R. Real-Time Cell Analysis System in Cytotoxicity Applications: Usefulness and Comparison with Tetrazolium Salt Assays. Toxicol. Rep. 2020, 7, 335–344. [Google Scholar] [CrossRef]
- Dowling, C.M.; Herranz Ors, C.; Kiely, P.A. Using Real-Time Impedance-Based Assays to Monitor the Effects of Fibroblast-Derived Media on the Adhesion, Proliferation, Migration and Invasion of Colon Cancer Cells. Biosci. Rep. 2014, 34, 415–427. [Google Scholar] [CrossRef]
- Bahattab, E.; Khatlani, T.; Abomaray, F.M.; Messaoudi, S.A.; Abumaree, M.H. Cancer Conditioned Medium Modulates Functional and Phenotypic Properties of Human Decidua Parietalis Mesenchymal Stem/Stromal Cells. Tissue Eng. Regen. Med. 2019, 16, 615–630. [Google Scholar] [CrossRef]
- Basmaeil, Y.; Rashid, M.A.; Khatlani, T.; AlShabibi, M.; Bahattab, E.; Abdullah, M.L.; Abomaray, F.; Kalionis, B.; Massoudi, S.; Abumaree, M. Preconditioning of Human Decidua Basalis Mesenchymal Stem/Stromal Cells with Glucose Increased Their Engraftment and Anti-Diabetic Properties. Tissue Eng. Regen. Med. 2020, 17, 209–222. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oncogenes | ||||
|---|---|---|---|---|
| Fold Change Expression as Compared to Control | ||||
| Gene Symbol | Gene Name | CVMSCs CM 25% Treatment | MDA231:CVMSCs (1:5) Treatment | |
| 1 | AKT1 | AKT Serine/Threonine Kinase 1 | 0.618622154 | 0.150137447 |
| 2 | ATM | Ataxia Telangiectasia Mutated | 1.778029701 | 0.000298889 |
| 3 | BIRC5 | Baculoviral IAP Repeat Containing 5 | 1.233057039 | 0.619744644 |
| 4 | CCND1 | Cyclin D1 | 0.290049141 | 0.231581661 |
| 5 | CCND2 | Cyclin D2 | 1.287981276 | 0.040808033 |
| 6 | CCNE1 | Cyclin E1 | 1.864428555 | 0.004532964 |
| 7 | CSF1 | Colony Stimulating Factor 1 | 0.348220492 | 0.086619766 |
| 8 | EGFR | Epidermal Growth Factor Receptor | 0.569847063 | 0.001123859 |
| 9 | ERBB2 | erb-b2 Receptor Tyrosine Kinase 2 | 0.598625231 | 0.00608788 |
| 10 | ESR1 | Estrogen Receptor Alpha | 1.379391323 | 0.070357461 |
| 11 | GRB7 | Growth Factor Receptor-Bound Protein-7 | 1.307511163 | 0.737863692 |
| 12 | GSTP1 | Glutathione-S-transferase Pi 1 | 0.187847314 | 0.41971974 |
| 13 | IDO | Indoleamine-2,3-Dioxygenase Enzyme | 0.285609401 | 0.269798431 |
| 14 | IGF1R | Insulin like Growth Factor 1 Receptor | 0.588756921 | 0.174552115 |
| 15 | IGFBP3 | Insulin-like Growth Factor Binding Protein 3 | 1.26035563 | 0.80957975 |
| 16 | IL6 | Interleukin-6 | 0.759431623 | 0.001836595 |
| 17 | JUN | Jun Proto-oncogene. | 0.479301652 | 0.003486001 |
| 18 | KRT18 | Keratin 18 | 0.578317336 | 0.535014722 |
| 19 | KRT8 | Keratin8 | 1.239435925 | 0.279858319 |
| 20 | MKI67 | Marker of Proliferation Ki-67 | 0.538599614 | 0.000380797 |
| 21 | MMP2 | Matrix Metalloproteinase 2 | 1.801857196 | 0.007483997 |
| 22 | MMP7 | Matrix Metalloproteinase 7 | 1.562817349 | 0.870375475 |
| 23 | MUC1 | Mucin 1 | 0.659100118 | 0.431684903 |
| 24 | NOTCH1 | Neurogenic Locus Notch Homolog Protein 1 | 1.081148284 | 0.350957369 |
| 25 | PLAU | Urokinase-Plasminogen Activator | 0.428671888 | 0.69880195 |
| 26 | PTGS2 | Cyclooxygenase 2 | 1.156198717 | 0.003074883 |
| 27 | SLC39A6 | Solute Carrier Family 39 Member 6 | 1.348045319 | 0.819709278 |
| 28 | TFF3 | Trefoil Factor 3 | 1.163580106 | 0.003049453 |
| 29 | TGFB1 | Transforming Growth Factor | 0.364712528 | 0.000123789 |
| Tumor Suppressor Genes | ||||
| 1 | CDKN1A | Cyclin-dependent Kinase Inhibitor A | 1.467611692 | 2.072603835 |
| 2 | CDKN1C | Cyclin Dependent Kinase Inhibitor 1C | 2.097303084 | 3.08391553 |
| 3 | IFN-γ | Interferon Gamma | 3.946022877 | 6.013921647 |
| 4 | CDH1 | E-Cadherin | 1.80114776 | 2.529275488 |
| 5 | RB1 | Retinoblastoma Protein | 1.210082072 | 2.477135473 |
| 6 | SFRP1 | Secreted Frizzled Related Protein 1 | 29.89537164 | 87.56450986 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Subayyil, A.A.; Basmaeil, Y.S.; Kulayb, H.B.; Alrodayyan, M.; Alhaber, L.A.A.; Almanaa, T.N.; Khatlani, T. Preconditioned Chorionic Villus Mesenchymal Stem/Stromal Cells (CVMSCs) Minimize the Invasive Phenotypes of Breast Cancer Cell Line MDA231 In Vitro. Int. J. Mol. Sci. 2023, 24, 9569. https://doi.org/10.3390/ijms24119569
Subayyil AA, Basmaeil YS, Kulayb HB, Alrodayyan M, Alhaber LAA, Almanaa TN, Khatlani T. Preconditioned Chorionic Villus Mesenchymal Stem/Stromal Cells (CVMSCs) Minimize the Invasive Phenotypes of Breast Cancer Cell Line MDA231 In Vitro. International Journal of Molecular Sciences. 2023; 24(11):9569. https://doi.org/10.3390/ijms24119569
Chicago/Turabian StyleSubayyil, Abdullah Al, Yasser S. Basmaeil, Hayaa Bin Kulayb, Maha Alrodayyan, Lama Abdulaziz A. Alhaber, Taghreed N. Almanaa, and Tanvir Khatlani. 2023. "Preconditioned Chorionic Villus Mesenchymal Stem/Stromal Cells (CVMSCs) Minimize the Invasive Phenotypes of Breast Cancer Cell Line MDA231 In Vitro" International Journal of Molecular Sciences 24, no. 11: 9569. https://doi.org/10.3390/ijms24119569
APA StyleSubayyil, A. A., Basmaeil, Y. S., Kulayb, H. B., Alrodayyan, M., Alhaber, L. A. A., Almanaa, T. N., & Khatlani, T. (2023). Preconditioned Chorionic Villus Mesenchymal Stem/Stromal Cells (CVMSCs) Minimize the Invasive Phenotypes of Breast Cancer Cell Line MDA231 In Vitro. International Journal of Molecular Sciences, 24(11), 9569. https://doi.org/10.3390/ijms24119569