
Selective Activation of D3 Dopamine Receptors Ameliorates DOI-Induced Head Twitching Accompanied by Changes in Corticostriatal Processing

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
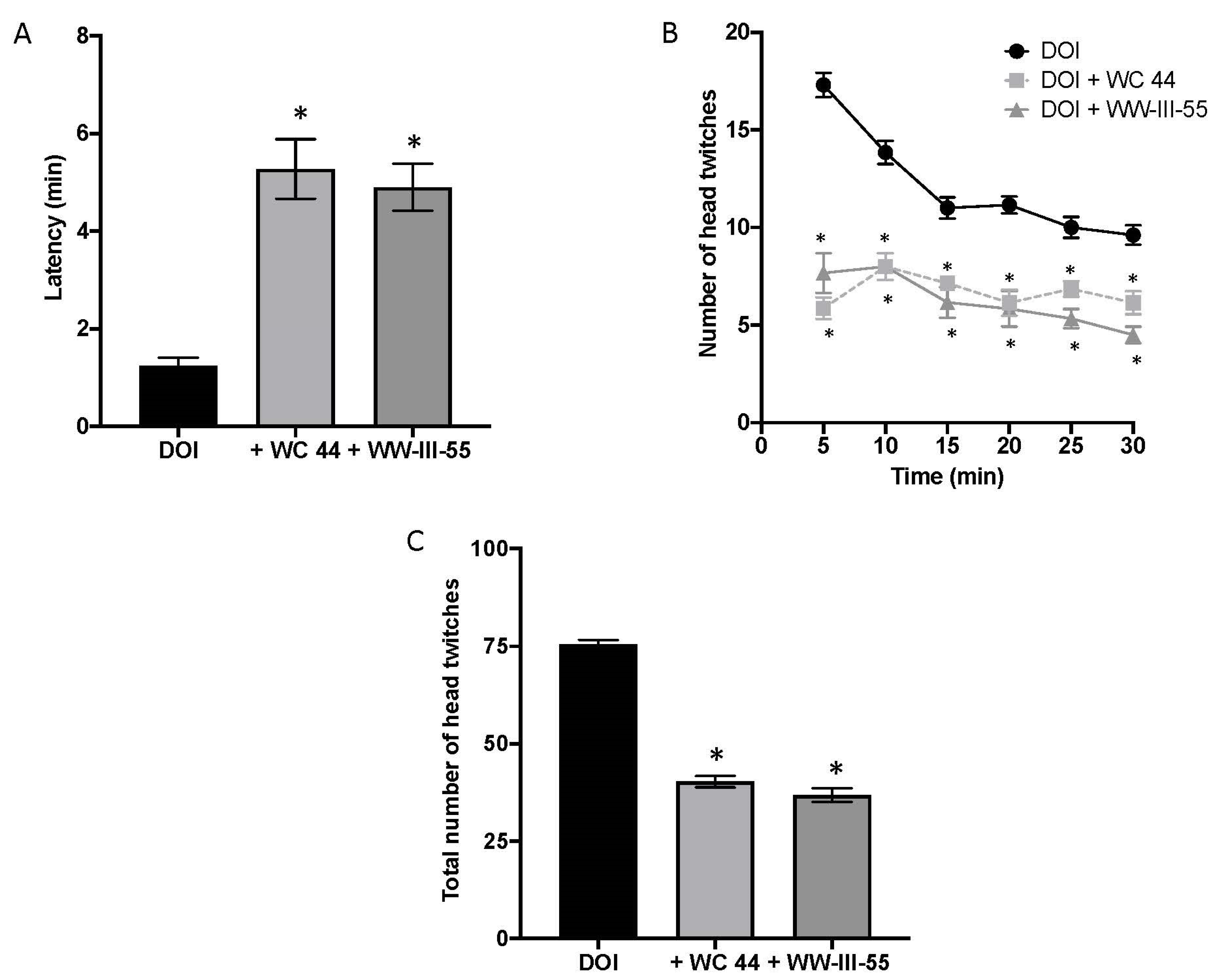
2.1. Dopamine D3 Receptor Agonists Reduce DOI-Induced Head Twitching
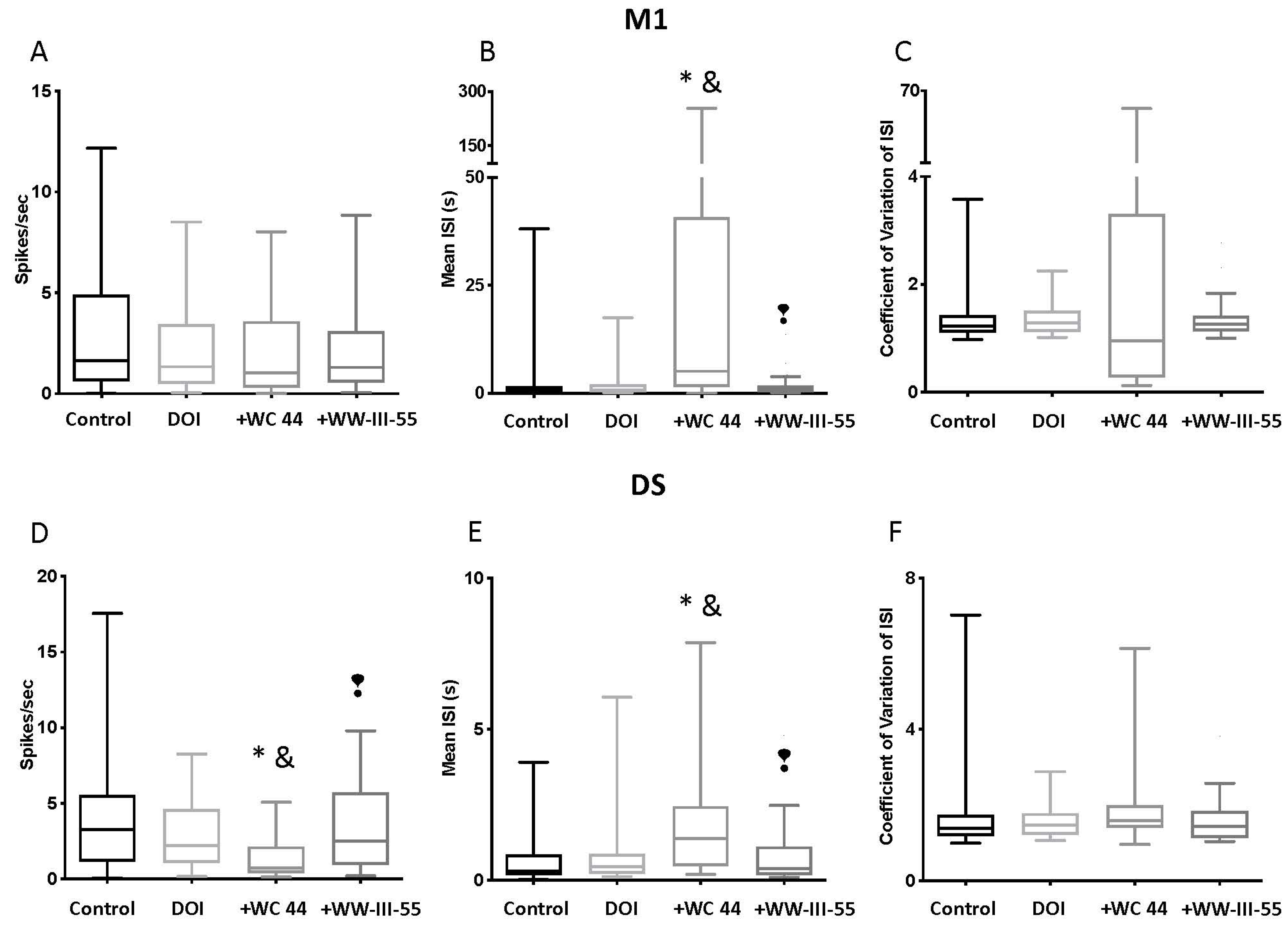
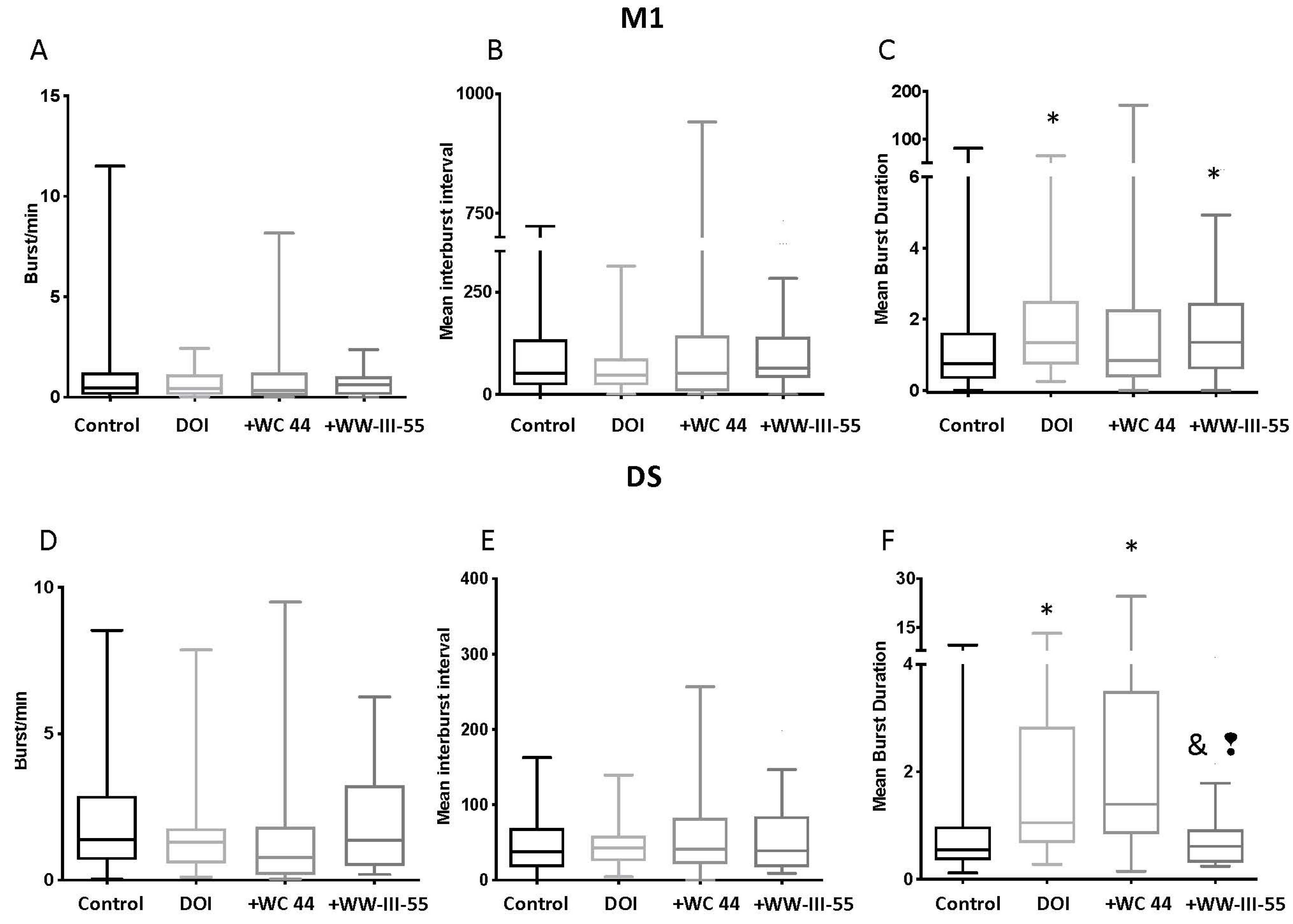
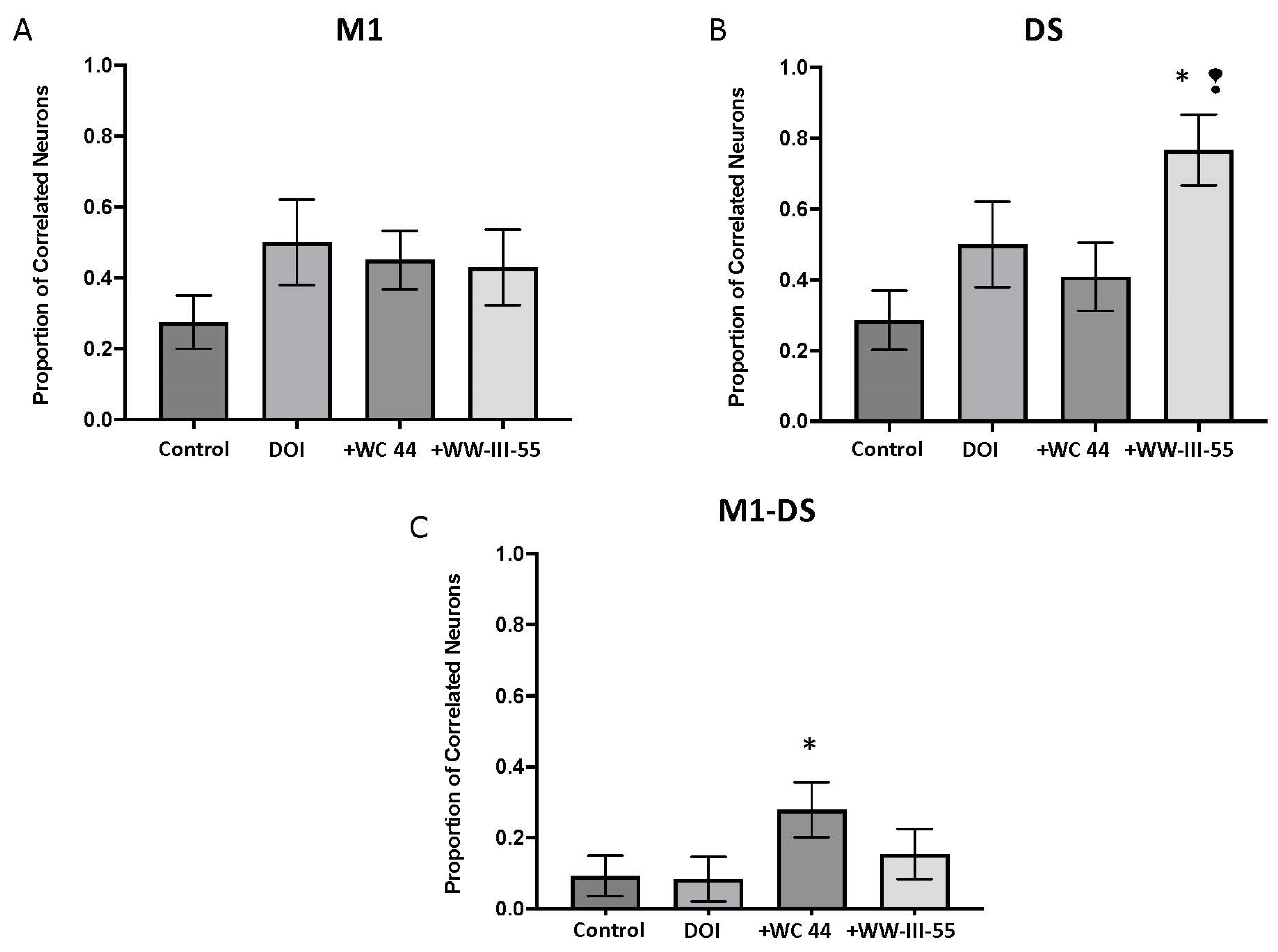
2.2. Effect of D3 Receptor Agonists on DOI-Induced Changes in M1 and DS Spike Activity
3. Discussion
4. Materials and Methods
4.1. Animal Care and Housing
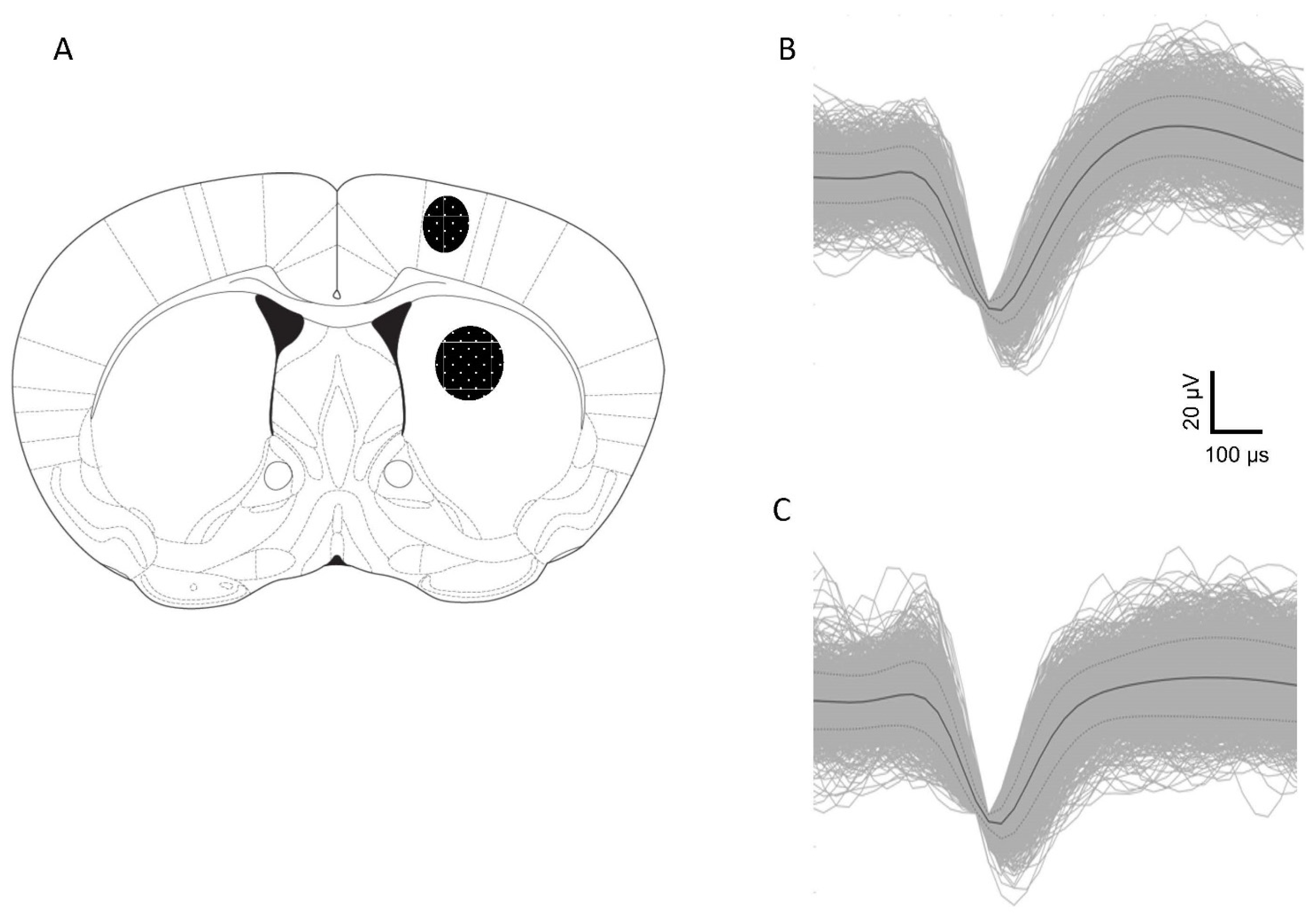
4.2. Electrodes and Implantation Surgery
4.3. Drugs and Experimental Protocol
4.4. Behavioral Electrophysiology
4.5. Spike-Train Analysis
4.6. Histology
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| DOI | 2,5-dimethoxy-4-iodoamphetamine |
| DS | Dorsal striatum |
| CV ISI | Coefficient of variation in inter-spike intervals |
| M1 | cortical motor area 1 |
| WC 44 | 4-(2-fluoroethyl)-N-[4-[4-(2-methoxyphenyl)piperazin 1-yl]butyl]benzamide |
| WW-III-55 | N-(4-(4-(4-methoxyphenyl)piperazin-1-yl)butyl)-4-(thiophen-3 yl)benzamide |
References
- Jinnai, K.; Matsuda, Y. Neurons of the motor cortex projecting commonly on the caudate nucleus and the lower brain stem in the cat. Neurosci. Lett. 1979, 13, 121–126. [Google Scholar] [CrossRef]
