1. Introduction
Regulated gene switches were designed and constructed to turn “on” and “off” the expression of a transgene using an inducer [
1,
2]. The development of these switches seeks to properly dose the delivery of a therapeutic gene product (RNA or protein), avoiding the toxic effects of overexpression when constitutive promoters are used. This feature makes the regulatable gene switches useful in basic research for studies requiring especially precise and acute control of the gene delivery to unveil its function. Furthermore, gene switches have promising uses in biomedicine to provide regulated dosed delivery of biopharmaceuticals or for use in gene replacement therapy, as reviewed in [
3].
A critical issue for gene switches is the choice of the appropriate inducer to provide high spatial and temporal resolution of the expression. Most of the regulatable gene switches initially used chemical inducers [
4], but they can be toxic in high doses and may freely diffuse in the system, being difficult to control its location or elimination and preventing the fine-tuning required for therapeutic gene expression. These features limit the temporal and spatial resolution of chemical-induced gene expression [
5]. Light-activated gene switches have been proposed as an alternative to chemical gene switches [
6,
7,
8,
9,
10]. Light has proven to be an ideal inducer to overcome chemical inducers, as it is cheap, easy to obtain, highly adjustable, and renders higher spatial and temporal control of expression [
11,
12,
13].
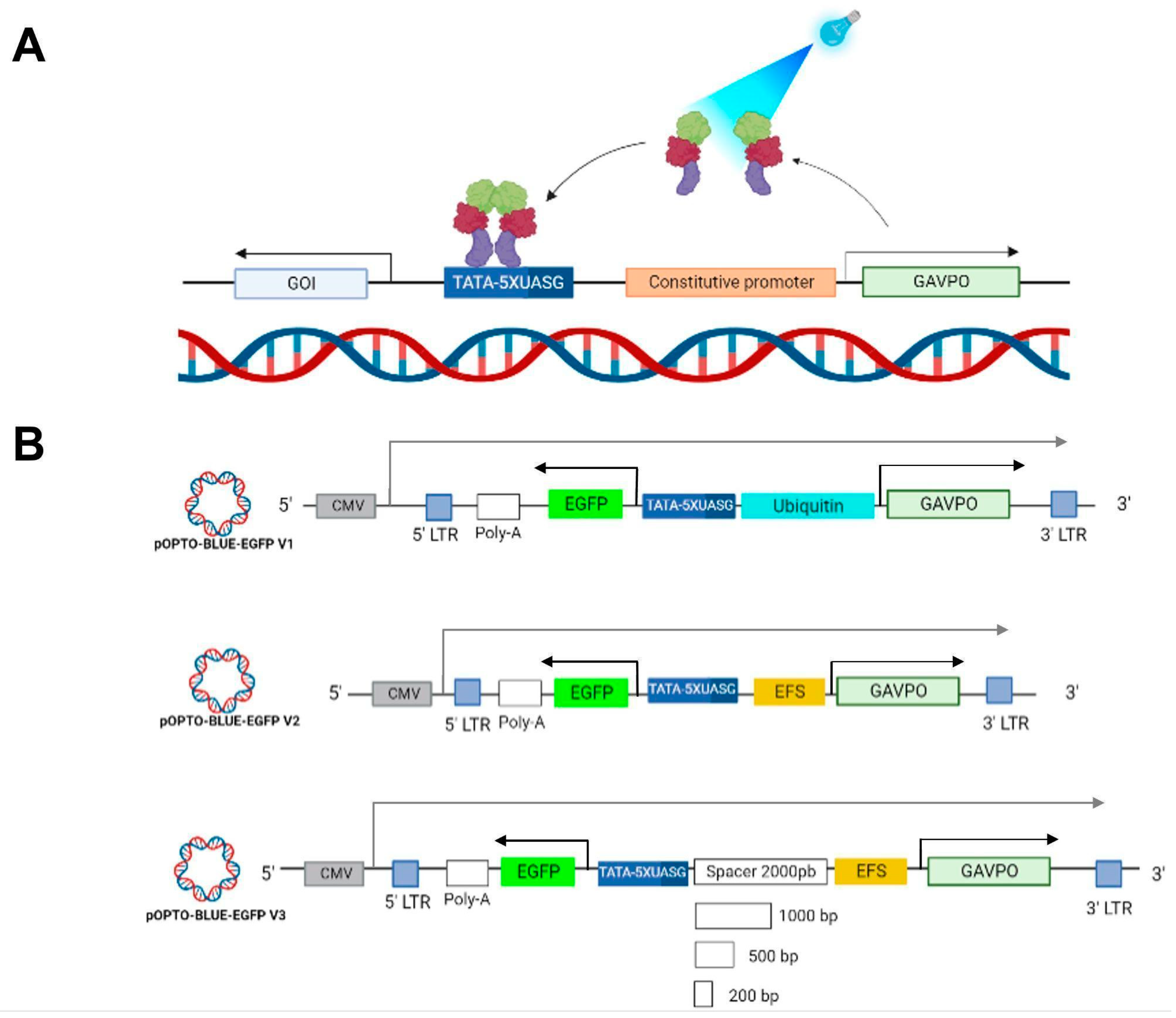
Distinct optogenetic gene switches have been developed to regulate transgene expression using different wavelengths and light-responsive proteins [
14,
15]. The LightOn system is an optogenetic gene switch developed by Wang. et al. [
16], composed of two components: (i) a blue light switchable chimeric transcription factor GAVPO containing a photosensitive LOV domain, which homodimerizes upon exposure to blue light, and (ii) a GAVPO responsive promoter composed of five repetitions of the upstream activation sequence of Gal4 (UAS), fused to a minimal adenovirus E1b promoter (UAS inducible promoter). The LightOn system has been shown to be able to control transgene expression in HEK293 cells and mice [
14]. In addition, it has been used to control the cell fate of neural progenitor cells and chick-limb mesenchyme cells [
17,
18], study cell ablation in zebrafish [
19], regulate blood-glucose homeostasis in type 1 diabetic mice [
20] and target a potential breast cancer tumor by a light-switchable gene expression system encapsulated in a nanoparticle delivery system [
20].
