
High-Throughput In Vitro Screening Identified Nemadipine as a Novel Suppressor of Embryo Implantation

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
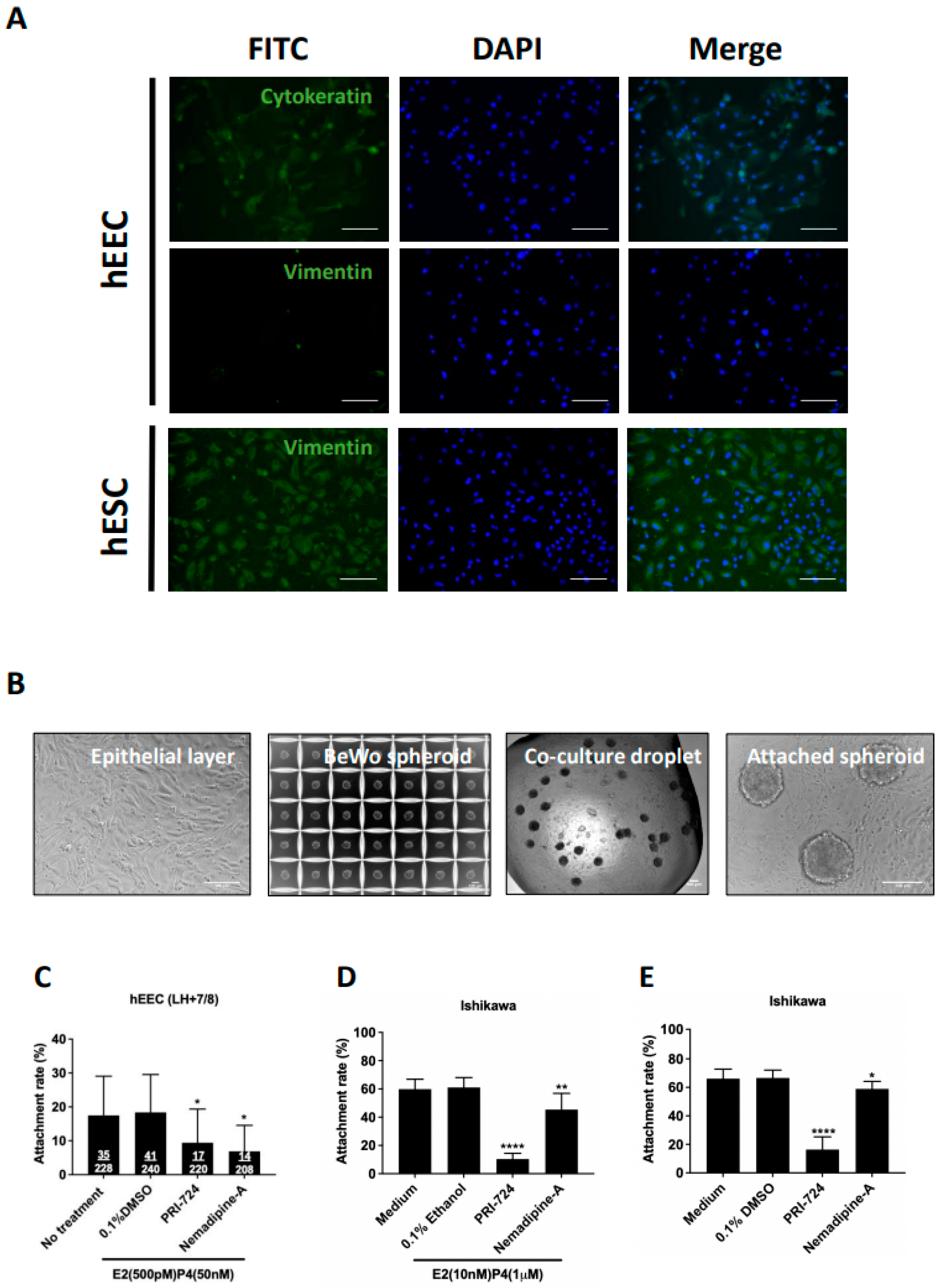
2.1. Establishment and Validation of a High-Throughput In Vitro Model
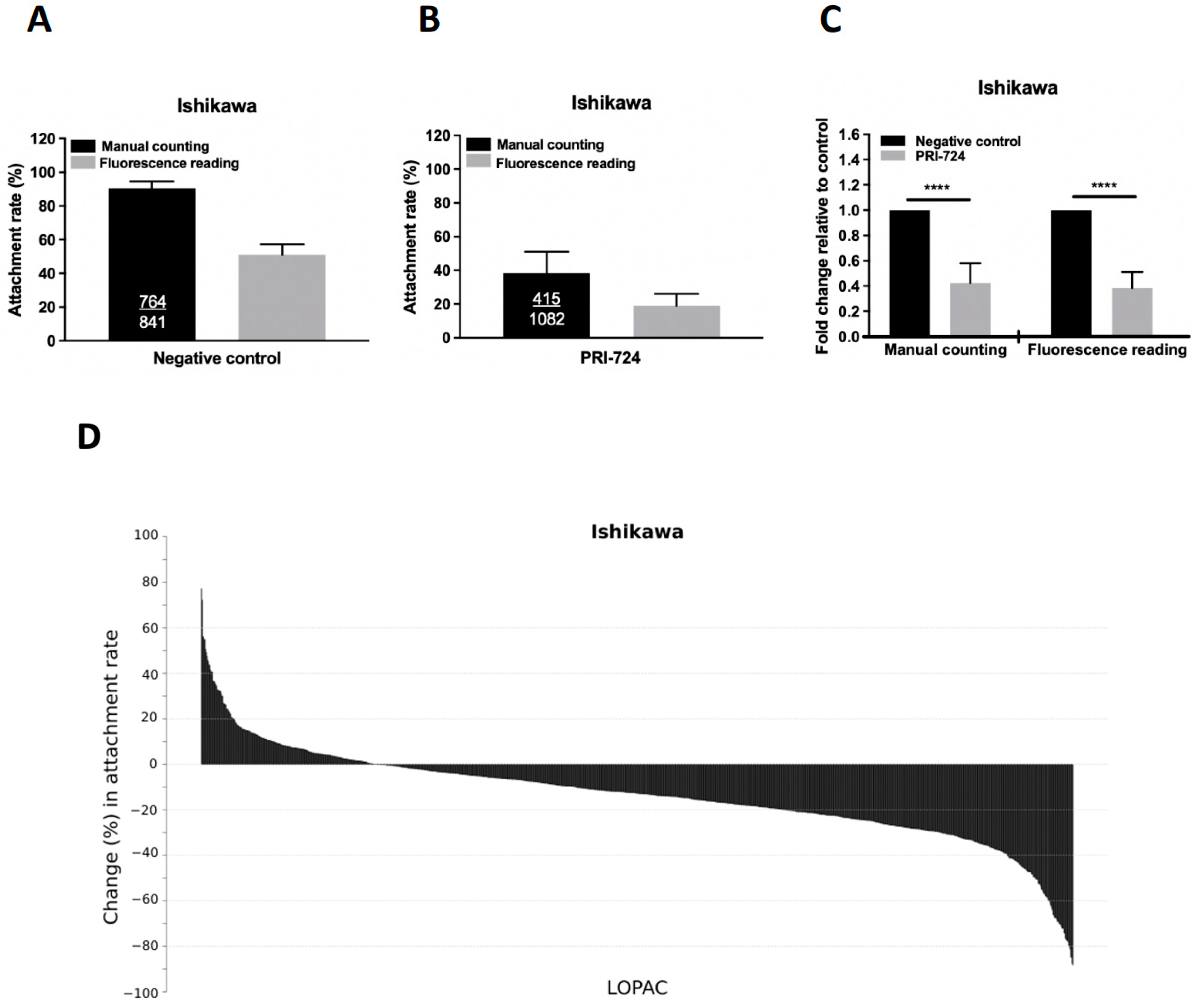
2.2. Primary Screening of LOPAC for Compounds That Suppress Spheroid Attachment
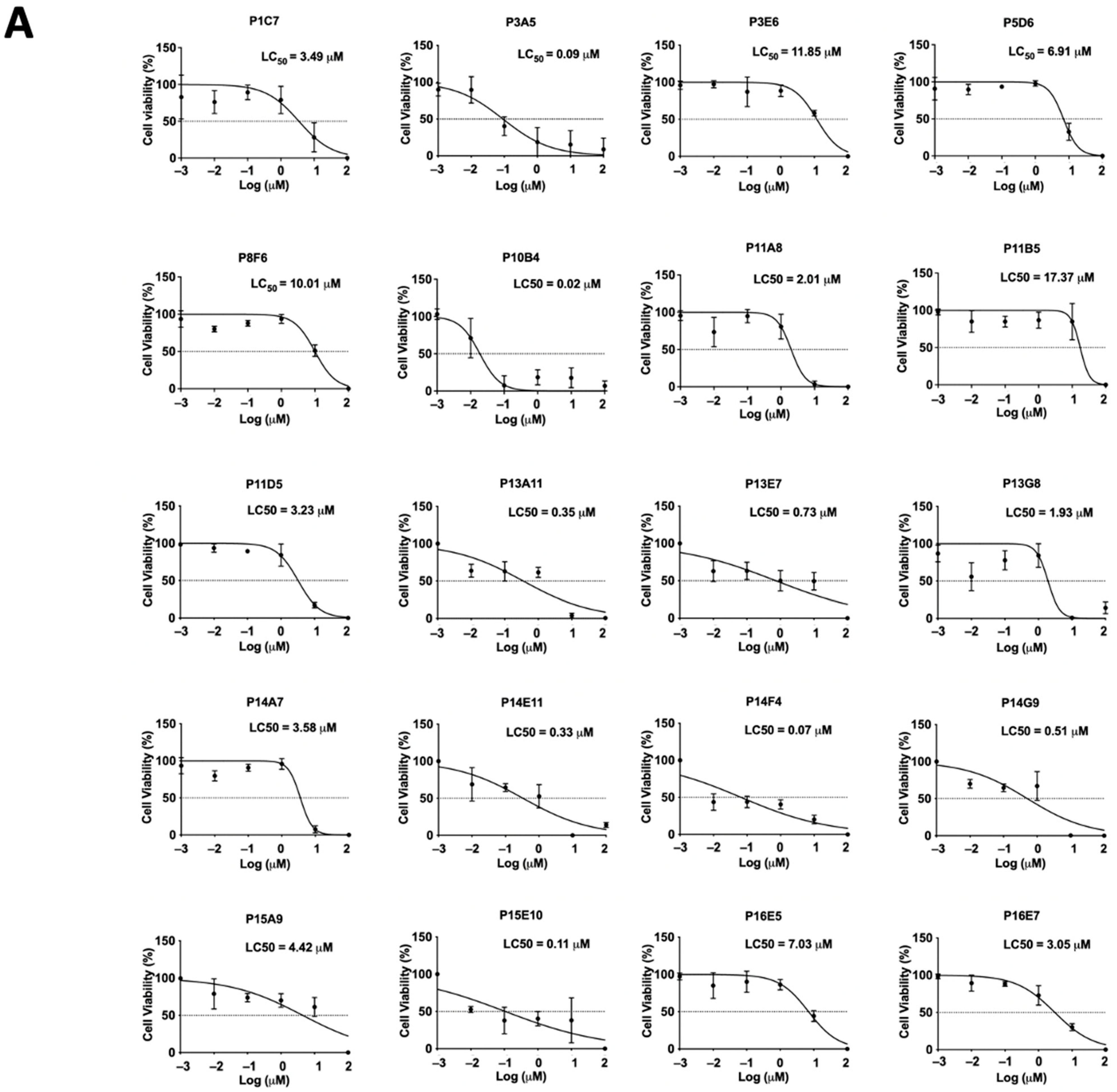
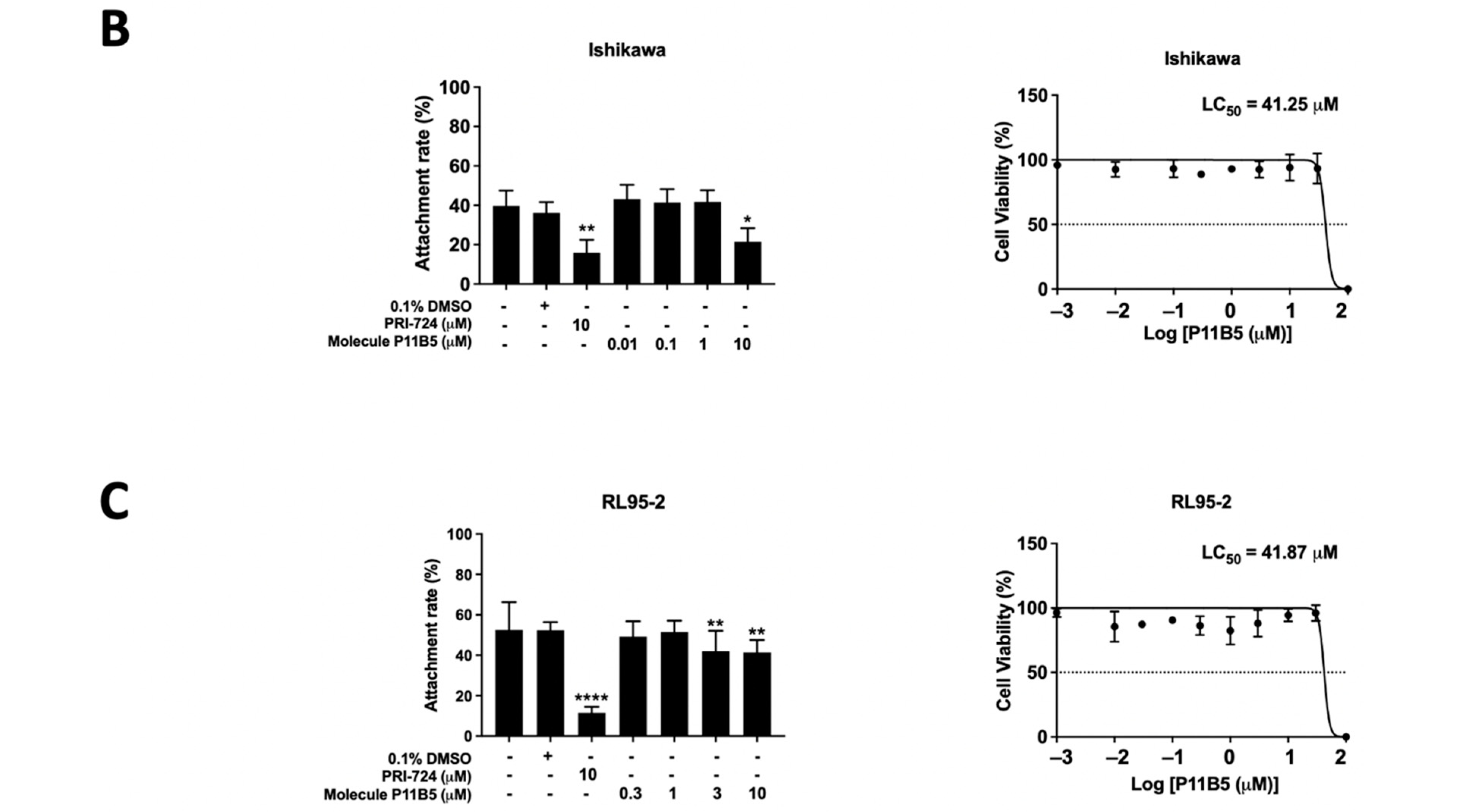
2.3. Secondary Screening of the Selected Compounds from LOPAC with Low Cytotoxicity
2.4. Nemadipine-A Suppressed Attachment in Receptive Primary Human Endometrial Epithelial Cells (hEECs) and Ishikawa Cells with or without Steroid Hormones
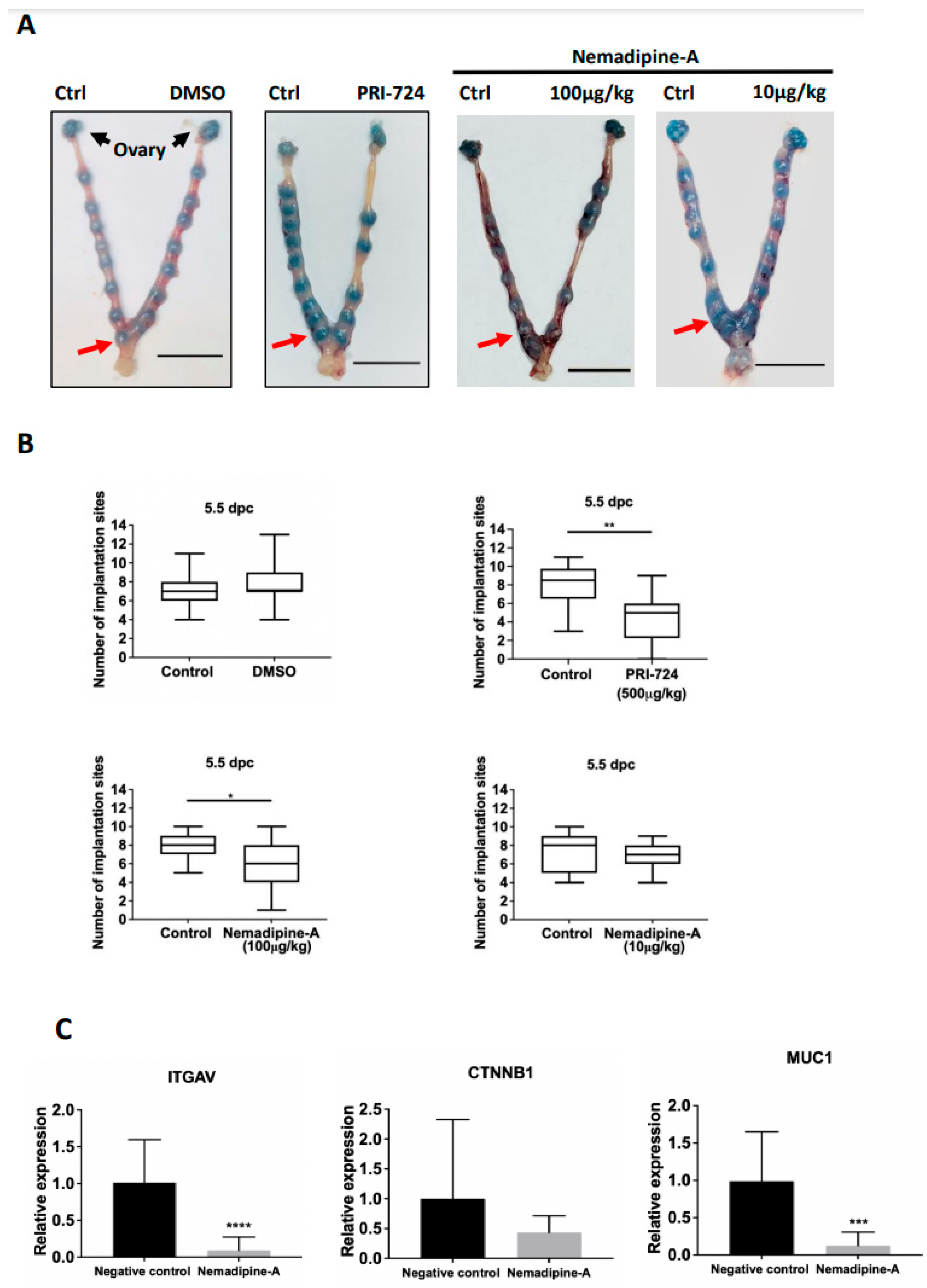
2.5. In Vivo Effects of PBS, DMSO, PRI-724, and Nemadipine-A on Mouse Embryo Implantation
2.6. Nemadipine-A Suppressed Endometrial Receptivity Marker Expression in Mice
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Treatment of Endometrial Epithelial Cells with Steroid Hormones and LOPAC
4.3. Isolation and Culture of Primary hEECs
4.4. Spheroid Attachment Assay
4.5. Cell Viability Assay
4.6. Immunofluorescence Staining
4.7. Murine Model and Transcervical Transfer of LOPAC
4.8. RNA Extraction and Quantitative Reverse Transcription-Polymerase Chain Reaction (qRT—PCR)
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simón, C.; Martín, J.C.; Pellicer, A. Paracrine regulators of implantation. Best Pract. Research. Clin. Obstet. Gynaecol. 2000, 14, 815–826. [Google Scholar] [CrossRef]
- Harper, M.J. The implantation window. Baillieres Clin. Obstet. Gynaecol. 1992, 6, 351–371. [Google Scholar] [CrossRef]
- Meseguer, M.; Pellicer, A.; Simon, C. MUC1 and endometrial receptivity. Mol. Hum. Reprod. 1998, 4, 1089–1098. [Google Scholar] [CrossRef] [Green Version]
- Singh, H.; Aplin, J.D. Adhesion molecules in endometrial epithelium: Tissue integrity and embryo implantation. J. Anat. 2009, 215, 3–13. [Google Scholar] [CrossRef]