- McGeorge, A.J.; Faull, R.L. The organization of the projection from the cerebral cortex to the striatum in the rat. Neuroscience 1989, 29, 503–537. [Google Scholar] [CrossRef] [PubMed]
- Leckman, J.F.; Bloch, M.H.; Smith, M.E.; Larabi, D.; Hampson, M. Neurobiological substrates of Tourette’s disorder. J. Child Adolesc. Psychopharmacol. 2010, 20, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Pogorelov, V.; Xu, M.; Smith, H.R.; Buchanan, G.F.; Pittenger, C. Corticostriatal interactions in the generation of tic-like behaviors after local striatal disinhibition. Exp. Neurol. 2015, 265, 122–128. [Google Scholar] [CrossRef]
- Julius, A.; Longfellow, K. Movement Disorders: A Brief Guide in Medication Management. Med. Clin. N. Am. 2016, 100, 733–761. [Google Scholar] [CrossRef]
- Bloch, M.H. Emerging treatments for Tourette’s disorder. Curr. Psychiatry Rep. 2008, 10, 323–330. [Google Scholar] [CrossRef]
- Neve, K.A.; Seamans, J.K.; Trantham-Davidson, H. Dopamine receptor signaling. J. Recept. Signal Transduct. Res. 2004, 24, 165–205. [Google Scholar] [CrossRef]
- Bouthenet, M.L.; Souil, E.; Martres, M.P.; Sokoloff, P.; Giros, B.; Schwartz, J.C. Localization of dopamine D3 receptor mRNA in the rat brain using in situ hybridization histochemistry: Comparison with dopamine D2 receptor mRNA. Brain Res. 1991, 564, 203–219. [Google Scholar] [CrossRef] [PubMed]
- Surmeier, D.J.; Song, W.J.; Yan, Z. Coordinated expression of dopamine receptors in neostriatal medium spiny neurons. J. Neurosci. 1996, 16, 6579–6591. [Google Scholar] [CrossRef]
