
Intronic Enhancer Is Essential for Nr5a1 Expression in the Pituitary Gonadotrope and for Postnatal Development of Male Reproductive Organs in a Mouse Model

 , ,
, ,
Abstract
1. Introduction
2. Results
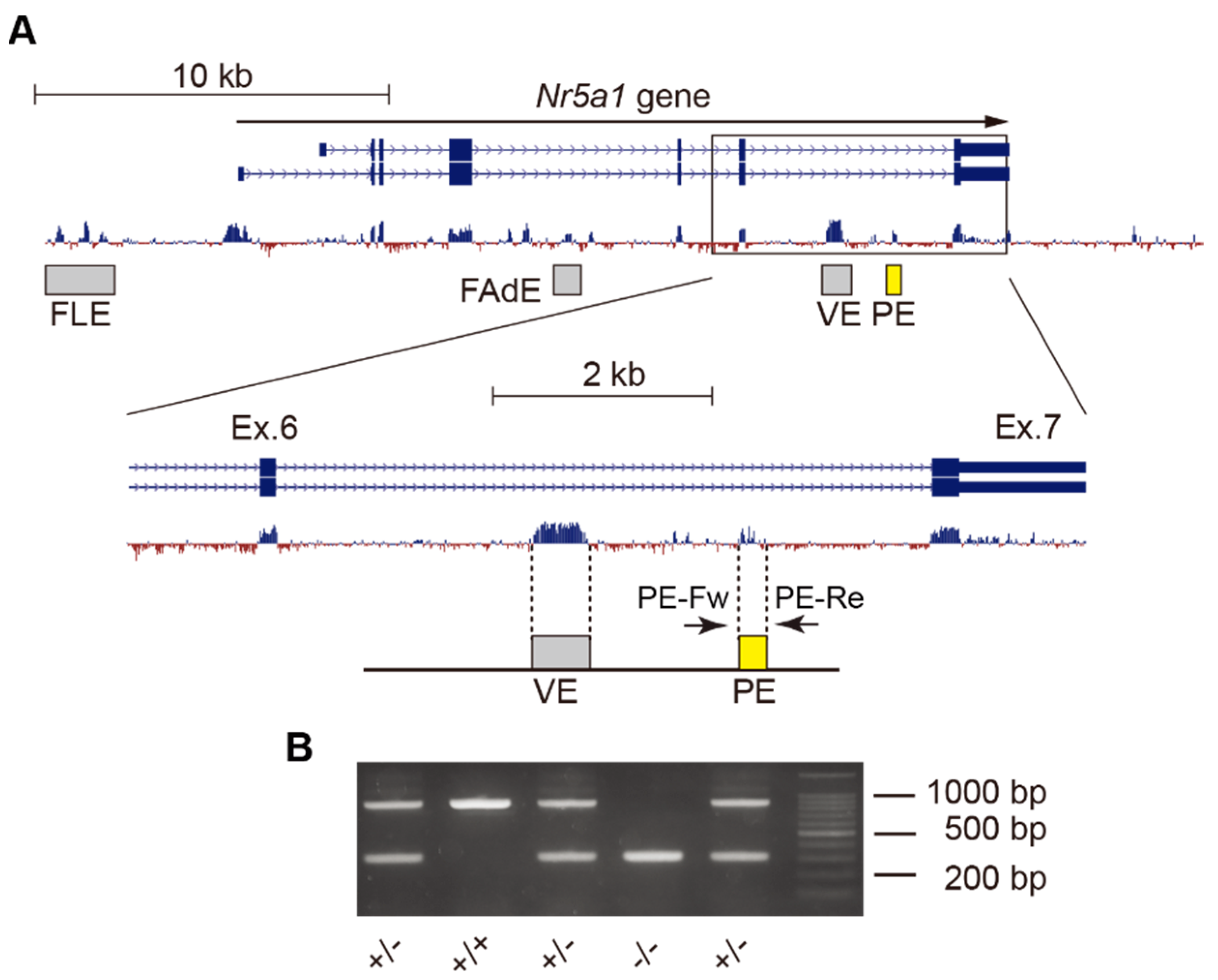
2.1. Deletion of Gonadotrope-Specific PE of Nr5a1
2.2. Normal Masculinization in Fetal Stages
2.2.1. Testis and Accessory Reproductive Organs
2.2.2. Steroids in Fetal Testes
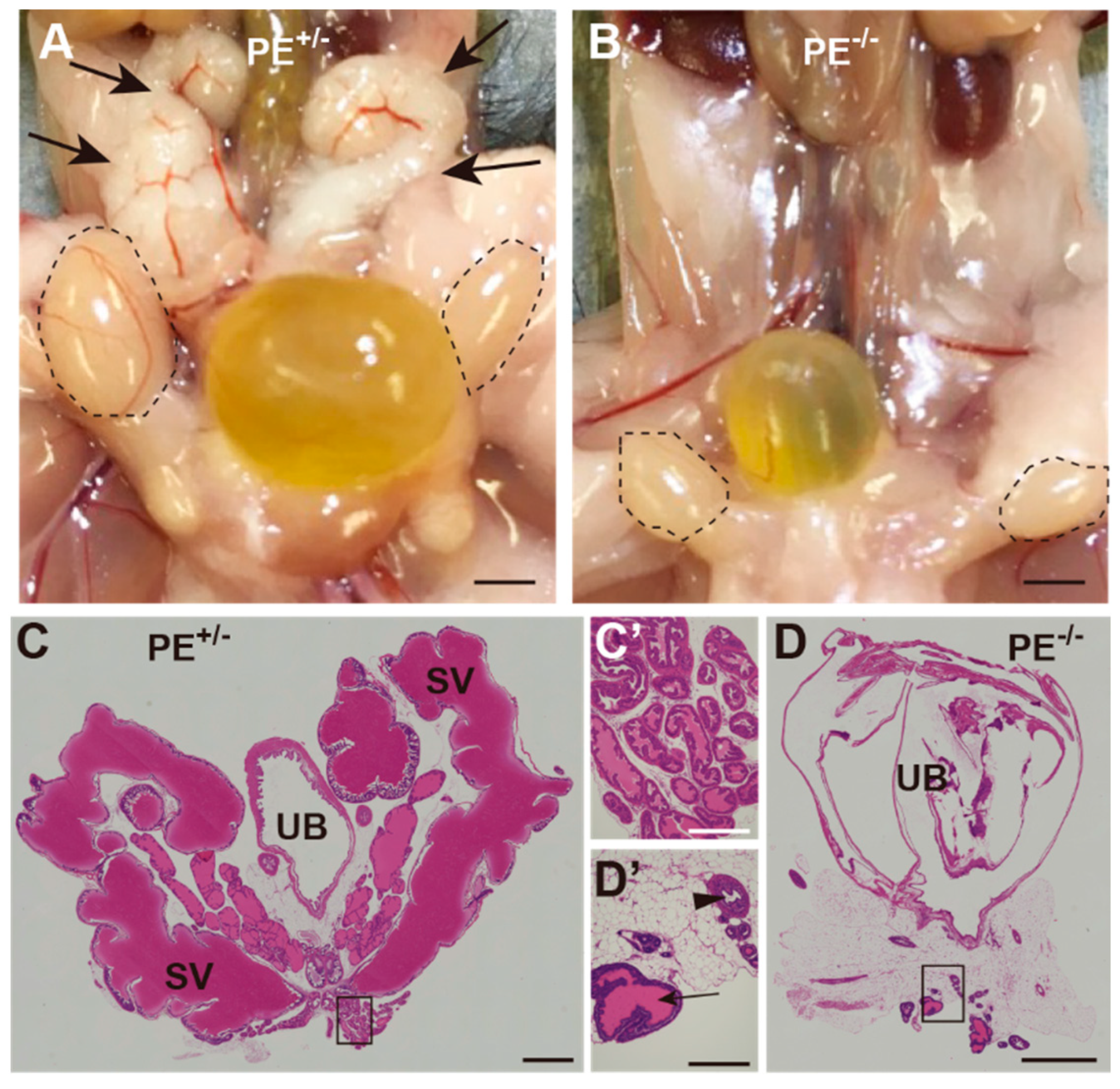
2.3. Impaired Development of Reproductive Organs at Adult Stages
2.3.1. Testis
2.3.2. Seminal Vesicles, Prostate Gland, and Penis
2.3.3. Steroid Levels in Adult Testes
2.3.4. Quantitative Reverse Transcription (qRT)-PCR and Gonadotropin Immunostaining
2.4. Transcriptome Analyses of Pituitaries and Isolated Gonadotropes
3. Discussion
3.1. Functional Importance of the PE in Nr5a1 Gene Regulation
3.2. Dependence of Fetal and Adult Leydig Cells on Pituitary Gonadotropins
3.3. Role of NR5A1 in the Pituitary Gonadotrope
3.4. Candidate NR5A1 Downstream Genes
4. Materials and Methods
4.1. Mice
4.2. Tissue Preparation, Histological Analyses, and Immunostaining
4.3. RNA Preparation and Quantitative RT-PCR
4.4. Measurement of Plasma Gonadotropin
4.5. Measurement of Testicular Steroids
4.6. mRNA Sequencing, Data Processing, and Differentially Expressed Gene Analyses
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ma, X.; Dong, Y.; Matzuk, M.M.; Kumar, T.R. Targeted disruption of luteinizing hormone beta-subunit leads to hypogonadism, defects in gonadal steroidogenesis, and infertility. Proc. Natl. Acad. Sci. USA 2004, 101, 17294–17299. [Google Scholar] [CrossRef] [PubMed]
- Lei, Z.M.; Mishra, S.; Zou, W.; Xu, B.; Foltz, M.; Li, X.; Rao, C.V. Targeted disruption of luteinizing hormone/human chorionic gonadotropin receptor gene. Mol. Endocrinol. 2001, 15, 184–200. [Google Scholar] [CrossRef] [PubMed]
- Kumar, T.R.; Wang, Y.; Lu, N.; Matzuk, M.M. Follicle stimulating hormone is required for ovarian follicle maturation but not male fertility. Nat. Genet. 1997, 15, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Morohashi, K. Gonadal and extragonadal functions of Ad4BP/SF-1: Developmental aspects. Trends Endocrinol. Metab. 1999, 10, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Parker, K.L.; Schimmer, B.P. Steroidogenic factor 1: A key determinant of endocrine development and function. Endocr. Rev. 1997, 18, 361–377. [Google Scholar] [CrossRef]