- Paule, S.G.; Heng, S.; Samarajeewa, N.; Li, Y.; Mansilla, M.; Webb, A.I.; Nebl, T.; Young, S.L.; Lessey, B.A.; Hull, M.L.; et al. Podocalyxin is a key negative regulator of human endometrial epithelial receptivity for embryo implantation. Hum. Reprod. 2021, 36, 1353–1366. [Google Scholar] [CrossRef]
- Achache, H.; Revel, A. Endometrial receptivity markers, the journey to successful embryo implantation. Hum. Reprod. Update 2006, 12, 731–746. [Google Scholar] [CrossRef] [Green Version]
- Lessey, B.A.; Castelbaum, A.J.; Wolf, L.; Greene, W.; Paulson, M.; Meyer, W.R.; Fritz, M.A. Use of integrins to date the endometrium. Fertil. Steril. 2000, 73, 779–787. [Google Scholar] [CrossRef]
- Tulac, S.; Nayak, N.R.; Kao, L.C.; Van Waes, M.; Huang, J.; Lobo, S.; Germeyer, A.; Lessey, B.A.; Taylor, R.N.; Suchanek, E.; et al. Identification, characterization, and regulation of the canonical Wnt signaling pathway in human endometrium. J. Clin. Endocrinol. Metab. 2003, 88, 3860–3866. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, J.; Ichigo, S.; Hori, M.; Tamaya, T. Alteration of E-cadherin, alpha- and beta-catenin mRNA expression in human uterine endometrium during the menstrual cycle. Gynecol. Endocrinol. 1996, 10, 187–191. [Google Scholar] [CrossRef]
- Valdez-Morales, F.J.; Gamboa-Dominguez, A.; Vital-Reyes, V.S.; Cruz, J.C.; Chimal-Monroy, J.; Franco-Murillo, Y.; Cerbon, M. Changes in receptivity epithelial cell markers of endometrium after ovarian stimulation treatments: Its role during implantation window. Reprod. Health 2015, 12, 45. [Google Scholar] [CrossRef] [Green Version]
- Kiewisz, J.; Wasniewski, T.; Kmiec, Z. Participation of WNT and β-Catenin in Physiological and Pathological Endometrial Changes: Association with Angiogenesis. Biomed. Res. Int. 2015, 2015, 854056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lessey, B.A. Endometrial integrins and the establishment of uterine receptivity. Hum Reprod 1998, 13 (Suppl. S3), 247–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lessey, B.A.; Damjanovich, L.; Coutifaris, C.; Castelbaum, A.; Albelda, S.M.; Buck, C.A. Integrin adhesion molecules in the human endometrium. Correlation with the normal and abnormal menstrual cycle. J. Clin. Investig. 1992, 90, 188–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lessey, B.A.; Castelbaum, A.J. Integrins and implantation in the human. Rev. Endocr. Metab. Disord. 2002, 3, 107–117. [Google Scholar] [CrossRef]