- Kassel, S.; Schwed, J.S.; Stark, H. Dopamine D3 receptor agonists as pharmacological tools. Eur. Neuropsychopharmacol. 2015, 25, 1480–1499. [Google Scholar] [CrossRef]
- Le Foll, B.; Collo, G.; Rabiner, E.A.; Boileau, I.; Merlo Pich, E.; Sokoloff, P. Dopamine D3 receptor ligands for drug addiction treatment: Update on recent findings. Prog. Brain Res. 2014, 211, 255–275. [Google Scholar] [CrossRef] [PubMed]
- Bordet, R.; Ridray, S.; Schwartz, J.C.; Sokoloff, P. Involvement of the direct striatonigral pathway in levodopa-induced sensitization in 6-hydroxydopamine-lesioned rats. Eur. J. Neurosci. 2000, 12, 2117–2123. [Google Scholar] [CrossRef] [PubMed]
- Millan, M.J.; Seguin, L.; Gobert, A.; Cussac, D.; Brocco, M. The role of dopamine D3 compared with D2 receptors in the control of locomotor activity: A combined behavioural and neurochemical analysis with novel, selective antagonists in rats. Psychopharmacology 2004, 174, 341–357. [Google Scholar] [CrossRef] [PubMed]
- Yarkov, A.V.; Hanger, D.; Reploge, M.; Joyce, J.N. Behavioral effects of dopamine agonists and antagonists in MPTP-lesioned D3 receptor knockout mice. Pharmacol. Biochem. Behav. 2003, 76, 551–562. [Google Scholar] [CrossRef]
- Rangel-Barajas, C.; Malik, M.; Mach, R.H.; Luedtke, R.R. Pharmacological modulation of abnormal involuntary DOI-induced head twitch response movements in male DBA/2J mice: II. Effects of D3 dopamine receptor selective compounds. Neuropharmacology 2015, 93, 179–190. [Google Scholar] [CrossRef]
