
Actin Stress Fibers Response and Adaptation under Stretch


Abstract
1. Introduction
Stress Fibers’ Mechanical Model
2. Results
2.1. Static Analysis of Stretched Micro Patterned Cells
2.1.1. Short Time Scale Analysis
2.1.2. Long Time Scale Analysis
2.2. Fresh Actin Recruitment during Compression and Extension of Actin Bundles
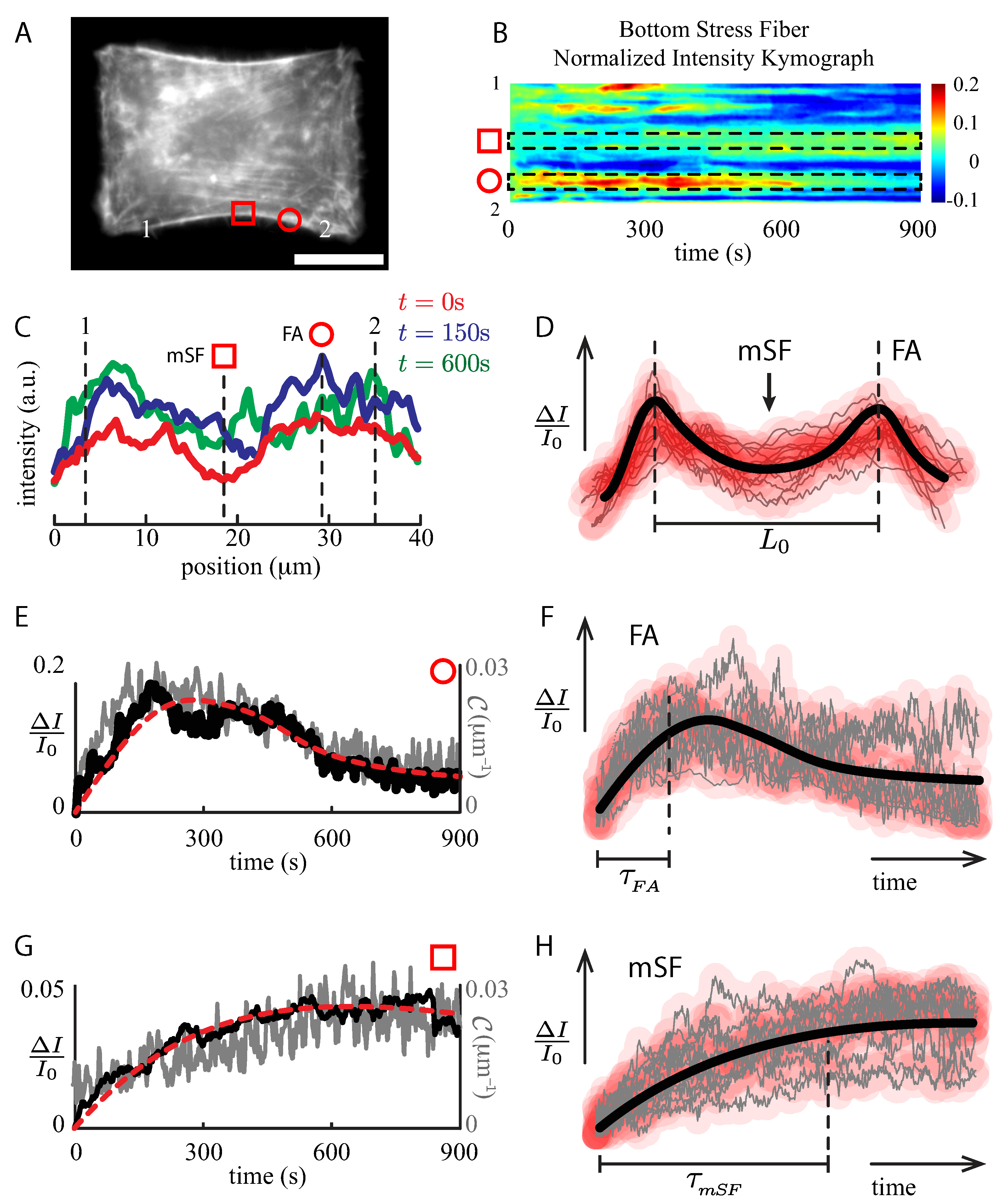
2.3. Global to Local Dynamics of Stretched Actin Bundles
2.4. Local Temporal Evolution Analysis of the SF under Stretch
3. Discussion
4. Materials and Methods
4.1. PDMS Micro Patterning
4.2. Cell Culture and Experiments
4.3. Image Acquisition and Analysis
4.4. Dynamics under Stretch
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Blanchoin, L.; Boujemaa-Paterski, R.; Sykes, C.; Plastino, J. Actin dynamics, architecture, and mechanics in cell motility. Physiol. Rev. 2014, 94, 235–263. [Google Scholar] [CrossRef] [PubMed]
- Mitrossilis, D.; Fouchard, J.; Pereira, D.; Postic, F.; Richert, A.; Saint-Jean, M.; Asnacios, A. Real-time single-cell response to stiffness. Proc. Natl. Acad. Sci. USA 2010, 107, 16518–16523. [Google Scholar] [CrossRef] [PubMed]
- Joanny, J.F.; Prost, J. Active gels as a description of the actin-myosin cytoskeleton. HFSP J. 2009, 3, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Faust, U.; Hampe, N.; Rubner, W.; Kirchgeßner, N.; Safran, S.; Hoffmann, B.; Merkel, R. Cyclic stress at mHz frequencies aligns fibroblasts in direction of zero strain. PLoS ONE 2011, 6, e028963. [Google Scholar] [CrossRef]
- Livne, A.; Bouchbinder, E.; Geiger, B. Cell reorientation under cyclic stretching. Nat. Commun. 2014, 5, 3938. [Google Scholar] [CrossRef]
- Pellegrin, S.; Mellor, H. Actin stress fibers. J. Cell Sci. 2007, 120, 3491–3499. [Google Scholar] [CrossRef]
- Kumar, S.; Maxwell, I.Z.; Heisterkamp, A.; Polte, T.R.; Lele, T.P.; Salanga, M.; Mazur, E.; Ingber, D.E. Viscoelastic retraction of single living stress fibers and its impact on cell shape, cytoskeletal organization, and extracellular matrix mechanics. Biophys. J. 2006, 90, 3762–3773. [Google Scholar] [CrossRef]
- Colombelli, J.; Besser, A.; Kress, H.; Reynaud, E.G.; Girard, P.; Caussinus, E.; Haselmann, U.; Small, J.V.; Schwarz, U.S.; Stelzer, E.H. Erratum: Mechanosensing in actin stress fibers revealed by a close correlation between force and protein localization (Journal of Cell Science vol. 122 (1665-1679)). J. Cell Sci. 2009, 122, 1928. [Google Scholar] [CrossRef]
- Peterson, L.J.; Rajfur, Z.; Maddox, A.S.; Freel, C.D.; Chen, Y.; Edlund, M.; Otey, C.; Burridge, K. Simultaneous stretching and contraction of stress fibers in vivo. Mol. Biol. Cell 2004, 15, 3497–3508. [Google Scholar] [CrossRef]