- Elnaggar, A.; Farag, A.H.; Gaber, M.E.; Hafeez, M.A.; Ali, M.S.; Atef, A.M. AlphaVBeta3 Integrin expression within uterine endometrium in unexplained infertility: A prospective cohort study. BMC Womens Health 2017, 17, 90. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.; Hu, L.; Zhang, T.; Wang, M.; Xu, H.; Li, T.C.; Sun, Y.; Wang, C.C. Effects of high progesterone in in-vitro fertilization cycle on DNA methylation and gene expression of adhesion molecules on endometrium during implantation window. J. Assist. Reprod. Genet 2020, 37, 33–43. [Google Scholar] [CrossRef]
- Mardon, H.; Grewal, S.; Mills, K. Experimental models for investigating implantation of the human embryo. Semin. Reprod. Med. 2007, 25, 410–417. [Google Scholar] [CrossRef]
- Weimar, C.H.E.; Post Uiterweer, E.D.; Teklenburg, G.; Heijnen, C.J.; Macklon, N.S. In-vitro model systems for the study of human embryo–endometrium interactions. Reprod. Biomed. Online 2013, 27, 461–476. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Kottawatta, K.S.A.; Kodithuwakku, S.P.; Fernando, T.S.; Lee, Y.L.; Ng, E.H.Y.; Yeung, W.S.B.; Lee, K.F. The fungicide Mancozeb reduces spheroid attachment onto endometrial epithelial cells through downregulation of estrogen receptor β and integrin β3 in Ishikawa cells. Ecotoxicol. Environ. Saf. 2021, 208, 111606. [Google Scholar] [CrossRef]
- Finer, L.B.; Zolna, M.R. Shifts in Intended and Unintended Pregnancies in the United States, 2001–2008. Am. J. Public Health 2014, 104, S43–S48. [Google Scholar] [CrossRef]
- Krisberg, K. Deaths due to unintended pregnancies on the rise.(Global Health Council report). Nation’s Health (1971) 2002, 32, 12. [Google Scholar]
- Black, K.I.; Hussainy, S.Y. Emergency contraception: Oral and intrauterine options. Aust. Fam. Physician 2017, 46, 722–726. [Google Scholar]