We previously optimized and adapted the LightOn system to a lentiviral platform [
21]. Lentiviruses allow the integration of genetic material into the host-cell genome so that genetic incorporation remains stable over time [
22]. The lentiviral plasmids are also an advantageous tool for manipulating the genomes of post-mitotic cells, such as neurons. In our previous work, we constructed two lentiviral plasmids bearing the two components of the LightOn system: (i) one that expresses GAVPO under the Ubiquitin constitutive promoter; and (ii) a second plasmid that expresses a gene of interest (GOI) driven by the UAS inducible promoter. We demonstrated the functionality of this system through the blue light-induced expression of the cerebral dopamine neurotrophic factor and mCherry by transfection and transduction of mammalian cell lines and neurons [
21]. As the LightOn lentiviral system is based on two lentiviruses, its efficient operation requires the co-transduction of the target cells in the tissue where the regulation of gene expression is desired. This co-transduction requirement decreases the efficient and equitable delivery of all LightOn system components due to possible interference between the lentiviruses to achieve the dual transduction of the target cells. To overcome this problem, we designed and constructed the OPTO-BLUE system, an integrated bidirectional lentiviral system containing all the components of the LightOn system in a single lentivirus, thus improving the mentioned disadvantages and favoring the equitable incorporation of all the components through a single transduction event. We constructed the pOPTO-BLUE-EGFP bidirectional lentiviral plasmid and optimized its design to minimize background activity in darkness. We transfected pOPTO-BLUE-EGFP in HEK293-T mammalian cells to evaluate the functionality of the OPTO-BLUE system and the optimal lighting parameters to achieve an efficient expression. The results showed that blue light-induced enhanced green fluorescent protein (EGFP) level expression could be regulated through intensity and lighting time exposure, reaching a two-fold induction after 16 h of illumination. We further demonstrated blue light-inducible EGFP expression by transducing HEK293-T cells. These findings prove that the OPTO-BLUE system allows the blue light control of a GOI in mammalian cells, suggesting its potential use in biomedical applications requiring dosed therapeutic gene expression.
3. Discussion
The Light-On inducible expression system is a highly regulated, light-inducible expression system with multiple applications in biology. Accordingly, its therapeutic approach could be of ample use in gene therapy [
11,
14]. This system allows spatiotemporal control of gene expression, where light could tightly control the dosed expression of therapeutic proteins or biologicals. For the effective use of this system in gene therapy, especially in the central nervous system, it is required to adapt the molecular components into the proper vector that efficiently could incorporate the Light-On system into the target cells or tissues. For that purpose, we have adapted the Light-On system to a single lentiviral platform that has shown to be very reliable and efficient in transferring genetic information in HEK293-T cells. We previously developed a lentiviral Light-On platform that relies on the co-transduction of two viral particles to regulate gene expression effectively [
21]. Although this system is appropriate in an experimental setting in cell biology research, it is not practical for applications in a therapeutic situation, due to the difficulty of transducing two viral particles to the same target cell, where the insertion of the expression cassettes of the GAVPO transactivator and the regulated light-induced UAS expression cassette with the GOI is required. This is much easier achieved with a two-plasmid system carrying each expression cassette, as recently demonstrated by the inclusion of both plasmids in a nanoparticle delivery system for the expression of diphtheria toxin for cancer treatment, where a spatiotemporal control is achieved [
20]. Here, we showed a functional and inducible Light-On system in a single lentiviral vector, coined OPTO-BLUE, where the cassette for the constitutive expression of the transactivator protein GAVPO and the GAVPO-regulated UAS minimal-promoter driving the expression of the GOI is arranged in a head-to-head configuration. This arrangement allows the generation of lentiviral particles fully capable of transducing mammalian cells carrying the LightOn system. Furthermore, we characterized the optimal separation between the promoters to decrease background expression in the absence of light, as well as the light-intensity and temporal induction of a reporter protein EGFP on this system.