- Tizabi, Y.; Russell, L.T.; Johnson, M.; Darmani, N.A. Nicotine attenuates DOI-induced head-twitch response in mice: Implications for Tourette syndrome. Prog. Neuropsychopharmacol. Biol. Psychiatry 2001, 25, 1445–1457. [Google Scholar] [CrossRef]
- Nespoli, E.; Rizzo, F.; Boeckers, T.M.; Hengerer, B.; Ludolph, A.G. Addressing the Complexity of Tourette’s Syndrome through the Use of Animal Models. Front. Neurosci. 2016, 10, 133. [Google Scholar] [CrossRef]
- Rangel-Barajas, C.; Estrada-Sanchez, A.M.; Barton, S.J.; Luedtke, R.R.; Rebec, G.V. Dysregulated corticostriatal activity in open-field behavior and the head-twitch response induced by the hallucinogen 2,5-dimethoxy-4-iodoamphetamine. Neuropharmacology 2017, 113, 502–510. [Google Scholar] [CrossRef]
- Larson, E.R.; Ariano, M.A. D3 and D2 dopamine receptors: Visualization of cellular expression patterns in motor and limbic structures. Synapse 1995, 20, 325–337. [Google Scholar] [CrossRef]
- Peters, S.K.; Dunlop, K.; Downar, J. Cortico-Striatal-Thalamic Loop Circuits of the Salience Network: A Central Pathway in Psychiatric Disease and Treatment. Front. Syst. Neurosci. 2016, 10, 104. [Google Scholar] [CrossRef]