- Besser, A.; Colombelli, J.; Stelzer, E.H.; Schwarz, U.S. Viscoelastic response of contractile filament bundles. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2011, 83, 1–12. [Google Scholar] [CrossRef]
- Stachowiak, M.R.; Smith, M.A.; Blankman, E.; Chapin, L.M.; Balcioglu, H.E.; Wang, S.; Beckerle, M.C.; O’Shaughnessy, B. A mechanical-biochemical feedback loop regulates remodeling in the actin cytoskeleton. Proc. Natl. Acad. Sci. USA 2014, 111, 17528–17533. [Google Scholar] [CrossRef] [PubMed]
- Huxley, A.F. Muscle structure and theories of contraction. Prog. Biophys. Biophys. Chem. 1957, 7, 255–318. [Google Scholar] [CrossRef]
- Lan, G.; Sun, S.X. Dynamics of myosin-driven skeletal muscle contraction: I. Steady-state force generation. Biophys. J. 2005, 88, 4107–4117. [Google Scholar] [CrossRef]
- Chin, L.; Yue, P.; Feng, J.J.; Seow, C.Y. Mathematical simulation of muscle cross-bridge cycle and force-velocity relationship. Biophys. J. 2006, 91, 3653–3663. [Google Scholar] [CrossRef] [PubMed]
- Loh, O.; Vaziri, A.; Espinosa, H.D. The potential of MEMS for advancing experiments and modeling in cell mechanics. Exp. Mech. 2009, 49, 105–124. [Google Scholar] [CrossRef]
- Goldyn, A.M.; Rioja, B.A.; Spatz, J.P.; Ballestrem, C.; Kemkemer, R. Force-induced cell polarisation is linked to RhoA-driven microtubule-independent focal-adhesion sliding. J. Cell Sci. 2009, 122, 3644–3651. [Google Scholar] [CrossRef]
- Kaverina, I.; Krylyshkina, O.; Small, J.V. Microtubule targeting of substrate contacts promotes their relaxation and dissociation. J. Cell Biol. 1999, 146, 1033–1043. [Google Scholar] [CrossRef]
- Théry, M.; Racine, V.; Pépin, A.; Piel, M.; Chen, Y.; Sibarita, J.B.; Bornens, M. The extracellular matrix guides the orientation of the cell division axis. Nat. Cell Biol. 2005, 7, 947–953. [Google Scholar] [CrossRef]
- Théry, M.; Racine, V.; Piel, M.; Pépin, A.; Dimitrov, A.; Chen, Y.; Sibarita, J.B.; Bornens, M. Anisotropy of cell adhesive microenvironment governs cell internal organization and orientation of polarity. Proc. Natl. Acad. Sci. USA 2006, 103, 19771–19776. [Google Scholar] [CrossRef]
- Fink, J.; Carpi, N.; Betz, T.; Bétard, A.; Chebah, M.; Azioune, A.; Bornens, M.; Sykes, C.; Fetler, L.; Cuvelier, D.; et al. External forces control mitotic spindle positioning. Nat. Cell Biol. 2011, 13, 771–778. [Google Scholar] [CrossRef]
- Zhang, H.; Cai, C.; Liu, W.; Li, D.; Zhang, J.; Zhao, N.; Xu, J. Recyclable polydimethylsiloxane network crosslinked by dynamic transesterification reaction. Sci. Rep. 2017, 7, 11833. [Google Scholar] [CrossRef] [PubMed]