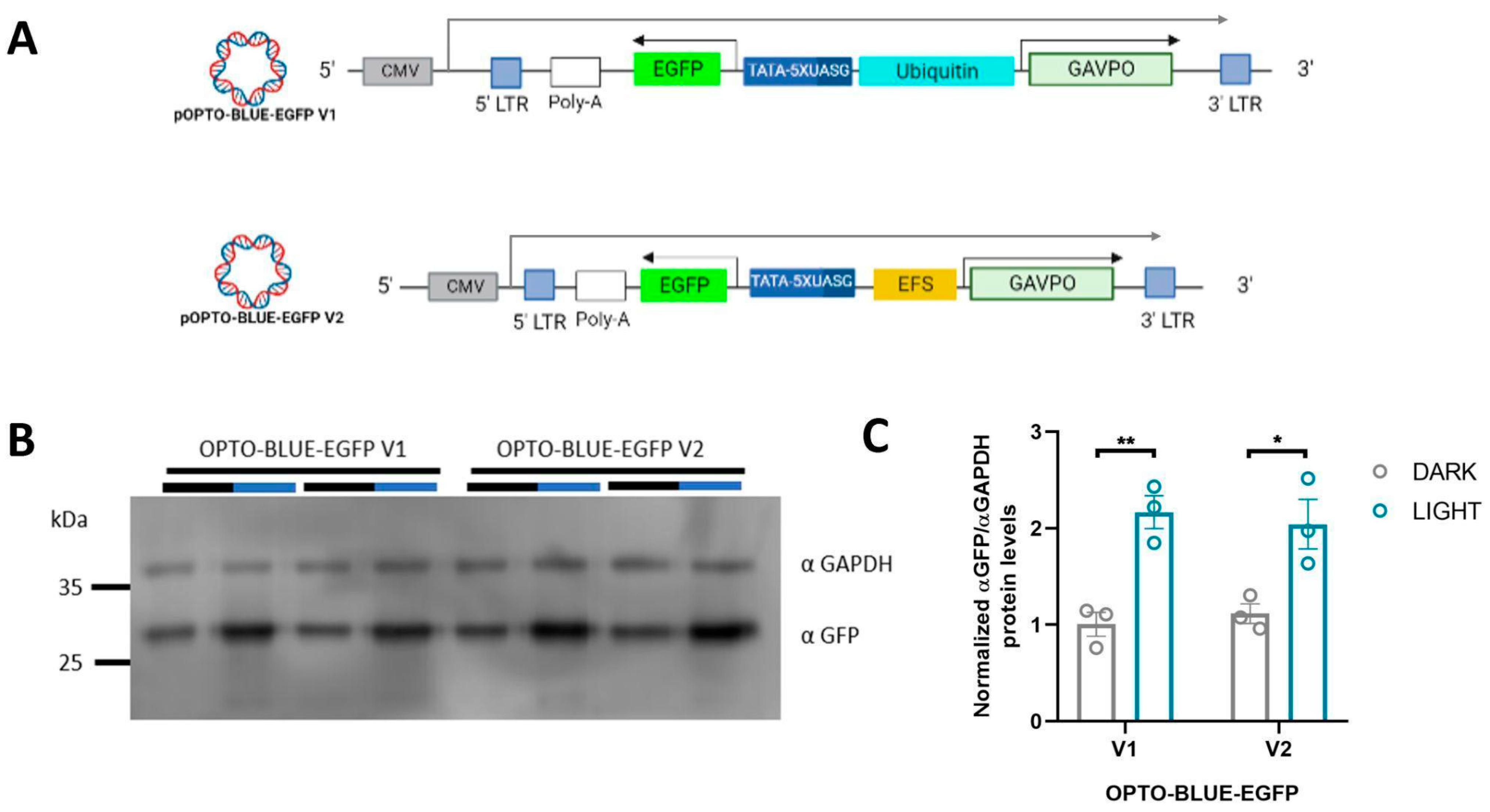
The OPTO-BLUE lentiviral system is a single integrated Light-On inducible expression system that combines the cassette for the expression of the GAVPO transactivator and the UAS cassette containing the GOI in a single viral transcript. Different hurdles must be overcome and tested to demonstrate that a functional and inducible lentiviral vector was obtained. We first attempted to compact most of the genetic elements required for GAVPO expression, reducing the size of the constitutive Ubiquitin promoter for the EFS promoter. The EFS promoter is robust and resistant to the silencing of the promoter in a lentiviral setting [
23], allowing a reduction of approximately 1 kb of the packaged viral transcript. Although the size of the maximal packaged lentiviral genome has not been defined, it is known that increasing the size of the transcript results in a lower viral titer [
24]. Therefore, the reduction of approximately 1 kb on the packaging transcripts should not only allow the expression of a larger GOI, but also could result in higher titers for those small viral particles that harbor smaller genomes. Furthermore, the decrease in the transfer plasmid size should also lead to better transfection efficiency during viral production, helping to obtain better viral titers [
25].
The mechanism of gene activation in the OPTO-BLUE system involves the dimerization of GAVPO, that is, transduced cells should be acting on a single site or a few sites, if considered a single or a few viral-integration events. Although the dynamics of GAVPO gene activation are not completely resolved, the LOV domain has an equilibrium–dissociation constant (Kd) in a micromolar range [
