
Chronic Intermittent Hypoxia Exposure Alternative to Exercise Alleviates High-Fat-Diet-Induced Obesity and Fatty Liver


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
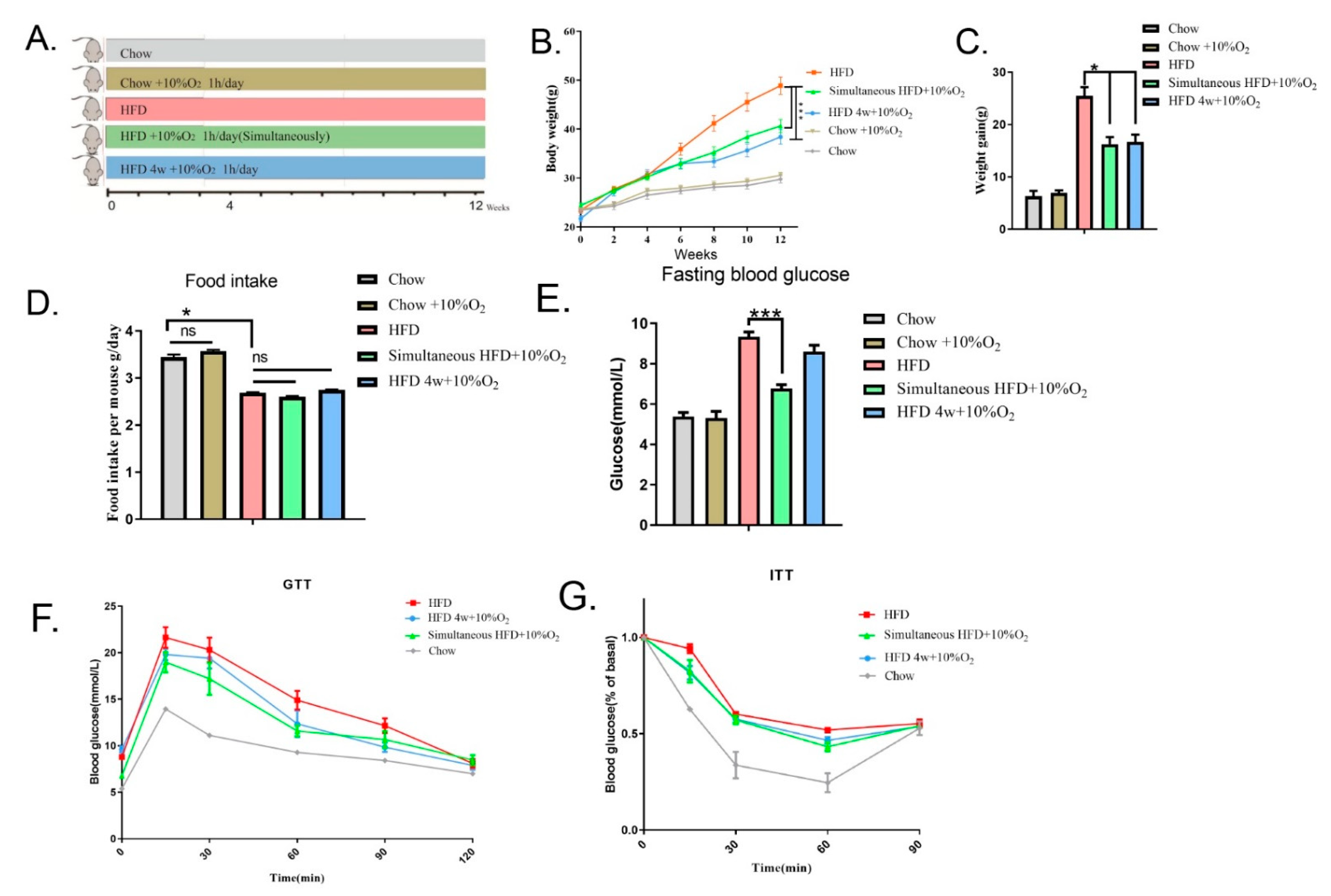
2.1. Chronic Intermittent Hypoxia Reduced Overweight/Obesity in Mice and Improved Glucose Tolerance
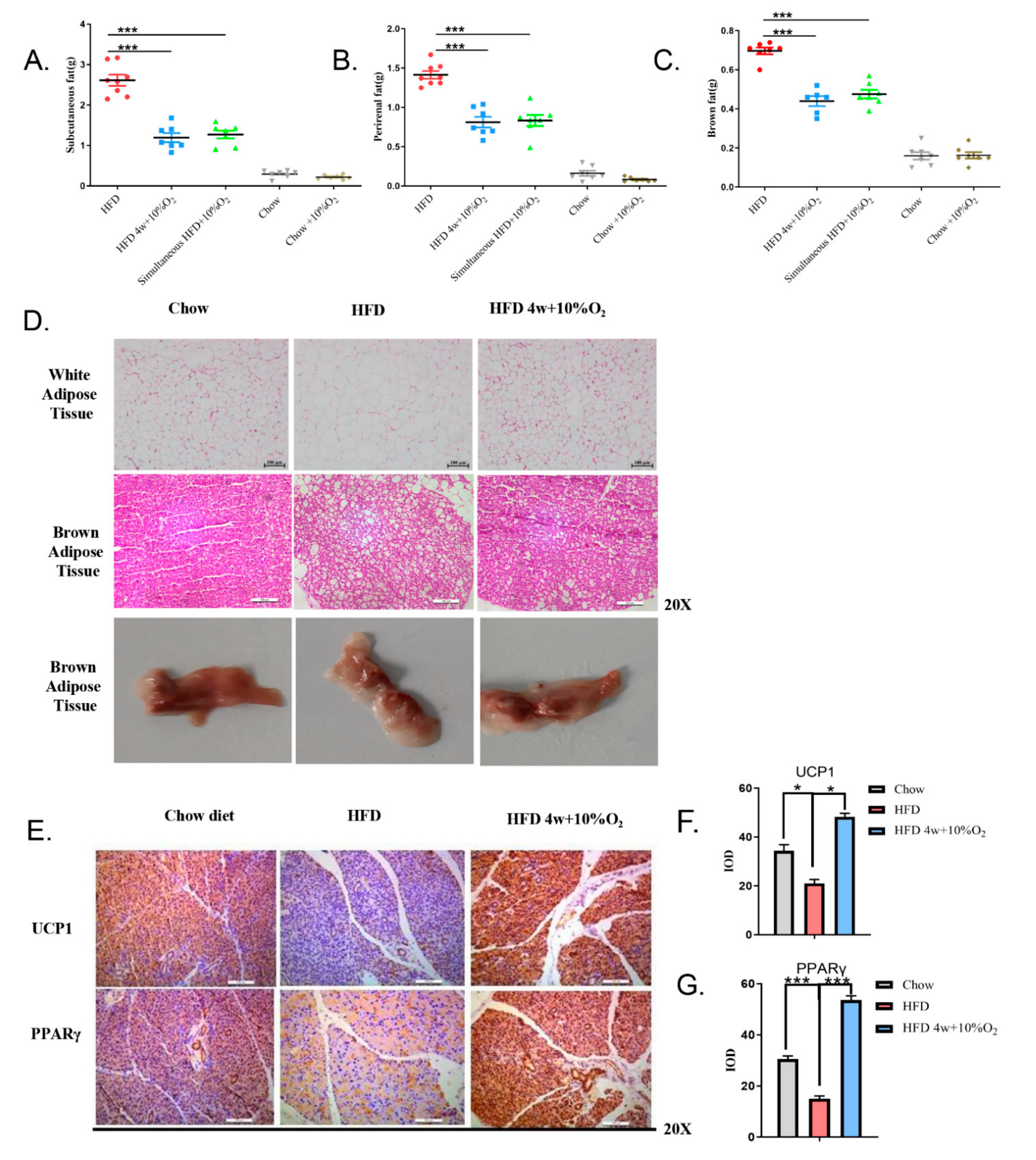
2.2. Chronic Intermittent Hypoxia Reduced the Fat Content of Mice and Promoted the Expression of Thermogenic Genes in Brown Fat
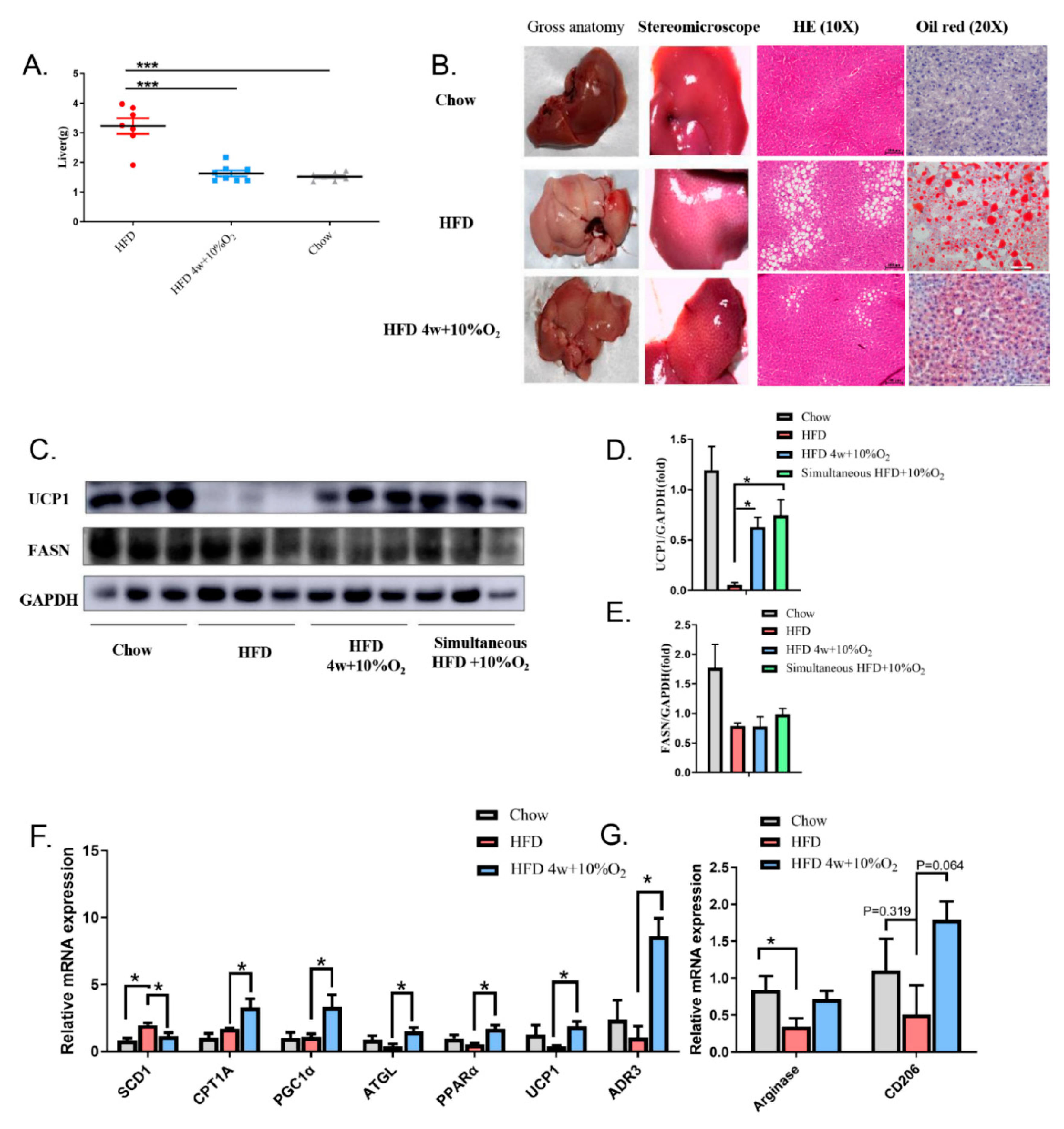
2.3. Chronic Intermittent Hypoxia Alleviated Fatty Liver and Altered Gene Expression
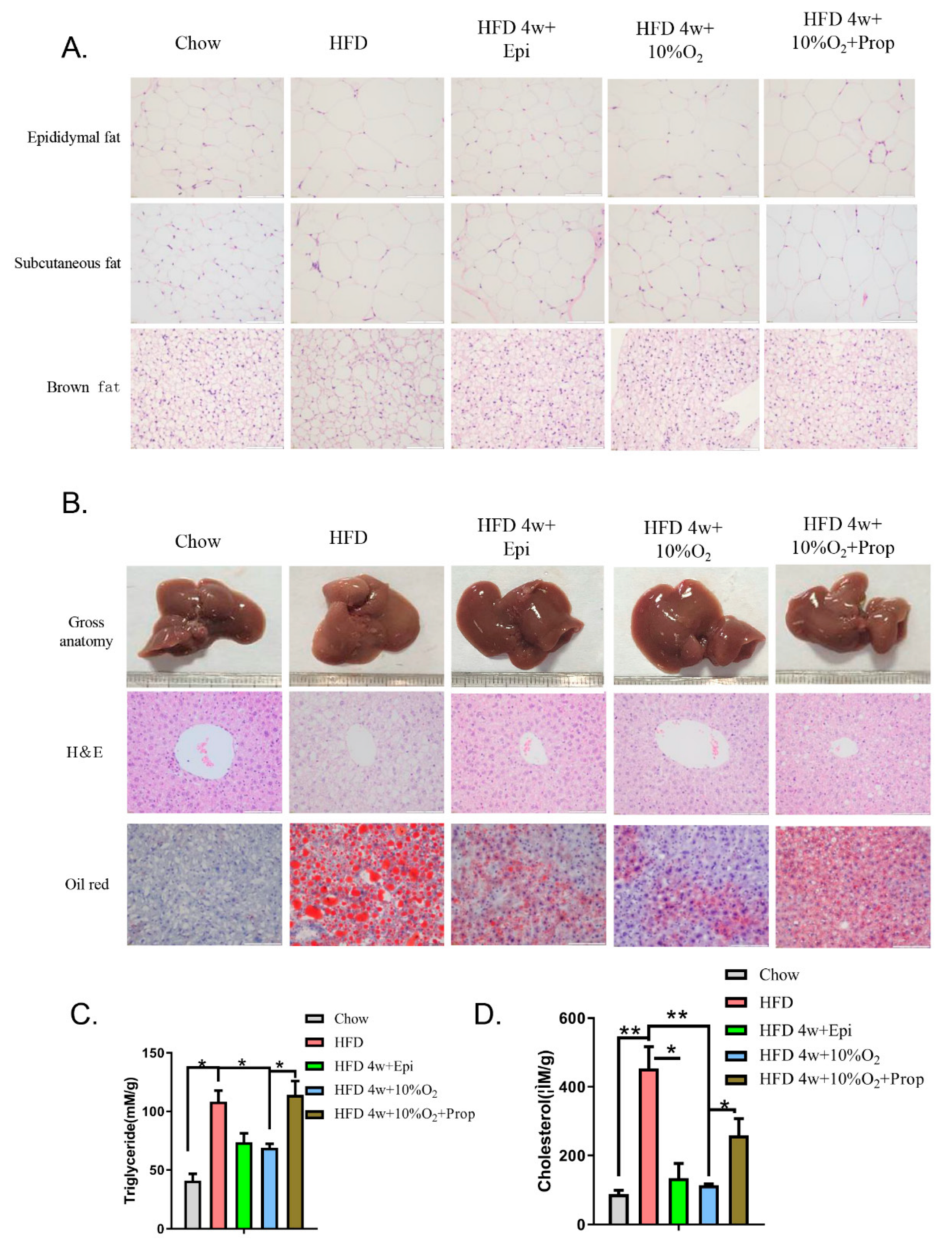
2.4. The Role of Epinephrine in Hypoxia-Induced Physiological Changes
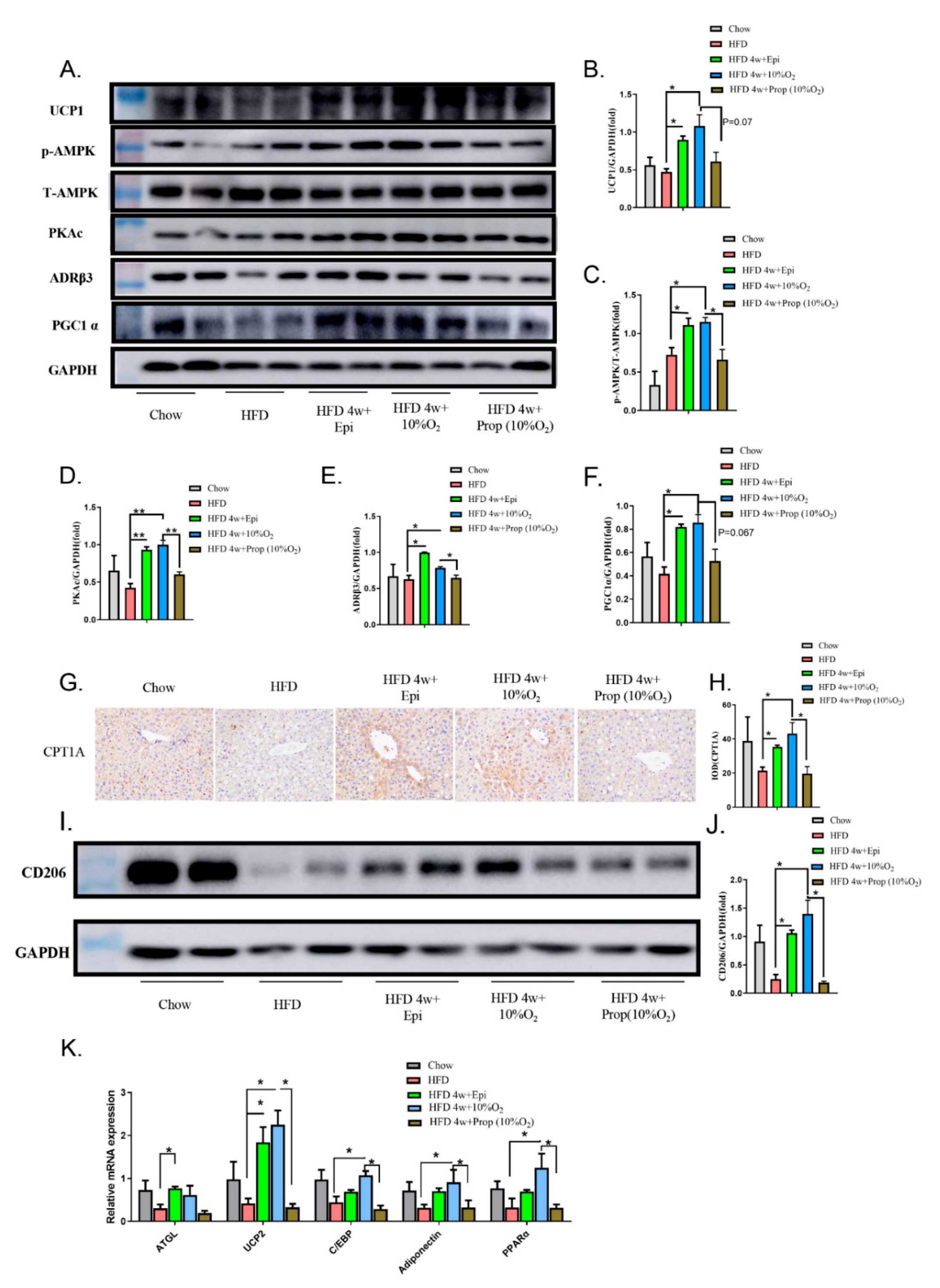
2.5. Epinephrine Mediated the Effect of Hypoxia on Fat Deposits in Liver and Adipose Tissues and Associated Molecular Changes
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Animal Model
4.3. Glucose Tolerance Test (GTT) and Insulin Tolerance Test (ITT)
4.4. Serum Epinephrine and Enzyme Assays
4.5. Tissue Homogenization
4.6. Measurement of Triacylglycerol and Total Cholesterol in Liver
4.7. Western Blot
4.8. Histochemistry
4.9. Quantitative Reverse Transcription–Polymerase Chain Reaction (RT–PCR)
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hill, J.J. Obesity: An Emerging Threat. J. Natl. Black Nurses’ Assoc. 2018, 29, 36–39. [Google Scholar]
- Frühbeck, G.; Toplak, H.; Woodward, E.; Yumuk, V.; Maislos, M.; Oppert, J.M. Obesity: The gateway to ill health—An EASO position statement on a rising public health, clinical and scientific challenge in Europe. Obes. Facts 2013, 6, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Polyzos, S.A.; Kountouras, J.; Mantzoros, C.S. Obesity and nonalcoholic fatty liver disease: From pathophysiology to therapeutics. Metabolism 2019, 92, 82–97. [Google Scholar] [CrossRef] [PubMed]