- Luo, X.; Ikeda, Y.; Lala, D.S.; Baity, L.A.; Meade, J.C.; Parker, K.L. A cell-specific nuclear receptor plays essential roles in adrenal and gonadal development. Endocr. Res. 1995, 21, 517–524. [Google Scholar] [CrossRef]
- Zhao, L.; Bakke, M.; Krimkevich, Y.; Cushman, L.J.; Parlow, A.F.; Camper, S.A.; Parker, K.L. Steroidogenic factor 1 (SF1) is essential for pituitary gonadotrope function. Development 2001, 128, 147–154. [Google Scholar] [CrossRef]
- Zhao, L.; Bakke, M.; Parker, K.L. Pituitary-specific knockout of steroidogenic factor 1. Mol. Cell Endocrinol. 2001, 185, 27–32. [Google Scholar] [CrossRef]
- Zubair, M.; Ishihara, S.; Oka, S.; Okumura, K.; Morohashi, K. Two-step regulation of Ad4BP/SF-1 gene transcription during fetal adrenal development: Initiation by a Hox-Pbx1-Prep1 complex and maintenance via autoregulation by Ad4BP/SF-1. Mol. Cell Biol. 2006, 26, 4111–4121. [Google Scholar] [CrossRef]
- Shima, Y.; Zubair, M.; Ishihara, S.; Shinohara, Y.; Oka, S.; Kimura, S.; Okamoto, S.; Minokoshi, Y.; Suita, S.; Morohashi, K. Ventromedial hypothalamic nucleus-specific enhancer of Ad4BP/SF-1 gene. Mol. Endocrinol. 2005, 19, 2812–2823. [Google Scholar] [CrossRef]
- Shima, Y.; Zubair, M.; Komatsu, T.; Oka, S.; Yokoyama, C.; Tachibana, T.; Hjalt, T.A.; Drouin, J.; Morohashi, K. Pituitary homeobox 2 regulates adrenal 4 binding protein/steroidogenic factor-1 gene transcription in the pituitary gonadotrope through interaction with the intronic enhancer. Mol. Endocrinol. 2008, 22, 1633–1646. [Google Scholar] [CrossRef] [PubMed]
- Shima, Y.; Miyabayashi, K.; Baba, T.; Otake, H.; Katsura, Y.; Oka, S.; Zubair, M.; Morohashi, K. Identification of an enhancer in the Ad4BP/SF-1 gene specific for fetal Leydig cells. Endocrinology 2012, 153, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Shima, Y.; Miyabayashi, K.; Sato, T.; Suyama, M.; Ohkawa, Y.; Doi, M.; Okamura, H.; Suzuki, K. Fetal Leydig cells dedifferentiate and serve as adult Leydig stem cells. Development 2018, 145. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, Y.; Luo, X.; Abbud, R.; Nilson, J.H.; Parker, K.L. The Nuclear Receptor Steroidogenic Factor 1 Is Essential for the Formation of the Ventromedial Hypothalamic Nucleus. Mol. Endocrinol. 