26], suggesting that a robust promoter must drive the expression of GAVPO. The EFS promoter reached similar levels of GAVPO expression to the Ubiquitin promoter. Therefore, the OPTO-BLUE system with the EFS promoter should achieve comparable functionality as the Ubiquitin promoter.
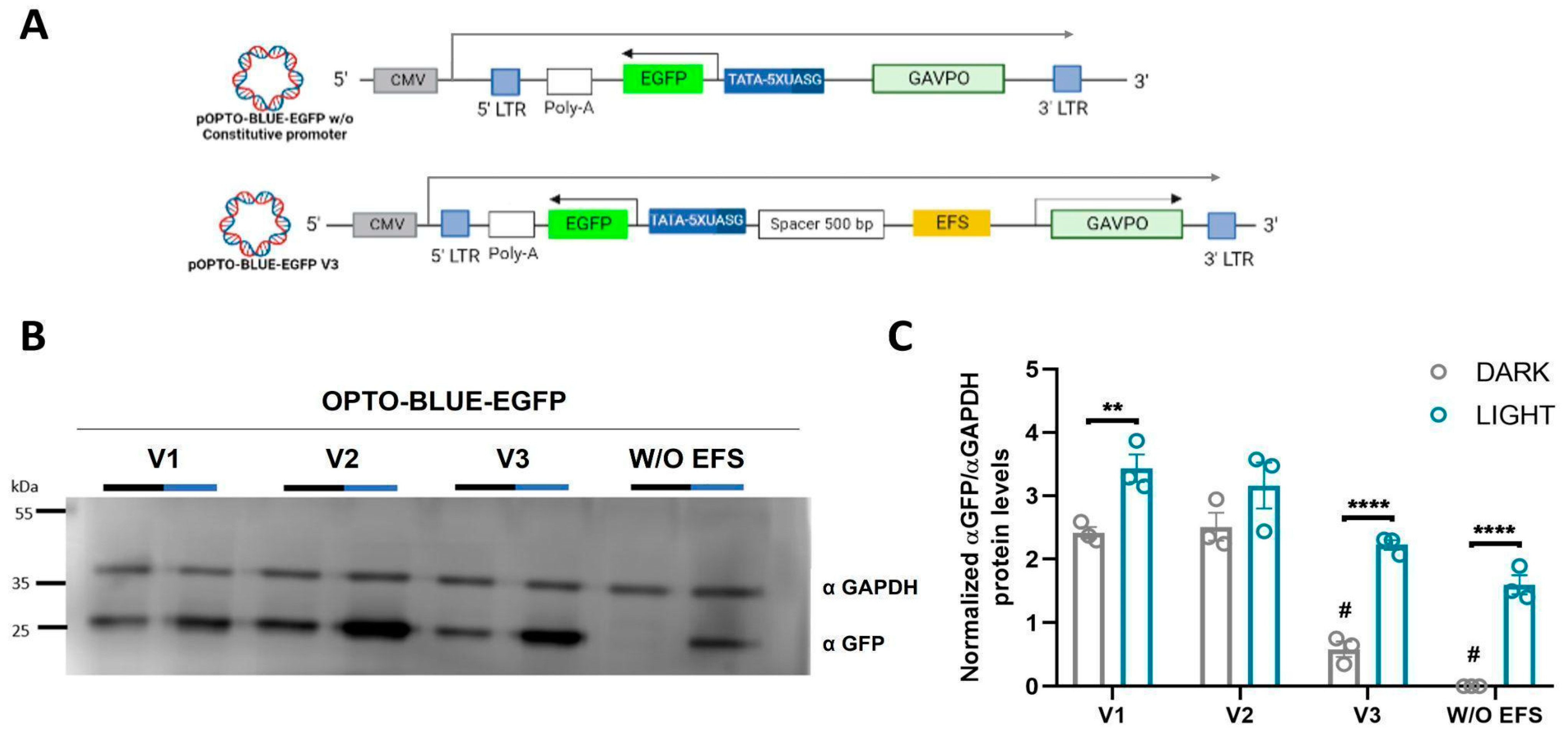
The second improvement to address was to design a transfer vector with both elements of the Light-On system as a pair of cis-acting genetic elements located on different expression cassettes. This design should permit the synthesis of full transfer/packaging transcripts containing both long terminal repeats (LTRs) in a configuration that allows the entire retro transcription process. This allows the insertion and transduction of eukaryotic cells by the lentiviral particles. The generation of the packaging transcript in many lentiviral systems is driven by a CMV promoter located upstream of the 5′LTR sequence. RNA pol II is recruited by this promoter, generating a capped transcript that is polyadenylated at the 3′ ends by the polyadenylation signal on the 3′LTR [
27]. The two components of the Light-On system consist of two expression cassettes, one for the expression of GAVPO and the second for the expression of GOI. Both cassettes are driven by RNA pol II; therefore, both cassettes must contain a polyadenylation signal. We hypothesize that the best arrangement of both expression cassettes, acting in
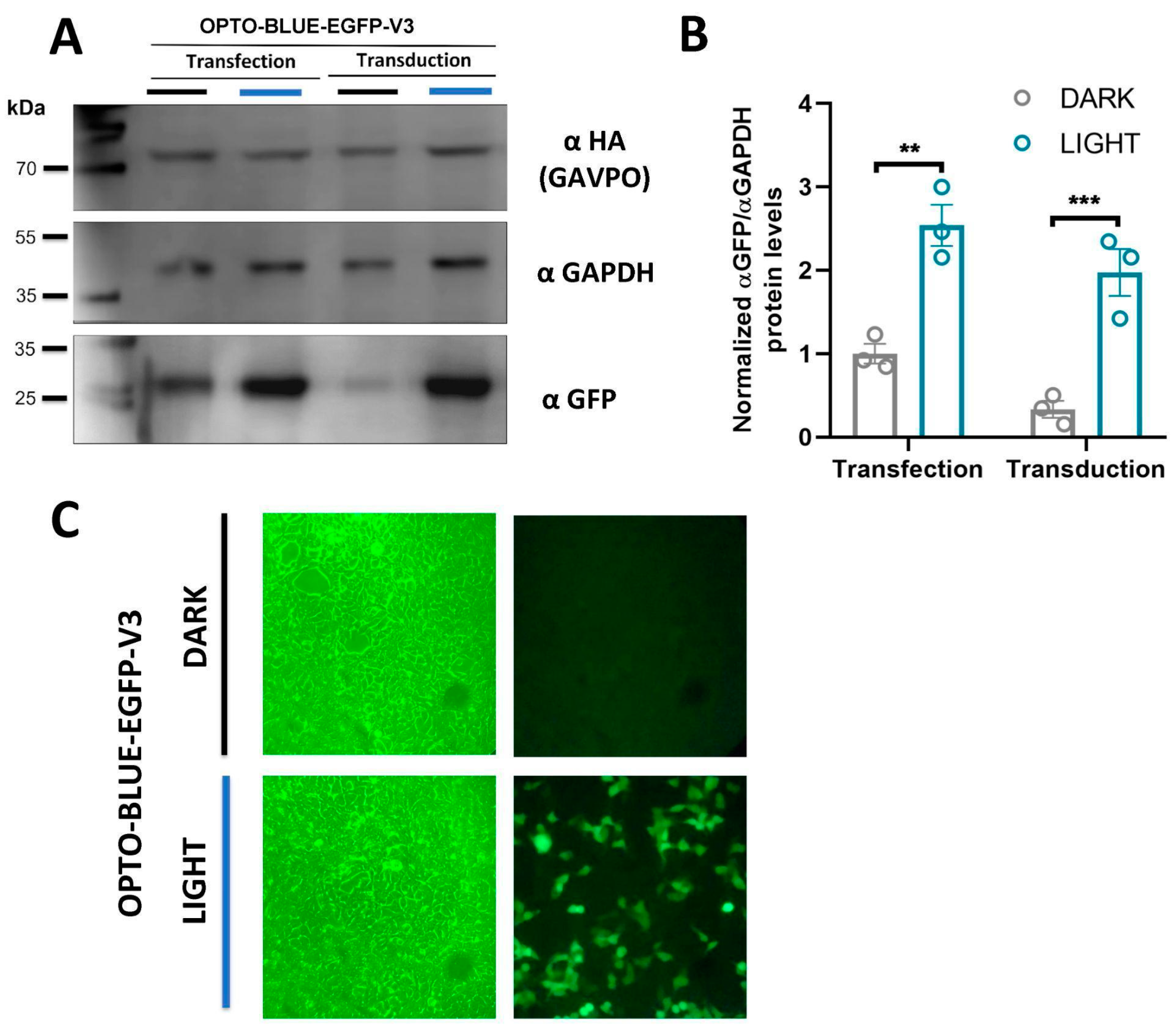
CIS, without interfering with the generation of a full transcript, was by designing the UAS-GOI-polyA signal expression cassette in the opposite direction of the EFS-GAVPO-LTR (polyA signal) cassette. The polyA signal in the UAS-GOI-polyA cassette is in the opposite strand and should not interfere with generating the full package