- Avenell, A.; Grant, A.M.; McGee, M.; McPherson, G.; Campbell, M.K.; McGee, M.A. The effects of an open design on trial participant recruitment, compliance and retention--a randomized controlled trial comparison with a blinded, placebo-controlled design. Clin. Trials 2004, 1, 490–498. [Google Scholar] [CrossRef]
- Dansinger, M.L.; Gleason, J.A.; Griffith, J.L.; Selker, H.P.; Schaefer, E.J. Comparison of the Atkins, Ornish, Weight Watchers, and Zone diets for weight loss and heart disease risk reduction: A randomized trial. JAMA 2005, 293, 43–53. [Google Scholar] [CrossRef]
- Woolcott, O.O.; Ader, M.; Bergman, R.N. Glucose homeostasis during short-term and prolonged exposure to high altitudes. Endocr. Rev. 2015, 36, 149–173. [Google Scholar] [CrossRef] [Green Version]
- Santos, C.; Bustamante, A.; Hedeker, D.; Vasconcelos, O.; Garganta, R.; Katzmarzyk, P.T.; Maia, J. Correlates of Overweight in Children and Adolescents Living at Different Altitudes: The Peruvian Health and Optimist Growth Study. J. Obes. 2019, 2019, 2631713. [Google Scholar] [CrossRef] [Green Version]
- Hao, G.S.; Fan, Q.L.; Hu, Q.Z.; Hou, Q. Research progress on the mechanism of cerebral blood flow regulation in hypoxia environment at plateau. Bioengineered 2022, 13, 6353–6358. [Google Scholar] [CrossRef]
- Wang, Y.; Shi, Y.; Li, W.; Wang, S.; Zheng, J.; Xu, G.; Li, G.; Shen, X.; Yang, J.A.-O. Gut microbiota imbalance mediates intestinal barrier damage in high-altitude exposed mice. FEBS J. 2022; early view. [Google Scholar] [CrossRef]
- Xu, Z.; You, W.; Zhou, Y.; Chen, W.; Wang, Y.; Shan, T. Cold-induced lipid dynamics and transcriptional programs in white adipose tissue. BMC Biol. 2019, 17, 74. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.Y.; Li, L.Y.; Jiao, X.L.; Jia, L.X.; Zhang, X.P.; Wang, Y.L.; Yang, S.; Li, J.; Du, J.; Wei, Y.X.; et al. Intermittent Hypoxia Alleviates beta-Aminopropionitrile Monofumarate Induced Thoracic Aortic Dissection in C57BL/6 Mice. Eur. J. Vasc. Endovasc. Surg. 2020, 59, 1000–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peacock, A.J. ABC of oxygen: Oxygen at high altitude. BMJ 1998, 317, 1063–1066. [Google Scholar] [CrossRef] [PubMed]