1995, 9, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Fowkes, R.C.; Desclozeaux, M.; Patel, M.V.; Aylwin, S.J.B.; King, P.; Ingraham, H.A.; Burrin, J.M. Steroidogenic Factor-1 and the Gonadotrope-Specific Element Enhance Basal and Pituitary Adenylate Cyclase-Activating Polypeptide-Stimulated Transcription of the Human Glycoprotein Hormone Alpha-Subunit Gene in Gonadotropes. Mol. Endocrinol. 2003, 17, 2177–2188. [Google Scholar] [CrossRef][Green Version]
- Kaiser, U.B.; Halvorson, L.M.; Chen, M.T. Sp1, Steroidogenic Factor 1 (SF-1), and Early Growth Response Protein 1 (Egr-1) Binding Sites Form a Tripartite Gonadotropin-Releasing Hormone Response Element in the Rat Luteinizing Hormone-Beta Gene Promoter: An Integral Role for SF-1. Mol. Endocrinol. 2000, 14, 1235–1245. [Google Scholar] [CrossRef][Green Version]
- Pacini, V.; Petit, F.; Querat, B.; Laverriere, J.N.; Cohen-Tannoudji, J.; L’Hôte, D. Identification of a pituitary ERα-activated enhancer triggering the expression of Nr5a1, the earliest gonadotrope lineage-specific transcription factor. Epigenet Chromatin 2019, 12, 48. [Google Scholar] [CrossRef]
- Chen, J.; Minabe, S.; Munetomo, A.; Magata, F.; Sato, M.; Nakamura, S.; Hirabayashi, M.; Ishihara, Y.; Yamazaki, T.; Uenoyama, Y.; et al. Kiss1-Dependent and Independent Release of Luteinizing Hormone and Testosterone in Perinatal Male Rats. Endocr. J. 2022, 69, 797–807. [Google Scholar] [CrossRef]
- Shima, Y. Development of Fetal and Adult Leydig Cells. Reprod. Med. Biol. 2019, 18, 323–330. [Google Scholar] [CrossRef]
- Shima, Y.; Matsuzaki, S.; Miyabayashi, K.; Otake, H.; Baba, T.; Kato, S.; Huhtaniemi, I.; Morohashi, K. Fetal Leydig cells persist as an androgen-independent subpopulation in the postnatal testis. Mol. Endocrinol. 2015, 29, 1581–1593. [Google Scholar] [CrossRef]
- Prince, F.P. The triphasic nature of Leydig cell development in humans, and comments on nomenclature. J. Endocrinol. 2001, 168, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Shima, Y. Functional importance of mini-puberty in spermatogenic stem cell formation. Front. Cell Dev. Biol. 2022, 10, 907989. [Google Scholar] [CrossRef] [PubMed]
- Bjelobaba, I.; Janjic, M.M.; Prévide, R.M.; Abebe, D.; Kucka, M.; Stojilkovic, S.S. Distinct expression patterns of osteopontin and dentin matrix protein 1 genes in pituitary gonadotrophs. Front. Endocrinol. 2019, 10, 248. [Google Scholar] [CrossRef] [PubMed]