- Azioune, A.; Carpi, N.; Fink, J.; Chehimi, M.M.; Cuvelier, D.; Piel, M. Robust method for high-throughput surface patterning of deformable substrates. Langmuir 2011, 27, 7349–7352. [Google Scholar] [CrossRef] [PubMed]
- Bischofs, I.B.; Klein, F.; Lehnert, D.; Bastmeyer, M.; Schwarz, U.S. Filamentous network mechanics and active contractility determine cell and tissue shape. Biophys. J. 2008, 95, 3488–3496. [Google Scholar] [CrossRef] [PubMed]
- Bischofs, I.B.; Schmidt, S.S.; Schwarz, U.S. Effect of adhesion geometry and rigidity on cellular force distributions. Phys. Rev. Lett. 2009, 103, 048101. [Google Scholar] [CrossRef]
- Burridge, K.; Wittchen, E.S. Stress fibers get a makeover. Biophys. J. 2012, 103, 2045–2046. [Google Scholar] [CrossRef][Green Version]
- Trichet, L.; Le Digabel, J.; Hawkins, R.J.; Vedula, S.R.K.; Gupta, M.; Ribrault, C.; Hersen, P.; Voituriez, R.; Ladoux, B. Evidence of a large-scale mechanosensing mechanism for cellular adaptation to substrate stiffness. Proc. Natl. Acad. Sci. USA 2012, 109, 6933–6938. [Google Scholar] [CrossRef]
- Oiwa, K.; Chaen, S.; Kamitsubo, E.; Shimmen, T.; Sugi, H. Steady-state force-velocity relation in the ATP-dependent sliding movement of myosin-coated beads on actin cables in vitro studied with a centrifuge microscope. Proc. Natl. Acad. Sci. USA 1990, 87, 7893–7897. [Google Scholar] [CrossRef]
- Schnitzer, M.J.; Visscher, K.; Block, S.M. Force production by single kinesin motors. Nat. Cell Biol. 2000, 2, 718–723. [Google Scholar] [CrossRef]
- Howard, J. Mechanics of Motor Proteins and the Cytoskeleton. Phys. Today 2002, 55, 63. [Google Scholar] [CrossRef]
- Bernal, R.; Pullarkat, P.A.; Melo, F. Mechanical properties of axons. Phys. Rev. Lett. 2007, 99, 018301. [Google Scholar] [CrossRef]
- Bernal, R.; Melo, F.; Pullarkat, P.A. Drag force as a tool to test the active mechanical response of PC12 neurites. Biophys. J. 2010, 98, 515–523. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Smith, M.A.; Blankman, E.; Gardel, M.L.; Luettjohann, L.; Waterman, C.M.; Beckerle, M.C. A Zyxin-mediated mechanism for actin stress fiber maintenance and repair. Dev. Cell 2010, 19, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Yoshigi, M.; Hoffman, L.M.; Jensen, C.C.; Yost, H.J.; Beckerle, M.C. Mechanical force mobilizes zyxin from focal adhesions to actin filaments and regulates cytoskeletal reinforcement. J. Cell Biol. 2005, 171, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Costa, K.D.; Hucker, W.J.; Yin, F.C. Buckling of actin stress fibers: A new wrinkle in the cytoskeletal tapestry. Cell Motil. Cytoskelet. 2002, 52, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Kassianidou, E.; Kumar, S. A biomechanical perspective on stress fiber structure and function. Biochim. Biophys. Acta Mol. Cell Res. 2015, 1853, 3065–3074. [Google Scholar] [CrossRef] [PubMed]