- Tomic, D.; Kemp, W.W.; Roberts, S.K. Nonalcoholic fatty liver disease: Current concepts, epidemiology and management strategies. Eur. J. Gastroenterol. Hepatol. 2018, 30, 1103–1115. [Google Scholar] [CrossRef] [PubMed]
- Lippl, F.J.; Neubauer, S.; Schipfer, S.; Lichter, N.; Tufman, A.; Otto, B.; Fischer, R. Hypobaric hypoxia causes body weight reduction in obese subjects. Obesity 2010, 18, 675–681. [Google Scholar] [CrossRef] [PubMed]
- Serebrovskaya, T.V. Intermittent hypoxia research in the former soviet union and the commonwealth of independent States: History and review of the concept and selected applications. High Alt. Med. Biol. 2002, 3, 205–221. [Google Scholar] [CrossRef] [Green Version]
- AIa, C.; Bludov, A.A. Efficiency of intermittent and resonance intermittent normobaric hypoxia therapy in patients with infection-dependent bronchial asthma. Vestn. Ross. Akad. Meditsinskikh Nauk. 2000, 9, 48–50. [Google Scholar]
- Glazachev, O.S.; Kryzhanovskaya, S.Y.; Zapara, M.A.; Dudnik, E.N.; Samartseva, V.G.; Susta, D. Safety and Efficacy of Intermittent Hypoxia Conditioning as a New Rehabilitation/Secondary Prevention Strategy for Patients with Cardiovascular Diseases: A Systematic Review and Meta-analysis. Curr. Cardiol. Rev. 2021, 17, 88–99. [Google Scholar] [CrossRef]
- Serebrovska, T.V.; Grib, O.N.; Portnichenko, V.I.; Serebrovska, Z.O.; Egorov, E.; Shatylo, V.B. Intermittent Hypoxia/Hyperoxia Versus Intermittent Hypoxia/Normoxia: Comparative Study in Prediabetes. High Alt. Med. Biol. 2019, 20, 383–391. [Google Scholar] [CrossRef]
- Serebrovskaya, T.; Karaban, I.; Mankovskaya, I.; Bernardi, L.; Passino, C.; Appenzeller, O. Hypoxic ventilatory responses and gas exchange in patients with Parkinson’s disease. Respiration 1998, 65, 28–33. [Google Scholar] [CrossRef]