- Rydze, R.T.; Patton, B.K.; Briley, S.M.; Salazar Torralba, H.; Gipson, G.; James, R.; Rajkovic, A.; Thompson, T.; Pangas, S.A. deletion of gremlin-2 alters estrous cyclicity and disrupts female fertility in mice. Biol. Reprod. 2021, 105, 1205–1220. [Google Scholar] [CrossRef] [PubMed]
- Brûlé, E.; Wang, Y.; Li, Y.; Lin, Y.-F.; Zhou, X.; Ongaro, L.; Alonso, C.A.I.; Buddle, E.R.S.; Schneyer, A.L.; Byeon, C.-H.; et al. TGFBR3L is an inhibin B co-receptor that regulates female fertility. Sci. Adv. 2021, 7, eabl4391. [Google Scholar] [CrossRef]
- Lin, Y.-F.; Schang, G.; Buddle, E.R.S.; Schultz, H.; Willis, T.L.; Ruf-Zamojski, F.; Zamojski, M.; Mendelev, N.; Boehm, U.; Sealfon, S.C.; et al. Steroidogenic factor 1 regulates transcription of the inhibin B coreceptor in pituitary gonadotrope cells. Endocrinology 2022, 163, bqac131. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.-J.; Wang, L. Bile acid-activated receptors: A review on FXR and other nuclear receptors. Handb. Exp. Pharmacol. 2019, 256, 51–72. [Google Scholar] [CrossRef]
- Ikeda, Y.; Takeda, Y.; Shikayama, T.; Mukai, T.; Hisano, S.; Morohashi, K.I. Comparative localization of Dax-1 and Ad4BP/SF-1 during development of the hypothalamic-pituitary-gonadal axis suggests their closely related and distinct functions. Dev. Dyn. 2001, 220, 363–376. [Google Scholar] [CrossRef]
- Kawabe, K.; Shikayama, T.; Tsuboi, H.; Oka, S.; Oba, K.; Yanase, T.; Nawata, H.; Morohashi, K. Dax-1 as one of the target genes of Ad4BP/SF-1. Mol. Endocrinol. 1999, 13, 1267–1284. [Google Scholar] [CrossRef]
- Yu, R.N.; Ito, M.; Jameson, J.L. The murine Dax-1 promoter is stimulated by SF-1 (steroidogenic factor-1) and inhibited by COUP-TF (chicken ovalbumin upstream promoter-transcription factor) via a composite nuclear receptor-regulatory element. Mol. Endocrinol. 1998, 12, 1010–1022. [Google Scholar] [CrossRef]
- Burris, T.P.; Guo, W.; Le, T.; McCabe, E.R. Identification of a putative steroidogenic factor-1 response element in the DAX-1 promoter. Biochem. Biophys. Res. Commun. 1995, 214, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Miyabayashi, K.; Tokunaga, K.; Otake, H.; Baba, T.; Shima, Y.; Morohashi, K. Heterogeneity of Ovarian Theca and Interstitial Gland Cells in Mice. PLoS ONE 2015, 10, e0128352. [Google Scholar] [CrossRef] [PubMed]