transcript. Transfection experiments indicate that both cassettes are functional, as EGFP expression is activated by light, and the GAVPO protein is detected. Despite this functionality, we observed a high-background expression in the darkness. Considering the minimal UAS promoter is of short size, and the EFS promoter is a robust promoter [
23], the head-to-head configuration of both promoters could result in a high-background expression due to the closeness of both promoters, in a manner similar to the design of bidirectional Light-On vectors [
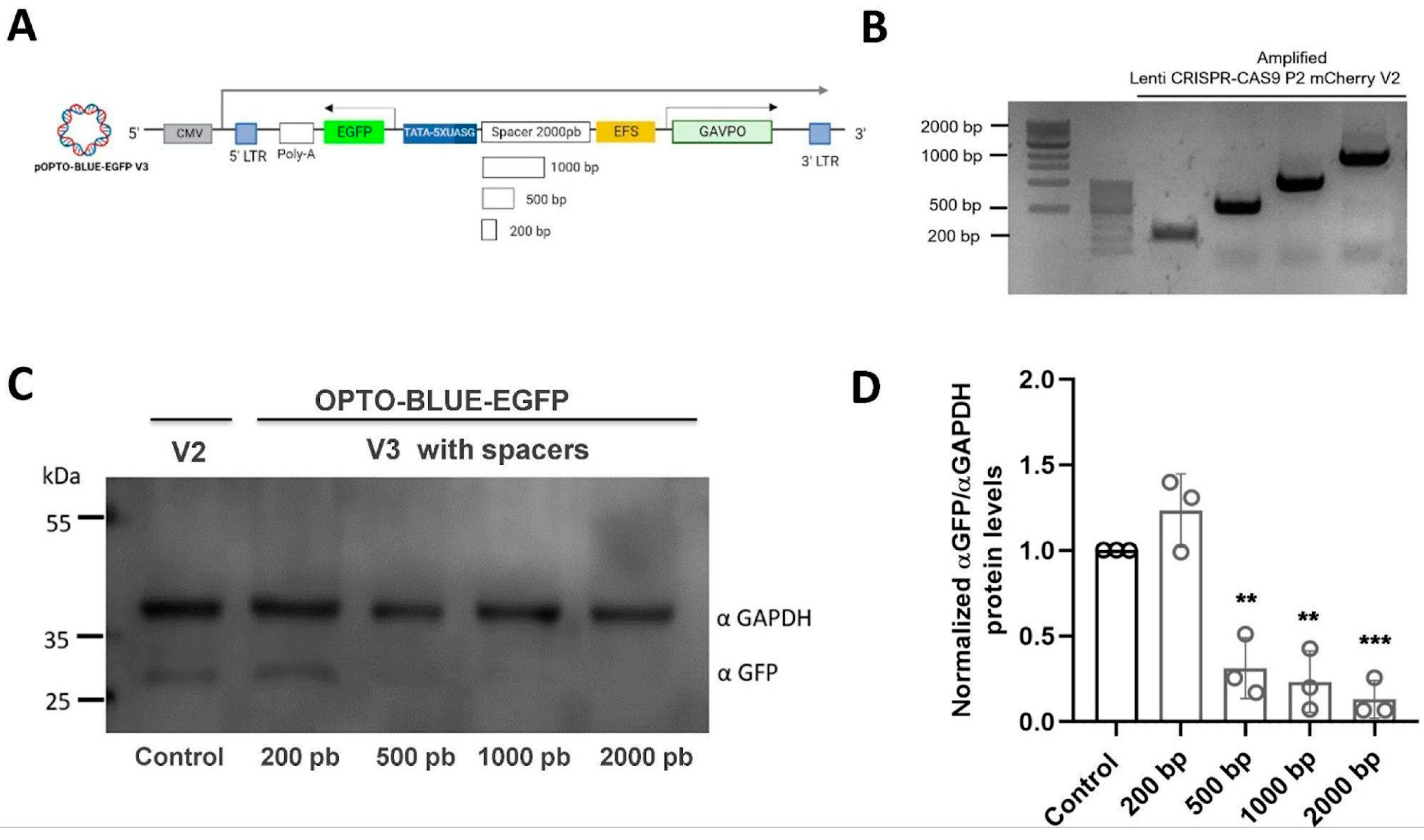
12]. Therefore, we constructed a series of OPTO BLUE vectors with different spacers to assess the background expression of the GOI. The stuffer fragment was amplified from ORF of Cas9 using the pLenti-CRISPR-Cas9-P2-mCherry-V2 vector and comprehensively analyzed to avoid the presence of regulatory gene elements. As expected, the background expression was decreased as a measure of the distance separating the promoters, highly suggesting that the assembly of the transcription initiation complex of the EFS promoter could be inducing the expression of the GOI. We tried stuffer sequences of different sizes, from 0.2–2.0 kb, and found that a separation of 0.5 kb is enough to decrease background expression significantly. The background expression is even lower with larger spacers, obtaining the lowest background with the largest spacer tested at 2.0 kb. However, we suggest that a spacer of 0.5 kb should be enough to be functional, offering a significant signal (light) to noise (background) ratio; but the interplay between the amount of background desired and the size of the GOI wanting to be expressed should be considered when choosing the proper OPTO BLUE plasmid.