- Miller, B.R.; Walker, A.G.; Barton, S.J.; Rebec, G.V. Dysregulated Neuronal Activity Patterns Implicate Corticostriatal Circuit Dysfunction in Multiple Rodent Models of Huntington’s Disease. Front. Syst. Neurosci. 2011, 5, 26. [Google Scholar] [CrossRef] [PubMed]
- Donzis, E.J.; Estrada-Sanchez, A.M.; Indersmitten, T.; Oikonomou, K.; Tran, C.H.; Wang, C.; Latifi, S.; Golshani, P.; Cepeda, C.; Levine, M.S. Cortical Network Dynamics Is Altered in Mouse Models of Huntington’s Disease. Cereb. Cortex 2020, 30, 2372–2388. [Google Scholar] [CrossRef] [PubMed]
- Uhlhaas, P.J.; Singer, W. Neural synchrony in brain disorders: Relevance for cognitive dysfunctions and pathophysiology. Neuron 2006, 52, 155–168. [Google Scholar] [CrossRef]
- Oh, T.; Daadi, E.S.; Kim, J.; Daadi, E.W.; Chen, P.J.; Roy-Choudhury, G.; Bohmann, J.; Blass, B.E.; Daadi, M.M. Dopamine D3 receptor ligand suppresses the expression of levodopa-induced dyskinesia in nonhuman primate model of parkinson’s disease. Exp. Neurol. 2022, 347, 113920. [Google Scholar] [CrossRef]
- Sato, K.; Ueda, H.; Okumura, F.; Misu, Y. 6-OHDA-induced lesion of the nigrostriatal dopaminergic neurons potentiates the inhibitory effect of 7-OHDPAT, a selective D3 agonist, on acetylcholine release during striatal microdialysis in conscious rats. Brain Res. 1994, 655, 233–236. [Google Scholar] [CrossRef]