- Serebrovskaya, T.V.; Manukhina, E.B.; Smith, M.L.; Downey, H.F.; Mallet, R.T. Intermittent hypoxia: Cause of or therapy for systemic hypertension? Exp. Biol. Med. 2008, 233, 627–650. [Google Scholar] [CrossRef]
- Kelly, K.R.; Williamson, D.L.; Fealy, C.E.; Kriz, D.A.; Krishnan, R.K.; Huang, H.; Ahn, J.; Loomis, J.L.; Kirwan, J.P. Acute altitude-induced hypoxia suppresses plasma glucose and leptin in healthy humans. Metabolism 2010, 59, 200–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, Q.; Sailan, W.; Ran, J.; Zhi, S.; Cen, L.; Yang, X.; Xiaoqun, Q. The effect of intermittent hypoxia on bodyweight, serum glucose and cholesterol in obesity mice. Pak. J. Biol. Sci. 2008, 11, 869–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, M.; Punjabi, N.M. Effects of acute intermittent hypoxia on glucose metabolism in awake healthy volunteers. J. Appl. Physiol. 2009, 106, 1538–1544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackenzie, R.; Maxwell, N.; Castle, P.; Brickley, G.; Watt, P. Acute hypoxia and exercise improve insulin sensitivity (S(I) (2*)) in individuals with type 2 diabetes. Diabetes/Metab. Res. Rev. 2011, 27, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Chiu, L.L.; Chou, S.W.; Cho, Y.M.; Ho, H.Y.; Ivy, J.L.; Hunt, D.; Wang, P.S.; Kuo, C.H. Effect of prolonged intermittent hypoxia and exercise training on glucose tolerance and muscle GLUT4 protein expression in rats. J. Biomed. Sci 2004, 11, 838–846. [Google Scholar] [CrossRef]
- Mitsui, T.; Nakamura, T.; Ito, T.; Umemoto, Y.; Sakamoto, K.; Kinoshita, T.; Nakagawa, M.; Tajima, F. Exercise significantly increases plasma adrenaline and oxidized low-density lipoprotein in normal healthy subjects but not in persons with spinal cord injury. Arch. Phys. Med. Rehabil. 2012, 93, 725–727. [Google Scholar] [CrossRef]