- Ikegami, K.; Goto, T.; Nakamura, S.; Watanabe, Y.; Sugimoto, A.; Majarune, S.; Horihata, K.; Nagae, M.; Tomikawa, J.; Imamura, T.; et al. Conditional Kisspeptin Neuron-Specific Kiss1 Knockout with Newly Generated Kiss1-Floxed and Kiss1-Cre Mice Replicates a Hypogonadal Phenotype of Global Kiss1 Knockout Mice. J. Reprod. Dev. 2020, 66, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Hirabayashi, M.; Watanabe, Y.; Sanbo, M.; Tomita, K.; Inoue, N.; Tsukamura, H.; Uenoyama, Y. Testosterone supplementation rescues spermatogenesis and in vitro fertilizing ability of sperm in Kiss1 knockout mice. Endocrinology 2020, 161. [Google Scholar] [CrossRef]
- Han, S.; Baba, T.; Yanai, S.; Byun, D.J.; Morohashi, K.-I.; Kim, J.-H.; Choi, M.H. GC-MS-based metabolic signatures reveal comparative steroidogenic pathways between fetal and adult mouse testes. Andrology 2021, 9, 400–406. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amount (pg/Whole Tissue) | |||||
|---|---|---|---|---|---|
| PE+/− | PE−/− | ||||
| Steroid Compound | Mean | SD | Mean | SD | p-Value 1 |
| Dehydroepiandrosterone | 28.4 | 3.1 | 24.5 | 8.5 | 0.5476 |
| Androstenediol | 36.7 | 1.2 | 39.7 | 8.7 | 0.5000 |
| Epitestosterone | 12.2 | 2.1 | 15.8 | 9.5 | 0.6905 |
| 5α-androstane-3β, 17β-diol | 18.7 | 11.2 | 14.9 | 1.8 | 0.6905 |
| Androstenedione | 324.0 | 133.2 | 381.4 | 156.0 | 0.5476 |
| Testosterone | 627.1 | 248.0 | 722.9 | 274.6 | 0.6905 |
| Pregnenolone | 18.8 | 4.6 | 15.5 | 0.6 | 0.2222 |
| Progesterone | 28.97 | 0.24 | 28.68 | 11.33 | 0.8413 |
| 17α-hydroxyprogesterone | 100.3 | 35.3 | 98.4 | 33.6 | >0.9999 |
| 11-deoxycortisol | 49.9 | 7.9 | 62.8 | 16.5 | 0.1508 |
| Concentration (pg/mg Tissue) | |||||
|---|---|---|---|---|---|
| PE+/− | PE−/− | ||||
| Steroid Compound | Mean | SD | Mean | SD | p-Value 1 |
| 3α-androstane-3α, 17β-diol | 2.1 | 0.3 | 2.3 | 1.2 | 0.6905 |
| Dehydroepiandrosterone | 0.6 | 0.1 | 1.8 | 0.4 | 0.0079 * |
| Androstenediol | 3.7 | 0.8 | 2.4 | 0.6 | 0.0317 * |
| Epitestosterone | 0.9 | 0.9 | 1.2 | 0.8 | >0.9999 |
| 3α-androstane-3β, 17β-diol | 1.1 | 0.5 | 1.6 | 0.5 | 0.2222 |
| 7α-hydroxyandrostenedione | 3.1 | 0.9 | 12.3 | 2.7 | 0.0079 * |
| Dihydrotestosterone | 1.7 | 0.8 | 2.3 | 0.5 | 0.2222 |
| Androstenedione | 54.0 | 21.7 | 10.9 | 5.5 | 0.0079 * |
| Testosterone | 58.3 | 57.4 | 9.7 | 8.1 | 0.0079 * |