- Castro-Hernandez, J.; Afonso-Oramas, D.; Cruz-Muros, I.; Salas-Hernandez, J.; Barroso-Chinea, P.; Moratalla, R.; Millan, M.J.; Gonzalez-Hernandez, T. Prolonged treatment with pramipexole promotes physical interaction of striatal dopamine D3 autoreceptors with dopamine transporters to reduce dopamine uptake. Neurobiol. Dis. 2015, 74, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Cuellar-Santoyo, A.O.; Ruiz-Rodriguez, V.M.; Mares-Barbosa, T.B.; Patron-Soberano, A.; Howe, A.G.; Portales-Perez, D.P.; Miquelajauregui Graf, A.; Estrada-Sanchez, A.M. Revealing the contribution of astrocytes to glutamatergic neuronal transmission. Front. Cell. Neurosci. 2022, 16, 1037641. [Google Scholar] [CrossRef]
- Lyon, K.A.; Allen, N.J. From Synapses to Circuits, Astrocytes Regulate Behavior. Front. Neural Circuits 2021, 15, 786293. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, I.; Asanuma, M.; Diaz-Corrales, F.J.; Miyoshi, K.; Ogawa, N. Direct evidence for expression of dopamine receptors in astrocytes from basal ganglia. Brain Res. 2004, 1029, 120–123. [Google Scholar] [CrossRef]
- Estrada-Sanchez, A.M.; Burroughs, C.L.; Cavaliere, S.; Barton, S.J.; Chen, S.; Yang, X.W.; Rebec, G.V. Cortical efferents lacking mutant huntingtin improve striatal neuronal activity and behavior in a conditional mouse model of Huntington’s disease. J. Neurosci. 2015, 35, 4440–4451. [Google Scholar] [CrossRef]
- Paxinos, G.; Franklin, K.B.J. The Mouse Brain in Stereotaxic Coordinates; Academic: San Diego, CA, USA; London, UK, 2001. [Google Scholar]