- Glasier, A.F.; Cameron, S.T.; Fine, P.M.; Logan, S.J.S.; Casale, W.; Van Horn, J.; Sogor, L.; Blithe, D.L.; Scherrer, B.; Mathe, H.; et al. Ulipristal acetate versus levonorgestrel for emergency contraception: A randomised non-inferiority trial and meta-analysis. Lancet 2010, 375, 555–562. [Google Scholar] [CrossRef]
- Tepekoy, F.; Tepekoy, F.; Akkoyunlu, G.; Akkoyunlu, G.; Demir, R.; Demir, R. The role of Wnt signaling members in the uterus and embryo during pre-implantation and implantation. J. Assist. Reprod. Genet 2015, 32, 337–346. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Zhang, Y.; Lu, J.; Wang, Q.; Wang, S.; Cao, Y.; Wang, H.; Duan, E. Embryo-uterine cross-talk during implantation: The role of Wnt signaling. Mol. Hum. Reprod. 2009, 15, 215–221. [Google Scholar] [CrossRef] [Green Version]
- Tokunaga, Y.; Osawa, Y.; Ohtsuki, T.; Hayashi, Y.; Yamaji, K.; Yamane, D.; Hara, M.; Munekata, K.; Tsukiyama-Kohara, K.; Hishima, T.; et al. Selective inhibitor of Wnt/β-catenin/CBP signaling ameliorates hepatitis C virus-induced liver fibrosis in mouse model. Sci. Rep. 2017, 7, 325. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Yoon, J.E.; Nikapitiya, C.; Kim, T.H.; Uddin, M.B.; Lee, H.C.; Kim, Y.H.; Hwang, J.H.; Chathuranga, K.; Chathuranga, W.A.G.; et al. Small Heterodimer Partner Controls the Virus-Mediated Antiviral Immune Response by Targeting CREB-Binding Protein in the Nucleus. Cell Rep. 2019, 27, 2105–2118.e5. [Google Scholar] [CrossRef] [Green Version]
- Li, H.W.R.; Li, Y.X.; Li, T.T.; Fan, H.; Ng, E.H.; Yeung, W.S.; Ho, P.C.; Lee, K.F. Effect of ulipristal acetate and mifepristone at emergency contraception dose on the embryo-endometrial attachment using an in vitro human trophoblastic spheroid and endometrial cell co-culture model. Hum. Reprod. 2017, 32, 2414–2422. [Google Scholar] [CrossRef]