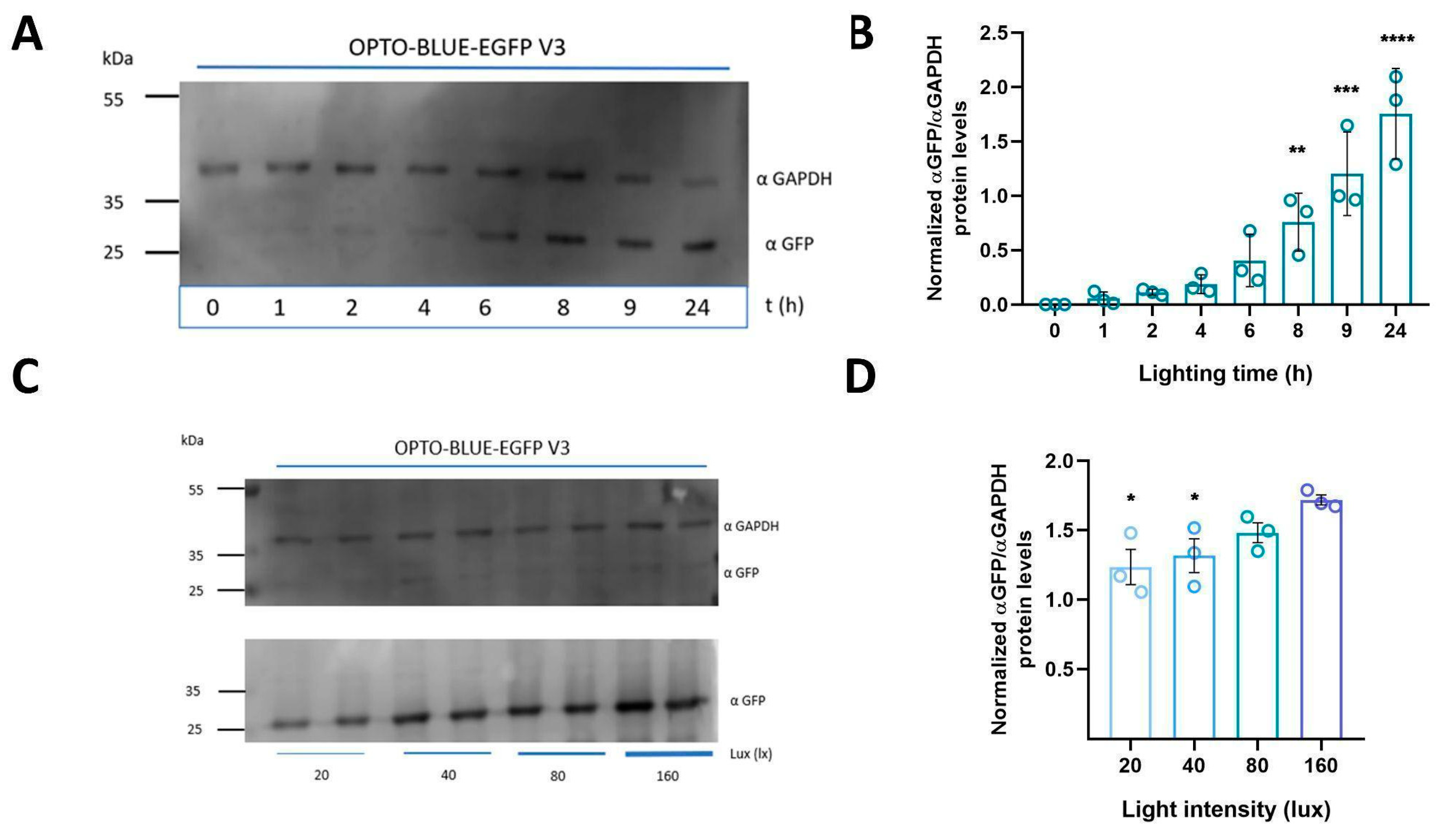
The Light-On system is quite sensible for light induction [
11]. The OPTO BLUE system seems to maintain this dynamic since it can increase the expression of the GOI after 2 h of induction, which becomes significant after 8 h of illumination. This expression keeps increasing as the light induction is maintained, similar to previous reports [
14,
21]. Furthermore, we observed that the expression of EGFP could be regulated by light intensity. Similar to the LuminOn system, the OPTO-BLUE system could allow pulsatile and quantitative activation of transgene expression in a light irradiance-dependent manner [
28]. Even a low intensity seems to be enough to activate the OPTO-BLUE system. The low blue-light intensity can induce the covalent cysteinyl-flavin adduct, which allows the dimerization of GAVPO [
29]. This characteristic has not been fully studied, but it is important for the therapeutic applications of this technology, since the lower the intensity required for gene activation, the less energy will be required by the light-generating devices to be utilized in a clinical setting.
To our knowledge, this is the first, fully integrated, lentiviral Light-On vector system to be functional. The arrangement of the genetic elements permits the induction of the GOI with a low-background expression. The improved Light-On platform described here is an improved version of the two-component lentiviral platform developed previously [
21], that simplified its use in many biological settings. Despite this simplification, some background expression in the absence of light is observed that could be avoided using a larger separation between the promoters. We suggest that a 0.5 kb separation should be optimal for most of the GOI to be used, but when a tighter control of the background is required, a larger separation of 2.0 kb is recommended. This configuration could be used with the improved versions of GAVPO [
30], permitting a lower background expression that could be used in an experimental or therapeutic setting.