- Tu, Z.; Li, S.; Cui, J.; Xu, J.; Taylor, M.; Ho, D.; Luedtke, R.R.; Mach, R.H. Synthesis and pharmacological evaluation of fluorine-containing D(3) dopamine receptor ligands. J. Med. Chem. 2011, 54, 1555–1564. [Google Scholar] [CrossRef] [PubMed]
- Cheung, T.H.; Loriaux, A.L.; Weber, S.M.; Chandler, K.N.; Lenz, J.D.; Schaan, R.F.; Mach, R.H.; Luedtke, R.R.; Neisewander, J.L. Reduction of cocaine self-administration and D3 receptor-mediated behavior by two novel dopamine D3 receptor-selective partial agonists, OS-3-106 and WW-III-55. J. Pharmacol. Exp. Ther. 2013, 347, 410–423. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Sanchez, A.M.; Barton, S.J.; Burroughs, C.L.; Doyle, A.R.; Rebec, G.V. Dysregulated striatal neuronal processing and impaired motor behavior in mice lacking huntingtin interacting protein 14 (HIP14). PLoS ONE 2013, 8, e84537. [Google Scholar] [CrossRef] [PubMed]
- Lewicki, M.S. A review of methods for spike sorting: The detection and classification of neural action potentials. Network 1998, 9, R53–R78. [Google Scholar] [CrossRef]
- Legendy, C.R.; Salcman, M. Bursts and recurrences of bursts in the spike trains of spontaneously active striate cortex neurons. J. Neurophysiol. 1985, 53, 926–939. [Google Scholar] [CrossRef]
- Miller, B.R.; Walker, A.G.; Shah, A.S.; Barton, S.J.; Rebec, G.V. Dysregulated information processing by medium spiny neurons in striatum of freely behaving mouse models of Huntington’s disease. J. Neurophysiol. 2008, 100, 2205–2216. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Estrada-Sánchez, A.M.; Rangel-Barajas, C.; Howe, A.G.; Barton, S.J.; Mach, R.H.; Luedtke, R.R.; Rebec, G.V. Selective Activation of D3 Dopamine Receptors Ameliorates DOI-Induced Head Twitching Accompanied by Changes in Corticostriatal Processing. Int. J. Mol. Sci. 2023, 24, 9300. https://doi.org/10.3390/ijms24119300
Estrada-Sánchez AM, Rangel-Barajas C, Howe AG, Barton SJ, Mach RH, Luedtke RR, Rebec GV. Selective Activation of D3 Dopamine Receptors Ameliorates DOI-Induced Head Twitching Accompanied by Changes in Corticostriatal Processing. International Journal of Molecular Sciences. 2023; 24(11):9300. https://doi.org/10.3390/ijms24119300
Chicago/Turabian StyleEstrada-Sánchez, Ana María, Claudia Rangel-Barajas, Andrew G. Howe, Scott J. Barton, Robert H. Mach, Robert R. Luedtke, and George V. Rebec. 2023. "Selective Activation of D3 Dopamine Receptors Ameliorates DOI-Induced Head Twitching Accompanied by Changes in Corticostriatal Processing" International Journal of Molecular Sciences 24, no. 11: 9300. https://doi.org/10.3390/ijms24119300
APA StyleEstrada-Sánchez, A. M., Rangel-Barajas, C., Howe, A. G., Barton, S. J., Mach, R. H., Luedtke, R. R., & Rebec, G. V. (2023). Selective Activation of D3 Dopamine Receptors Ameliorates DOI-Induced Head Twitching Accompanied by Changes in Corticostriatal Processing. International Journal of Molecular Sciences, 24(11), 9300. https://doi.org/10.3390/ijms24119300