- Watamoto, T.; Egusa, H.; Sawase, T.; Yatani, H. Screening of Pharmacologically Active Small Molecule Compounds Identifies Antifungal Agents Against Candida Biofilms. Front. Microbiol. 2015, 6, 1453. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Lee, E.M.; Wen, Z.; Cheng, Y.; Huang, W.K.; Qian, X.; Tcw, J.; Kouznetsova, J.; Ogden, S.C.; Hammack, C.; et al. Identification of small-molecule inhibitors of Zika virus infection and induced neural cell death via a drug repurposing screen. Nat. Med. 2016, 22, 1101–1107. [Google Scholar] [CrossRef]
- Sherman, S.P.; Bang, A.G. High-throughput screen for compounds that modulate neurite growth of human induced pluripotent stem cell-derived neurons. Dis Model Mech 2018, 11, dmm031906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chamoun, A.M.; Chockalingam, K.; Bobardt, M.; Simeon, R.; Chang, J.; Gallay, P.; Chen, Z. PD 404,182 is a virocidal small molecule that disrupts hepatitis C virus and human immunodeficiency virus. Antimicrob. Agents Chemother. 2012, 56, 672–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Haj Zen, A.; Nawrot, D.A.; Howarth, A.; Caporali, A.; Ebner, D.; Vernet, A.; Schneider, J.E.; Bhattacharya, S. The Retinoid Agonist Tazarotene Promotes Angiogenesis and Wound Healing. Mol. Ther. 2016, 24, 1745–1759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vliet, S.M.; Ho, T.C.; Volz, D.C. Behavioral screening of the LOPAC(1280) library in zebrafish embryos. Toxicol. Appl. Pharmacol. 2017, 329, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T.; Satow, R.; Kato, A.; Tamura, M.; Murayama, Y.; Saya, H.; Kojima, H.; Nagano, T.; Okabe, T.; Fukami, K. Identification of novel small compounds that restore E-cadherin expression and inhibit tumor cell motility and invasiveness. Biochem. Pharmacol. 2013, 86, 1419–1429. [Google Scholar] [CrossRef] [PubMed]
- Hannan, N.J.; Paiva, P.; Dimitriadis, E.; Salamonsen, L.A. Models for study of human embryo implantation: Choice of cell lines? Biol. Reprod. 2010, 82, 235–245. [Google Scholar] [CrossRef] [Green Version]
- Matthews, C.J.; Redfern, C.P.; Hirst, B.H.; Thomas, E.J. Characterization of human purified epithelial and stromal cells from endometrium and endometriosis in tissue culture. Fertil. Steril. 1992, 57, 990–997. [Google Scholar] [CrossRef]