- Blegen, M.; Cheatham, C.; Caine-Bish, N.; Woolverton, C.; Marcinkiewicz, J.; Glickman, E. The immunological and metabolic responses to exercise of varying intensities in normoxic and hypoxic environments. J. Strength Cond. Res. 2008, 22, 1638–1644. [Google Scholar] [CrossRef]
- Kjaer, M.; Bangsbo, J.; Lortie, G.; Galbo, H. Hormonal response to exercise in humans: Influence of hypoxia and physical training. Am. J. Physiol. 1988, 254, R197–R203. [Google Scholar] [CrossRef]
- Lee, S.B.; Kim, H.G.; Lee, J.S.; Kim, W.Y.; Lee, M.M.; Kim, Y.H.; Lee, J.O.; Kim, H.S.; Son, C.G. Intermittent restraint-induced sympathetic activation attenuates hepatic steatosis and inflammation in a high-fat diet-fed mouse model. Am. J. Physiol.-Gastrointest. Liver Physiol. 2019, 317, G811–G823. [Google Scholar] [CrossRef]
- Larsson, S.; Jones, H.A.; Göransson, O.; Degerman, E.; Holm, C. Parathyroid hormone induces adipocyte lipolysis via PKA-mediated phosphorylation of hormone-sensitive lipase. Cell. Signal. 2016, 28, 204–213. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.Y.; Yu, L.; Chen, J.H.; Yang, L.N.; Lin, C.; Shi, X.Q.; Qin, H. Sesamol Alleviates Obesity-Related Hepatic Steatosis via Activating Hepatic PKA Pathway. Nutrients 2020, 12, 329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herzig, S.; Shaw, R.J. AMPK: Guardian of metabolism and mitochondrial homeostasis. Nat. Rev. Mol. Cell Biol. 2018, 19, 121–135. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Xiao, L.; Hou, Y.; He, Q.; Zhu, J.; Li, Y.; Wu, J.; Zhao, J.; Yu, S.; Zhao, Y. Sestrin2 Silencing Exacerbates Cerebral Ischemia/Reperfusion Injury by Decreasing Mitochondrial Biogenesis through the AMPK/PGC-1α Pathway in Rats. Sci. Rep. 2016, 6, 30272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujisaka, S.; Usui, I.; Nawaz, A.; Takikawa, A.; Kado, T.; Igarashi, Y.; Tobe, K. M2 macrophages in metabolism. Diabetol. Int. 2016, 7, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Garcia, D.; Hellberg, K.; Chaix, A.; Wallace, M.; Herzig, S.; Badur, M.G.; Lin, T.; Shokhirev, M.N.; Pinto, A.F.M.; Ross, D.S.; et al. Genetic Liver-Specific AMPK Activation Protects against Diet-Induced Obesity and NAFLD. Cell Rep. 2019, 26, 192–208.