4. Materials and Methods
4.1. Lentiviral Plasmid Construction
We previously developed pFUG-1D/2A-HA-GAVPO-W (12 kb) plasmid that expresses EGFP and GAVPO under the Ubiquitin constitutive promoter, and pF-UAS(s)-mCherry-W (10 kb) plasmid that expresses mCherry under the 5xUASG-TATA (UAS) inducible promoter. Standard molecular cloning and recombinant DNA techniques were used for plasmid construction. Lentiviral OPTO-BLUE expression plasmids were assembled from the FUGW plasmid [
22], as detailed below: the GAVPO sequence was obtained from pGAVPO plasmid using
PacI and
BsrGI restriction enzymes (New England Biolabs, Ipswich, MA, USA). Then, the EGFP sequence from the FUGW plasmid was replaced by the GAVPO sequence to obtain pFU-GAVPO-W (11 kb). This plasmid was digested with
NspI enzyme (New England Biolabs, Ipswich, MA, USA) to reduce the size. The reduced version of pFU-GAVPO-W (psFU-GAVPO-W) was achieved by releasing expendable sequences (~2 kb). The 9 kb psFU-GAVPO-W plasmid was used to construct the first version of pOPTO-BLUE. The UAS inducible promoter with a multiple cloning site (UAS-MCS) was obtained from pUAS-mcherry plasmid by PCR using the following primers sets flanked by
AsiSI and
PacI restriction sites: FP_AsiSI_UAS: 5’-GCGTTAATTAAAAGTGCAGGTGCCAGAAC-3′ and RP_PacI_UAS: 5′-CGCGCCGCGGGCGATCGCTAAGATACATTGATGAGT-3′. Then, the psFU-GAVPO-W plasmid was linearized using the
PacI enzyme (New England Biolabs, Ipswich, MA, USA), and the UAS-MCS amplicon previously treated with the
AsiSI and
PacI enzymes were introduced. A restriction-enzyme assay was performed to corroborate the insert orientation, obtaining the pOPTO-BLUE bidirectional plasmid. This plasmid expresses GAVPO driven by the Ubiquitin promoter in one direction and a GOI driven by the UAS inducible promoter in the opposite direction. From the pOPTO-BLUE plasmid, the pOPTO-BLUE-EGFP plasmids were constructed. To achieve this, the coding sequence for EGFP was obtained from pFUGW plasmid by PCR using the following primer sets flanked by
AscI and
BstBI restriction sites: FP_AscI_GFP: 5′-ATATGGCGCGCCATGGTGAGCAAGG-3′ and RP_BstBI_GFP: 5′-CGCGTTCGAACTACTTGTACAGCTCGT-3′. Then, the EGFP amplicon was introduced between
AscI and
BstBI restriction sites in the pOPTO-BLUE plasmid to obtain pOPTO-BLUE-EGFP-V1.
To reduce the size of the pOPTO-BLUE-EGFP-V1 plasmid even more, the Ubiquitin promoter was replaced by the EFS promoter to obtain pOPTO-BLUE-EGFP-V2. The EFS sequence was obtained by PCR from the pLenti-CRISPR-Cas9-P2A-mCherry plasmid using the following sets of primers: FP_EFS: 5′-ATATTTAATTAATGGCTCCGGT-3′ and RP_EFS: 5′-TGGCAGCGCTCTAGAACCGGT-3′. The EFS amplicon was treated with AgeI and PacI enzymes (New England Biolabs, Ipswich, MA, USA) and introduced into the pOPTO-BLUE-V1 plasmid between AgeI and PacI restriction sites. The new plasmid version, constructed with an EFS promoter, was named pOPTO-BLUE-EGFP-V2. Moreover, a pOPTO-BLUE-EGFP-V2_W/O plasmid without Ubiquitin/EFS promoter was generated. Since the Ubiquitin sequence is flanked by PacI sites, its extraction was performed by enzymatic digestion with PacI. Then, the ligation of their compatible ends was performed. Finally, different sizes of spacer sequences were introduced into the pOPTO-BLUE-EGFP-V2 plasmid to generate pOPTO-BLUE-EGFP-V3. DNA spacers of 200, 500, 1000, and 2000 bp were obtained from pLenti CRISPR-Cas9-P2A-mCherry, using the following set of primers: FP_200bp: 5′-CGCTTAATTAAGCTGTACGAGTACTTCAC-3′ and RP_200bp: 5′-CGCTTAATTAAAGATTTCCACGGAGTCGA-3′; FP_500bp: 5′-CGCTTAATTAAGCTGTACGAGTACTTCAC-3′and RP_500bp: 5′-CGCTTAATTAAAGATTTCCACGGAGTCGA-3′; FP_1000bp: 5′-CGCTTAATTAAGCTGTACGAGTACTTCAC-3′ and RP_1000bp: 5′-CGCTTAATTAAGTCGATGGAGTCGTCCT-3′; FP_2000bp: 5′-CGCTTAATTAAGCTGTACGAGTACTTCAC-3′ and RP_2000bp: 5′-CGCTTAATTAACTCGAAGCTGCTTCTTTC-3′). The plasmid versions obtained were pOPTO-BLUE-EGFP-V3_200bp, pOPTO-BLUE-EGFP-V3_500bp, pOPTO-BLUE-EGFP-V3_1000bp, pOPTO-BLUE-EGFP-V3_2000pb (pOPTO-BLUE-EGFP-V3 + spacers).