- Kobayashi, R.; Terakawa, J.; Omatsu, T.; Hengjan, Y.; Mizutani, T.; Ohmori, Y.; Hondo, E. The Window of Implantation Is Closed by Estrogen via Insulin-Like Growth Factor 1 Pathway. J. Reprod. Infertil. 2017, 18, 231–241. [Google Scholar]
- Xu, L.; Sun, L.; Xie, L.; Mou, S.; Zhang, D.; Zhu, J.; Xu, P. Advance in L-Type Calcium Channel Structures, Functions and Molecular Modeling. Curr. Med. Chem. 2020, 27, 514–524. [Google Scholar] [CrossRef]
- Morales, D.; Hermosilla, T.; Varela, D. Calcium-dependent inactivation controls cardiac L-type Ca2+ currents under β-adrenergic stimulation. J. Gen. Physiol. 2019, 151, 786–797. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.; Jang, H.; Seo, H.; Yoo, I.; Han, J.; Kim, M.; Lee, S.; Ka, H. Changes in calcium levels in the endometrium throughout pregnancy and the role of calcium on endometrial gene expression at the time of conceptus implantation in pigs. Mol. Reprod. Dev. 2019, 86, 883–895. [Google Scholar] [CrossRef] [PubMed]
- Ebner, T.; Oppelt, P.; Wober, M.; Staples, P.; Mayer, R.B.; Sonnleitner, U.; Bulfon-Vogl, S.; Gruber, I.; Haid, A.E.; Shebl, O. Treatment with Ca2+ ionophore improves embryo development and outcome in cases with previous developmental problems: A prospective multicenter study. Hum. Reprod. 2014, 30, 97–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, Y.C.; Guo, J.H.; Liu, X.; Zhang, R.; Tsang, L.L.; Dong, J.D.; Chen, H.; Yu, M.K.; Jiang, X.; Zhang, X.H.; et al. Activation of the epithelial Na+ channel triggers prostaglandin E(2) release and production required for embryo implantation. Nat. Med. 2012, 18, 1112–1117. [Google Scholar] [CrossRef]
- Ruan, Y.C.; Chen, H.; Chan, H.C. Ion channels in the endometrium: Regulation of endometrial receptivity and embryo implantation. Hum. Reprod. Update 2014, 20, 517–529. [Google Scholar] [CrossRef] [Green Version]
- He, J.-P.; Zhao, M.; Zhang, W.-Q.; Huang, M.-Y.; Zhu, C.; Cheng, H.-Z.; Liu, J.-L. Identification of Gene Expression Changes Associated With Uterine Receptivity in Mice. Front. Physiol. 2019, 10, 125. [Google Scholar] [CrossRef]