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oo, S.M.; Oo, H.K.; Takayama, H.; Ishii, K.A.; Takeshita, Y.; Goto, H.; Nakano, Y.; Kohno, S.; Takahashi, C.; Nakamura, H.; et al. Selenoprotein P-mediated reductive stress impairs cold-induced thermogenesis in brown fat. Cell Rep. 2022, 38, 110566. [Google Scholar] [CrossRef] [PubMed]
- Ishigaki, Y.; Katagiri, H.; Yamada, T.; Ogihara, T.; Imai, J.; Uno, K.; Hasegawa, Y.; Gao, J.; Ishihara, H.; Shimosegawa, T.; et al. Dissipating excess energy stored in the liver is a potential treatment strategy for diabetes associated with obesity. Diabetes 2005, 54, 322–332. [Google Scholar] [CrossRef] [Green Version]
- Kataoka, N.; Takeuchi, T.; Kusudo, T.; Li, Y.; Endo, Y.; Yamashita, H. Lack of UCP1 stimulates fatty liver but mediates UCP1-independent action of beige fat to improve hyperlipidemia in Apoe knockout mice. Biochim. Biophys. Acta BBA—Mol. Basis Dis. 2020, 1866, 165762. [Google Scholar] [CrossRef]
- Yang, Q.; Liang, X.; Sun, X.; Zhang, L.; Fu, X.; Rogers, C.J.; Berim, A.; Zhang, S.; Wang, S.; Wang, B.; et al. AMPK/α-Ketoglutarate Axis Dynamically Mediates DNA Demethylation in the Prdm16 Promoter and Brown Adipogenesis. Cell Metab. 2016, 24, 542–554. [Google Scholar] [CrossRef] [Green Version]
- Pollard, A.E.; Martins, L.; Muckett, P.J.; Khadayate, S.; Bornot, A.; Clausen, M.; Admyre, T.; Bjursell, M.; Fiadeiro, R.; Wilson, L.; et al. AMPK activation protects against diet induced obesity through Ucp1-independent thermogenesis in subcutaneous white adipose tissue. Nat. Metab. 2019, 1, 340–349. [Google Scholar] [CrossRef]
- Flowers, M.T.; Ntambi, J.M. Role of stearoyl-coenzyme A desaturase in regulating lipid metabolism. Curr. Opin. Lipidol. 2008, 19, 248–256. [Google Scholar] [CrossRef] [Green Version]