- Åström, J.A.; Kumar, P.B.; Vattulainen, I.; Karttunen, M. Strain hardening, avalanches, and strain softening in dense cross-linked actin networks. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2008, 77, 1–6. [Google Scholar] [CrossRef]
- Lee, H.; Ferrer, J.M.; Lang, M.J.; Kamm, R.D. Molecular origin of strain softening in cross-linked F-actin networks. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2010, 82, 1–9. [Google Scholar] [CrossRef]
- Wolff, L.; Fernández, P.; Kroy, K. Resolving the stiffening-softening paradox in cell mechanics. PLoS ONE 2012, 7, e040063. [Google Scholar] [CrossRef]
- Ahmed, W.W.; Fodor, É.; Betz, T. Active cell mechanics: Measurement and theory. Biochim. Biophys. Acta Mol. Cell Res. 2015, 1853, 3083–3094. [Google Scholar] [CrossRef]
- Lu, L.; Oswald, S.J.; Ngu, H.; Yin, F.C. Mechanical properties of actin stress fibers in living cells. Biophys. J. 2008, 95, 6060–6071. [Google Scholar] [CrossRef]
- Cochet-Escartin, O.; Ranft, J.; Silberzan, P.; Marcq, P. Border forces and friction control epithelial closure dynamics. Biophys. J. 2014, 106, 65–73. [Google Scholar] [CrossRef] [PubMed]
- López-Gay, J.M.; Nunley, H.; Spencer, M.; Di Pietro, F.; Guirao, B.; Bosveld, F.; Markova, O.; Gaugue, I.; Pelletier, S.; Lubensky, D.K.; et al. Apical stress fibers enable a scaling between cell mechanical response and area in epithelial tissue. Science 2020, 370, eabb2169. [Google Scholar] [CrossRef] [PubMed]
- Peukes, J.; Betz, T. Direct measurement of the cortical tension during the growth of membrane blebs. Biophys. J. 2014, 107, 1810–1820. [Google Scholar] [CrossRef] [PubMed]
- Gárate, F.; Betz, T.; Pertusa, M.; Bernal, R. Time-resolved neurite mechanics by thermal fluctuation assessments. Phys. Biol. 2015, 12, 66020. [Google Scholar] [CrossRef] [PubMed]
- Gárate, F.; Pertusa, M.; Arana, Y.; Bernal, R. Non-invasive neurite mechanics in differentiated PC12 cells. Front. Cell. Neurosci. 2018, 12, 194. [Google Scholar] [CrossRef]
- Burridge, K.; Wittchen, E.S. The tension mounts: Stress fibers as force-generating mechanotransducers. J. Cell Biol. 2013, 200, 9–19. [Google Scholar] [CrossRef]
- Labouesse, C.; Gabella, C.; Meister, J.J.; Vianay, B.; Verkhovsky, A.B. Microsurgery-aided in situ force probing reveals extensibility and viscoelastic properties of individual stress fibers. Sci. Rep. 2016, 6, 23722. [Google Scholar] [CrossRef]
- Goldmann, W.H. Mechanotransduction and focal adhesions. Cell Biol. Int. 2012, 36, 649–652. [Google Scholar] [CrossRef]
- Kuo, J.C. Mechanotransduction at focal adhesions: Integrating cytoskeletal mechanics in migrating cells. J. Cell. Mol. Med. 2013, 17, 704–712. [Google Scholar] [CrossRef]