- Illera, M.J.; Cullinan, E.; Gui, Y.; Yuan, L.; Beyler, S.A.; Lessey, B.A. Blockade of the alpha(v)beta(3) integrin adversely affects implantation in the mouse. Biol. Reprod. 2000, 62, 1285–1290. [Google Scholar] [CrossRef] [Green Version]
- Jha, R.K.; Titus, S.; Saxena, D.; Kumar, P.G.; Laloraya, M. Profiling of E-cadherin, β-catenin and Ca2+ in embryo–uterine interactions at implantation. FEBS Lett. 2006, 580, 5653–5660. [Google Scholar] [CrossRef] [Green Version]
- Braga, V.M.; Gendler, S.J. Modulation of Muc-1 mucin expression in the mouse uterus during the estrus cycle, early pregnancy and placentation. J. Cell Sci. 1993, 105 Pt 2, 397–405. [Google Scholar] [CrossRef]
- Gendler, S.J. MUC1, the renaissance molecule. J. Mammary Gland Biol. Neoplasia 2001, 6, 339–353. [Google Scholar] [CrossRef]
- Brayman, M.; Thathiah, A.; Carson, D.D. MUC1: A multifunctional cell surface component of reproductive tissue epithelia. Reprod. Biol. Endocrinol. 2004, 2, 4. [Google Scholar] [CrossRef] [Green Version]
- Hey, N.A.; Li, T.C.; Devine, P.L.; Graham, R.A.; Saravelos, H.; Aplin, J.D. MUC1 in secretory phase endometrium: Expression in precisely dated biopsies and flushings from normal and recurrent miscarriage patients. Hum. Reprod. 1995, 10, 2655–2662. [Google Scholar] [CrossRef]
- Lee, Y.L.; Fong, S.W.; Chen, A.C.; Li, T.; Yue, C.; Lee, C.L.; Ng, E.H.; Yeung, W.S.; Lee, K.F. Establishment of a novel human embryonic stem cell-derived trophoblastic spheroid implantation model. Hum. Reprod. 2015, 30, 2614–2626. [Google Scholar] [CrossRef] [Green Version]
- Yue, C.; Chen, A.C.H.; Tian, S.; Fong, S.W.; Lee, K.C.; Zhang, J.; Ng, E.H.Y.; Lee, K.F.; Yeung, W.S.B.; Lee, Y.L. Human embryonic stem cell-derived blastocyst-like spheroids resemble human trophectoderm during early implantation process. Fertil. Steril. 2020, 114, 653–664.e6. [Google Scholar] [CrossRef]
- Ho, H.; Singh, H.; Aljofan, M.; Nie, G. A high-throughput in vitro model of human embryo attachment. Fertil. Steril. 2012, 97, 974–978. [Google Scholar] [CrossRef]
- Cui, L.; Zhang, Z.; Sun, F.; Duan, X.; Wang, M.; Di, K.; Li, X. Transcervical embryo transfer in mice. J. Am. Assoc. Lab. Anim. Sci. 2014, 53, 228–231. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecule ID | Change (%) in | p-Value | LC50 (µM) |