4.2. Cell Culture and Transfection
HEK293-T cells were cultured in DMEM (Dulbecco’s modified Eagle’s medium), supplemented with 10% (v/v) fetal-bovine serum, 100 U/mL penicillin, and 100 μg/mL streptomycin and L-glutamine (2 mM). All culture reagents were from Gibco. Cell cultures were maintained at 37 °C in an atmosphere of 95% air and 5% CO2. Cells were transfected with a Calfectin agent following the manufacturer’s recommendations (Calbiotech, El Cajon, CA, USA) using the following plasmid: psFEGW, pOPTO-BLUE-EGFP-V1, pOPTO-BLUE-EGFP-V2, pOPTO-BLUE-EGFP-V2_W/O, and pOPTO-BLUE-EGFP-V3 + spacers.
4.3. Lentivirus Production and Transduction
HEK293-T cells were transfected with Lipofectamine 2000 (Thermo Fisher Scientific, Waltham, MA, USA) using the packaging plasmids pCMV-ΔR8.9, pCMV-VSVg, and the corresponding transfer plasmid pOPTO-BLUE-EGFP-V3_500bp to produce a functional lentivirus, as described previously [
22]. HEK293-T cells were grown in Dulbecco’s Modified Eagle Medium DMEM supplemented with 10% fetal-bovine serum and 100 U/mL penicillin, and 100 μg/mL streptomycin (Thermo Fisher Scientific, Waltham, MA, USA). Lentiviral particles were collected from the supernatants 48 h after transfection and the lentiviral particles were centrifuged, passed through a 0.45-μm filter, and stored at −80 °C. Briefly, transduction was performed by adding 250 μL of lentiviral particles to HEK293-T cells, previously cultivated in 24 well plates. HEK293-T cell transduction was determined by fluorescence analysis and Western blot of EGFP.
4.4. Blue Light Induction Assay
After transfection or transduction, a group of plates with HEK293-T cells was maintained in darkness, while other groups of plates were exposed to blue light using a light intensity of 160 lux at 37 °C with 5% CO2. For all the induction experiments (except where indicated), cells were transfected for 8 h followed by 16 h of continuous illumination with blue light (LED, 460 nm). To perform time exposure of blue light experiments, 8 h post-transfection, different plates were exposed to continuous blue-light illumination for 0.25, 0.5, 1, 2, 4, 6, 8, 9, or 24 h. For light-intensity experiments, cells were transfected for 8 h and induced with blue light for 16 h at different light intensities (20, 40, 60, 80, and 160 lux). Finally, for transduction experiments, 76 h after transduction with lentiviruses, cells were exposed to 16 h of continuous illumination with blue light.
4.5. Western Blotting Analysis
For immunoblotting experiments, cell cultures were homogenized with a lysis buffer (50 mM sodium acetate, 150 mM sodium chloride, 10% glycerol (
v/
v), dH
2O) containing protein inhibitors, then the lysate was centrifuged at 10,000×
g for 5 min to remove debris. The lysate supernatant was preserved at −80 °C. Protein concentration was estimated by the DC protein assay method (Bio-Rad). The equivalent of 30 µg of protein extract was separated by SDS-PAGE and then transferred to a nitrocellulose or polyvinylidene difluoride (PVDF) membrane (GE Healthcare Bio-Sciences Corpstate, Piscataway, NJ, USA). Membranes were treated with a blocking buffer (3% Bovine Serum Albumin (BSA)) for 1 h at room temperature (RT). Then, membranes were probed with the appropriate primary antibodies, 2 h at RT for anti-GAPDH or overnight at 4 °C for anti-GFP. Then, the membranes were washed with a Tris Buffer Solution with 0.1% Tween (TBS-T buffer) and incubated with HRP-conjugated secondary anti-mouse/rabbit IgG antibodies (1:5000, Li-COR) for 1 h at RT. The blots were revealed by an enhanced ECL chemiluminescence system (LICOR detection system) and detected by a C-Digit Blot Scanner (Li-COR). Bands were quantified by densitometry using Fiji software Version 1.53t24 (NIH, Bethesda, MD, USA). The primary antibodies included an anti-GFP rabbit polyclonal antibody (A-11122, 1:2000, Thermo Fisher Scientific, Waltham, MA, USA), anti-GAPDH mouse monoclonal antibody (sc-32233, 1:2000, Santa Cruz Biotechnology, Dallas, TX, USA), and anti-HA mouse monoclonal antibody (sc-7392, 1:2000, Santa Cruz Biotechnology, Dallas, TX, USA) [
31].
4.6. Epifluorescence Microscopy Imaging
Cells were fixed using 4% paraformaldehyde for 10 min at room temperature. After fixation, three washes with PBS 1× solution were performed, and a final wash in distilled water was made to remove excess salt. Finally, the coverslips were mounted with a Vectashield/DAPI solution (Vector). Images were acquired in dark and light conditions with a DS-Fi2 epifluorescence microscope (Olympus, Shinjuku-ku, Tokyo, Japan) equipped with a Nikon DS-Fi2 camera (Nikon, Minato-ku, Tokyo, Japan) operated with QCapture Suite PLUS 3.1.3.10 (Q-Imaging).
4.7. Statistical Analysis
The data obtained with the Fiji program were analyzed and processed with the GraphPad Prism 8 program. The values shown in the figures represent the mean ± SEM (Standard Error of the Mean) results obtained from at least three independent experiments. The Shapiro–Wilk test for normality was used to determine whether parametric or nonparametric statistical tests were to be used. When two or more experimental groups with one factor were compared, the one-way analysis of variance (“one-way ANOVA”) was used, followed by Dunnet’s multiple-range comparison as a post-hoc test. A two-way ANOVA with Dunn’s analysis as a post-hoc test was used to compare data of multiple groups with two independent factors. The differences between the experimental groups were considered statistically significant from a confidence level greater than 95% (p < 0.05).



 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