- Cryer, P.E. Adrenaline: A physiological metabolic regulatory hormone in humans? Int. J. Obes. Relat. Metab. Disord. J. Int. Assoc. Study Obes. 1993, 17 (Suppl. 3), S43–S46, discussion S68. [Google Scholar]
- Gore, C.J.; Clark, S.A.; Saunders, P.U. Nonhematological mechanisms of improved sea-level performance after hypoxic exposure. Med. Sci. Sports Exerc. 2007, 39, 1600. [Google Scholar] [CrossRef] [PubMed]
- Vogt, M.; Puntschart, A.; Geiser, J.; Zuleger, C.; Billeter, R.; Hoppeler, H. Molecular adaptations in human skeletal muscle to endurance training under simulated hypoxic conditions. J. Appl. Physiol. 2001, 91, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Hoppeler, H.; Vogt, M. Muscle tissue adaptations to hypoxia. J. Exp. Biol. 2001, 204, 3133–3139. [Google Scholar] [CrossRef] [PubMed]
- Mesarwi, O.A.; Moya, E.A.; Zhen, X.; Gautane, M.; Zhao, H.; Wegbrans Giró, P.; Alshebli, M.; McCarley, K.E.; Breen, E.C.; Malhotra, A. Hepatocyte HIF-1 and Intermittent Hypoxia Independently Impact Liver Fibrosis in Murine Nonalcoholic Fatty Liver Disease. Am. J. Respir. Cell Mol. Biol. 2021, 65, 390–402. [Google Scholar] [CrossRef]
- Lee, M.; Hwang, J.T.; Yun, H.; Kim, E.J.; Kim, M.J.; Kim, S.S.; Ha, J. Critical roles of AMP-activated protein kinase in the carcinogenic metal-induced expression of VEGF and HIF-1 proteins in DU145 prostate carcinoma. Biochem. Pharmacol. 2006, 72, 91–103. [Google Scholar] [CrossRef]
- Treins, C.; Murdaca, J.; Van Obberghen, E.; Giorgetti-Peraldi, S. AMPK activation inhibits the expression of HIF-1alpha induced by insulin and IGF-1. Biochemical Biophys. Res. Commun. 2006, 342, 1197–1202. [Google Scholar] [CrossRef]
- Siques, P.; Brito, J.; Flores, K.; Ordenes, S.; Arriaza, K.; Pena, E.; León-Velarde, F.; López de Pablo, Á.L.; Gonzalez, M.C.; Arribas, S. Long-Term Chronic Intermittent Hypobaric Hypoxia Induces Glucose Transporter (GLUT4) Translocation Through AMP-Activated Protein Kinase (AMPK) in the Soleus Muscle in Lean Rats. Front. Physiol. 2018, 9, 799. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Tsai, Y.-L.; Kao, C.-L.; Lee, S.-S.; Wu, M.-C.; Mallikarjuna, K.; Liao, Y.-H.; Ivy, J.-L.; Kuo, C.-H. Effect of mild intermittent hypoxia on glucose tolerance, muscle morphology and AMPK-PGC-1alpha signaling. Chin. J. Physiol. 2010, 53, 62–71. [Google Scholar] [CrossRef]
- Regué, L.; Minichiello, L.; Avruch, J.; Dai, N. Liver-specific deletion of IGF2 mRNA binding protein-2/IMP2 reduces hepatic fatty acid oxidation and increases hepatic triglyceride accumulation. J. Biol. Chem. 2019, 294, 11944–11951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, J.; Li, C.; Jiang, S.; Luo, L.; Yan, X.; Huang, D.; Luo, Z. AMPK inhibits Smad3-mediated autoinduction of TGF-β1 in gastric cancer cells. J. Cell. Mol. Med. 2021, 25, 2806–2815. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, Y.; Chen, Q.; Zou, J.; Fan, J.; Li, Y.; Luo, Z. Chronic Intermittent Hypoxia Exposure Alternative to Exercise Alleviates High-Fat-Diet-Induced Obesity and Fatty Liver. Int. J. Mol. Sci. 2022, 23, 5209. https://doi.org/10.3390/ijms23095209
Luo Y, Chen Q, Zou J, Fan J, Li Y, Luo Z. Chronic Intermittent Hypoxia Exposure Alternative to Exercise Alleviates High-Fat-Diet-Induced Obesity and Fatty Liver. International Journal of Molecular Sciences. 2022; 23(9):5209. https://doi.org/10.3390/ijms23095209
Chicago/Turabian StyleLuo, Yunfei, Qiongfeng Chen, Junrong Zou, Jingjing Fan, Yuanjun Li, and Zhijun Luo. 2022. "Chronic Intermittent Hypoxia Exposure Alternative to Exercise Alleviates High-Fat-Diet-Induced Obesity and Fatty Liver" International Journal of Molecular Sciences 23, no. 9: 5209. https://doi.org/10.3390/ijms23095209
APA StyleLuo, Y., Chen, Q., Zou, J., Fan, J., Li, Y., & Luo, Z. (2022). Chronic Intermittent Hypoxia Exposure Alternative to Exercise Alleviates High-Fat-Diet-Induced Obesity and Fatty Liver. International Journal of Molecular Sciences, 23(9), 5209. https://doi.org/10.3390/ijms23095209