|---|---|---|---|
| Attachment Rate (at 10 μM) | |||
| P14A7 | −88.3 | 0.0142 | 3.58 |
| P3E6 | −87.5 | 0.0286 | 11.85 |
| P14F4 | −84.8 | 0.004 | 0.07 |
| P11A8 | −84.7 | 0.004 | 2.01 |
| P15A9 | −81.3 | 0.0095 | 4.42 |
| P13A11 | −80.3 | 0.004 | 0.35 |
| P1C7 | −79.8 | 0.0286 | 3.49 |
| P14G9 | −78.1 | 0.0402 | 0.51 |
| P13G8 | −77.6 | 0.004 | 1.93 |
| P15E10 | −77.3 | 0.004 | 0.11 |
| P11D5 | −77.1 | 0.004 | 3.23 |
| P3A5 | −76.5 | 0.0286 | 0.09 |
| P8F6 | −74.4 | 0.0286 | 10.01 |
| P11B5 | −74.0 | 0.004 | 17.37 |
| P10B4 | −72.1 | 0.0286 | 0.02 |
| P13E7 | −72.1 | 0.004 | 0.73 |
| P16E7 | −71.9 | 0.004 | 3.05 |
| P16E5 | −71.3 | 0.004 | 7.03 |
| P14E11 | −70.5 | 0.004 | 0.33 |
| P5D6 | −55.2 | 0.0286 | 6.91 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Fernando, S.R.; Lee, Y.-L.; Yeung, W.S.-B.; Ng, E.H.-Y.; Li, R.H.-W.; Lee, K.-F. High-Throughput In Vitro Screening Identified Nemadipine as a Novel Suppressor of Embryo Implantation. Int. J. Mol. Sci. 2022, 23, 5073. https://doi.org/10.3390/ijms23095073
Chen X, Fernando SR, Lee Y-L, Yeung WS-B, Ng EH-Y, Li RH-W, Lee K-F. High-Throughput In Vitro Screening Identified Nemadipine as a Novel Suppressor of Embryo Implantation. International Journal of Molecular Sciences. 2022; 23(9):5073. https://doi.org/10.3390/ijms23095073
Chicago/Turabian StyleChen, Xian, Sudini Ranshaya Fernando, Yin-Lau Lee, William Shu-Biu Yeung, Ernest Hung-Yu Ng, Raymond Hang-Wun Li, and Kai-Fai Lee. 2022. "High-Throughput In Vitro Screening Identified Nemadipine as a Novel Suppressor of Embryo Implantation" International Journal of Molecular Sciences 23, no. 9: 5073. https://doi.org/10.3390/ijms23095073
APA StyleChen, X., Fernando, S. R., Lee, Y.-L., Yeung, W. S.-B., Ng, E. H.-Y., Li, R. H.-W., & Lee, K.-F. (2022). High-Throughput In Vitro Screening Identified Nemadipine as a Novel Suppressor of Embryo Implantation. International Journal of Molecular Sciences, 23(9), 5073. https://doi.org/10.3390/ijms23095073