| 6β-hydroxyandrostenedione | 1.3 | 0.3 | ND | ND | NA |
| 6β-hydroxytestosterone | 1.5 | 1.5 | ND | ND | NA |
| Pregnenolon | 4.7 | 1.1 | 2.8 | 2.0 | 0.0952 |
| Progesterone | 8.61 | 3.11 | 9.95 | 3.73 | 0.6905 |
| 16α-hydroxytestosterone | 5.9 | 6.8 | ND | ND | NA |
| 16α-hydroxyandrostenedione | 1.0 | 0.7 | ND | ND | NA |
| 17α-hydroxypregnenolone | 1.4 | 0.3 | ND | ND | NA |
| 17α-hydroxyprogesterone | 9.6 | 2.9 | 7.1 | 2.9 | 0.3095 |
| Tetrahydrodeoxycorticosterone | ND | ND | 3.9 | 2.1 | NA |
| Allo-tetrahydrodeoxycorticosterone | 0.9 | 0.4 | 1.0 | 0.3 | 0.5476 |
| 11-deoxycortisol | 2.9 | 3.0 | 3.8 | 2.5 | 0.1508 |
| Corticosterone | 13.3 | 12.4 | 7.7 | 5.7 | 0.3095 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shima, Y.; Miyabayashi, K.; Mori, T.; Ono, K.; Kajimoto, M.; Cho, H.L.; Tsuchida, H.; Uenoyama, Y.; Tsukamura, H.; Suzuki, K.; et al. Intronic Enhancer Is Essential for Nr5a1 Expression in the Pituitary Gonadotrope and for Postnatal Development of Male Reproductive Organs in a Mouse Model. Int. J. Mol. Sci. 2023, 24, 192. https://doi.org/10.3390/ijms24010192
Shima Y, Miyabayashi K, Mori T, Ono K, Kajimoto M, Cho HL, Tsuchida H, Uenoyama Y, Tsukamura H, Suzuki K, et al. Intronic Enhancer Is Essential for Nr5a1 Expression in the Pituitary Gonadotrope and for Postnatal Development of Male Reproductive Organs in a Mouse Model. International Journal of Molecular Sciences. 2023; 24(1):192. https://doi.org/10.3390/ijms24010192
Chicago/Turabian StyleShima, Yuichi, Kanako Miyabayashi, Takami Mori, Koji Ono, Mizuki Kajimoto, Hae Lim Cho, Hitomi Tsuchida, Yoshihisa Uenoyama, Hiroko Tsukamura, Kentaro Suzuki, and et al. 2023. "Intronic Enhancer Is Essential for Nr5a1 Expression in the Pituitary Gonadotrope and for Postnatal Development of Male Reproductive Organs in a Mouse Model" International Journal of Molecular Sciences 24, no. 1: 192. https://doi.org/10.3390/ijms24010192
APA StyleShima, Y., Miyabayashi, K., Mori, T., Ono, K., Kajimoto, M., Cho, H. L., Tsuchida, H., Uenoyama, Y., Tsukamura, H., Suzuki, K., Choi, M. H., & Toida, K. (2023). Intronic Enhancer Is Essential for Nr5a1 Expression in the Pituitary Gonadotrope and for Postnatal Development of Male Reproductive Organs in a Mouse Model. International Journal of Molecular Sciences, 24(1), 192. https://doi.org/10.3390/ijms24010192