- Riveline, D.; Zamir, E.; Balaban, N.Q.; Schwarz, U.S.; Ishizaki, T.; Narumiya, S.; Kam, Z.; Geiger, B.; Bershadsky, A.D. Focal contacts as mechanosensors: Externally applied local mechanical force induces growth of focal contacts by an mDia1-dependent and ROCK-independent mechanism. J. Cell Biol. 2001, 153, 1175–1185. [Google Scholar] [CrossRef]
- Schaedel, L.; John, K.; Gaillard, J.; Nachury, M.V.; Blanchoin, L.; Thery, M. Microtubules self-repair in response to mechanical stress. Nat. Mater. 2015, 14, 1156–1163. [Google Scholar] [CrossRef] [PubMed]
- Rückerl, F.; Lenz, M.; Betz, T.; Manzi, J.; Martiel, J.L.; Safouane, M.; Paterski-Boujemaa, R.; Blanchoin, L.; Sykes, C. Adaptive Response of Actin Bundles under Mechanical Stress. Biophys. J. 2017, 113, 1072–1079. [Google Scholar] [CrossRef] [PubMed]
- Déjardin, T.; Carollo, P.S.; Sipieter, F.; Davidson, P.M.; Seiler, C.; Cuvelier, D.; Cadot, B.; Sykes, C.; Gomes, E.R.; Borghi, N. Nesprins are mechanotransducers that discriminate epithelial–mesenchymal transition programs. J. Cell Biol. 2020, 219, e201908036. [Google Scholar] [CrossRef] [PubMed]
- Miura, K. Bleach correction ImageJ plugin for compensating the photobleaching of time-lapse sequences. F1000Research 2020, 9, 1494. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Mechanical Parameter | Value | Model | Units |
|---|---|---|---|---|
| Initial cortical tension | 35 ( 15) | — | pN/m | |
| Initial line tension | 1.9 ( 1.4) | — | nN | |
| Passive time scale | 320 ( 90) | ∼300 | s | |
| Active time scale | 240 ( 50) | ∼200 | s | |
| Elasticity/cortex tension ratio | 2–10 | 1 | — | |
| Motor activity/motor dissipation ratio | 0.3–1.4 | 1 | — |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernal, R.; Van Hemelryck, M.; Gurchenkov, B.; Cuvelier, D. Actin Stress Fibers Response and Adaptation under Stretch. Int. J. Mol. Sci. 2022, 23, 5095. https://doi.org/10.3390/ijms23095095
Bernal R, Van Hemelryck M, Gurchenkov B, Cuvelier D. Actin Stress Fibers Response and Adaptation under Stretch. International Journal of Molecular Sciences. 2022; 23(9):5095. https://doi.org/10.3390/ijms23095095
Chicago/Turabian StyleBernal, Roberto, Milenka Van Hemelryck, Basile Gurchenkov, and Damien Cuvelier. 2022. "Actin Stress Fibers Response and Adaptation under Stretch" International Journal of Molecular Sciences 23, no. 9: 5095. https://doi.org/10.3390/ijms23095095
APA StyleBernal, R., Van Hemelryck, M., Gurchenkov, B., & Cuvelier, D. (2022). Actin Stress Fibers Response and Adaptation under Stretch. International Journal of Molecular Sciences, 23(9), 5095. https://doi.org/10.3390/ijms23095095